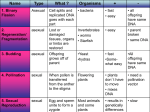
Survey
* Your assessment is very important for improving the workof artificial intelligence, which forms the content of this project
Some Implications of Paleoecology for Contemporary Ecology Author(s): Peter K. Schoonmaker and David R. Foster Source: Botanical Review, Vol. 57, No. 3 (Jul. - Sep., 1991), pp. 204-245 Published by: Springer on behalf of New York Botanical Garden Press Stable URL: http://www.jstor.org/stable/4354170 . Accessed: 19/08/2011 11:50 Your use of the JSTOR archive indicates your acceptance of the Terms & Conditions of Use, available at . http://www.jstor.org/page/info/about/policies/terms.jsp JSTOR is a not-for-profit service that helps scholars, researchers, and students discover, use, and build upon a wide range of content in a trusted digital archive. We use information technology and tools to increase productivity and facilitate new forms of scholarship. For more information about JSTOR, please contact [email protected]. New York Botanical Garden Press and Springer are collaborating with JSTOR to digitize, preserve and extend access to Botanical Review. http://www.jstor.org BOTANICAL THE VOL. 57 REVIEW JULY-SEPrEMBER, 1991 No. 3 Some Implications of Paleoecology for Contemporary Ecology AND DAVID R. FOSTER PETER K. SCHOONMAKER HarvardUniversity HarvardForest,Petersham,Massachusetts01366 I. Abstract...--------------------------------------------204 Sommaire.........................205 LI. Introduction .........................205 III QuaternaryEnvironmentsand Vegetation ............. . 206 .......... A. ClimaticChange Duringthe Quaternary.................................................. 206 ................................ B. Evidence for Biotic Change ................................................. 211 C. VegetationChangeDuringthe Late-Glacialand Holocene ....................................... 215 IV. PaleoecologicalInsightsfor CommunityEcology ................................................ 220 A. The Nature of the Biotic Community ................................................ 220 B. Evidenceof Disturbancein the PaleoecologicalRecord . ................................. 223 C. CommunityStabilityand Equilibrium............... .................................225 D. EvolutionaryConsiderations 227 ............................. E. Refugiain the PaleoecologicalRecord ...... 230 ........................ V. Prescriptionsand Applications............................................................................................................. 233 VI. Summary .-............. ......................................----------.....235 VII. Acknowledgments............... 236 ,,,.... . VIII. LiteratureCited........................................................................................... . ................. ..... 236 I. Abstract The paleoecologicalrecordallows contemporaryecologists to put currentphenomena into the context of a longer time-frame,therebyprovidingthe opportunityto evaluatethe importanceof slowly operatingprocesses,past cyclic or unusualevents, disturbanceregimes,and historicallyconstrainedphenomena.We brieflyoutline the environmentalhistoryof the Quatemary,discuss the spatialand temporalresolution of the paleoecologicalevidence for biotic change, and summarize data relevant to such currentissues as the nature of the biotic community, the role of disturbance, stabilityversus rapid change, evolutionarytheory, explanationsof species diversity, and refugiatheory. Finally, we offer examples of the utility of paleoecologicaltechniques for ecologists and environmentalscientists. Copies ofthis issue [57(3)] may be purchasedfrom the Scientific PublicationsDepartment,The New York BotanicalGarden, Bronx, NY 10458-5126 USA. Please inquireas to prices. TheBotanicalReview57: 204-245, Jul.-Sep., 1991 C 1991 The New York Botanical Garden 204 IMPLICATIONSOF PALEOECOLOGY 205 Sommaire Le dossier paleoecologiquepeut aider les ecologistes contemporainesa mettre les phenomenes actuels dans le contexte d'une periode plus longue, ainsi fournissant l'occasion d'evaluer l'importance des processus qui s'effectuentlentement, d'evenements cycliques ou inhabituelsdu passe de regimes perturbatrices,et du phenomenes historiquementcontraints.Nous traconsbrievementl'histoire du milieu environnement du Quaternaire, discutons la resolution spaciale le temporelle de l'evidence paleoecologiquedu changementbiotique et resumons les donnees applicable a des issues actuels tel que la nature de la communautebiotique, le