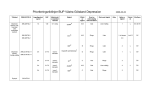
Survey
* Your assessment is very important for improving the work of artificial intelligence, which forms the content of this project
Digital Comprehensive Summaries of Uppsala Dissertations from the Faculty of Medicine 1115 Amoebae as Hosts and Vectors for Spread of Campylobacter jejuni JENNY OLOFSSON ACTA UNIVERSITATIS UPSALIENSIS UPPSALA 2015 ISSN 1651-6206 ISBN 978-91-554-9276-2 urn:nbn:se:uu:diva-255804 Dissertation presented at Uppsala University to be publicly examined in A1:111a, Biomedicinskt Centrum, Husargatan 3, Ing C7:2, Uppsala, Thursday, 10 September 2015 at 09:00 for the degree of Doctor of Philosophy (Faculty of Medicine). The examination will be conducted in Swedish. Faculty examiner: Docent Hans Lindmark (Livsmedelsverket, Uppsala). Abstract Olofsson, J. 2015. Amoebae as Hosts and Vectors for Spread of Campylobacter jejuni. Digital Comprehensive Summaries of Uppsala Dissertations from the Faculty of Medicine 1115. 50 pp. Uppsala: Acta Universitatis Upsaliensis. ISBN 978-91-554-9276-2. Campylobacter jejuni is the leading bacterial cause of gastrointestinal diarrheal disease in humans worldwide. This zoonotic pathogen has a complex epidemiology due to its presence in many different host organisms. The overall aim of this thesis was to explore the role of amoebae of the genus Acanthamoeba as an intermediate host and vector for survival and dissemination of C. jejuni. Earlier studies have shown that C. jejuni can enter, survive and replicate within Acanthamoebae spp. In this thesis, I have shown that C. jejuni actively invades Acanthamoeba polyphaga. Once inside, C. jejuni could survive within the amoebae by avoiding localization to degradative lysosomes. We also found that A. polyphaga could protect C. jejuni in acid environments with pH levels far below the range in which the bacterium normally survives. Furthermore, low pH triggered C. jejuni motility and invasion of A. polyphaga. In an applied study I found that A. polyphaga also could increase the survival of C. jejuni in milk and juice both at room temperature and at +4ºC, but not during heating to recommended pasteurization temperatures. In the last study we found that forty environmental C. jejuni isolates with low bacterial concentrations could be successfully enriched using the Acanthamoeba-Campylobacter coculture (ACC) method. Molecular genetic analysis using multilocus sequence typing (MLST) and sequencing of the flaA gene, showed no genetic changes during coculture. The results of this thesis have increased our knowledge on the mechanisms behind C. jejuni invasion and intracellular survival in amoebae of the genus Acanthamoeba. By protecting C. jejuni from acid environments, Acanthamoebae could serve as important reservoirs for C. jejuni e.g. during acid sanitation of chicken stables and possibly as vectors during passage through the stomach of host animals. Furthermore, Acanthamoeba spp. could serve as a vehicle and reservoir introducing and protecting C. jejuni in beverages such as milk and juice. Validation of the ACC method suggests that it is robust and could be used even in outbreak investigations where genetic fingerprints are compared between isolates. In conclusion, Acanthamoeba spp. are good candidates for being natural hosts and vectors of C. jejuni. Keywords: Amoebae-Bacteria interactions; Campylobacter jejuni; epidemiology; environmental stress; low pH; Acanthamoeba; coculture; intracellular trafficking; bacterial survival; beverages; ACC-method; enrichment; genetic stability; Trojan horse; reservoir; host; vector Jenny Olofsson, Department of Medical Sciences, Akademiska sjukhuset, Uppsala University, SE-75185 Uppsala, Sweden. © Jenny Olofsson 2015 ISSN 1651-6206 ISBN 978-91-554-9276-2 urn:nbn:se:uu:diva-255804 (http://urn.kb.se/resolve?urn=urn:nbn:se:uu:diva-255804) Till Mikael ”Inget är omöjligt. Det omöjliga tar bara lite längre tid.” Winston Churchill List of Papers This thesis is based on the following papers, which are referred to in the text by their Roman numerals. I. Axelsson-Olsson D, Svensson L, Olofsson J, Salomon P, Waldenström J, Ellström P, Olsen B. (2010) Increase in Acid Tolerance of Campylobacter jejuni through Coincubation with Amoebae. APPLIED AND ENVIRONMENTAL MICROBIOLOGY 76(13): 4194–4200. II. Olofsson J, Griekspoor Berglund P, Olsen B, Ellström P, Axelsson-Olsson D. (2015) The Abundant Free Living Amoeba, Acanthamoeba polyphaga, Increases the Survival of Campylobacter jejuni in Milk and Orange Juice. Submitted III. Olofsson J, Axelsson-Olsson D, Brudin L, Olsen B, Ellström P. (2013) Campylobacter jejuni Actively Invades the Amoeba Acanthamoeba polyphaga and Survives within Non Digestive Vacuoles. PLoSONE 8(11): e78873. IV. Griekspoor P, Olofsson J, Axelsson-Olsson D, Waldenström J, Olsen B. (2013) Multilocus Sequence Typing and FlaA Sequencing Reveal the Genetic Stability of Campylobacter jejuni Enrichment during Coculture with Acanthamoeba polyphaga. APPLIED AND ENVIRONMENTAL MICROBIOLOGY 79(7): 2477–2479. Reprints were made with permission from the respective publishers. Additional work Liljekvist-Soltic I, Olofsson J, Johansson K. (2008) Progenitor cell-derived factors enhance photoreceptor survival in rat retinal explants. BRAIN RESEARCH 1227: 226-233. Ellström P, Latorre-Margalef N, Griekspoor P, Waldenström J, Olofsson J, Wahlgren J, Olsen B. (2008) Sampling for low-pathogenic avian influenza A virus in wild Mallard ducks: Oropharyngeal versus cloacal swabbing. Vaccine 26(35): 4414-4416. Axelsson-Olsson D, Olofsson J, Ellström P, Waldenström J, Olsen B. (2009) A simple method for long-term storage of Acanthamoeba species. Parasitol Research 104(4): 935-937. Axelsson-Olsson D, Olofsson J, Svensson L, Griekspoor P, Waldenström J, Ellström P, Olsen B. (2010) Amoebae and algae can prolong the survival of Campylobacter species in coculture. Experimental Parasitology 126(1): 5964. Mohlin C, Liljekvist-Soltic I, Olofsson J, Johansson K. (2011) Chapter VIII: Neuropathology of Cultured Retinas: Degenerative Events and Rescue Paradigms. Advances in Eye Research 2: 177-190. Contents Introduction ................................................................................................... 11 Campylobacter ......................................................................................... 11 C. jejuni ............................................................................................... 11 C. jejuni and environmental stress ....................................................... 13 C. jejuni detection and species identification ...................................... 14 Multilocus sequence typing (MLST) ................................................... 15 Protozoa/Unicellular eukaryotes .............................................................. 15 Amoeba................................................................................................ 15 Amoeba-Bacteria interactions ............................................................. 16 Acanthamoeba ..................................................................................... 16 Intracellular trafficking pathways in Acanthamoeba ........................... 18 Unicellular eukaryotes as reservoir for Campylobacter ........................... 19 The ACC – method .............................................................................. 20 Aims .............................................................................................................. 22 Methods ........................................................................................................ 23 Bacterial and amoebic cultures (Paper I-IV) ............................................ 23 Coincubation of C. jejuni with Acanthamoeba spp. in acidic environments or in beverages (Paper I, II) ............................................... 23 Pasteurization of C. jejuni cells co-incubated with A. polyphaga in milk and orange juice (Paper II) .......................................................... 24 Quantification of adhered/internalized C. jejuni into A. polyphaga (Paper I) ............................................................................................... 24 Determination of C. jejuni motility (Paper I) ...................................... 25 Uptake and intracellular survival of C. jejuni in A. polyphaga (Paper III) ............................................................................................................ 25 Coculturing of viable and heat killed C. jejuni with A. polyphaga...... 25 Determination of intracellular localization of viable and heat killed C. jejuni in A. polyphaga ..................................................................... 26 Investigation of the genetic stability of C. jejuni after enrichment in A. polyphaga (Paper IV) ............................................................................... 26 C. jejuni enrichment by A. polyphaga cocultures ................................ 26 MLST and flaA typing of C. jejuni isolates ......................................... 26 Results and discussion .................................................................................. 28 Coincubation of C. jejuni with Acanthamoeba spp. in acidic environments (Paper I) ............................................................................. 28 Evaluation of A. polyphaga as a Trojan horse for protection of C. jejuni in beverages (Paper II) ............................................................................. 30 Uptake and intracellular survival of C. jejuni in A. polyphaga (Paper III) ............................................................................................................ 31 Genetic stability of C. jejuni after enrichment in A. polyphaga (Paper IV) ............................................................................................................ 33 Conclusions ................................................................................................... 35 Future perspectives ....................................................................................... 36 Svensk sammanfattning (Swedish summary) ............................................... 37 Acknowledgements ....................................................................................... 39 References ..................................................................................................... 41 Abbreviations spp. CFU MALDI-TOF PCR PFGE BLAST MLST ST CC FLA ACC VBNC PYG HCl CTC flaA SVR PBS ANOVA Cia dot icm MFI Species Colony forming units Matrix-assisted laser desorption ionisation-time of flight Polymerase chain reaction Pulsefield gel electrophoresis Basic Local Alignment Search Tool Multilocus sequence typing Sequence type Clonal complexes Free-living amoebae Acanthamoeba–Campylobacter coculture Viable but non-culturable Peptone-yeast-glucose Hydrochloric acid Cyanoditolyl tetrazolium chloride Flagellin A Short variable region Phosphate buffered saline Analysis of variance Campylobacter invasion antigens Defect in organelle trafficking Intracellular multiplication Mean fluorescence intensity Introduction Campylobacter Campylobacter spp. are a leading cause of bacterial foodborne diarrheal disease both in industrialized countries and in the developing world, where it causes morbidity and mortality and accounts for significant healthcare costs (1-3). Campylobacter bacteria are small (0.2-0.8 μm x 0.5-5 μm) Gram negative curved rods with flagella at one or both ends of the cell (4). The Campylobacter genus includes approximately 18 species and subspecies (5-7), and most of them are microaerophilic and able to grow between 37 - 42˚C (4, 5). The most commonly isolated species from human cases of gastroenteritis are C. jejuni, C. coli and C. lari. However, C. jejuni is the single most common species constituting more than 90% of all human infections and the work in this thesis has focused on C. jejuni (8, 9). C. jejuni C. jejuni is highly infectious and a leading cause of human bacterial gastroenteritis throughout the developed world (10, 11). It has a very low infective dose and as little as 500-800 colony forming units (CFU) have been shown to cause clinical infection in humans (12, 13). Molecular mechanisms behind the infection process are still not well understood, but flagellamediated motility (14), bacterial attachment to intestinal epithelial cells (15), invasive capability (16) and toxin production (17) have been identified as virulence factors (4, 18). Symptoms appear