Survey
* Your assessment is very important for improving the work of artificial intelligence, which forms the content of this project
* Your assessment is very important for improving the work of artificial intelligence, which forms the content of this project
Nitrilases
Self-terminating, homo-oliogomeric
spirals with industrial applications
Trevor Sewell
University of Cape Town
with lots of help from:
Mark Berman (Cape Town)
Paul Chang (Cape Town)
Dakshina M. Jandhyala and
Michael Benedik (Houston)
Paul Meyers (Cape Town)
Ed Egelman (Virginia)
Dennis Burford (Cape Town)
Helen Saibil (London)
and the EMU at UCT:
Mohamed Jaffer
Brendon Price
Miranda Waldron
James Duncan
William Williams
The Wellcome Trust
Establishing the principles
underlying the oligomeric
structure of the nitrilases.
Insights into the structures of
nitrilases and GroEL from 3D
electron microscopy
Trevor Sewell
with lots of help from:
Mark Berman (Cape Town)
Dakshina M. Jandhyala and
Michael Benedik (Houston)
Paul Meyers (Cape Town)
Ed Egelman (Virginia)
Dennis Burford (Cape Town)
Helen Saibil (London)
The Wellcome Trust
and the EMU at UCT:
Mohamed Jaffer
Brendon Price
Miranda Waldron
James Duncan
William Williams
Why nitrilases are interesting:
Cleave non-peptide C-N bonds
Used in industrial processes e.g. manufacture of
acrylic acid - efficient and environmentally friendly
Detoxification of cyanide waste - bioremediation
Role in plants - in synthesis of auxin - is one of
few biological roles properly documented
Variety of different reported sizes of apparently
homogeneous material
Apparent link between quaternary structure and
activity in some enzymes
What we know:
Cysteine, lysine and glutamic acid at active site
pH optimum 7.6 - 8.0
Molecular weight of subunit = 37kD
Close relatives all have large molecular weights reported number of subnits varies in different species
from monomers and dimers, to tens and occasionally
hundreds.
Sequences of over 400 members of the nitrilase
superfamily
Atomic structure of two (now four) distant members of
the superfamily.
The B. pumilus enzyme complex measures 10nm x
10nm x 20nm
The Structure of Nitrilases
Self-terminating, homo-oliogomeric
spirals with industrial applications
Trevor Sewell,
Biotechnology Department UWC and
EMU, University of Cape Town
Ndoriah Thuku (UWC)
Margot Scheffer(UCT)
Mark Berman (UCT)
Paul Chang (UCT)
Dakshina M. Jandhyala(Houston)
Xing Zhang (Houston)
Michael Benedik (Tamu)
Paul Meyers (Cape Town)
Ed Egelman (Virginia)
Arvind Varsani(Cape Town)
Helen Saibil (London)
and the EMU at UCT:
Mohamed Jaffer
Brandon Weber
Brendon Price
Miranda Waldron
James Duncan
Sean Karriem
The Wellcome Trust
Carnegie Corporation
Useful Industrial
Enzymes
Nicotinic Acid
Mandelic Acid
Ibuprophen
Detoxification of
cyanide
Reactions catalysed
Nitrilase - cyanide dihydratase - B. pumilus, P.stutzeri
Cyanide hydratase - G. sorghi
Nit active site
Putative catalytic mechanism
Topology diagram of
the a-b-b-a-a-b-b-a To Fhit
domain
dimer structure
found in both DCase
and Nit.
Nit labelling.
Pace et al (2000)
To Fhit
domain
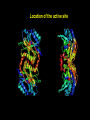
Location of the active site
Two questions concerning the
quaternary structure :
Homologous nitrilases have subunit molecular
weights around 40 kDa but are generally reported
to occur in complexes with 2 - 18 subunits. Why is
this?
Nitrilases from several Rhodococcus species are
inactive as dimers but form active decamers or
dodecamers on incubation with substrate. Why is
this?
What we did:
Reconstructed a 3D map from negatively stained
images to a resolution of 2.5nm using SPIDER
Located homologues in the PDB and aligned
them to our sequences with GENthreader.
Developed a dimer model for our enzymes
based on the non-spiral forming homologues.
Located the dimer model within the density with
CoLoRes in SITUS and O.
