Survey
* Your assessment is very important for improving the workof artificial intelligence, which forms the content of this project
From www.bloodjournal.org by guest on December 22, 2014. For personal use only.
Molecular Analysis of Polish Patients With Factor VI1 Deficiency
By Arnaldo A. Arbini, Dinah Bodkin, Stanislaw Lopaciuk, and Kenneth A. Bauer
We analyzed the mutations in patients from 10 Polish kindreds with a bleeding diathesis due
to factor VI1 deficiency.
Patients from eight families had plasma levels of factorVI1
coagulant activity (VII:C) and factorVI1 antigen (VIkAg) that
were less than 4% of normal. The coding sequence of the
factor VI1 gene was amplified from genomic DNA by polymerase chain reaction (PCR). Sequencing demonstrated a C
to T transition at position 10798 resultingin Ala294Va1, a G
to A transition at 10976 resultingin Arg353Gln. and a single
bp deletionat 11125 to 11 128 causing a frameshift
mutation
in the triplet encoding amino acid 404. Homozygosity for
the three sequence alterations was confirmed with the restriction enzymes Awall and MSpr and allele specificPCR,
respectively. A homozygous
patient from a ninth family with
levels of VkC and VII:Ag of 4% and 17%. respectively, had
Ala294Val andthe frameshift mutation, but not Arg353Gln.
Investigation of a homozygouspatient from a tenth kindred
with VkC and VII:Ag of 11% and 47%. respectively, demonstrated Ala294Val and Arg353Gln.
but not the frameshift mutation. Based on the abovedata, we conclude that the
frameshift mutation in the codon for amino acid 404 is associated with marked reductions in VkC, Arg353Gln can decrease plasma levels of factor VI1 in the presence of other
mutations in the factor VI1 gene, and Ala294Val resultsin a
dysfunctional factor VI1 molecule.
0 1994 by The American Societyof Hematology.
H
structurally abnormal factor VI1 molecules, and CRMR defects may result from a combination of these mechanisms.
Some CRMR or CRM+ factor VI1 molecules give variable
VI1:C results when assayed with tissue factor from different
species (ie, rabbit, ox, simian, and human
Based
on clinical studies using these test procedures, it seems certain that the molecular basis of factor VI1 deficiency will
turn outto be very heterogeneous. The autosomal inheritance
pattern of the disorder implies that severely affected individuals can be homozygous or doubly heterozygous.
The molecular basis of factor VI1 deficiency, in contrast
to the relatively common sex-linked hemophilias, has only
recently begun to be investigated, and most of the reported
mutations have been in CRM’ patients with either no bleeding history ora mild hemorrhagic diathe~is.”~’~
In this report,
we describe the molecular defects in ten Polish patients with
homozygous factor VI1 deficiency who were not known to
be related, and had moderate or severe bleeding manifestations. Our results show that a common polymorphism in the
factor VI1 gene, Arg353Gln, can result in a further reduction
in VI1:C and VII:Ag levels in patients with other mutations
in the factor V11 gene.
UMAN FACTOR VI1 is a vitamin K-dependent glycoprotein with a molecular weight of 50 k D . It is composed of 406 amino acids,’ and is present in normal plasma
at a concentration of 0.5 pg/mL.’ In association with tissue
factor, factor VI1 initiates blood coagulation by virtue of its
ability to activate factor IX, as well as factor X.’,4Thrombin,
factor XIIa, factor Xa, factor IXa, and factor VIIa itself are
capable of activating factor VI1 to factor VIIa.”’”
Factor VI1 deficiency has an estimated incidence of one
per 500,000 in the general population and an autosomal recessive pattern of inheritance. The hemorrhagic predispositionin affected patients ishighly variable and correlates
poorlywith plasma factor VI1 activity levels.”~‘*Patients
with levels less than 1% of normal can experience severe
bleeding episodes, including hemarthroses and crippling arthropathy comparable to patients with the classic hemophilias.
The laboratory evaluation of factor VI1 deficiency relies
on assays of plasma factor VI1 coagulant activity (V1I:C)
using animal thromboplastins, as well as immunologic quantitation of factor VI1 antigen.’.I3 Patients have been categorized as to the plasma level of factor VI1 antigen (V1I:Ag)
or cross-reacting material (CRM-, low or absent antigen;
CRMR, reduced antigen; C M + ,normal antigen). CRMdefects presumably result from defective factor VI1 biosynthesis or accelerated clearance in vivo, CRM+ defects are
Fromthe Hematology-Oncology Section, Department of Medicine, Brockton-West Roxbury Department of Veterans Affairs Medical Center, and Beth Israel Hospital, Harvard Medical School, Boston, MA; A. BianchiBonomi Hemophilia and Thrombosis Center,
Institute of Internal Medicine, IRCCS Ospedale Maggiore, University of Milan, Milan, Italy; and kboratory of Blood Coagulation,
Institute of Hematology and Blood Transfusion, Warsaw, Poland.
Submitted January 28, 1994; accepted June 6, 1994.
Supported by the Medical Research Service of the Department of
Veterans Affairs. K.A.B. is an Established Investigator of the American Heart Association.
Address reprint requests to Kenneth A. Bauer, MD, Department
of Veterans Affairs Medical Center, 1400 VFW Pkwy, West Roxbury,
MA 02132.
The publication costs of this article were defrayed in part by page
charge payment. This article must therefore be hereby marked
“advertisement” in accordance with 18 U.S.C. section 1734 solely to
indicate this fact.
0 1994 by The American Society of Hematology.