two to four days after ingestion and include fever, vomiting, headaches and watery or bloody diarrhea with abdominal pain (19). The disease is normally resolved within one week, but severe post infectious complications including reactive arthritis, GuillainBarré-(GBS) and Miller Fischer (MFS) syndromes occasionally occur (8, 20). The GBS is a form of paralysis that can result in respiratory and severe neurological dysfunction, sometimes leading to death (21), while MFS is characterized by ocular muscle defects (22). Most patients recover from Campylobacter infections without any specific treatment other than replacing lost fluids and electrolytes. Some severe cases need antibiotic treatment but due to emerging antibiotic resistance in Campylobacter, antibiotics should be reserved for these cases (18, 23). The most common antibiotics used in the treatment of Campylobacter infections 11 are macrolides, such as erythromycin, and fluoroquinolones, such as ciprofloxacin (5, 24). Humans are infected by contaminated food, primarily undercooked chicken but also unchlorinated water and unpasteurized milk can contain Campylobacter (25). Recent studies have indicated that water is a more important risk factor for Campylobacter infection than previously thought. In the Nordic countries, contamination of drinking water and swimming in natural water sources has been identified as important risk factors for sporadic human campylobacteriosis (26-28). C. jejuni colonizes the intestinal tract of several warm-blooded animals including swine, cattle, sheep, companion animals and birds, where they can exist as a commensal organism without giving any symptoms (29). Wild birds have been found to be natural reservoirs of Campylobacter spp., (30-32) and are considered possible vectors for transmission of Campylobacter to poultry (33, 34). The transmission modes between hosts are poorly understood. The source of infection remains unknown for a large proportion of reported human cases, and the epidemiological pathways have not been fully elucidated, even for the contained environments of poultry stables (35). Poultry is recognized as the main source of human campylobacteriosis (36-38) and therefore reducing the levels of Campylobacter in poultry is an important step to decrease the number of human cases. Several hygiene practices have been implemented in broiler production facilities to reduce C. jejuni carriage in live birds. Such measures include hygiene barriers such as changing clothes before entering the broiler houses and disinfection of the interior of the building with acid between flock rotations (39, 40). These efforts may reduce the number of C. jejuni organisms, but the bacterium is still difficult to eradicate from contaminated farms, and subsequent outbreaks at the same farm are not rare (41). A possible explanation for this eradication problem is the existence of an environmental reservoir that provides shelter for C. jejuni. 12 Fig.1 C. jejuni, with permission from Chris Pooley (Electron and Confocal Microscopy Laboratory, Agricultural Research Service, U. S. Department of Agriculture). C. jejuni and environmental stress Despite the high prevalence of C. jejuni in many animals, this bacterium is generally regarded as a fragile organism. Unlike other foodborne pathogens, C. jejuni is highly sensitive to environmental stresses including acidity, disinfectants, oxygen exposure, osmotic stress, desiccation and heating (42, 43). Campylobacter jejuni grow optimally at pH 6.5-7.5 and cannot survive below a pH of 4.9 or above 9.0 (18). The sensitive nature of C. jejuni observed in vitro, stands in contrast to the fact that it is difficult to eradicate from the poultry production and that the bacterium is able to survive the acidic passage through the human stomach (fasting conditions pH ~2.0, but may range from 1.5 to 5.5 depending on food intake) to the lower intestinal tract, where infection is established (44, 45). This is illustrated by the very low infectious dose for both broiler chickens (46, 47) and humans (12, 13) and indicates that the bacterium has developed strategies to avoid or withstand low pH in order to survive the transit. C. jejuni are sensitive to temperature changes, and below 30˚C they are incapable to grow. The fact that C. jejuni is both microaerophilic and has a narrow growth range, place severe restrictions on the ability of the organism to multiply outside an animal host. Consequently, it is not capable of multiplication in food during either processing or storage and cannot survive pasteurization and adequate cooking 13 (42, 48). Like many other pathogens C. jejuni can enter a viable but nonculturable (VBNC) state in response to stress. Bacteria in this state cannot be detected via conventional culturing methods (49). It has been questioned whether VBNC cells can survive until environmental conditions become more favorable for growth and cell division, or if the VBNC state is a stage on the way to irreversible cell death. Therefore, the role of the VBNC state in the survival and transmission of C. jejuni remains to be elucidated (50). Although C. jejuni is thought to account for as much as 400-500 million cases of gastroenteritis throughout the world every year (51), it is unclear how C. jejuni succeed to survive environmental conditions encountered during transmission between hosts. In the light of this, attention has been paid to the potential role of an environmental vector and/or reservoir for this pathogen. C. jejuni detection and species identification There are many methods available for detection and species determination of Campylobacter spp. including direct plating, biochemical testing, microscopy as well as newer methods including Polymerase chain reaction (PCR), sequence based methods and matrix-assisted laser desorption ionisation-time of flight (MALDI-TOF) mass spectrometry (52-56). The source and the condition of the sample are both of importance when selecting detection method. Typically, fecal samples from patients with acute campylobacteriosis contain large numbers of bacteria. At this stage of infection, direct plating on Campylobacter-selective agar plates is a convenient method used in most clinical laboratories. In contrast to fecal samples, isolation of Campylobacter from food or environmental water samples can be complicated since these samples often contain low numbers of bacteria as well as damaged cells. Therefore, detection of Campylobacter in non-clinical samples often involves an enrichment step using broth media (Preston or Bolton) before plating (57, 58). The development of different PCR based assays, which are accurate, cost effective and relatively easy to use, are promising tools for subtyping Campylobacter spp. The atpA based species identification method by Miller et al is a god example of such a new method (59-61). Following species identification, C. jejuni can be further subtyped for phylogenetic analysis. Two widely used methods are Pulsefield gel electrophoresis (PFGE), and serotyping using the Penner serotyping scheme (62, 63). However, Penner serotyping is an expensive and complex method and is nowadays largely replaced by other methods (64). In recent years, multilocus sequence typing (MLST) has become the method of choice for phylogenetic analysis of C. jejuni. 14 Multilocus sequence typing (MLST) Campylobacter jejuni MLST measures sequence variation in seven loci distributed in house-keeping genes (essential genes) across the chromosome. The seven loci used for C. jejuni MLST are (protein products in parentheses): aspA (aspartase A), glnA (glutamine synthetase), gltA (citrate synthase), glyA (serine hydroxymethyltransferase), pgm (phosphoglucomutase), tkt (transketolase) and uncA (ATP synthase alfa subunit) (65). Methodically, the nucleotide sequences (~500 bp) from each locus are amplified with PCR and the amplified products are purified and sequenced (65). The sequences are run through an international database, The Basic Local Alignment Search Tool (BLAST), (http://pubmlst.org/campylobacter/), where each sequence will generate an allelic number. The allelic profile composed of the seven loci is considered as a sequence type (ST). C. jejuni isolates with STs that share 4 or more alleles can be grouped into clonal complexes (CC), which demonstrate close genetic relationship (66, 67). Protozoa/Unicellular eukaryotes Molecular phylogenetic studies have increased the knowledge about evolutionary relationships of eukaryotes and there are now different opinions on how unicellular eukaryotes should be taxonomically ordered (68) (www.dyntaxa.se, http://eol.org/). The classical scheme of Bütschli (18801889), which grouped amoebae, sporozoa, flagellates and ciliates into protozoa, has been abandoned by protistologists for decades (68). However, the term protozoa is still frequently used in scientific articles as an umbrella term for a diverse group of single celled eukaryotic organisms. In this thesis I will use the term “protozoa” only when citing articles by others and instead use the term “unicellular eukaryotes”. Amoeba Free-living amoebae (FLA) are unicellular eukaryotes that are widely distributed in the environment and have been isolated from soil, air and water, including both natural and man-made water systems (69, 70). The majority of FLA are harmless, but there are some species that can cause disease in humans such as Entamoeba histolytica, Naegleria fowleri, Balamuthia mandrillaris, Sappinia diploidea and several Acanthamoeba species that can cause life threatening disease. For their diverse roles in the ecosystems and their role in causing serious and sometimes fatal human infections, freeliving amoeba have gained increased attention from the scientific and the medical community (71). 15 Amoeba-Bacteria interactions The importance of amoebae-bacteria interactions was brought to attention by T.J. Rowbotham during the early 1980s. He found that Legionella pneumophila, the causative organism of legionnaire´s disease, could colonize Acanthamoeba polyphaga, and that the bacteria benefited from the interaction (72). Today it is well established that such interactions can constitute ways for the bacteria to survive in the environment. This is illustrated by the detection of L. pneumophila and some of its amoeba hosts in floating biofilms from anthropogenic and natural aquatic environments (73, 74). Legionella spp. can survive and replicate within amoebal vacuoles, and they are also able to survive within the cyst form of amoebae (75-79). Cysts with Legionella can be transmitted by aerosols over great distances and Legionella within amoebae has been proposed as the dominant mode of transmission to humans rather than free Legionella (72, 80). Other bacteria that can interact with amoebae include Listeria monocytogenes, Mycobacterium avium, and many gastrointestinal pathogens such as Vibrio cholera, Helicobacter pylori, Salmonella spp., Shigella sonnei and Campylobacter spp. (81-90). In many cases where interactions between amoebae and pathogenic bacteria have been demonstrated, the amoeba is believed to be the natural host (91). Most of the described interactions between bacteria and amoebae issue intracellular survival of the bacteria, but there are an increasing number of studies demonstrating that bacteria are able to replicate within the amoeba (80, 82, 89, 92, 93). By evolving mechanisms to survive within different species of amoebae (or other unicellular eukaryotes), bacteria may not only escape the threat of being preyed upon, but could also benefit from protection from unfavorable conditions outside the host. Acanthamoeba The most common FLA are Acanthamoeba spp. which can be found in a very wide variety of environments including seawater, fresh water lakes, public tap- and bottled water, ventilation systems, soil, sand, dust, air, chlorinated swimming pools, lens cleaning solutions and hospital equipment (71, 94). More than 24 different Acanthamoeba spp. have been identified, of which the majority have been associated with human infections. They have been isolated from different parts of the body, including human skin lesions, cornea and lung tissue. Acanthamoeba castellanii and A. polyphaga are the most commonly isolated species causing human infections. They are known to cause two different diseases: granulomatous amoebic encephalitis, most often diagnosed in immunocompromised hosts, and amoebic keratitis. Keratitis is a rare infection of the eye, and it primarily affects otherwise healthy contact lens wearers, with the risk of degradation of vision, and in rare cases even blindness (94, 95). 