Survey
* Your assessment is very important for improving the work of artificial intelligence, which forms the content of this project
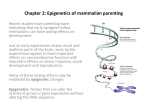
Epigenetics Immunology of Histocompatibility May 2nd 2006 Jonathan Duke-Cohan (DFCI/HMS) This presentation draws heavily upon some superb images prepared by others and freely available on the Web. In every instance, the copyright of the original artists is acknowledged, and the artwork is being used here solely for teaching purposes. To all the anonymous authors of these graphics, thank you! Epigenetics Epigenesis: • • the unfolding development of an organism, and in particular the development of a plant or animal from an egg or spore through a sequence of steps in which cells differentiate and organs form; in contrast to “preformation” or the “homunculus” theory Epigenetics: • C.H. Waddington in 1942 defined it as “the branch of biology which studies the causal interactions between genes and their products which bring the phenotype into being”. Epigenetic inheritance: • Epigenetic inheritance is the transmission of information from a cell or multicellular organism to its descendants without that information being encoded in the nucleotide sequence of the gene. Epigenetic phenomena Example - Cellular proliferation: • The progeny of a fibroblast cell division are fibroblasts • The progeny of a stem cell may or may not be a stem cell How? • Transcriptional factor presence determined by parent cell? • Mechanisms of maintaining gene repression/activation through generations? • Environmental cues? (e.g. hormones, cytokines, pO2, osmolarity) Phenotype or “trait” is the end result, but we’re really talking about inherited control of gene expression Epigenetic phenomena Example 2 – Chromosomal dosage and compensation • • Women are XX, men are XY How are levels of all essential X-encoded gene products similar between men and women if women have twice the number of alleles? Mary Lyon – 1961 • • • • in cells with multiple X chromosomes, all but one is inactivated during mammalian embryogenesis – the “Lyon effect” X-inactivation; which X? Usually random … but always paternal in marsupials and variable in calico cats representing regional expression of differing pigmentation genes on alternate X chromosomes Epigenetic phenomena X-inactivation • The repressed X-chromosome condenses to form a Barr body • In fact, 10-15% of repressed alleles are active – to compensate for recessive alleles of sex-linked disorders including haemophilia and colour blindness • How does the cell “count” X chromosomes? Epigenetics and Disease Prader-Willi Syndrome Angelman Syndrome Beckwith-Wiedemann Syndrome del-15q11.2-13 del-15q11.2-13 del-11p15.5 • Prader-Willi: obesity, muscular hypotonia, mental retardation, short stature, hypogonadism, small limbs • Up to 4Mb deleted, primary genes affected are: • SNRPN (small nuclear ribonucleoprotein polypeptide N) • NDN • MKRN3 • IPW (necdin) (makorin) (imprinted in Prader-Willi syndrome) (!!!) but • Deletions account for ~70%, and always of the paternal chromosome • 28% are uniparental disomy*, always maternal, with NO genomic deletion • <2% are small mutations on the paternal side, affecting the whole region *usually follows trisomic rescue – 47 chromosomes in fertilised ovum, one lost on cell division. (correction by two mistakes) Epigenetics and Disease Prader-Willi: Independent of primary gene sequence, there are genomic modifications that can be passed on from the parental environment – “Epigenetic marks” Epigenetic marks Modification of primary sequence • CpG methylation Modification of amino terminal of histones: • Methylation of lysines/arginines • Acetylation/ubiquitination of lysines • Serine phosphorylation DNA methylation CpG – Cytosine phosphate Guanine • • • • Strongly represented in repetitive sequence associated with retroviralderived sequence Can be methylated to generate 5-methylcytosine Spontaneously deaminates to form thymine Poorly recognised by DNA repair systems thus: • CG→TG mutation is propagated • CpG levels are less frequent than predicted 1/16 • May contribute to relative inactivity of retroelements DNA methylation CpG–islands CpG frequency CpG islands in Rb gene (180kb) • • low frequency 70-80% methylated promoter-related CpG are usually unmethylated methylation-free state is essential for transcription of the associated gene DNA methylation 3 human DNA methyltransferases • • • DNMT1 DNMT3A DNMT3B maintenance