0006-497I/94/8407-06$3.00/0
2214
MATERIALS AND METHODS
Patients. Patients from families of Polish ancestry with factor
VI1 deficiency were investigated (Table 1). Eight of the patients (B
through H and J) resided in Poland, and were followed by physicians
at the Institute of Hematology and Blood Transfusion in Warsaw,
Poland. Two patients (A and I) reside in the United States.
The hemorrhagic symptoms of the patients with factor VI1 deficiency were categorized as follows: mild, a definite history of excessive bleeding following hemorrhagic challenges that did not require
clinical intervention (eg, easy bruising, epistaxis, mild to moderate
menorrhagia, moderately delayed bleeding following tooth extractions); moderate, recurrent bleeding following hemostatic challenges
or menstrual periods requiring transfusion of red bloodcells or factor
VII-containing products; and severe, one or more spontaneous hemorrhages (eg, joints, deep muscles). None of the patients had histories
of intracranial hemorrhage.
Collection and processing of blood samples. Bloodwas collected by atraumatic venipuncture into plastic tubes containing 1/10
volume 0.129 molL buffered trisodium citrate. Plasma was obtained
by centrifugation at 2,0001: for 15 minutes at 4°C. transferred into
plastic tubes, and stored along with the cellular elements at -80°C
until use.
Factor VI1 assays. Plasma VII:C levels were measured by onestage clotting assay using rabbit brain thromboplastin (Automated
Simplastin; Organon Teknika Corp. Durham. NC), recombinant huBlood, Vol 84, No 7 (October l), 1994 pp 2214-2220
From www.bloodjournal.org by guest on December 22, 2014. For personal use only.
MOLECULARANALYSISOFFACTOR
2215
VI1 DEFICIENCY
Table 1. Hemostatic Characteristicsof Patients From 10 Polish
Kindreds With Factor VI1 Deficiency
Patient
Sex/Age
~
A
B
C
D
E
F
G
H
Father
Mother
Sibling
I
Father
Mother
Sibling
J
Father
Mother
Bleeding
VII:C (%l
(rabbiVhumanI
VII:Ag 1%)
~~~
F140
M/31
F152
F126
F119
F147
MI39
MI19
MI46
F148
MI16
F122
MI53
F150
F130
MI33
MI62
F160
Moderate
Moderate
Moderate
Severe
Moderate
Moderate
Moderate
Moderate
Absent
Mild
Absent
Moderate
Absent
Absent
Moderate
Moderate
Absent
Absent
<2/<2
<2/<2
<2/<2
<2/<2
<2/<2
<2/<2
<2/<2
<2/<2
44/40
44/52
27/38
413
36/40
62/60
414
1115
30145
75/73
2
3
3
3
1
1
4
2
45
43
34
17
42
56
12
47
50
82
VII:C and VII:Ag results are expressed as percentage of normal
plasma pool. Rabbit and human refer to the type of thromboplastin
used for the VII:C measurements.
man tissue factor (RecombiPlasTin; Ortho Diagnostic Systems, Inc,
Raritan, NJ), and bovine brain thromboplastin (Thrombotest; Immuno AG, Vienna, Austria). Unless otherwise stated, VI1:C refers
to levels obtained using rabbit thromboplastin. Plasma VI1:Ag levels
were determined with an enzyme-linked immunoabsorbent assay
using a commercially available kit (American Bioproducts CO,Parsippany, NJ). A normal plasma poolwas constructed by mixing
equal volumes of plasma from greater than 30 control subjects. This
population consisted of healthy laboratory and medical personnel
between the ages of 20 and 50 years, who gave a negative history
for bleeding, as well as thrombosis, and were not taking any medications at the time of sample collection.
DNA isolacion and in vitro amplification using polymerase chain
reaction. DNAwaspurified from leukocyte nuclei obtained from
whole blood by standard techniques.*’ Oligonucleotides containing
intronic sequences2*flanking exons l a through 8 were synthesized
on an Applied Biosystems 381A DNA Synthesizer (Foster City,
CA). Polymerase chain reaction (PCR) amplifications were performed using a DNA Thermal Cycler (Perkin Elmer Cetus, Norwalk,
CT).” For single-strand conformation polymorphism (SSCP) analysis, PCR products were generated in 20-pL reaction mixtures that
initially contained 160 ngof genomic DNA, 0.4 U of Taq DNA
polymerase (Perkin Elmer Cetus), oligonucleotide primers at a concentration of 0.5 pmollL each, dNTPs at a concentration of 100
pmollL each, 2 pCi of [(Y-’~P]dCTP (3,000 mCi/mmol, 10 mCil
mL; New England Nuclear, Billerica, MA), 1 to 1.5 mmoUL MgC12,
10 mmol/L Tris-HCI, pH 8.3 (at 25”C), 50 mmol/L KCl, and 0.01
mglmL autoclaved gelatin. For subcloning, sequencing, or performing restriction enzyme analysis, PCR products were amplified
in 50-pL reaction mixtures with the aforementioned reaction components excluding radiolabeled dCTP. Each sample was subjected to
30 cycles of denaturation (30 S at 95°C). annealing, and extension
(30 S at 72°C). A 5-minute denaturation step in the first cycle, a 1minute denaturation step in the next three cycles, and a 5-minute
extension step in the last cycle were always included.