16 Acanthamoebae have two stages in its life cycle; a vegetative trophozoite stage and a resistant cyst stage. The trophozoites are in the size range of 1235 μm (Figure 2), while cysts are smaller (5-20 μm), spherical and equipped with a double cell wall. The transformation from trophozoites to cysts is a response to harsh conditions (96). The double-walled cyst is highly resistant to chlorination, antimicrobials and disinfectants, as well as to changes in pH and osmolarity (97). Moreover, it has been shown that cysts can survive transient heating even up to 95˚C (98) and temperatures below 10˚C (99). When the environmental conditions are favorable, an amoeba in the cyst form can reverse to its trophozoite stage (100). Acanthamoeba spp. can colonize biofilms where they normally prey on different microorganisms such as algae, yeast or bacteria. Acanthapodia, extended parts of their cell membrane, are used for prey capturing but also for surface attachment and cellular movements (94). Captured prey are taken up by endocytosis and directed to food vacuoles where the amoeba releases digestive enzymes to degrade the food (96). Several studies have described that some bacteria are able to avoid degradation within different amoebae and establish a symbiotic relationship with the organism, especially with protozoa of the genus Acanthamoeba (80). There are several shared functions and similarities between the amoebal cells and human phagocytic cells that make Acanthamoeba species attractive as a model organism in protistology and in interaction studies with pathogenic bacteria (71, 101). In this thesis, species of the genus Acanthamoebae were used for amoeba-bacteria interaction studies. Fig.2 A. polyphaga trophozoite. 17 Intracellular trafficking pathways in Acanthamoeba Many bacterial species are taken up via phagocytosis and are used as food source by Acanthamoeba (71, 102). Bacteria, yeasts and latex beads have been used to study the phagocytic process (102, 103).The studies show that yeast and Escherichia coli K12 adhere to a mannose binding protein (MBP) on the Acanthamoeba surface (104, 105). The adherence to the Acanthamoeba surface and the following invagination of the Acanthamoeba plasma membrane is an actin dependent process (104). Once inside the Acanthamoeba, phagosomes containing yeast or E.coli K12 are acidified and eventually degraded. Latex beads cannot be degraded but are instead exocytosed upon presentation of new beads on the Acanthamoeba surface (96). Some bacterial species can escape the intracellular killing mechanisms, and survive and even multiply inside the Acanthamoeba (Figure 3). Bacterial multiplication can lead to lysis of the Acanthamoeba, and released bacteria can infect new Acanthamoeba or spread to infect other hosts. For a few bacteria, the strategies to evade lysosomal killing in Acanthamoeba have been studied (106). The human pathogens L. pneumophila, and Mycobacterium avium survive within amoebae by their ability to inhibit fusion of the phagosome with lysosomes, whereas Burkholderia cepacia survives within acidic vacuoles that are distinct from the lysosomal compartment (82, 107, 108). The intra protozoan fate of the bacteria might even differ within a bacterial species, as for Escherichia coli. The virulent serotypes E. coli O157 (109), E. coli K1 O187 (110, 111) and E. coli K5 (111) are able to invade and multiply within Acanthamoeba spp. whereas the avirulent laboratory strain K12, HB101, is phagocytosed and killed (110, 111). Both L. pneumophila and M. avium are known to use a similar survival strategy in Acanthamoeba castellanii as in human macrophages (82, 107, 112, 113). Different bacteria show different mechanisms for: (i) bacterial entrance of the Acanthamoeba (ii) their intracellular survival and (iii) the escape form the Acanthamoeba (101). These events can also be affected by bacterial viability. For L. pneumophila, viability was shown to affect the level of phagosome-lysosome fusion. The level of fusion increased for formalin-treated an UV- killed L. pneumophila compared to viable L. pneumophila, suggesting an active process from the bacterial point of view. On the other hand, viability had no effect on the uptake of L. pneumophila into Acanthamoeba as both viable and non-viable bacteria were taken up by coiling phagocytosis (107), suggesting that the uptake process is more dependent on amoebal activity. Mapping out the entrance mechanism and the following intracellular faith of a pathogenic bacterium open up for the possibility of blocking the bacteria-amoebae interaction, and thereby removing the amoebae as a safe haven for the pathogen. Studying intracellular trafficking pathways of bacteria in amoebae can also lead to valuable insights into the infection process in human cells. 18 Fig.3 Intracellular trafficking pathways for bacteria in Acanthamoeba, with permission from (106). Unicellular eukaryotes as reservoir for Campylobacter In 1988, a study by King et al showed that C. jejuni within the amoeba A. castellanii and the ciliate Tetrahymena pyriformis obtained protection from free chlorine residuals. The increased resistance was presumed to be due to survival within protozoan cells (114). In 2005, our research group demonstrated that C. jejuni was able to survive and multiply within A. polyphaga (88). This observation was confirmed with the related species A. castellanii as well as in the ciliate T. pyriformis (87). Later on, Snelling et al reported that C. jejuni internalized within A. castellanii were able to colonize broilers (115). In our group, we have studied a number of different Campylobacter spp. (C. jejuni, C. lari, C. coli and C. hyointestinalis) in coculture with different unicellular eukaryotes (Naegleria americana, Hartmanella vermiformis, A. polyphaga, A. rhysodes, A. castellanii and T. pyriformis). Prolonged survival was generally observed in cocultures, compared to Campylobacter incubated in media alone (90). The knowledge about C. jejuni survival and replication within A. polyphaga led to the Acanthamoeba-Campylobacter coculture method (ACC) (89). 19 The ACC – method The ACC (Acanthamoeba-Campylobacter coculture) method (Figure 4) was presented in 2007 as a novel method to isolate and enrich low concentrations of Campylobacter spp. (89). Four strains relevant to human infection and colonization of livestock, C. jejuni, C. coli, C. lari, and C. hyointestinalis, were effectively enriched by the method. The method is based on the intracellular survival and replication of Campylobacter spp. in the amoeba A. polyphaga. Basically, samples suspected to contain Campylobacter are seeded in peptone-yeast-glucose (PYG) medium containing A. polyphaga trophozoites and incubated aerobically at 37°C for 24 h. During incubation, the Campylobacter cells enter and proliferate at a high rate inside the amoebae. Transferring a small volume of the cocultured sample to a blood agar plate, combined with additional microaerobic incubation for 24 h, results in a large number of Campylobacter colonies for detection and further characterization. The method has shown to perform excellently with inoculum containing as little as 1-10 bacteria, and even in a background of competing flora. Isolation of C. jejuni from environmental samples can sometimes be difficult with conventional culture methods. Such samples normally contain a high microbiological flora and growth of other bacteria could outcompete low densities of C. jejuni cells. Filtering of generous amounts of water and a following enrichment step is usually required to detect C. jejuni. This is labor intensive and can generate false negative results. Detection with polymerase chain reaction (PCR) is usually more sensitive but has the draw-back that bacterial isolates are not available for further studies and the method does not discriminate between viable and dead C. jejuni. With its sensitivity and selectivity, the ACC method is a promising tool to detect C. jejuni and other thermotolerant Campylobacter cells at concentrations far below the detection limit of blood agar culture. The sensitivity and selectivity of the ACC method was equal to, or even better than Preston broth and Campylobacter Enrichment Broth (89). 20 Fig.4 Description of the ACC–method, with the permission from (89). 21 Aims ¾ To investigate if Acanthamoeba spp. can protect C. jejuni in acidic environments and to study the effect of low pH on C. jejuni association with A. polyphaga. ¾ To investigate if A. polyphaga can protect C. jejuni in milk and orange juice at different temperatures. ¾ To investigate the mechanisms behind the uptake and intracellular survival of C. jejuni in A. polyphaga ¾ To investigate if C. jejuni maintain genetic stability during enrichment in A. polyphaga 22 Methods Bacterial and amoebic cultures (Paper I-IV) Three Acanthamoeba species, A. polyphaga (strain Linc Ap-1, kindly provided by Bernard La Scola, Université de la Méditerranée, Marseille, France), A. castellanii, and A. rhysodes (Both originally isolated from Swedish patients with keratitis and kindly provided by J. Winiecka-Krusnell, Swedish Institute for Infectious Disease Control (SMI), Sweden), were used in Paper I. In Paper II-IV the A. polyphaga strain Linc Ap-1was used. C. jejuni strain CCUG 11284 was used in Paper I-II, and strain 81-176 was used in Paper III. C. jejuni strain CCUG 11284 and 40 MLST-characterized wild bird- or food animal associated C. jejuni strains from previously published studies, chosen to represent C. jejuni from diverse sources (116-118), were used in Paper IV. Acanthamoeba spp. were maintained in peptoneyeast-glucose (PYG) medium at 27°C. For the experiments, Acanthamoeba spp. in PYG medium were seeded into 12-well (Paper I, II, IV) or 24-well culture plates (Paper III) and incubated at 27°C until the trophozoites formed confluent layers at the bottom of the wells. Culture plates with confluent Acanthamoeba layers were inoculated with C. jejuni into each well. Bacterial strains were stored at -80°C. Before each experiment, bacteria were grown on conventional blood agar plates at 42°C for 20-24 h in a micro aerobic environment using GasPak incubation containers (Beckton Dickinson (BD), Franklin Lakes, NJ) and CampyGen paper sachets (Oxoid, Basingstoke, United Kingdom). Bacterial cells were harvested and diluted in PYG medium to create stock solutions suitable for each individual experiment. Coincubation of C. jejuni with Acanthamoeba spp. in acidic environments or in beverages (Paper I, II) In Paper I acidification and adjustment of media (water and PBS) to different pH levels (pH 1, 2, 4 and 5) was done using hydrochloric acid (HCl). To test whether the presence of amoebae in the acidified medium influenced the survival of C. jejuni at different pH, the following three treatments were used: C. jejuni preincubated with A. polyphaga before acidification of the 23 medium (treatment A), C. jejuni added to A. polyphaga after acidification of the medium (treatment B), and C. jejuni in acidified medium without A. polyphaga (treatment C). Plates were incubated in an aerobic environment at 32˚C and sampled at 0, 5, and 20 h. The pH level of the fluid in each well was continuously measured during the experiment. In Paper II the C. jejuni survival was studied in two beverage products, milk (pH 6.4) and orange juice (pH 3.9), in the presence or absence of A. polyphaga. To test whether the presence of amoeba in milk and juice influenced the survival of C. jejuni, the following three treatments were used: C. jejuni preincubated with A. polyphaga before the addition of product (treatment A), C. jejuni added to A. polyphaga after the addition of product (treatment B), and C. jejuni in product without A. polyphaga (treatment C). Plates were incubated in an aerobic environment at room temperature and 4˚C and sampled at 0, 3, 6, 18, 24 and 48 h. All samples were 10-fold serially diluted in PYG medium and spread on blood agar for colony counting. Controls without amoeba, only C. jejuni, were treated in the same way as cocultures. Pasteurization of C. jejuni cells co-incubated with A. polyphaga in milk and orange juice (Paper II) To test whether the presence of amoeba in milk and juice, influenced the survival of C. jejuni when heated to recommended Swedish pasteurization temperatures, the following three treatments were used: C. jejuni preincubated with A. polyphaga before the addition