methyltransferases de novo methyltransferases – highly expressed at embryo implantation when waves of de novo methylation are occurring in the genome ◄daughter strand ◄daughter strand DNA methylation CpG–island methylation – how does it affect transcription? • • • • methylated-DNA binding proteins (MECP2, methyl CpG binding protein 2 ) bind to DNA this recruits a complex of histone deacetylases and SIN3A induces a closed chromatin structure → gene silencing in contrast to usual deacetylation-related silencing, when methylation is involved, it’s (almost) irreversible gene Histone modification ▲ ▲ active silenced Histone modification Mechanism: • • • Acetylation of H3 or H4 leads to unfolding and accessibility of chromatin (histone acetyltransferases) Methylation of K4 of H3 = active gene expression Methylation of K9 of H3 = gene silencing Prader-Willi revisited…. • Presentation with no genomic deletion • Uniparental maternal disomy ◄ paternal ◄ maternal Imprinting centre ◄ maternal ◄ maternal ▼ ◄ paternal ◄ maternal and Angelman syndrome • Presentation with no genomic deletion • Uniparental paternal disomy • Imprinting center defect • leading to loss of maternal UBE3A function • UBE3A paternal allele silenced in specific brain regions, maternal allele active almost exclusively in hippocampus and cerebellum ▲ Epigenetics in embryonic development • In each generation, the imprint inherited from the parent of the opposite sex must be erased and re-established in developing germ cells so that maternal or paternal imprint is appropriate for the sex of the individual • During early embryogenesis, a wave of demethylation followed by a wave of sex-appropriate remethylation (“resetting”) • Is “resetting” universal? And when does it occur? • In spermatazoa, all potential methylation sites for SNRPN and NDN are unmethylated • In oocytes, near complete methylation of these genes at the germinal stage and metaphases I and II • ~50% methylation in the pre-implantation embryo Epigenetics in embryonic development Implications for assisted reproductive technology (ART)? • in vitro fertilisation by intracytoplasmic sperm injection: girls maintained paternal imprint New England Journal of Medicine 346: 725-730 (2002) Hansen M, Kurinczuk JJ, Bower C, Webb S The risk of major birth defects after intracytoplasmic sperm injection and in vitro fertilization Following and ICS, 26/301 (8.6%) IVF, 75/837 (9.0%) have major birth defects compared with 168/4000 naturally-conceived infants (4.2%) ART often required for sperm malfunction, but Angelman (for example) which has increased incidence following IVF/ICS is a result of loss of maternal methylation? Increased risk more a reflection of in vitro culture effects? X-inactivation revisited • How does the cell “know” it has more than one X-chromosome, and needs to inactivate one? • At the four cell stage, the paternally-derived X chromosome is inactivated. The extra-embryonic tissue ( →placenta etc.) retain this imprinting i.e. only the maternal X is active • In the early blastocyst, this initial imprinted paternal X-inactivation is reversed (a wave of demethylation) and both X are active • Random inactivation, maintained in all descendants of that cell. Thus mosaicism in calico cats, reflecting X-linked heterozygosity • In the female germline, all X-inactivation is reversed X-inactivation revisited (continued) • Initially hypothesised that there were limiting levels of a blocker – enough to block one X but not enough to block more than this. • But this cannot be so since in cells with more than two X chromosomes, all except one were blocked • XIC – the X inactivation centre • required for X-inactivation • introduction of this region to ANY chromosome leads to silencing • encodes two genes: XIST, and TSIX • XIST: X (inactive)-specific transcript • TSIX: X (inactive)-specific transcript-antisense • No XIST, no inactivation – encodes a large RNA, not protein X-inactivation revisited (continued) • XIST is the only gene expressed from the inactive X (Xi) and not expressed from the active X (Xa) • Initially, both X weakly express XIST, then on the future Xi, dramatic upregulation, and on future Xa, no expression • XIST coats the Xi chromosome, moving out from the XIC • The activity of TSIX is reciprocal to that of XIST, expressed by Xa it suppresses XIST – alleles bearing a deletion of TSIX are much more likely to be inactivated XIST: X (inactive)-specific transcript ► ◄TSIX: (inactive)-specific transcript-antisense