For amplifying the exons of the factor VI1 gene, the sequences
of the sense and antisense oligonucleotide primers, the sizes of the
product obtained, and the annealing temperatures of the PCR reac-
tion, respectively, were as follows: la, 5’-ACAGGCAGGGGCAGCACTGC-3‘, 5’-ACCAAGmATGGAGAAAAC-3’, 159 bp, 50°C;
5”CCACGCGGCCTG2, 5’-GGCGGTCTCCGAGGCACTGG-3’,
G’ITCAC-3’; 288 bp,70°C; 3 + 4, 5”GC’ITACCGTTGGGTGCTCTG-3’, 5”G’ITGGCCAGGCCACCTCCAC-3’, 339 bp,60°C;
5, 5’-AGCTCATGCCACClTCCAGGC-3’, 5”CTAGTGGGACAGGGACTGGT-3’, 314 bp, 55°C; 6, 5”GCCTCXCAGAGGATGGGTGTT-3’, 5“TGGAGCTG’lTG‘lTCACATTCA-3’, 273 bp, 55°C; 7,
5‘-CAGAGAAACAATGACAGCAAT-3’, 5”GATGTCTGTCTGTCTGTGGA-3‘, 470 bp, 55°C;8.5”AATGGCCACAGCCCATCC3’, 5’-CCAGGACAGTTCGACGCA-3‘, 732 bp, 54°C.For the SSCP
analysis of exon 8, additional sets of internal primers were synthesized to obtain two overlapping PCR fragments encoding this domain
of the factor VI1 molecule (8a, 8b). These products were made in
secondary PCR reaction mixtures containing 0.02 pmol of the fragment encoding exon 8, which had been previously generated from
genomic DNA and purified from low melt agarose. The sense and
antisense oligonucleotides, PCR product sizes, and annealing temperatures,respectively,wereasfollows: 8% 5”AATGGCCACAGC305bp, 54OC;
CCATCC-3’, 5”CCGCTGACCAATGAGAAGCG-3’,
8b, 5’-CGCTTCTCA’ITGGTCAGCGGC-3’, 5”CCACAGGCCAGGGCTGCTGG-3’, 414 bp, 64°C. All primer sets were annealed for 1
minute except exon 2, for whicha 2-minute annealing cycle wasused.
SSCP analysis. The method of SSCP analysis described by Orita
et a124.2S was used to screen for unknown mutations in the factor VI1
gene with the following modifications. We ensured that the purity
and quantity of the various fragments generated by PCR were adequate by electrophoresing an aliquot in a 1.5% (wt/vol) agarose gel
containing ethidium bromide and visualizing it under UV transillumination. A small quantity (1 to 2 pL) of the PCR reaction mixture
was mixed with 9 pL of a solution containing 95% (wt/vol) formamide, 10 mmollL NaOH, 0.05% bromophenol blue, and 0.05%
xylene cyanol. Immediately before electrophoresis, samples were
heat-denaturated at 95°C for 2 minutes and placed on an ice bath
for 15 minutes. Three microliters were then loaded onto a 35 cm X
45 cm X 0.4 mm gel cast using Hydrolink-MDE Gel (AT Biochem,
Malvern, PA) in 54 mmol/L Tris-borate, pH 8.3, and 1.2 mmol/L
Electrophoresis was performed at 6 to 8 W constant
power at room temperature for 14 to 20 hours. The gels were then
transferred onto Whatman 3M paper (Whatman International Ltd,
Markstone, England), dried under vacuum at 70°C for 1 hour, and
exposed to x-rayfilm (XAR-5; Kodak, Rochester, NY) at -80°C
for 36 hours using an intensifying screen.
Cloning and sequencing of PCR fragments. The amplified PCR
fragments were gel purified from low-melt agarose and ligated into
PT7Blue T-vectors (Novagen, Madison, WI). The inserts were sequenced by the dideoxy chain termination method2*on an Applied
Biosystems 373A DNA Sequencer. Allthe mutations were confirmed in at least two independent clones. Sequence analyses were
performed using the GCG Sequence Analysis Software Package
(Genetics Computer Group, Inc. Madison, WI) from the Molecular
Biology Computer Research Resource (Boston University, Boston,
MA).
Restriction enzyme analysis. Restriction enzyme digestion of
PCR fragments was used to depict segregation patterns of mutations
that were found by sequencing to introduce or abolish a restriction
site. All restriction enzymes andDNA molecular weight markers
were purchased from New England Biolabs (Beverly, MA). Approximately 200 ng of PCR product purified from low-melt agarose were
digested with 5 U of enzyme in a final volume of 20 pL. Restriction
fragments were extracted with phenol-chloroform, and 10-pL aliquots were electrophoresed in a 6% (wt/vol) polyacrylamide gel
containing 1 m o w urea. After completion of the electrophoretic
procedure, gels were stained with ethidium bromide andphotographed under UV transillumination. For fragments, in which restriction enzymes created products under 40 bp, the products were radio-
From www.bloodjournal.org by guest on December 22, 2014. For personal use only.
2216
ARBlNl ET AL
labeled with [CY-’~P]
dCTP in the PCR reaction as described above.
After completion of the electrophoretic procedure, these gels were
fixed, dried, and autoradiographed for 24 hours at -80°C using an
intensifying screen.
Allele speciJc PCR. Primer-directed allele specific amplificationZ9was used to depict mutations that could not be tracked using
restriction enzymes. Fragments spanning a mutation site were amplified by PCR and used astemplates for allele-specific PCR (ASPCR).
Oligonucleotides of 14 bp containing the mutant or normal base at
its 3’ end were used with a common second primer at concentrations
of 0.3 pmol/L. PCR reactions were performed in a total volume of
20 pL with 0.02 pm01 template, 0.4 U Taq DNA polymerase, dNTPs
at a concentration of 70 ,umol/L each, and MgClz at 1.0 mmol/L.