of product (treatment A), C. jejuni added to A. polyphaga after the addition of product (treatment B), and C. jejuni in product without A. polyphaga (treatment C). Directly after the addition of product, samples of 100 μl were taken from the different treatments, A, B, and C and added to tubes containing 500 μl of milk or juice. The tubes were gently shaken at 1400 rpm (MS2 Minishaker IKA®, Germany) and then incubated in a water bath (Heto DT Hetotherm, Denmark). Incubation conditions for milk tubes were 72-74˚C for 15 sec and 85˚C for 15 sec for juice tubes. After heating, the sample tubes were put on ice and thereafter 100-μl samples were directly spread on blood agar for colony counting. Quantification of adhered/internalized C. jejuni into A. polyphaga (Paper I) The effect of pH (pH 2, 3, 4, 5 and 7) on adhesion/internalization of C. jejuni into A. polyphaga was measured by flow cytometry (FACSCalibur, Becton Dickinson, CA). The flow cytometer was set up to detect samples containing A. polyphaga cells with or without stained C. jejuni. Staining of the bacteria was performed using a 50 mM cyanoditolyl tetrazolium chloride (CTC) (Polyscience, Eppelheim, Germany) solution according to the manu- 24 facturer’s instructions. Data acquisition and analysis were done using the CellQuest v. 3.3 software. The same samples were also analyzed by fluorescence microscopy (Axioskop microscope; Zeiss, West Germany). For each sample, 100 amoebae were analyzed, and the number of stained C. jejuni cells adhered to or internalized in each amoeba was counted. In the flow cytometry analysis it was not possible to distinguish C. jejuni that adhered to the A. polyphaga surface, from those residing intracellularly, and therefore the term “adhered/internalized” was used throughout the article. Determination of C. jejuni motility (Paper I) The effect of pH on C. jejuni motility was assessed by a swarming assay. C. jejuni pre-exposed to different pH levels was inoculated onto soft agar (0.4% thioglycolate) plates. 5 μl was inoculated in the center of the plate and the zone diameter was measured after incubation at 37˚C for 48 h. Uptake and intracellular survival of C. jejuni in A. polyphaga (Paper III) Coculturing of viable and heat killed C. jejuni with A. polyphaga To determine the importance of bacterial viability for internalization and intracellular trafficking in the amoebal species A. polyphaga we compared these processes between viable and heat killed cells of the C. jejuni strain 81-176. Heat inactivation of bacteria was done in a water bath (Heto DT Hetotherm, Denmark) for 45 min at 70ͼC. To separate viable and heat killed cells, bacteria were stained using two different approaches depending on the experiment. For the quantification experiment bacterial suspensions, both viable and heat killed, were stained using the Live/Dead stain (Bacterial Viability Kit, Molecular probes) according to the manufacturer’s instructions. Bacteria with intact membranes fluoresced green (live), whereas heat killed bacteria with damaged membranes fluoresced red (dead). For intracellular localization experiments, bacterial suspensions were stained using CTC solution. CTC was used instead of the Live/Dead stain because it has red fluorescence which gives a better contrast to the green Alexafluor-488dextran (Molecular Probes, Eugene, Oregon) containing lysosomes, described below. For the experiments, A. polyphaga in PYG medium were seeded into 24-well culture plates. Plates with confluent Acanthamoeba layers were inoculated with viable and/or heat killed C. jejuni into each well. These plates were cocultured in an aerobic environment at room temperature and 10 μl samples were taken at the time intervals 1, 24, 48, 72 and 96 h for quantification of adhered/internalized bacteria and at 1, 24, 48 and 72 h for 25 determination of intracellular localization of C. jejuni in A. polyphaga trophozooites. Coculture samples were analyzed by fluorescence microscopy (Axioskop Zeiss, West Germany). For each sample, 50-100 amoebae were analyzed, and the number of stained viable or heat killed C. jejuni cells adhered to or internalized in each amoeba was counted. Determination of intracellular localization of viable and heat killed C. jejuni in A. polyphaga The intracellular localization of viable or heat killed C. jejuni was determined using Alexafluor-488 labeled dextran (Molecular Probes). The dextran is internalized and processed via the endocytic pathway ending up in degradative phagolysosomes both in human cells and the widely used amoebozoa species Dictyostelium discoideum (119, 120). Coculture samples were analyzed by fluorescence microscopy (Axioskop Zeiss, West Germany) and the number of stained viable and heat killed cells outside and inside dextran filled vacuoles were counted in each amoeba. For each sample, 50-100 amoebae were analyzed. Investigation of the genetic stability of C. jejuni after enrichment in A. polyphaga (Paper IV) C. jejuni enrichment by A. polyphaga cocultures Forty MLST-characterized wild bird- or food animal associated C. jejuni strains were chosen to represent C. jejuni from diverse sources. The reference strain CCUG 11284 was also included. Each strain had a unique sequence type (ST), and the collection of strains was chosen to reflect a high genetic diversity. Each strain was cocultured with A. polyphaga as described above. Cocultures were incubated aerobically for 24 h at 37ͼC. To mimic the conditions of environmental samples with low numbers of C. jejuni, an inoculum giving a final concentration of approximately 1 to 10 CFU/ml was chosen, as determined by colony counting on blood agar plates. Microscopic observations assessed bacterial amplification by lysis of amoebae and entrance of C. jejuni into the surrounding medium. Samples with low C. jejuni concentrations after coculture were incubated an additional 24 h. MLST and flaA typing of C. jejuni isolates Following passage and subsequent amplification in amoebae, the STs and CCs were determined for each of the 41 C. jejuni strains. The DNA was 26 extracted using the boil lysate method. Fresh overnight cultures were harvested and one loop-full of cells was resolved in ddH2O and boiled for 8 minutes. The lysates were directly used as template for PCR reactions. PCR amplification and nucleotide sequencing were performed using standard C. jejuni MLST protocols (65). To increase the discriminatory power, the virulence gene, flaA, with its short variable region (SVR-fla), was sequenced and used as a complementary target. The 321-bp sequence containing the SVRfla was determined for each isolate prior to and after amoeba passage (65, 121, 122). Acquired forward and reverse sequences were aligned to create contiguous fragments. The primers used for the seven MLST loci and the flaA gene are available at (http://pubmlst.org/campylobacter/). After amplification, the PCR products were purified according to the following protocol: The supernatant was discarded by inverting the PCR-plate on a paper towel and spun for 1min at 500 g. The pellet was then washed with100 μl ice cold 70% EtOH followed by centrifugation at 2800 g for 10min and removal of supernatant. This washing step was repeated and the dried PCR-product was resuspended in10μl ddH2O. Purified PCR products were sent to Macrogen Inc. (South Korea) for sequencing using internally separated nested primer pairs. The retrieved sequences from Macrogen Inc. were then used for genotyping of C. jejuni through BLAST in the PubMLST database (http://pubmlst.org/campylobacter/). 27 Results and discussion Coincubation of C. jejuni with Acanthamoeba spp. in acidic environments (Paper I) Campylobacter jejuni are sensitive to acidity, disinfectants, oxygen exposure, osmotic stress, desiccation and heating (42). On the other hand, coincubation with unicellular eukaryotes have been shown to increase C. jejuni survival during exposure to free chlorine (114), disinfectants (87) and aerobic conditions (88). In vitro studies have shown that C. jejuni poorly survive low pH levels (123), but in this thesis we have found that, in the presence of Acanthamoebae spp. the bacterium can survive pH levels far below its normal range (Paper I, II). When C. jejuni cells were co-incubated with A. polyphaga in acidified PBS or tap water, the bacteria survived at pH 4 for 20 h (Figure 5) (Paper I). We also found that moderately acidic conditions (pH 4 and 5) triggered C. jejuni motility and increased the adhesion/internalization of bacteria into A. polyphaga. This was shown by significantly increased swarming when C. jejuni were pretreated with acidified PBS at pH 5 for 1 h (1.9 ± 0.03 cm; P < 0.001), compared to that of the control (1.5 ± 0.03 cm). The results from flow cytometric analysis showed an increase in the geometric mean fluorescence intensity (MFI) of A. polyphaga cells infected with CTC-stained bacteria at pH 4 and 5 compared to at pH 7. Also the microscopy data showed that the association of C. jejuni with A. polyphaga was increased at pH 4 and 5, compared to that at pH 7. At pH 5, 6.65 ± 0.94 bacteria were found inside or on the surface of each A. polyphaga trophozoite (P < 0.0001 [Tukey’s test]), and at pH 4, the number was 5.87 ± 0.96 bacteria/amoeba (P < 0.0001 [Tukey’s test]). The percentage of amoebae associated with bacteria was also dependent on the surrounding pH, and significant differences in association compared to that at pH 7 were seen at pH 4 and 5. At pH 7, 37.0% ± 18% of A. polyphaga trophozoites had C. jejuni cells internalized or on the surface, compared to 78.0% ± 12% at pH 5 (P = 0.016 [Tukey’s test]) and 80.0% ± 8.7% at pH 4 (P = 0.012 [Tukey’s test]). 28 Fig.5 Survival of C. jejuni in PBS at pH 4. The box plots show survival as the % of initial CFU after 5 h and 20 h. C. jejuni were treated in three different ways: treatment A (white boxes), C. jejuni added prior to acidification; treatment B (striped boxes), C. jejuni added after acidification; and treatment C (dotted boxes), C. jejuni without A. polyphaga, as a control. These findings show that amoebae of the genus Acanthamoeba can protect C. jejuni in acidic environments where they normally cannot survive alone. The findings can have implications for the understanding of C. jejuni survival during passage through the gastrointestinal tract of host animals and humans and for hygiene routines in chicken stables. C. jejuni have been shown difficult to eradicate from chicken stables even after sanitation (124). Acid treatment is a commonly used sanitation method in chicken stables and the results from this study suggest that Acanthamoeba spp. might protect C. jejuni from such treatment. 