The Process
Negative stain (uranyl acetate on carbon film)
Image using low dose
Digitize film
Select images
Classify images
Starting model using a common-lines based method
Match images to projections of model
iterat
Reconstruct » new model
e
Check resolution of structure
Negatively stained native B. pumilus nitrilase, pH8
Multi-reference alignment
Iterative 3D reconstruction
Averages of the 84 image sets used in the reconstriction
of the cyanide dihydratase from P. stutzeri AK61
The refinement of the structure of the
nitrilase from Pseudomonas stutzeri
(7008 images)
video made by Paul
Chang
The refinement of the structure of the
nitrilase from Bacillus pumilus
(11661 images)
video made by Paul
Chang
B. pumilus nitrilase (pH 6)
bulge
ridge
P. stutzeri nitrilase (pH 8)
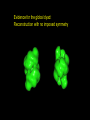
Evidence for the global dyad:
Reconstruction with no imposed symmetry
Cylindrical projection of P. stutzeri nitrilase
32
1.6 nm vertical
displacement
between local
two fold axes
z (nm)
0
-180
96.5
0
f (°)
76.5 70.5 70.5 76.5
180
96.5
Angular offset between local two-fold axes (°)
The cylindrical projection shows that successive
local two fold axes are separated by increasing
angular rotations but a constant shift along the
helix axis. The projections of the subunits also
appear increasingly elongated along v, because
they are closer to the helix axis.
We know the sequences of the B. pumilus
enzyme, thanks to Michael Benedik and Dakshina
Jandhyala at the University of Houston, and the P.
stutzeri enzyme due to Atsushi Watanabe et al,
(1998) BBA, 1382, 1-4.
They have 70% sequence homology.
A search for structurally homologous enzymes
in the Protein Data Bank using GenTHREADER
produced two enzymes: Nit and DCase.
These have less than 20% sequence homology
to our enzymes.
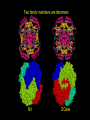
Two family members are tetramers
Nit
DCase
In the tetramer there are two interacting surfaces
almost at right angles to one another
Surface A
alpha helix
Nit
Surface B
beta sheet
DCase
Topology diagram of
the a-b-b-a-a-b-b-a To Fhit domain
dimer structure found
in both DCase and
Nit.
Nit labelling.
Pace et al (2000)
To Fhit domain
Superposition of the alpha carbons of DCase and Nit
DCase
Nit
cys 169, lys 127, glu 54 catalytic triad
An alignment of the nitrilase sequences
with Nit and DCase by GenTHREADER
From the sequence comparisons we
conclude that:
The insertions and deletions in our enzymes
relative to NIT and DCase are in outer loops and
will not impinge on the tertiary structure that is
crucial to the fold.
A major difference between our enzymes and the
tetramers is the existence two significant
insertions and the C-terminal extension.
Need to fit model into density
The two fold axes must coincide
Dimer with A surface associating
modeled on residues 10-291 of Nit
Surface A
Cterminal
Surface B
Surface B
Cterminal
Dimer with B surface associating
modeled on residues 10-280 of Nit
Surface A
Cterminal
Cterminal
Surface A
Surface B
4 ways to align global dyad to dimer axis
A
surface
mating
B
surface
mating
This was repeated for the other handedness
What is wrong with the B surface models?
Steric clash between NH5 and
NS13 and NH3 in the neighbouring
dimer
Poor
fits
Unexplainable gaps in density
The final, left-handed, 14-subunit model
Termination of the helix
The C surface is flexible and operates as a hinge
between the subunits.
As subunits are added at terminus of the spiral
new opportunities arise for interactions across the
groove.
The addition of a further subunit will occur if the
energetic considerations favour this in preference
to interactions across the groove which result in
steric hindrance which would prevent the addition
of a further subunit.
Contacts a and b result in the terminal dimer having an inwards tilt of 12
degrees thus preventing the addition of a further dimer. .
I
B
a
Contacts c and d are between helices NH2. The
contact area has a local pseudo-dyad axis.
M
d
d
D
K
glu 82
c
B
c
lys 86
N
b
L
M
d
J
K
c
b d
H
I
a
F
G
c
D
E
a
B
C
A
(a)
Cylindrical projection
32
z (nm)
0
-320
(b)
-244
-147
-71
0
f
(°)
71
147
244
320
Superposition of the P. stutzeri nitrilase dimer model
onto the A surface Nit dimer
Insertions
thought to be
responsible
for the C
surface
interactions
Deletion: causes steric
hindrance and would
prevent C surface
interactions
A prominent ridge on the outer surface was not filled by the
initial model. A four stranded segment of sheet from bovine
superoxide dismutase fills the density has the correct number of
residues and mates with the ends in left handed models only.
Crosslinking with
glutaraldehyde:
the protein from the
column was diluted
32 fold and
crosslinked
with the
glutaraldehyde
concentration
indicated for 1.25 hrs.
Incompletely unfolded
conformational isomers?
10x(?)
8x(?)
6x(?)