Optimal annealing temperatures for each set of primers were previously determined, and amplifications were carried out for 25 cycles. After completion of the PCR, 6 pL ofthe reaction mixture
were electrophoresed in a 1.5% agarose gel containing 0.5 pg/mL
ethidium bromide and photographed under UV transillumination.
Informed consent. Informed consent was obtained from the patients. The study was approved by the Human Studies Committee
of Brockton-West Roxbury Department of Veterans Affairs Medical
Center, Beth Israel Hospital (Boston, MA), and the Institute of Hematology and Blood Transfusion (Warsaw, Poland).
RESULTS
We have investigated the molecular defects underlying
factor VI1 deficiency in patients from 10 kindreds of Polish
ancestry (Table 1). One affected patient from each family
was studied in detail. Patients A through H had moderate to
severe bleeding diathesis and levels of plasma VI1:C and
VI1:Ag that were less than 4%of normal. Patients I and J
exhibited a milder hemorrhagic diathesis than the other patients, andhadVII:C levels of 4% and l l % and V1I:Ag
levels of 17% and47%, respectively. Similar VI1:C measurements were obtained using rabbit brain thromboplastin or
human tissue factor (Table 1) or bovine brain thromboplastin
(data not shown). We were also able to investigate both
parents of patients A, I, and J, and their results are also
shown in Table 1. None of the 10 patients were known to
be related, and a history of consanguinity was obtained only
from patient I, in which her parents were first cousins.
The SSCP te~hnique’~.*~
was used to screen the exons of
the factor VI1 gene for sequence alterations. This method is
based on the observation that single-stranded DNA molecules, differing in sequence by as little as a single base, can
migrate differently on polyacrylamide gels under nondenaturing conditions. However, the method can lose sensitivity
when large PCR fragments are used, and the coding sequence
Table 2. Coagulation Data and Genetic Alterations in the Factor
VI1 Gene of Polish Patients With Factor VI1 Deficiency
11125-
11128
Patient
<4A-H
I
J
VII:C (%)
VII:Ag (%)
<2
4
11
17
47
Ala294Val
Arg353Gln
Frameshih
+l+
+l+
+l+
+I+
-l+l+
+It
+It
-1-
VII:C and VII:Ag resultsareexpressedaspercentage
of normal
plasma pool. VII:C values refer t o those obtained using rabbit thromboplastin. Presence or absence of the sequence alteration i n each
allele is denoted by + or - signs, respectively.
of exon 8 is almost 600 bp in length. Therefore, exon 8 was
amplified by PCR after subdividing it into two overlapping
pieces of 305 bp (8a) and 414 bp (8b). In all 10 patients,
we identified electrophoretic migration patterns distinct from
those of normal individuals in the second portion of exon 8
(8b), as well as exon 5. For exon 8b, patients A through
H demonstrated the same electrophoretic pattern, whereas
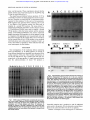
patients I and J had their own distinctive pattern (Fig 1j.
These findings in conjunction with the levels of VII:C and
VI1:Ag suggested that patients A through H had the same
molecular defect, whereas patients I and J were different.
To determine the molecular abnormalities in exon 8, PCR
fragments encoding the entire exon from patients H, I, and
J were subcloned and sequenced (Table 2). The following
alterations were identified in two independent clones from
patient H: a C to T transition at position 10798 in the gene
(GCC to GTC) resulting in Ala294Val; a G to A transition
at position TO976 (CGG to CAG) resulting in Arg353Gln;
and a single base deletion of ;C between positions 1 1 125
through 11 128(GCC CCA to GCC CAT). This deletion
causes a frameshift mutation in the triplet encoding P1-0404,
and abolishes the TAG stop codon following amino acid
406. Two independent clones from patient I demonstrated
the same abnormalities at positions 10798 and11125 to
11128, but did not have the substitution at position 10976
resulting in Arg353Gln. Four independent clones from patient J showed sequence alterations resulting in Ala294Val
and Arg353Gln, but he did not have the single bp deletion
at position 11125 to 1 1 128. Clones encoding exon 8 from
patients A and B demonstrated sequences that were identical
to patient H (Table 2).
For exon 5 , we observed that the abnormally migrating
bands on SSCP analysis in patients A through J were present
in three normal subjects, who also displayed bands with
the normal electrophoretic pattern (datanot shown). This
suggested the presence of a sequence polymorphism in homozygous form, whichwasconfirmed by subcloning and
sequencing PCR fragments containing this exon from patients H, I, andJ. A C to T transition was observed at position
7880 in the gene, which isa neutral dimorphism in the codon
for His1 15.30Sequencing was then performed on subcloned
PCR fragments containing the exons la, 2, 3 + 4, 5 , 6, and
7 in patients H, I, and J, and no additional substitutions were
detected.
We next examined if patients C through G had the same
sequence alterations as patients A, B, and H, as was suggested by the SSCP analysis. To accomplish this, we used
the fact that the C to T transition at position 10798 creates
a new AvuII restriction site, and the G to A transition at
10976 abolishes a MspI restriction site. Restriction digests
of the PCR product containing the second portion of exon
8 (8bj demonstrated that patients A through G, as well as
J, all had the Ala294Val (Fig 2, Table 2) and Arg353Gln
substitutions (Fig 3). In patients H and J, homozygosity for
these sequence alterations was proven by performing restriction enzyme digests with AvuII (Fig 2) and MspI (Fig 3,
Table 2) on parental DNA. The brother of patient H was
heterozygous for the two sequence alterations. A similar
series of restriction enzyme digests were carried out on PCR
fragments of exon 8b obtained from patient I, her affected
”
“
From www.bloodjournal.org by guest on December 22, 2014. For personal use only.