29 Evaluation of A. polyphaga as a Trojan horse for protection of C. jejuni in beverages (Paper II) Several outbreaks where unpasteurized milk has been the source of C. jejuni infections have been documented. Many of those outbreaks have arisen from farmers that serve their visitors raw milk (125-130). There is an increasing trend of drinking “natural” unpasteurized milk, which can contribute to an increased number of bacterial infections, such as Campylobacteriosis (131). Most C. jejuni infections are sporadic, and due to the high number of human infections with sometimes neuronal complications and in the worst case; fatal outcomes (4), identification of all possible infection sources are important. In recent decades a number of human pathogens have caused outbreaks through acidic fruit juices (132). The low pH in juice is an inhospitable environment for C. jejuni and therefore juice has not been of interest when looking for potential infection sources. In this thesis I studied the survival of C. jejuni in milk and orange juice in the presence of A. polyphaga (Paper II). In milk stored at room temperature, coculture with A. polyphaga were shown to increase the C. jejuni survival at all time points studied. When stored at refridgerator temperature (4ͼC), coculture increased the C. jejuni survival at all time points except at 48 h, where C. jejuni alone exceeded the survival in cocultures. This is consistent with previous studies reporting good survival of Campylobacter in refrigerated milk (123, 133). In orange juice (pH 3.9), the survival of C. jejuni was increased when cocultured with A. polyphaga, both when incubated at room temperature (Figure 6) and at 4ͼC, compared to cultures with C. jejuni alone. Statistically significant differences in relative C. jejuni survival between cocultures, treatment A and B, and C. jejuni without A.polyphaga, treatment C, were seen at 18 h to 48 h when the treatments were incubated at room temperature (Treatment A and C: Kruskal-Wallis test; (18 h) p < 0.0001, (24 h) p < 0.0001) (Treatment B and C: Kruskal-Wallis test; (18 h) p = 0.0428, (24 h) p = 0.0428, (48h) p = 0.0029) (Figure 6). When treatments were incubated at 4ͼC, significant differences in relative survival between treatment A, bacteria pre-incubated with amoebae before addition of juice, and treatment C, C. jejuni without A.polyphaga, were seen at 18 h to 48 h (KruskalWallis test; (18 h) p = 0.0001, (24 h) p = 0.0002, (48 h) p = 0.0489. Amoebae could come in contact with C. jejuni contaminated milk and orange juice in different ways. Amoebae are common in water supply systems (134, 135) and drinking water is used both within the industrial process (136) and in the preparation of juice in households when adding water to juice concentrate. Amoebae can attach to different surfaces that can be found in juice squeezing machines or other utensils e.g. baby bottles (73, 137, 138). The pasteurized beverage can also be post infected with Campylobacter. In a 30 British study it was established that Jackdaws and Magpies that attacked the foil caps of milk bottles caused people to become infected with C. jejuni when they consumed the remaining milk later on (139, 140). In summary my findings show that the presence of A. polyphaga can increase the C. jejuni survival in milk and orange juice. However, since A. polyphaga did not show any protective effect on C. jejuni when heated to recommended pasteurization temperatures, neither in milk nor in orange juice (Paper II), the pasteurized milk and orange juice would then make a safer alternative for the consumer compared to unpasteurized. Fig.6 Survival of C. jejuni in orange juice at pH 3.9 at room temperature. The graph shows survival as the % of initial CFU after 0, 3, 6, 18, 24 and 48 h. C. jejuni were treated in three different ways: treatment A (dots), C. jejuni preincubated with A. polyphaga before the addition of product; treatment B (squares), C. jejuni inoculated to A. polyphaga after the addition of product; and treatment C (triangles), C. jejuni in product without A. polyphaga as a control. Uptake and intracellular survival of C. jejuni in A. polyphaga (Paper III) Many bacteria act as food source for amoebae. They are taken up via phagocytosis and are degraded in lysosomes. However, a number of bacterial species including several human pathogens can avoid lysosomal degradation in protozoa and for some species, the evasion strategy has been studied 31 in more detail (82, 107, 108). In this thesis we studied the uptake and intracellular trafficking of C. jejuni in the amoeba A. polyphaga. (Paper III). We found that viable C. jejuni entered A. polyphaga in higher numbers and at a higher rate compared to non-viable C. jejuni. The internalization of viable bacteria into A. polyphaga was significantly higher compared to heat killed bacteria after 1, 24 and 48 h of coincubation (repeated measures ANOVA with Duncans post test; (1 h) p < 0.001, (24 h) p < 0.001, (48 h) p < 0.001). After 1 h the average number of amoeba associated viable C. jejuni were 9.4 bacteria per amoeba, compared to 0.5 bacteria/amoeba for the heat killed control (Paper III). In addition, viable C. jejuni associated with a greater number of trophozites than non-viable. After one hour of coincubation, 96% of the A. polyphaga trophozoites were associated with viable C. jejuni compared to 0% of the trophozoites associated with heat killed C. jejuni (Liddells exact test; (1 h) p < 0.001). When the proportion of viable/heat killed C. jejuni was analyzed among trophozoites of this fraction, the amount of viable bacteria were dominating at all time points except at 96 h where the number of heat killed C. jejuni exceeded the number of viable (Liddells exact test; (1 h) p < 0.001, (24 h) p < 0.001, (48 h) p < 0.001, (96 h) p = 0.027) (Paper III). Once inside A. polyphaga, most viable C. jejuni were gathered in non-acidic vacuoles, and the majority of the heat killed bacteria were gathered in big acidic vacuoles, with the characteristics of amoebal lysosomes (Figure 7). Absolute numbers of bacteria observed inside non-digestive vacuoles were significantly higher for viable bacteria than for heat killed bacteria at all time points (repeated measures ANOVA with Duncans post test, (1 h) p < 0.001, (24 h) p < 0.001, (48 h) p < 0.001, (72 h) p < 0.001) (Paper III). Studies in human intestinal epithelial cells have also shown that the majority of C. jejuni cells are found in non-acidic vacuoles (141). Together these findings indicate that viability is important for bacterial entrance and intracellular survival in A. polyphaga. Further investigations are needed to identify proteins or bacterial functions that are critical for amoebal entrance. It could be that a functional flagella or the secretion of proteins is needed for binding to the amoebal surface and for subsequent internalization. A C. jejuni mutant (K2-32) with a kanamycin-resistance cassette inserted into the flagellin gene, flaA was shown to have greatly reduced motility and was not capable to adhere to or invade epithelial cells (142). It has also been shown that C. jejuni invasion of epithelial cells is dependent on the secretion of virulence proteins, termed the Campylobacter invasion antigens (Cia), from the bacterium’s flagellar Type III Secretion System (143-146). Epithelial cell invasion by C. jejuni and intracellular survival has been proposed to be the consequences of the Cia proteins’ modification of host cell regulatory pathways (147). In the case of L. pneumophila, the usage of different bacterial mechanisms to attach to and invade different 32 protozoan hosts, show how complex and specific these interactions can be (148). Once inside A. polyphaga, we showed that the viability of C. jejuni was also important for the escape of lysosomal degradation. The intracellular survival of C. jejuni in A. polyphaga can be due to effector proteins that prevent the fusion of phagosomes with lysosomes and in L. pneumophila this process is regulated by the dot/icm genes (149). Genes involved in C. jejuni invasion and intracellular survival in eukaryotic cells remains poorly studied. However, despite many features shared by amoebae and macrophages, it appears that C. jejuni, in contrast to L. pneumophila, are unable to avoid lysosomal degradation in human macrophages (141, 150). Fig.7 Left: Viable C. jejuni (red) are gathered in non acidic vacuoles and not in dextran filled acidic vacuoles (green). Right: Heat killed C. jejuni (yellow) are gathered in dextran filled acidic vacuoles (green). Genetic stability of C. jejuni after enrichment in A. polyphaga (Paper IV) In our research group we have developed a method for enrichment of environmental samples with low C. jejuni concentrations based on coculture with A. polyphaga, the ACC-method, described above (89). However, for the ACC method to be used in outbreak investigations and to enable identification of the source of infection, it is important to evaluate if the propagation of C. jejuni in the amoeba has any effect on the genetic stability of the bacterial strains. Recombination is frequent in C. jejuni (151), indicating that the bacterium has a high genetic instability and a high risk for genetic changes. C. jejuni has also been shown to be naturally competent for transformation (152). 33 In this thesis, the genetic stability of C. jejuni during coculture with A. polyphaga was assessed by multilocus sequence typing (MLST) and flaA short variable region (SVR) sequencing of 40 environmental C. jejuni isolates before and after coculture. We found that all isolates could be successfully enriched by the ACC method when added at a bacterial concentration of approximately 1 to 10 CFU/ml. With MLST and SVR-fla sequencing, it was assessed that all 40 C. jejuni isolates retained their genetic profile after passage in A. polyphaga (Paper IV). This suggests that the ACC-method is suitable for amplification and detection of C. jejuni in environmental samples and that this propagation does not induce genetic changes as determined by MLST or SVR-fla sequencing. The MLST method with SVR-fla sequencing is a robust and widely used method for typing of C. jejuni isolates. However, as the method only provides information about genetic changes in the seven house-keeping genes and the SVR-fla, we cannot rule out that genetic changes occur in other regions of the C. jejuni genome during passage in A. polyphaga. Furthermore, if bacteria compete to enter the amoebae and/or if replication is selective when more than one type of C. jejuni is present, needs further study. 34 Conclusions I conclude that: I Amoebae of the genus Acanthamoeba can prolong C. jejuni survival at low pH. (Paper I) II Low pH triggers C. jejuni motility as well as adhesion and internalization of the bacterium into A. polyphaga. (Paper I) III A. polyphaga can increase the survival of C. jejuni in milk and orange juice, but cannot protect the bacteria from temperatures recommended for pasteurization. (Paper II) IV Bacterial viability is important for C. jejuni adhesion, uptake and intracellular survival within A. polyphaga. (Paper III) V Viable C. jejuni escape degradation in A. polyphaga by avoiding localization to digestive vacuoles. (Paper III) VI The ACC method is suitable for enrichment of C. jejuni strains of different origin and propagation of C. jejuni by this method does not affect the genetic profile, as determined by MLST and SVR-fla sequencing. (Paper IV) 35 Future perspectives In this thesis I found that Acanthamoeba spp. are good candidates for being an environmental C. jejuni reservoir. Further interaction studies between C. jejuni and unicellular eukaryotes are needed to better establish the role of such organisms in the epidemiology of C. jejuni. How C. jejuni invade and survive intracellularly within Acanthamoebae is a complicated puzzle to lay. Further molecular studies are needed to identify amoebal-or bacterial surface receptors as well as bacterial proteins or pathways involved in avoiding lysosomal destruction. Such knowledge could be useful for strategies to block the invasion and the intracellular survival. To investigate if Acanthamoeba spp. can act as reservoirs for C. jejuni in poultry stables, I have built a water supply system in miniature to mimic the systems currently used in many chicken farms in Sweden. This system will be experimentally colonized with Acanthamoeba spp. and inoculated with C. jejuni to study if the bacteria can be protected by the amoebae under seminatural conditions. In this system we will also study the effect of the currently used treatment with mild acids (e g citric acid) as sanitizer. 