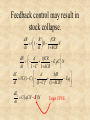Survey
* Your assessment is very important for improving the work of artificial intelligence, which forms the content of this project
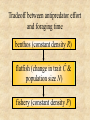
Self-extinction due to adaptive change in foraging and anti-predator effort • Matsuda H, Abrams PA (1994a) Runaway evolution to self-extinction under asymmetric competition. Evolution 48:1764-1772. • Matsuda H, Abrams PA (1994b) Timid consumers: self-extinction due to adaptive change in foraging and anti-predator effort. Theor Pop Biol 45:76-91. • Matsuda H, Abrams PA (2004) Effects of predator-prey interactions and adaptive change on sustainable yield. Can J Fish Aq Sci in press Matsuda & Abrams (1994a, b) • Frequency dependent selection may decrease the population size and the population growth rate. • Therefore, self-extinction due to frequencydependent selection is possible. • e.g., Timid herbivores (Matsuda & Abrams 1994, Theor. Pop. Biol. ) Tradeoff between antipredator effort and foraging time benthos (constantdensity densityR)R) plant (constant flatfish (change herbivore (changeinintrait traitĈĈ&& population size N) fishery (constant carnivore (constantdensity densityP)P) Model I: Harbivore’s fitness W • W(C) = B(CR) - M(C,P) - d W (C ) 1 bCR eCP d • Optimal foraging time W bR 0 C 2 2 C 4e P –always decreases as predator increases; Optimal foraging time dN CR CP ( D N ) N dt 1 bCR 1 hCN • I = ĈR: Foraging intake rate I = (individual’s foraging time)(plant density), • B = ĈR/(1 + bĈR) Benefit B from intake saturates with intake, • Risk M of predation (type II functional response) M = ĈP / (1 + hCN) • C is population mean trait value h: handling time Population & evolutionary dynamics dN CP CR (W * 1) N (D N ) N dt 1 bCR 1 hCN dC dW g (C ) dt dCˆ Cˆ C • Equilibrium population – N* = Ns (stable level) – N* = Nu (unstable critical level) – N* = 0 (extinct) figures ESS ESS ESS ESS A model for exploitation of predator dR R fCR r 1 R N dt 1 hCR K dN d bfCR e1qC N dt 1 C 1 hCR dC d bfR VC (1 C ) e1q 2 2 dt (1 C ) (1 hCR) R: prey density; N: consumer density; e1: fishing effort; C: foraging time; q: catchability; V; evolution velocity; Stock & yield Non-standard fisheriesstock/yield relationship fishing effort may increase stock. P Y Fishing effort Stock△ and yield○ are maximized just before stock collapse. Feedback control may result in stock collapse. dR R fCR r 1 R N dt 1 hCR K dN d bfCR E1qC N dt 1 C 1 hCR dC d bfR VC (1 C ) E1q 2 2 dt (1 C ) (1 hCR) dE1 U qCN S N dt Target CPUE Feedback control may result in stock collapse. We should take account of adaptation in managing endangered species. • Does evolutionary response of species always increase its population size? – No • “the fish may become poorer foragers as the result of fishing, and that this may result in extinction, or at least contribute to reducing their population size.” (Matsuda & Abrams 1994b)