MOLECULARANALYSISOFFACTOR
VI1 DEFICIENCY
2217
sister, and her parents. These experiments showed that the
patient and her sister were homozygous for Ala294Val (Fig
2), but not Arg353Gln (Fig 3).
The single base-pair deletion between positions 1I125 to
11 128 did not result in an altered restriction enzyme recognition site. Therefore, we used ASPCR29
to demonstrate homozygosity for this mutation. To this end, we first generated a
PCR fragment from genomic DNA containing all of exon
8. An aliquot of this reaction mixture was thenused as
template in a second PCR reaction. The sense primer 5’CGCTTCTCATTGGTCAGCGGC and the antisense primer
5”CTAGGGAAATGGGG were used to amplify a normal
395-bp product, while the former primer and the antisense
primer 5”CTAGGGAAATGGGC were used to separately
amplify the mutant 394 bp product. In all our patients, except
J, a fragment of the appropriate size was generated only with
the primers designed for the mutant allele, but not withthose
for the normal allele (Fig 4). PCR products were produced
with both primer sets in the parents of patients H and I, as
well as the brother of patient H, which indicated that they
were heterozygous for the deletion.
15
B
FAMILY H
FAMILY I
FAMILY J
DISCUSSION
Our investigations of the molecular defects underlying
factor VI1 deficiency in 10 Polish patients (Athrough J )
from different kindreds have identified four alterations in the
coding sequence of the gene. These include substitutions of
Val for Ala and Gln for Arg at amino acids 294 and 353,
respectively, of the gene product. A single base-pair deletion
near the carboxy-terminus of factor VI1 produces a
N I N ~ N ~ N ~ NBs AC D E
F G H I J
1
348
36
30
15
Fig 2. Avdl restriction enzyme analysisshowing the presence of
Ala294Val. Radiolabeled PCR fragments of 414 bp were amplified
from genomic DNAusing the primers for the second portion of exon
8 (8b). After digestion with Avall, the products were subjected to
electrophoresis in a 6% polyacylamide gel and autoradiographed.A
restriction digest of a normal individual (N) is shown in the last lane
of each gel. This fragment normally has two Avall restriction sites
resulting in fragments of 348, 36, and 30 bp.
A C to T transition at
position 10798 producesa new digestion site in the 30-bp fragment
resulting in two 15-bp fragments. Heterozygosityis indicated bythe
presence ofboth the 30 bp and the 15 bp fragments, while homozygosity results in the absence ofthe 30-bp product. In (A), patients A
through G show homozygosity for Ala294. In (B), completely blackened circles and squares
in the pedigrees denoteindividualswho are
homozygous for the sequence alteration, while heterozygotes are
partially blackened. Patients H, 1, and J are homozygous for the defect, as is the sister of patient l. The parents of these patients and
the brother of patient H are heterozygous.
Fig 1. SSCP analysis of PCR products containing the second portion of exon 8 (8b). Fragments of414 bp (position 10746 t o position
11159 bp in the factor VI1gene) were amplified from five normal
individuals and l 0 Polish patients with factor VI1 deficiency(A
through J1. Patients A through H displayed two major bands that
migrated differently from the normal group. PatientsI and J showed
migration patterns that were distinct from patients A through H and
the normal controls.
frameshift mutation that is predicted to add 28 additional
amino acids to the protein. We also detected a neutral dimorphism in the codon for His1 15 in all of our patients.
Patients A through H were homozygous for Ala294Va1,
From www.bloodjournal.org by guest on December 22, 2014. For personal use only.
ARBlNl ET AL
2218
A
A B C D E F G J N
253
186
80-81
67
B
FAMILY OF
PATIENT H
FAMILY OF
PATIENT I
PARENTS OF
PATIENT J
IK)
N
253
186
80-81
67
Fig 3. Mspl restriction enzyme analysis showing
the presence of Arg353Gln. PCR fragments of 414 bp
were amplifiedfromgenomic DNA using the primers
for thesecond portion of exon (8b).
8 After digestion
with MS@,the products were subjected t o electrophoresis in a 6% polyacylamide gels and stained with
ethidium bromide. A restriction digest of a normal
individual (N) is shown in the last lane of each gel.
There are normally three restriction sites resulting
in four fragments of186,81,80, and 67 bp. The G t o
A substitution at position 10976 abolishes the restriction site between the
186-bp and 67-bp fragments, resulting in a new fragment of 253bp. In
(A), restriction digests of patients A throughG and
J show homozygosity for Arg353Gln. In (B),completely blackened circles and squares in the pedigrees denote individuals who are homozygous for
the sequence alteration, while heterozygotes are
partially blackened. Patient H is homozygous for
Arg353Gln, as is the fatherof patient J. The parents
and brother of patient H, and the motherof patient
J are heterozygous. Patient I and her family have the
normal sequence at this restriction site. Note that
the 81-bp and 80-bp fragments run very closely t o
one another. The DNA molecular weight markers are
an Haelll digest of 6x174.
FAMILY
1.r
A
Fig 4. ASPCR analysis showing thepresence of a
deletion of a single C between positions 11125 t o
11128 of the factorVI1 gene. Completely blackened
circles and squares in the pedigrees denote individuals who are homozygous for thedeletion, while heterozygotes are partially blackened. PCR fragments
of 658 bp spanning all of exon 8 were first
generated
from genomic DNA, and used as templates in the
subsequent ASPCR reactions. The sense primer used
t o amplify the second portion of exon 8 (8b) was
used with 14-mer primers containing the normal or
the mutant sequence at its 3' ends. The expected
size of the fragments amplified from the normal and
mutant alleles were 395 and 394 bp, respectively.