36 Svensk sammanfattning (Swedish summary) Campylobacter är en vanlig orsak till bakteriell diarrésjukdom världen över. Den orsakar lidande och död i utvecklingsländer och stora samhällskostnader i form av bl.a. sjukvård i i-länder. Bland de cirka 18 arter av Campylobacter som identifierats är Campylobacter jejuni den som orsakar flest infektioner hos människa. Cirka 7000 fall rapporteras årligen i Sverige och ca 200 000 inom EU. Inkubationstiden är två till fyra dagar, och de flesta insjuknar i feber, kräkningar, huvudvärk och diarréer, som kan vara blodtillblandade. En campylobakterinfektion kräver sällan antibiotikabehandling, men en del patienter kan behöva vätskeersättning. Normalt sett är en campylobakterinfektion över på en vecka men i sällsynta fall kan den leda till allvarliga neurologiska komplikationer som t.ex. Guillain–Barrés syndrom (GBS), och till och med till dödsfall. De flesta campylobakterfall är sporadiska och är associerade med dåligt tillagad kyckling, opastöriserad mjölk och kontaminerat vatten. C. jejuni finns i mag-tarm kanalen hos flera varmblodiga djur, inklusive gris, nötkreatur, får, sällskapsdjur och fåglar, där de kan existera utan att ge några symptom. Bakterien har visat sig vara känslig mot olika miljöfaktorer såsom syre, förändringar i pH och temperatur. Hur C. jejuni överlever och sprids i naturen är ännu inte helt klarlagt, men sannolikt existerar olika reservoarer utöver varmblodiga djur. I tidigare studier har C. jejuni visats kunna överleva och tillväxa inuti amöbor av släktet Acanthamoeba. Encelliga eukaryoter såsom amöbor, ciliater och alger är vanligt förekommande i jord och olika vattenmiljöer, där de till stor del livnär sig på bakterier. Flera bakterierarter har visat sig kunna överleva inuti encelliga eukaryoter. Det mest kända exemplet är Legionella pneumophila som använder sig av amöbor för sin överlevnad och spridning. Genom att undersöka vilka encelliga eukaryoter som kan fungera som värdorganism för bakterier kan vi få en ökad förståelse för hur sjukdomsalstrande (patogena) bakterier överlever i naturen. Amöbor är tåliga organismer och bakterier som befinner sig inuti dem kan dra fördel av den barriär de utgör mot den kringliggande omgivningen. Med denna avhandling har jag visat att amöbor av släktet A. polyphaga kan skydda C. jejuni i miljöer med lågt pH. Sur miljö visade sig öka C. jejuni´s förmåga att binda till och invadera amöbor. Det är sannolikt att bakterien på liknande sätt kan skydda sig mot lågt pH i epitelet i den sura magsäcken. Det är även möjligt att amöbor i vissa fall kan skydda bakterierna 37 under denna passage då amöbor innehållande C. jejuni intagits. Fenomenet kan sannolikt även förklara varför syrabehandling av vattenledningssystem i kycklingstallar ofta är en ineffektiv saneringsmetod mot Campylobacter. Jag fann också att samodling med A. polyphaga generellt ökade C. jejuni’s förmåga att överleva i både mjölk och apelsinjuice, gentemot när bakterierna inkuberades utan amöbor. Detta tyder på att, i de fall då amöbor finns närvarande i dessa produkter, skulle de kunna öka kontaminerande Campylobacter’s livslängd. Vidare visar avhandlingen på att levande C. jejuni internaliseras i A. polyphaga i mycket högre grad än avdödade bakterier. Detta har betydelse för hur vi ser på interaktionen mellan dessa bakterier och amöban, där C. jejuni i motsats till en del andra bakterier, sannolikt aktivt invaderar A. polyphaga. Jag fann även att C. jejuni överlever inuti amöban genom att undvika vakuoler för nedbrytning. Analyserna visade också att det vid samodling av C. jejuni med amöbor, inte sker några förändringar i gener som används för fylogenetisk analys av bakterien. Multilocus sequence typing (MLST) är en metod för fylogenetisk analys av bakterier. Här studeras den genetiska variationen i ett antal gener. Förändringar i genomet är vanliga hos C. jejuni. Då samodling med A. polyphaga används som en anrikningsmetod för C. jejuni är det dock av stor vikt att känna till huruvida det vid sådan anrikning kan ske genetiska förändringar i dessa gener som studeras med MLST-metoden. Sådana förändringar skedde dock inte. Sammantaget visar studierna på att amöbor av släktet A. polyphaga kan fungera som en reservoar för C. jejuni i naturen och kan i olika sammanhang sannolikt bidra till ökad överlevnad och spridning av denna bakterie. Studierna visar också att samodling med A. polyphaga är en robust anrikningsmetod för C. jejuni. 38 Acknowledgements Ett stort tack till Patrik Ellström, som under min doktorandtid har gått från att ha varit biträdande handledare till huvudhandledare. Du har hela tiden stått mig nära och alltid hjälpt mig när det behövts. En omtänksammare människa får man leta efter! Jag kommer att minnas trevliga jobbmöten i Uppsala och din gästfrihet. Tack för att jag fick vara din doktorand. Ett stort tack till min f.d. huvudhandledare Björn Olsen (nu biträdande), som gav mig chansen och förtroendet till att doktorera. Du gav mig fria tyglar från dag ett, vilket varit mycket nyttigt och lärorikt (men för ett kontrollfreak som jag var det en smula påfrestande i början-). Trots avståndet till Uppsala har din inspiration och energi alltid smittat av sig. Din entusiasm och framåtanda är något som jag verkligen uppskattar! Ett stort tack min biträdande handledare Diana Axelsson-Olsson, som jag alltid kunnat vända mig till för att diskutera stort som smått. Ditt engagemang har varit ovärderligt! Tack för att du tidiga morgnar och sena vinterkvällar hjälpte mig i labbet då mjölk och juicemanuset krävde fyra händer. Din tro på mig och din idérikedom har varit viktig under min doktorandtid. Tack till hela ”Zoonosgruppen” på Linnéuniversitetet i Kalmar! En samling människor med olika bakgrunder och kunskaper. Kul att få ha varit med i denna gemenskap. Människor har kommit och gått i denna grupp, men jag vill rikta ett särskilt tack till: Jonas W. för att du alltid erbjudit din hjälp (även om jag ibland varit dålig på att ta emot den…) och för att du fått mig att känna mig trygg i Kalmargruppen. Conny, för att livet blir mycket roligare med en Cönny Tölf vid sin sida. Din humor är oslagbar och ditt engagemang vad gäller att lösa andras problem har jag sällan skådat. Jag ser framemot uppföljaren till ”Albert och Herbert”, ”Det bästa med Conny & Jenny”. Johan, för att du är en riktig god kamrat som man kan dela livets vedermödor med. Jag saknar snacket vi hade under pizzaluncherna. Maria ”Marita” Blomqvist, för att jobbet blev extra roligt att gå till när du var i Kalmar och för att jag nu har en vän i Uppsala att besöka. Sara L, för alla pratstunder och sällskap till mikrobiologen. Neus, för din positiva inställning och för att du är duktig på att anordna trevliga tillställningar utanför jobbet. Petra, för alla diskussioner och för gott resesällskap till Canada, Schweiz och Lund. Abbtesaim, för din omtänksamhet. Jorge, för fina expeditionsbilder från Arktis. Daniel, för din positiva anda och för att du alltid bryr dig. 39 Michelle, Alexis and Joanne, for the joy and help you have been giving me during my PhD. Especially thanks to Alexis for your statistically advices. Lovisa, f.d. zoonosmedlem och medlem i Calmare vinsällskap, som alltid generöst delat med sig av sin kunskap. Lycka till med livet i Värmland! Tack till alla övriga kollegor på Linnéunivesitet som förgyllt fikastunder och luncher. Ingen nämnd ingen glömd! Dock riktar jag ett extra tack till vaktmästarna Anders Månsson och Henrik Hallberg som fixar det mesta (och dessutom med ett leende) och Sara Gunnarsson som vänligt hjälpt mig med kemikalier/labutrustning. Ett stort tack till Birgitta Sembrant och Anna Foyer vid Institutionen för Medicinska vetenskaper, Uppsala Universitet. Ni har tålmodigt och snabbt svarat på alla mina administrativa frågor. Tack till min mentor Ivar Tjernström. Jag har uppskattat våra måndagsträffar på sjukhuset i Kalmar där jag kravlöst kunnat prata om min tillvaro som doktorand. Tack för att du tog dig tid, för att du lyssnat och delgett mig dina doktoranderfarenheter. Tack till Berit på mikrobiologen i Kalmar som alltid så vänligt försett mig med blodplattor. Tack till Ingrid Hansson på SVA som hjälpte mig med planering och materiel till vattenledningsförsöket. Även om inte studien roddes i hamn så hoppas jag att systemet en dag ska komma till användning. Tack till Ulf Danielsson på Ebbetorps Gård i Smedby, för inblicken i hur ett kycklingstall fungerar och bedrivs. Tack till Mamma Rut och Pappa Tommy som alltid stöttat mig och låtit mig gå mina egna vägar. Jag har väl inte alltid valt de enklaste, men ni har hållit god min ändå. Tack Farmor Allis och Farfar Henry för att dörren till Almgatan 6 alltid står öppen och för alla släktträffar som ordnas där. Jag kan inte tacka dig nog farmor för alla telefonsamtal, all mat och många härliga insjöbad. Tack Storebror Linus och Linda för att ni hjälper syster yster när hon hamnar på hal is….. er kan man alltid räkna med. Tack till den stora familjen i Skäveryd som alltid ställer upp. Hos er känner jag mig alltid välkommen och platsen har varit ett andningshål för en orolig doktorandsjäl. Tack mina små godingar Thelma och Beata för att ni ger mig trygghet och glädje varje dag. Nu ska mamma inte tillbringa mer tid i det provisoriska kontoret i källaren. Tack Min Mikael, för att du är den du är! En kämpe helt enkelt. Med din inställning och ditt ödmjuka motto: ”Sluta lipa, kavla upp ärmarna och gör något åt saken”, kommer man långt har jag lärt mig. Tack för att du finns, livet blir allt bra mycket enklare med dig vid min sida! 40 References 1. 2. 3. 4. 5. 6. 7. 8. 9. 10. 11. 12. 13. 14. 15. Curtis D, Hill A, Wilcock A, Charlebois S. Foodborne and waterborne pathogenic bacteria in selected Organisation for economic cooperation and Development (OECD) countries. Journal of food science. 2014 Oct;79(10):1871-6. Hoffmann S, Batz MB, Morris JG, Jr. Annual cost of illness and qualityadjusted life year losses in the United States due to 14 foodborne pathogens. Journal of food protection. 2012 Jul;75(7):1292-302. Platts-Mills JA, Kosek M. Update on the burden of Campylobacter in developing countries. Current opinion in infectious diseases. 2014 Oct;27(5):444-50. Bolton DJ. Campylobacter virulence and survival factors. Food Microbiology. 2015 6//;48(0):99-108. Campylobacter (3rd Edition). Washington, DC, USA: ASM Press; 2008. pp.1215, 111. Fernández H, Vera F, Villanueva MP, García A. Occurrence of campylobacter species in healthy well-nourished and malnourished children. Brazilian Journal of Microbiology. 2008;39:56-8. On SL. Taxonomy of Campylobacter, Arcobacter, Helicobacter and related bacteria: current status, future prospects and immediate concerns. Symposium series (Society for Applied Microbiology). 2001(30):1S-15S. Janssen R, Krogfelt KA, Cawthraw SA, van Pelt W, Wagenaar JA, Owen RJ. Host-pathogen interactions in Campylobacter infections: the host perspective. Clinical microbiology reviews. 2008 Jul;21(3):505-18. McCARTHY ND, GILLESPIE IA, LAWSON AJ, RICHARDSON J, NEAL KR, HAWTIN PR, et al. Molecular epidemiology of human Campylobacter jejuni shows association between seasonal and international patterns of disease. Epidemiology & Infection. 2012;140(12):2247-55. ECDC. Annual epidemiological report 2014-food-and waterborne diseases and zoonoses European Centre for Disease Prevention and Control, Stockholm. 2014:20-4. CDC. Morbidity and Mortality Weekly Report. 2013 April 19;62(15):283-7. Black RE, Levine MM, Clements ML, Hughes TP, Blaser MJ. Experimental Campylobacter jejuni infection in humans. The Journal of infectious diseases. 1988 Mar;157(3):472-9. Robinson DA. Infective dose of Campylobacter jejuni in milk. British medical journal (Clinical research ed). 1981 May 16;282(6276):1584. Guerry P. Campylobacter flagella: not just for motility. Trends in microbiology. 2007 Oct;15(10):456-61. Jin S, Joe A, Lynett J, Hani EK, Sherman P, Chan VL. JlpA, a novel surfaceexposed lipoprotein specific to Campylobacter jejuni, mediates adherence to host epithelial cells. Molecular microbiology. 2001 Mar;39(5):1225-36. 41 16. Russell RG, O'Donnoghue M, Blake DC, Jr., Zulty J, DeTolla LJ. Early colonic damage and invasion of Campylobacter jejuni in experimentally challenged infant Macaca mulatta. The Journal of infectious diseases. 1993 Jul;168(1):2105. 17. Johnson WM, Lior H. A new heat-labile cytolethal distending toxin (CLDT) produced by Campylobacter spp. Microbial pathogenesis. 1988 Feb;4(2):11526. 18. Silva J, Leite D, Fernandes M, Mena C, Gibbs PA, Teixeira P. Campylobacter spp. as a foodborne pathogen: a review. Frontiers in Microbiology. 2011 2011September-27;2. 19. Peterson MC. Clinical aspects of Campylobacter jejuni infections in adults. The Western journal of medicine. 1994;161(2):148-52. 20. Ang CW, De Klerk MA, Endtz HP, Jacobs BC, Laman JD, van der Meche FG, et al. Guillain-Barre syndrome- and Miller Fisher syndrome-associated Campylobacter jejuni lipopolysaccharides induce anti-GM1 and anti-GQ1b Antibodies in rabbits. Infection and immunity. 2001 Apr;69(4):2462-9. 21. Nyati KK, Nyati R. Role of Campylobacter jejuni infection in the pathogenesis of Guillain-Barre syndrome: an update. BioMed research international. 2013;2013:852195. 22. Mori M, Kuwabara S. Fisher syndrome. Current treatment options in neurology. 2011 Feb;13(1):71-8. 23. Moore JE, Barton MD, Blair IS, Corcoran D, Dooley JS, Fanning S, et al. The epidemiology of antibiotic resistance in Campylobacter. Microbes and infection / Institut Pasteur. 2006 Jun;8(7):1955-66. 24. Xia Q, Muraoka WT, Shen Z, Sahin O, Wang H, Wu Z, et al. Adaptive mechanisms of Campylobacter jejuni to erythromycin treatment. BMC microbiology. 2013;13:133. 25. Wilson DJ, Gabriel E, Leatherbarrow AJ, Cheesbrough J, Gee S, Bolton E, et al. Tracing the source of campylobacteriosis. PLoS genetics. 2008;4(9):e1000203. 26. Hanninen ML, Haajanen H, Pummi T, Wermundsen K, Katila ML, Sarkkinen H, et al. Detection and typing of Campylobacter jejuni and Campylobacter coli and analysis of indicator organisms in three waterborne outbreaks in Finland. Applied and environmental microbiology. 2003 Mar;69(3):1391-6. 27. Nygard K, Andersson Y, Rottingen JA, Svensson A, Lindback J, Kistemann T, et al. Association between environmental risk factors and campylobacter infections in Sweden. Epidemiology and infection. 