4x
3x
2x
{
nitrilase
monomer
0 .002.005 .01 .02 .05 .1 .2%
The flexible C surface
The location of the active site and B surface
Does the quaternary structure have functional
significance?
Nagasawa et al (2000) have found that isolated
dimers of the related nitrilase from Rhodococcus
rhodochrous J1 are inactive. However in the
presence of certain substrates they assemble to form
an active decamer. ( A decamer is required to
produce one turn of the spiral.) We do not yet know
whether this occurs in our case as we don't yet know
how not to produce the spiral in our enzymes.
The enzyme from B. pumilus forms long fibres at
pH 5.4
Unidirectional shadowing shows that the long helices
are left handed.
The handedness of the spiral
Defined length oligomers from B. pumilus form long
helices at pH 5.4. These are shown by shadowing to be
left handed.
Our dimer model fits better into left handed spirals than
right handed spirals as shown by SITUS correlation coefficients.
Only in left handed spirals is there empty space in the
map to accommodate the insertions relative to non
spiral-forming homologues.
What came out of the study?
A new, defined size, short, spiral, homo-oligomeric
quaternary structure
The handedness of the spiral
The conserved interface (A surface)
The residues involved in a previously undiscovered
interface (C surface)
A model of this interface which would explain its
flexibility
A reason for the termination of the spiral
A reason for the variety of subunit sizes in the
enzymes
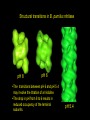
Structural transitions in B. pumilus nitrilase
pH 8
pH 6
The transitions between pH 6 and pH 5.4
may involve the titration of a histidine.
The drop in pH from 8 to 6 results in
reduced occupancy of the terminal
subunits.
pH 5.4
Regular helix having 9.4 residues per
turn ( for dimer model: Dv=76.7 ,
Dz=1.58 nm )
B. pumilus
P. stutzeri
G. sorghi
Potential for two salt bridges in pumilus
Repulsion in stutzeri - no long fibres
One salt bridge in sorghi - always fibres
Activity increases when structural transition occurs.
Could this mean that 2 extra sites per 18mer become active?
The Effect of Surface Mutations on Activity
Mutant
Surface
Change and location
Activity
B pumilus
1. Delta 303
A
Vgtg->stop
2. Delta 293
A
Matg->stop
Full
activity
Partial
activity
Inactive
Inactive
3. Delta 279
A
4.
A
Y201D/A204D
Ytat->stop
Ytat->Dgac, Agcg->Dgac
5. Delta 219233
6. 90
Inactive
D
MKEMICLTQEQRDYF was deleted.
235 Egaa->Naac
EAAKRNE->AAARKNK
A
A
A
A
A
A
Sagt->stop
Vgtg->stop
Qcag->stop
Ytat->stop
Kaaa->stop
Ytac->Dgac, Ctgc->Dgac
Inactive
Inactive
Inactive
Inactive
Inactive
Inactive
MKDMLCETQEERDYF deleted.
Inactive
P stutzeri
7. Delta 310
8. Delta 302
9. Delta 296
10. Delta 285
11. Delta 276
12.
Y200D/C203D
C
13. Delta 220- C
234
Hybrids
14. Pum – Stu A
15. Stu – Pum A
Full
activity
Residues 1-286 from B. pumilus, 287- Full
end from P. stutzeri
Activity
Residues 1-286 from P. stutzeri, 287Inactive
end from B. pumilus
The only histidines in pumilus
that are not in stutzeri.
The ATCC pumilus has no histidines in the tail - its properties are being
studied
Rhodococcus rhodochrous J1
20nm
Negatively stained fibres of J1 nitrilase (0.45mg/ml) buffered in 20mM KH2PO4, 50mM NaCl at pH 7.8.
Magnification 50000x
G. sorghi CHT reconstructions
WT1 (film)
WT2 (CCD)
Mutant R87Q(CCD)
Gloeocercospora sorghi cyanide hydratase
Surprise! Quaternary helix is right handed
What's empty?
C terminal extension
C surface linker as before
What interactions stabilize the spiral?
B. pumilus
P. stutzeri
G. sorghi
R91Q
E82V
charge
mutant
active
-
E82V
no
+
R91Q
no
-
D92Q
yes
a-surface
Y217D
no
a-surface
Y217E
no
We think we know where all the bits of the
molecule are at coarse resolution.
We think we know what stabilizes the spiral
and causes its termination.
We think that the spiral is essential for activity.
Biotechnological uses?
Can the knowlege we have gained be used to
enhance:
Stability
Activity
Ease of Purification
Ease of Immobilization
????
B. pumilus has a complex internal structure which changes during its
life cycle. It is therefore relevant to ask where the nitrilase is located in
the hope that it may give a clue to its function.