Amplification products were analyzed in 1.5% agarose gels containing 0.5 pg/mL ethidium bromide.
The result in a normal individual (N) with each set
in the lastlane of each gel. In (A),
of primers is shown
using the primers for the normal
allele, fragments of
the correct size were amplified in patient J, the parents of patients H and 1, and the brother of patient
H. In (B),using the primers for the mutant
allele,
fragments of the correct size were generated in all
patients except J, and in the parents and siblings of
patients H and 1. The DNA molecular weight markers
are an Haelll digest of &X174.
FAMILY
I
A B C D E F G J
395
B
FAMILY
H
A B C D E F G J &&N
FAMILY
I
From www.bloodjournal.org by guest on December 22, 2014. For personal use only.
MOLECULARANALYSIS
2219
OF FACTOR VI1 DEFICIENCY
Arg353Gln, and the frameshift mutation, and all had a moderate to severe bleeding diathesis with levels of VII:C and
VII:Ag that were generally less than 2%. While these individuals resided in different areas of Poland and are known
to be related, it is highly likely that a founder effect is responsible for the high frequency of this genotype. Patient I and
her sister were homozygous for Ala294Val and the
frameshift mutation, but not Arg353Gln. These women were
moderate bleeders with levels of VII:C and VII:Ag of4%
and 12% to 17%, respectively. It is tempting to speculate
that they had a distant ancestor in common with the former
group of patients. This would have required a transition
within a CG dinucleotide, which are known mutation hotspots,3’in the codon for amino acid 353 of a single ancestor
of patient I. Patient J was homozygous for Ala294Val and
Arg353Gln, but did not have the frameshift mutation. He
had a moderate bleeding history with levels ofVI1:C and
V1I:Ag of 11% and 47%, respectively. Based on the above
data, we conclude that the frameshift mutation in the codon
for amino acid 404 is associated with marked reductions in
VII:C, Arg353Gln can decrease plasma levels of factor VI1
in the presence of other mutations in the factor VI1 gene, and
Ala294Val results in a dysfunctional factor VI1 molecule.
From a previous study seeking to identify sequence alterations associated with differences in VI1:C levels, it is known
that Arg353Gln is a polymorphism associated with a modest
reduction in VII:C.32It occurs with an allelic frequency of
0.1 in normal individuals in theUnitedKingdomand
the
United States.’2333 Individuals heterozygous
for the Arg353Gln
allele have a significant reduction in their mean VII:C levels
to 75% of normal, while subjects homozygous for this polymorphism have levels of approximately 50%. It is unclear
whether these individuals have VI1:C levels that are reduced
relative to V1I:Ag. Green et a13’ postulated that Arg353Gln
induces an alteration in charge leading to a conformational
change in the factor VI1 molecule, which affects intracellular
processing, and reduces hepatic secretion or accelerates its
clearance rate from the circulation.
It should be noted that patient J and his father were both
homozygous for Arg353Gln and had V1I:Ag levels of approximately 50%. Since this patient and his father are homozygous and heterozygous for Ala294Va1, respectively, our
data suggests that this substitution has little effect on plasma
VI1:Ag measurements.
To understand the effect that Ala294Val and Arg353Gln
might have on factor VI1 function, we compared the amino
acid sequences of factor VI1 with those of human factor IX,34
factor X,35pr~thrombin,’~
protein C,37and chymotrypsinoged8 using the GCG software package. The aligned sequences were analyzed using a model for trypsin-like domains of bovine coagulation serine proteases proposed by
Furie et al.39This model is based on the known three dimensional structure of pancreatic serine proteases and sequence
homologies between these enzymes and serine proteases involved in blood coagulation. It recognizes seven constant
regions (CR) and six variable regions (VR) that are generally
well preserved in chymotrypsin, trypsin, thrombin, factor
Xa, and factor IXa.
According to the model for factor IX, Ala294 is one of
eight amino acids in VR4. The predicted location of this
region is on the surface of the molecule surrounding the
active site where it may play an important role in substrate
r e ~ o g n i t i o n . Evidence
~~.~
supporting this putative function
can also be inferred from the fact that substitution of the
factor VI1 residue homologous to Ala294 in factor IX,
Ala320, by the polar amino acid Asp results in a complete
loss of factor IX f~nction.~’
The patient with this mutation
had levels of factor IX coagulant activity and factor IX antigen of less than 1% of normal and 90%, respectively. The
substitution of Ala294 in factor VI1 with another nonpolar
amino acid, such as Val, is a relatively conservative change,
and might be expected to have a moderate effect on the
function of the protein.
The Arg353 residue is located within a group of 10 residues with limited homology denoted as VR6. This region is
also predicted to be on the surface of the molecule, but
distant from the active site, suggesting that it may not
be
involved in the substrate recognition process. To date, substitutions affecting four different residues have been reported
to occur in VR6 of factor IX, and all of these patients had
concordant reductions in their plasma level of factor IX coagulant activity and factor IX antigen.4’
The purification and biochemicalcharacterization of
mutant factor VI1 proteinscontainingAla294Val
and
Arg353Gln from stably transfected cell lines will ultimately allow us to definitively address the functional importance of the various sequence abnormalities.