2004 Apr;132(2):317-25. 28. Schonberg-Norio D, Takkinen J, Hanninen ML, Katila ML, Kaukoranta SS, Mattila L, et al. Swimming and Campylobacter infections. Emerging infectious diseases. 2004 Aug;10(8):1474-7. 29. Horrocks SM, Anderson RC, Nisbet DJ, Ricke SC. Incidence and ecology of Campylobacter jejuni and coli in animals. Anaerobe. 2009;15(1–2):18-25. 30. Waldenstrom J, Broman T, Carlsson I, Hasselquist D, Achterberg RP, Wagenaar JA, et al. Prevalence of Campylobacter jejuni, Campylobacter lari, and Campylobacter coli in different ecological guilds and taxa of migrating birds. Applied and environmental microbiology. 2002 Dec;68(12):5911-7. 31. Griekspoor P, Colles FM, McCarthy ND, Hansbro PM, Ashhurst-Smith C, Olsen B, et al. Marked host specificity and lack of phylogeographic population structure of Campylobacter jejuni in wild birds. Molecular ecology. 2013 Mar;22(5):1463-72. 42 32. Weis AM, Miller WA, Byrne BA, Chouicha N, Boyce WM, Townsend AK. Prevalence and pathogenic potential of campylobacter isolates from free-living, human-commensal american crows. Applied and environmental microbiology. 2014 Mar;80(5):1639-44. 33. Craven SE, Stern NJ, Line E, Bailey JS, Cox NA, Fedorka-Cray P. Determination of the incidence of Salmonella spp., Campylobacter jejuni, and Clostridium perfringens in wild birds near broiler chicken houses by sampling intestinal droppings. Avian diseases. 2000 Jul-Sep;44(3):715-20. 34. Studer E, Lüthy J, Hübner P. Study of the presence of Campylobacter jejuni and C. coli in sand samples from four Swiss chicken farms. Research in Microbiology. 1999 4//;150(3):213-9. 35. Snelling WJ, Moore JE, Dooley JSG. The colonization of broilers with Campylobacter. Worlds Poult Sci J. 2005 Dec;61(4):655-62. 36. Boysen L, Rosenquist H, Larsson JT, Nielsen EM, Sorensen G, Nordentoft S, et al. Source attribution of human campylobacteriosis in Denmark. Epidemiology and infection. 2014 Aug;142(8):1599-608. 37. Torjesen I. Supermarkets with the most Campylobacter infected chickens will be named and shamed next week. BMJ (Clinical research ed). 2014;349:7008. 38. Ogden ID, Dallas JF, MacRae M, Rotariu O, Reay KW, Leitch M, et al. Campylobacter excreted into the environment by animal sources: prevalence, concentration shed, and host association. Foodborne pathogens and disease. 2009 Dec;6(10):1161-70. 39. Pattison M. Practical intervention strategies for Campylobacter. Symposium series (Society for Applied Microbiology). 2001(30):121S-5S. 40. Jansen W, Reich F, Klein G. Large-scale feasibility of organic acids as a permanent preharvest intervention in drinking water of broilers and their effect on foodborne Campylobacter spp. before processing. Journal of applied microbiology. 2014 Jun;116(6):1676-87. 41. Hansson I, Vagsholm I, Svensson L, Olsson Engvall E. Correlations between Campylobacter spp. prevalence in the environment and broiler flocks. Journal of applied microbiology. 2007 Sep;103(3):640-9. 42. Park SF. The physiology of Campylobacter species and its relevance to their role as foodborne pathogens. Int J Food Microbiol. 2002 Apr;74(3):177-88. 43. Wang WL, Powers BW, Leuchtefeld NW, Blaser MJ. Effects of disinfectants on Campylobacter jejuni. Applied and environmental microbiology. 1983 Apr;45(4):1202-5. 44. Simonian HP, Vo L, Doma S, Fisher RS, Parkman HP. Regional postprandial differences in pH within the stomach and gastroesophageal junction. Digestive diseases and sciences. 2005 Dec;50(12):2276-85. 45. Kong F, Singh RP. Disintegration of solid foods in human stomach. Journal of food science. 2008 Jun;73(5):67-80. 46. Cawthraw SA, Wassenaar TM, Ayling R, Newell DG. Increased colonization potential of Campylobacter jejuni strain 81116 after passage through chickens and its implication on the rate of transmission within flocks. Epidemiol Infect. 1996 Aug;117(1):213-5. 47. Stern NJ, Bailey JS, Blankenship LC, Cox NA, McHan F. Colonization characteristics of Campylobacter jejuni in chick ceca. Avian diseases. 1988 Apr-Jun;32(2):330-4. 48. Humphrey TJ. Campylobacter jejuni: Some Aspects of Epidemiology, Detection and Control. British Food Journal. 1992;94(1):21-5. 43 49. Jackson DN, Davis B, Tirado SM, Duggal M, van Frankenhuyzen JK, Deaville D, et al. Survival mechanisms and culturability of Campylobacter jejuni under stress conditions. Antonie Van Leeuwenhoek International Journal of General and Molecular Microbiology. 2009 Nov;96(4):377-94. 50. Bronowski C, James CE, Winstanley C. Role of environmental survival in transmission of Campylobacter jejuni. FEMS microbiology letters. 2014 Jul;356(1):8-19. 51. Backert S, Hofreuter D. Molecular methods to investigate adhesion, transmigration, invasion and intracellular survival of the foodborne pathogen Campylobacter jejuni. Journal of Microbiological Methods. 2013 10//;95(1):823. 52. On SL. Isolation, identification and subtyping of Campylobacter: where to from here? J Microbiol Methods. 2013 Oct;95(1):3-7. 53. Randall L, Lemma F, Rodgers J, Vidal A, Clifton-Hadley F. Development and evaluation of internal amplification controls for use in a real-time duplex PCR assay for detection of Campylobacter coli and Campylobacter jejuni. Journal of medical microbiology. 2010 Feb;59(Pt 2):172-8. 54. Arnold ME, Jones EM, Lawes JR, Vidal AB, Clifton-Hadley FA, Rodgers JD, et al. Bayesian analysis of culture and PCR methods for detection of Campylobacter spp. in broiler caecal samples. Epidemiology and infection. 2015 Jan;143(2):298-307. 55. Xiao D, Tao XX, Wang P, Liu GD, Gong YN, Zhang HF, et al. Rapid and high-throughput identification of recombinant bacteria with mass spectrometry assay. Biomedical and environmental sciences : BES. 2014 Apr;27(4):250-8. 56. Bessède E, Solecki O, Sifré E, Labadi L, Mégraud F. Identification of Campylobacter species and related organisms by matrix assisted laser desorption ionization–time of flight (MALDI-TOF) mass spectrometry. Clinical Microbiology and Infection. 2011 11//;17(11):1735-9. 57. Garcia S, Heredia NL. Campylobacter. Guide to Foodborne Pathogens: John Wiley & Sons; 2013. p. 188-96. 58. Pitkanen T. Review of Campylobacter spp. in drinking and environmental waters. J Microbiol Methods. 2013 Oct;95(1):39-47. 59. Miller WG, Yee E, Jolley KA, Chapman MH. Use of an improved atpA amplification and sequencing method to identify members of the Campylobacteraceae and Helicobacteraceae. Letters in applied microbiology. 2014 Jun;58(6):582-90. 60. Cornelius AJ, Gilpin B, Carter P, Nicol C, On SL. Comparison of PCR binary typing (P-BIT), a new approach to epidemiological subtyping of Campylobacter jejuni, with serotyping, pulsed-field gel electrophoresis, and multilocus sequence typing methods. Applied and environmental microbiology. 2010 Mar;76(5):1533-44. 61. On SL, Jordan PJ. Evaluation of 11 PCR assays for species-level identification of Campylobacter jejuni and Campylobacter coli. Journal of clinical microbiology. 2003 Jan;41(1):330-6. 62. Nakari U-M, Hakkinen M, Siitonen A. Identification of Persistent Subtypes of Campylobacter jejuni by Pulsed-Field Gel Electrophoresis in Finland. Foodborne pathogens and disease. 2011;8(10):1143-5. 63. Pike BL, Guerry P, Poly F. Global Distribution of Penner Serotypes: A Systematic Review. PloS one. 2013;8(6):e67375. 44 64. Guerry P, Poly F, Riddle M, Maue AC, Chen YH, Monteiro MA. Campylobacter polysaccharide capsules: virulence and vaccines. Frontiers in cellular and infection microbiology. 2012;2:7. 65. Dingle KE, Colles FM, Wareing DR, Ure R, Fox AJ, Bolton FE, et al. Multilocus sequence typing system for Campylobacter jejuni. Journal of clinical microbiology. 2001 Jan;39(1):14-23. 66. Dingle KE, Colles FM, Ure R, Wagenaar JA, Duim B, Bolton FJ, et al. Molecular characterization of Campylobacter jejuni clones: a basis for epidemiologic investigation. Emerging infectious diseases. 2002 Sep;8(9):94955. 67. Sails AD, Swaminathan B, Fields PI. Utility of multilocus sequence typing as an epidemiological tool for investigation of outbreaks of gastroenteritis caused by Campylobacter jejuni. Journal of clinical microbiology. 2003 Oct;41(10):4733-9. 68. Adl SM, Simpson AG, Farmer MA, Andersen RA, Anderson OR, Barta JR, et al. The new higher level classification of eukaryotes with emphasis on the taxonomy of protists. The Journal of eukaryotic microbiology. 2005 SepOct;52(5):399-451. 69. Thomas V, McDonnell G, Denyer SP, Maillard J-Y. Free-living amoebae and their intracellular pathogenic microorganisms: risks for water quality. FEMS Microbiology Reviews. 2010;34(3):231-59. 70. Rodríguez-Zaragoza S. Ecology of Free-Living Amoebae. Critical Reviews in Microbiology. 1994;20(3):225-41. 71. Siddiqui R, Khan NA. Biology and pathogenesis of Acanthamoeba. Parasites Vectors. 2012a Jan;5. 72. Rowbotham TJ. Preliminary report on the pathogenicity of Legionella pneumophila for freshwater and soil amoebae. Journal of clinical pathology. 1980 Dec;33(12):1179-83. 73. Thomas V, Bouchez T, Nicolas V, Robert S, Loret JF, Levi Y. Amoebae in domestic water systems: resistance to disinfection treatments and implication in Legionella persistence. Journal of applied microbiology. 2004;97(5):950-63. 74. Declerck P, Behets J, van Hoef V, Ollevier F. Replication of Legionella pneumophila in floating biofilms. Current microbiology. 2007 Nov;55(5):43540. 75. Anand CM, Skinner AR, Malic A, Kurtz JB. Interaction of L. pneumophilia and a free living amoeba (Acanthamoeba palestinensis). The Journal of hygiene. 1983 Oct;91(2):167-78. 76. Kilvington S, Price J. Survival of Legionella pneumophila within cysts of Acanthamoeba polyphaga following chlorine exposure. The Journal of applied bacteriology. 1990 May;68(5):519-25. 77. Kuchta JM, Navratil JS, Shepherd ME, Wadowsky RM, Dowling JN, States SJ, et al. Impact of Chlorine and Heat on the Survival of Hartmannella vermiformis and Subsequent Growth of Legionella pneumophila. Applied and environmental microbiology. 1993 Dec;59(12):4096-100. 78. Bigot R, Bertaux J, Frere J, Berjeaud JM. Intra-amoeba multiplication induces chemotaxis and biofilm colonization and formation for Legionella. PloS one. 2013;8(10):e77875. 79. Li Z, Solomon JM, Isberg RR. Dictyostelium discoideum strains lacking the RtoA protein are defective for maturation of the Legionella pneumophila replication vacuole. Cellular microbiology. 2005 Mar;7(3):431-42. 45 80. Greub G, Raoult D. Microorganisms resistant to free-living amoebae. Clinical Microbiology Reviews. 2004 Apr;17(2):413-433. 81. Ly TM, Muller HE. Ingested Listeria monocytogenes survive and multiply in protozoa. Journal of medical microbiology. 1990 Sep;33(1):51-4. 82. Cirillo JD, Falkow S, Tompkins LS, Bermudez LE. Interaction of Mycobacterium avium with environmental amoebae enhances virulence. Infection and immunity. 1997 Sep;65(9):3759-67. 83. Thom S, Warhurst D, Drasar BS. Association of Vibrio cholerae with fresh water amoebae. Journal of medical microbiology. 1992 May;36(5):303-6. 84. Winiecka-Krusnell J, Wreiber K, von Euler A, Engstrand L, Linder E. Freeliving amoebae promote growth and survival of Helicobacter pylori. Scandinavian journal of infectious diseases. 2002;34(4):253-6. 85. Gaze WH, Burroughs N, Gallagher MP, Wellington EM. Interactions between Salmonella typhimurium and Acanthamoeba polyphaga, and observation of a new mode of intracellular growth within contractile vacuoles. Microbial ecology. 2003 Oct;46(3):358-69. 86. Jeong HJ, Jang ES, Han BI, Lee KH, Ock MS, Kong HH, et al. Acanthamoeba: could it be an environmental host of Shigella? Experimental parasitology. 2007 Feb;115(2):181-6. 87. Snelling WJ, McKenna JP, Lecky DM, Dooley JS. Survival of Campylobacter jejuni in waterborne protozoa. Applied and environmental microbiology. 2005 Sep;71(9):5560-71. 88. Axelsson-Olsson D, Waldenstrom J, Broman T, Olsen B, Holmberg M. Protozoan Acanthamoeba polyphaga as a potential reservoir for Campylobacter jejuni. Applied and environmental microbiology. 2005 Feb;71(2):987-92. 89. Axelsson-Olsson D, Ellstrom P, Waldenstrom J, Haemig PD, Brudin L, Olsen B. Acanthamoeba-Campylobacter coculture as a novel method for enrichment of Campylobacter species. Applied and environmental microbiology. 2007 Nov;73(21):6864-9. 90. Axelsson-Olsson D, Olofsson J, Svensson L, Griekspoor P, Waldenström J, Ellström P, et al. Amoebae and algae can prolong the survival of Campylobacter species in co-culture. Experimental Parasitology. 2010;126(1):59-64. 91. Winiecka-Krusnell J, Linder E. Bacterial infections of free-living amoebae. Res Microbiol. 