ACKNOWLEDGMENT
We thank Dr Yale Nemerson (Mt Sinai School of Medicine, New
York, NY) for facilitating this collaborative study of Polish patients
with factor VI1 deficiency.
REFERENCES
1. Hagen FS, Gray CL, O’Hara P, Grant FJ, Saari G C , Woodbury
RC, Hart CE, Insley M, Kisiel W, Kurachi K, Davie EW: Characterization of a cDNA coding for human factor VII. Roc Natl Acad Sci
USA 83:2412, 1986
2. Fair DS: Quantitation of factor VI1 intheplasma of normal
and
warfarin-treated
individuals
by
radioimmunoassay.
Blood
62:784, 1983
3. Osterud B, Rapaport SI: Activation of factor IX by the reaction
product of tissue factor and factor VII: Additional pathway for initiating blood coagulation. Proc Natl Acad Sci USA 745260, 1977
4. Bauer KA, Kass BL, ten Cate H, Hawiger JJ, Rosenberg RD:
Factor IX is activated in vivo by the tissue factor mechanism. Blood
76:731, 1990
5. Radcliffe R, Nemerson Y: The activation and control of factor
VI1 by activated factor X and thrombin. Isolation and characterization of a single chain form of factor VII. J Biol Chem 250:388, 1975
6. Kisiel W, Fujikawa K, Davie EW: Activation of bovine factor
VI1 (proconvertin) by factor XIIa (activated Hageman factor). Biochemistry 16:4189, 1977
7. Radcliffe R,Bagdasarian A, Colman R, Nemerson Y: ActivaBlood
tion of bovine factor VI1 byHagemanfactorfragments.
50:611, 1977
8. Seligsohn U, Osterud B, Brown SF, GriffinJH,Rapaport SI:
Activation of human factor VI1 in plasma and in purified systems.
Roles of activated factor IX, kallikrein, and activated factor XII. J
Clin Invest 64:1056, 1979
9. Nakagaki T, Foster DC, Berkner KL, Kisiel W: Initiation of
the extrinsic pathway of blood coagulation: Evidence for the tissue
From www.bloodjournal.org by guest on December 22, 2014. For personal use only.
2220
factor dependent autoactivation of human coagulation factor VII.
Biochemistry 30:10819, 1991
IO. Pedersen A, Lund-Hansen T, Bisgaard-Frantzen H, Olsen F,
Petersen LC: Autoactivation of human recombinant coagulation factor VII. Biochemistry 28:9331, 1989
11. Ragni MV, Lewis JH, Spero JA, Hasiba U: Factor VI1 deficiency. Am J Hematol 10:79, 1981
12. Triplett DA, Brandt JT, McGann Batard MA, Schaeffer Dixon
JL, Fair DS: Hereditary factor VI1 deficiency: Heterogeneity defined
by combined functional and immunochemical analysis. Blood
66:1284, 1985
13. Boyer C, Wolf M, Rothschild C, Migaud M, Amiral J, Mannucci PM, Meyer D, Larrieu MJ: An enzyme immunoassay (ELISA)
for the quantitation of human factor VII. Thromb Haemost 56:250,
l986
14. Girolami A, Falezza G, Patrassi G, Stenico M, Vettore L:
Factor VI1 Verona coagulation disorder: Double heterozygosis with
an abnormal factor VI1 and heterozygous factor-VI1 deficiency.
Blood 50:603, 1977
15. Girolami A, Fabris F, Dal Bo Zanon R, Ghiotto G, B u d A:
Factor VI1 Padua: A coagulation disorder due to an abnormal factor
VI1 with a peculiar activation pattern. J Lab Clin Med 91:387, 1978
16. Girolami A, Cottarozzi G, Dal Bo Zanon R, Cella G, Toffanin
F: Factor VI1 Padua2: Another factor VI1 abnormality with defective
ox brain thromboplastin activation and a complex hereditary pattern.
Blood 54:46, 1979
17. O'Brien DP, Gale KM, Anderson J, McVey JH, Miller GJ,
Meade W ,Tuddenham EGD: Purification and characterization of
factor VI1 304-Gln: A variant molecule with reduced activity isolated
from a clinically unaffected male. Blood 78: 132, 1991
18. Takamiya 0, Kemball-Cook G, Martin DMA, Cooper DN,
von Felten A, Meili E, Hann I,Prangnell DR, Lumley H, Tuddenham
EGD, McVey JH: Detection of missense mutations by single-strand
conformational polymorphism (SSCP) analysis in five dysfunctional
variants of coagulation factor VII. Hum Mol Gen 2:1355, 1993
19. Bernardi F, Liney DL, Patracchini P, Gemmati D, Legnani
C, Arcieri P, Pinotti M, Redaelli R, Ballerini G, Pemberton S, Wacey
AI, Mariani G, Tuddenham EGD, Marchetti G: Molecular defects
in CRM+ factor VI1 deficiencies: Modelling of missense mutations
in the catalytic domain of FVII. Br J Haematol 86:610, 1994
20. Matsushita T, Kojima T, Emi N, Takahashi I, Saito H: Impaired human tissue factor-mediated activity in blood clotting factor
VII,,,,,, (Arg""-Trp). J Biol Chem 269:7355, 1994
21. Kunkel LM, Smith KD, Boyer SH, Borgaonkar DS, Wachtel
SS, Miller OJ, Breg WR, Jonew HW, Rary JM: Analysis of human
Y chromosome-specific reiterated DNAin chromosome variants.