2001 Sep;152(7):613-9. 92. Molmeret M, Horn M, Wagner M, Santic M, Abu Kwaik Y. Amoebae as training grounds for intracellular bacterial pathogens. Applied and environmental microbiology. 2005 Jan;71(1):20-8. 93. Abd H, Johansson T, Golovliov I, Sandstrom G, Forsman M. Survival and growth of Francisella tularensis in Acanthamoeba castellanii. Applied and environmental microbiology. 2003 Jan;69(1):600-6. 94. Trabelsi H, Dendana F, Sellami A, Sellami H, Cheikhrouhou F, Neji S, et al. Pathogenic free-living amoebae: Epidemiology and clinical review. Pathologie Biologie. 2012 Dec;60(6):399-405. 95. Marciano-Cabral F, Cabral G. Acanthamoeba spp. as agents of disease in humans. Clinical Microbiology Reviews. 2003 Apr;16(2):273-307. 96. Khan NA. Acanthamoeba: biology and increasing importance in human health. FEMS Microbiology Reviews. 2006;30(4):564-95. 97. Siddiqui R, Dudley R, Khan NA. Acanthamoeba differentiation: a two-faced drama of Dr Jekyll and Mr Hyde. Parasitology. 2012;139(7):826-34. 46 98. Chang C-W, Lu L-W, Kuo C-L, Hung N-T. Density of environmental Acanthamoeba and their responses to superheating disinfection. Parasitology research (1987). 01/2013;112(11):3687-96. 99. Axelsson-Olsson D, Olofsson J, Ellstrom P, Waldenstrom J, Olsen B. A simple method for long-term storage of Acanthamoeba species. Parasitol Res. 2009 Mar;104(4):935-7. 100. Khunkitti W, Lloyd D, Furr JR, Russell AD. Acanthamoeba castellanii: Growth, encystment, excystment and biocide susceptibility. J Infect. 1998 Jan;36(1):43-8. 101. Siddiqui R, Khan NA. Acanthamoeba is an evolutionary ancestor of macrophages: A myth or reality? Experimental Parasitology. 2012b;130(2):957. 102. Khan NA. Pathogenesis of Acanthamoeba infections. Microbial pathogenesis. 2003 Jun;34(6):277-85. 103. Avery SV, Harwood JL, Lloyd D. Low temperature-induced adaptations in lipid metabolism and physiological function in Acanthamoeba castellanii cultures of different ages. Biochemical Society transactions. 1994 Aug;22(3):257S. 104. Alsam S, Sissons J, Dudley R, Khan NA. Mechanisms associated with Acanthamoeba castellanii (T4) phagocytosis. Parasitology research. 2005 Aug;96(6):402-9. 105. Kim JH, Matin A, Shin HJ, Park H, Yoo KT, Yuan XZ, et al. Functional roles of mannose-binding protein in the adhesion, cytotoxicity and phagocytosis of Acanthamoeba castellanii. Experimental parasitology. 2012 Oct;132(2):287-92. 106. Siddiqui R, Khan NA. War of the microbial worlds: who is the beneficiary in Acanthamoeba-bacterial interactions? Experimental parasitology. 2012 Apr;130(4):311-3. 107. Bozue JA, Johnson W. Interaction of Legionella pneumophila with Acanthamoeba castellanii: uptake by coiling phagocytosis and inhibition of phagosome-lysosome fusion. Infection and immunity. 1996 Feb;64(2):668-73. 108. Lamothe J, Thyssen S, Valvano MA. Burkholderia cepacia complex isolates survive intracellularly without replication within acidic vacuoles of Acanthamoeba polyphaga. Cellular microbiology. 2004 Dec;6(12):1127-38. 109. Barker J, Humphrey TJ, Brown MW. Survival of Escherichia coli O157 in a soil protozoan: implications for disease. FEMS microbiology letters. 1999 Apr 15;173(2):291-5. 110. Alsam S, Jeong SR, Sissons J, Dudley R, Kim KS, Khan NA. Escherichia coli interactions with Acanthamoeba: a symbiosis with environmental and clinical implications. Journal of medical microbiology. 2006 Jun;55(Pt 6):689-94. 111. Matin A, Jung SY. Interaction of Escherichia coli K1 and K5 with Acanthamoeba castellanii trophozoites and cysts. The Korean journal of parasitology. 2011 Dec;49(4):349-56. 112. Horwitz MA. The Legionnaires' disease bacterium (Legionella pneumophila) inhibits phagosome-lysosome fusion in human monocytes. The Journal of experimental medicine. 1983 Dec 1;158(6):2108-26. 113. Frehel C, de Chastellier C, Lang T, Rastogi N. Evidence for inhibition of fusion of lysosomal and prelysosomal compartments with phagosomes in macrophages infected with pathogenic Mycobacterium avium. Infection and immunity. 1986 Apr;52(1):252-62. 114. King CH, Shotts EB, Wooley RE, Porter KG. SURVIVAL OF COLIFORMS AND BACTERIAL PATHOGENS WITHIN PROTOZOA DURING 47 CHLORINATION. Applied and Environmental Microbiology. 1988 Dec;54(12):3023-33. 115. Snelling WJ, Stern NJ, Lowery CJ, Moore JE, Gibbons E, Baker C, et al. Colonization of broilers by Campylobacter jejuni internalized within Acanthamoeba castellanii. Arch Microbiol. 2008 Feb;189(2):175-9. 116. Griekspoor P, Engvall EO, Olsen B, Waldenstrom J. Multilocus sequence typing of Campylobacter jejuni from broilers. Veterinary microbiology. 2010 Jan 6;140(1-2):180-5. 117. Waldenstrom J, On SL, Ottvall R, Hasselquist D, Olsen B. Species diversity of campylobacteria in a wild bird community in Sweden. Journal of applied microbiology. 2007 Feb;102(2):424-32. 118. Broman T, Waldenstrom J, Dahlgren D, Carlsson I, Eliasson I, Olsen B. Diversities and similarities in PFGE profiles of Campylobacter jejuni isolated from migrating birds and humans. Journal of applied microbiology. 2004;96(4):834-43. 119. Pryor PR. Analyzing lysosomes in live cells. Methods in enzymology. 2012;505:145-57. 120. Rodriguez-Paris JM, Nolta KV, Steck TL. Characterization of lysosomes isolated from Dictyostelium discoideum by magnetic fractionation. The Journal of biological chemistry. 1993 Apr 25;268(12):9110-6. 121. Colles FM, Jones TA, McCarthy ND, Sheppard SK, Cody AJ, Dingle KE, et al. Campylobacter infection of broiler chickens in a free-range environment. Environmental microbiology. 2008 Aug;10(8):2042-50. 122. Meinersmann RJ, Helsel LO, Fields PI, Hiett KL. Discrimination of Campylobacter jejuni isolates by fla gene sequencing. Journal of clinical microbiology. 1997 Nov;35(11):2810-4. 123. Blaser MJ, Hardesty HL, Powers B, Wang WLL. SURVIVAL OF CAMPYLOBACTER-FETUS SUBSP JEJUNI IN BIOLOGICAL MILIEUS. J Clin Microbiol. 1980;11(4):309-13. 124. Bare J, Sabbe K, Van Wichelen J, van Gremberghe I, D'Hondt S, Houf K. Diversity and habitat specificity of free-living protozoa in commercial poultry houses. Applied and environmental microbiology. 2009 Mar;75(5):1417-26. 125. Korlath JA, Osterholm MT, Judy LA, Forfang JC, Robinson RA. A pointsource outbreak of campylobacteriosis associated with consumption of raw milk. The Journal of infectious diseases. 1985 Sep;152(3):592-6. 126. Evans MR, Roberts RJ, Ribeiro CD, Gardner D, Kembrey D. A milk-borne campylobacter outbreak following an educational farm visit. Epidemiology and infection. 1996 Dec;117(3):457-62. 127. Heuvelink AE, van Heerwaarden C, Zwartkruis-Nahuis A, Tilburg JJ, Bos MH, Heilmann FG, et al. Two outbreaks of campylobacteriosis associated with the consumption of raw cows' milk. International journal of food microbiology. 2009 Aug 31;134(1-2):70-4. 128. Longenberger AH, Palumbo AJ, Chu AK, Moll ME, Weltman A, Ostroff SM. Campylobacter jejuni infections associated with unpasteurized milk--multiple States, 2012. Clinical infectious diseases : an official publication of the Infectious Diseases Society of America. 2013 Jul;57(2):263-6. 129. Taylor EV, Herman KM, Ailes EC, Fitzgerald C, Yoder JS, Mahon BE, et al. Common source outbreaks of Campylobacter infection in the USA, 1997-2008. Epidemiology and infection. 2013 May;141(5):987-96. 48 130. Oliver SP, Boor KJ, Murphy SC, Murinda SE. Food safety hazards associated with consumption of raw milk. Foodborne pathogens and disease. 2009 Sep;6(7):793-806. 131. Mungai EA, Behravesh CB, Gould LH. Increased outbreaks associated with nonpasteurized milk, United States, 2007-2012. Emerging infectious diseases. 2015 Jan;21(1):119-22. 132. Burnett SL, Beuchat LR. Human pathogens associated with raw produce and unpasteurized juices, and difficulties in decontamination. Journal of industrial microbiology & biotechnology. 2000 Dec;25(6):281-7. 133. Doyle MP, Roman DJ. Prevalence and survival of Campylobacter jejuni in unpasteurized milk. Applied and environmental microbiology. 1982 Nov;44(5):1154-8. 134. Wang H, Edwards MA, Falkinham JO, 3rd, Pruden A. Probiotic approach to pathogen control in premise plumbing systems? A review. Environmental science & technology. 2013 Sep 17;47(18):10117-28. 135. Delafont V, Brouke A, Bouchon D, Moulin L, Hechard Y. Microbiome of freeliving amoebae isolated from drinking water. Water research. 2013 Dec 1;47(19):6958-65. 136. Averbeck M, Schieberle P. Characterisation of the key aroma compounds in a freshly reconstituted orange juice from concentrate. Eur Food Res Technol. 2009 2009/08/01;229(4):611-22. 137. Erdem AK, Sanli-Yürüdü NO, Arslan-Aydoğdu EÖ, Dogruoz N, Zeybek Z, Türetgen I, et al. Quantitative microbiological analysis of biofilm communities from the surfaces of different cooling tower materials. IUFS Journal of Biology. 2010;67(1):9-16. 138. Sospedra I, Rubert J, Soriano JM, Mañes J. Incidence of microorganisms from fresh orange juice processed by squeezing machines. Food Control. 2012 1//;23(1):282-5. 139. Hudson SJ, Sobo AO, Russel K, Lightfoot NF. Jackdaws as potential source of milk-borne Campylobacter jejuni infection. Lancet. 1990 May 12;335(8698):1160. 140. Southern JP, Smith RM, Palmer SR. Bird attack on milk bottles: possible mode of transmission of Campylobacter jejuni to man. Lancet. 1990 Dec 8;336(8728):1425-7. 141. Watson RO, Galan JE. Campylobacter jejuni survives within epithelial cells by avoiding delivery to lysosomes. PLoS pathogens. 2008 Jan;4(1):e14. 142. Yao R, Burr DH, Doig P, Trust TJ, Niu H, Guerry P. Isolation of motile and non-motile insertional mutants of Campylobacter jejuni: the role of motility in adherence and invasion of eukaryotic cells. Molecular microbiology. 1994 Dec;14(5):883-93. 143. Konkel ME, Kim BJ, Rivera-Amill V, Garvis SG. Identification of proteins required for the internalization of Campylobacter jejuni into cultured mammalian cells. Advances in experimental medicine and biology. 1999;473:215-24. 144. Konkel ME, Klena JD, Rivera-Amill V, Monteville MR, Biswas D, Raphael B, et al. Secretion of virulence proteins from Campylobacter jejuni is dependent on a functional flagellar export apparatus. Journal of bacteriology. 2004 Jun;186(11):3296-303. 145. Malik-Kale P, Parker CT, Konkel ME. Culture of Campylobacter jejuni with sodium deoxycholate induces virulence gene expression. Journal of bacteriology. 2008 Apr;190(7):2286-97. 49 146. Christensen JE, Pacheco SA, Konkel ME. Identification of a Campylobacter jejuni-secreted protein required for maximal invasion of host cells. Molecular microbiology. 2009 Aug;73(4):650-62. 147. Eucker TP, Konkel ME. The cooperative action of bacterial fibronectin-binding proteins and secreted proteins promote maximal Campylobacter jejuni invasion of host cells by stimulating membrane ruffling. Cellular microbiology. 2012 Feb;14(2):226-38. 148. Harb OS, Venkataraman C, Haack BJ, Gao LY, Kwaik YA. Heterogeneity in the attachment and uptake mechanisms of the Legionnaires' disease bacterium, Legionella pneumophila, by protozoan hosts. Applied and environmental microbiology. 1998 Jan;64(1):126-32. 149. Gao LY, Harb OS, Abu Kwaik Y. Utilization of similar mechanisms by Legionella pneumophila to parasitize two evolutionarily distant host cells, mammalian macrophages and protozoa. Infection and immunity. 1997 Nov;65(11):4738-46. 150. Bruckert WM, Abu Kwaik Y. Complete and ubiquitinated proteome of the legionella-containing vacuole within human macrophages. Journal of proteome research. 02/2015;14(1):236-48. 151. Fearnhead P, Smith NG, Barrigas M, Fox A, French N. Analysis of recombination in Campylobacter jejuni from MLST population data. Journal of molecular evolution. 2005 Sep;61(3):333-40. 152. Vegge CS, Brondsted L, Ligowska-Marzeta M, Ingmer H. Natural transformation of Campylobacter jejuni occurs beyond limits of growth. PloS one. 2012;7(9):e45467. 50 Acta Universitatis Upsaliensis Digital Comprehensive Summaries of Uppsala Dissertations from the Faculty of Medicine 1115 Editor: The Dean of the Faculty of Medicine A doctoral dissertation from the Faculty of Medicine, Uppsala University, is usually a summary of a number of papers. A few copies of the complete dissertation are kept at major Swedish research libraries, while the summary alone is distributed internationally through the series Digital Comprehensive Summaries of Uppsala Dissertations from the Faculty of Medicine. (Prior to January, 2005, the series was published under the title “Comprehensive Summaries of Uppsala Dissertations from the Faculty of Medicine”.) Distribution: publications.uu.se urn:nbn:se:uu:diva-255804 ACTA UNIVERSITATIS UPSALIENSIS UPPSALA 2015