Proc Natl Acad Sci USA 74:1245, 1977
22. O'Hara PJ, Grant FJ, Haldeman BA, Gray CL, Insley MY,
Hagen FS, Murray MJ: Nucleotide sequence of the gene coding for
human factor VII, a vitamin K-dependent protein participating in
blood coagulation. Proc Natl Acad Sci USA 84:5158, 1987
23. Saiki RK, Gelfand DH, Stoffel S, Scharf SJ, Higuchi R, Horn
GT, Mullis KB, Erlich HA: Primer-directed enzymatic amplification
ofDNA with a thermostable DNA polymerase. Science 239:487,
1988
24. Orita M, Iwahana H, Kanazawa H, Hayashi K, Sekiya T:
Detection of polymorphisms of human DNA by gel electrophoresis
as single-strand conformation polymorphisms. Proc Natl Acad Sci
USA 86:2766, 1989
ARBlNl ET AL
25. Orita M, Suzuki Y, Sekiya T, Hayashi K: Rapid and sensitive
detection of point mutations and DNA polymorphisms usingthe
polymerase chain reaction. Genomics 5:874, 1989
26. Perry DJ, Carrell RW: HydroLink gels: a rapid and simple
approach to the detection ofDNA mutations in thromboembolic
disease. J Clin Path 45: 158, 1992
27. Soto D, Sukumar S: Improved detection of mutations in the
p53 gene in human tumors as single-stranded conformation polymorphs and double-stranded heteroduplex DNA. PCR Methods and
Applications 2:96, 1992
28. Sanger F, Nicklen S, Coulsen AR:DNA sequencing with
chain terminating inhibitors. Proc Natl Acad Sci USA 74:5463, 1977
29. Wu DY, Ugozzoli L, Pal BK, Wallace RB: Allele-specific
enzymatic amplification of P-globin genomic DNA for diagnosis of
sickle cell anemia. Proc Natl Acad Sci USA 86:2757, 1989
30. Marchetti G, Ferrati M, Patracchini P, Redaelli R, Bernardi
F: A missense mutation ('7XCys-Tyr)and two neutral dimorphisms
('I5Hist and "?ier) in the human coagulation factor VI1 gene. Hum
Mol Gen 2: 1055, I993
31. YoussoufianH, Kazazian HH Jr, Phillips DG, Aronis S,
Tsiftis G, Brown VA,Antonarakis SE: Recurrent mutations in hemophilia A give evidence for CpG mutation hotspots. Nature 324:380,
1986
32. Green F, Kelleher C, Wilkes H, Temple A, Meade T, Humphries S: A common genetic polymorphism associated with lower
coagulation factor VI1 levels in healthy individuals. Arterioscler and
Thrombosis 11540, 1991
33. Humphries SE, Lane A, Dawson S, Green FR: The study of
gene-envirnoment interactions that influence thrombosis and fibrinolysis. Genetic variation at the loci for factor VI1 and plasminogen
activator inhibitor-l. Arch Pathol Lab Med 116: 1322, 1992
34. Kurachi K, Davie EW: Isolation and characterization of a
cDNA coding for human factor IX. Proc NatlAcadSciUSA
79:6461, 1982
35. Leytus SP, Chung DW, Kisiel W, Kurachi K, Davie EW:
Characterization of a cDNA coding for factor X. Proc NatlAcad
Sci USA 81:3699, 1984
36. Degen SJF, MacGillivray RTA, Davie EW: Characterization
of the complementary deoxyribonucleic acid and gene coding for
human prothrombin. Biochemistry 22:2087, 1983
37. Foster DC, Davie EW: Characterization of a cDNA coding
for protein C. Proc Natl Acad Sci USA 81:4766, 1984
38. Tomita N, Izumoto Y, Horii A, Doi S, Yokouchi H, Ogawa
M, Mori T, Matsubara K: Molecular cloning and nucleotide sequence
of human pancreatic prechymotrypsinogen cDNA. Biochem Biophys
Res Commun 158:569, 1989
39. Furie B, Bing DH, Feldmann RJ, Robison DJ, Bumier JP,
Furie BC: Computer-generated models of blood coagulation factor
Xa, factor IXa, and thrombin based upon structural homology with
other serine proteases. J Biol Chem 257:3875, 1982
40. Kumar A, Fair DS: Specific molecular interaction sites on
factor VI1 involved in factor X activation. Eur J Biochem 217:509,
1993
41. Giannelli F, Green PM, High KA, Sommer S, Lillicrap DP,
Ludwig M, Olek K, Reitsma PH, Goossens M,YoshiokaA,
Brownlee GG: Haemophilia B: Database ofpoint mutations and
short additions and deletions. Nucleic Acids Res 20:2027, 1992
(SUPPI)
From www.bloodjournal.org by guest on December 22, 2014. For personal use only.
1994 84: 2214-2220
Molecular analysis of Polish patients with factor VII deficiency
AA Arbini, D Bodkin, S Lopaciuk and KA Bauer
Updated information and services can be found at:
http://www.bloodjournal.org/content/84/7/2214.full.html
Articles on similar topics can be found in the following Blood collections
Information about reproducing this article in parts or in its entirety may be found online at:
http://www.bloodjournal.org/site/misc/rights.xhtml#repub_requests
Information about ordering reprints may be found online at:
http://www.bloodjournal.org/site/misc/rights.xhtml#reprints
Information about subscriptions and ASH membership may be found online at:
http://www.bloodjournal.org/site/subscriptions/index.xhtml
Blood (print ISSN 0006-4971, online ISSN 1528-0020), is published weekly by the American
Society of Hematology, 2021 L St, NW, Suite 900, Washington DC 20036.
Copyright 2011 by The American Society of Hematology; all rights reserved.