role du derangement,la stabilite contre le changement rapide, la theorie d'evolution, les explicationsde la diversite des especes, et la theorie d'evolution, les explicationsde la diversite des especes, et la theorie de refugia. Finalement, nous presentons des exemplesde l'utilitedes techniquespaleoecologiquepour les ecologisteset les savants qui etudient l'environnement. II. Introduction The earth's biota responds to environmental changes that range in time from millisecondsto millions of years and affectareas from 10-6 to 10'4 m2(Fig. 1). Most ecologists study organisms on a time scale of less than 100 years (and are usually limited to the time span of a field season, grant or dissertation)but many patterns and processesin naturecan be better understoodover a longerperiod. By providing information on vegetation and environmental change, especially during the past 15,000 years, paleoecologycan broaden the interpretationof issues that are specifically relevant to modern ecologists, e.g., the prevalence of stability versus rapid change in ecosystems, the concept of an integratedcommunity, the importanceof disturbancein regulatingcommunityorganization,alternativeexplanationsof species diversity, and refugia theory. Paleoecologicalapproachesalso have many uses for applied ecologists and environmentalscientists concernedwith landscapemanagement, restorationecology, and the design of naturepreserves. The ecologicalimplicationsof paleoecologicaldata have been noted both by community ecologists and paleoecologists (cf. various authors in Barbour & Billings, 1988; Braun, 1950; Clements, 1924; Davis, 1983; Delcourtet al. 1983; Foster et al., 1990; Ogden, 1985; Walker, 1982; Watts, 1970; West, 1964). Becausepaleoecology is essentially community ecology stretchedbackwardthroughthe fourth dimension of time, it offers the ecologist a vast temporal vista from which to view present patterns,processesand phenomena.While the numberof paleoecologicalstudiesand the variety of temporaland spatial scales thereinhas increased,contemporarycommunity ecologistshave become more awareof the importanceof processesoccurring over extended periods of time. In the past two decades enough paleoecologicaldata have accumulatedto allow analysis and mapping of multiple data sets, which has resultedin temporal/spatialmaps relevant to many ecologists. As ecologistsarebeginningto extend the scalesat which contemporaryphenomena are addressed(Hubbell & Foster, 1986; Ricklefs, 1987; Urban et al., 1987), and as paleoecologistsare contributingto ongoingdiscussionsof ecologicaltheory (Bennett, 1988; Davis, 1986; Delcourt & Delcourt, 1987; Prentice, 1988; Huntley & Webb, 1989), the interrelationshipsof the two fields are becoming more apparent. This paper emphasizes this interdependencewith examples drawn mainly from REVIEW THEBOTANICAL 206 0/S WRBANCEREIG/AES E-NVIRONMLNTAL 1O 1* 104 1012 100 uio ric l04 RESPONSES lOS Joe 00o PATTERNS VEGoETArIONAL O 1S o04 LOALrERRESTRIAL ~~~~~~~~Plate fA N EGA - SCA LE MACRO-SCALE -S~ 13 MICRO 0 Rgme- 4 SCALEt s CIiai Evolution of the Sliota Tectonics t lollEOIitFmRMA@t lnftorgl@oclI c limant ic4 M ZONME OIA soeclatlot EstlnliasTION lcutos--TP o STAND Hu on: a A Secondary Succesesyn: Gap-phoest Replaoement tmpetltion cllie..tlvlfiot if o.ptlOen Outbrsah ------* l *Oitu'rboncs Events' 00 o0 10)0 Examples: Wildtire Wind Danmage Clear Cut Flood Earthguok. YP P REE Prdulii 11 10 14 ia SPATIAL SCALE 112 (in 2 l00 0 104 o lo,; SHSRUB ~~~~~~~~~~~~~H.IHERBS Fig. 1. Spatial and temporal scales at which disturbance,biotic responsesand vegetation pattems can be detected. Paleoecologicaldata can detect micro- and macro-scaledisturbance regimes, and are useful at all but the finest scales of biotic responses(productivityand competition) and vegetationpattems (individualorganisms).(From Delcourt et al., 1983.) eastern North America and western Europe. A brief outline of the environmental history of the last 1.6 million years (the QuaternaryEra)sets the stage for describing the response of vegetation to environmentalchange duringthe last 18,000 years of the recent post-glacialperiod. Within this context we will examine the relevanceof recent developments in the field of paleoecologyto the fields of community ecology, evolutionarybiology, and environmentalscience. III. QuaternaryEnvironmentsand Vegetation A. CLIMATICCHANGEDURING THE QUATERNARY The QuaternaryEra encompasses the last 1.6 million years and some 24 glacial events (van Donk, 1976), and is recognizedas 'apredominantlycold periodfollowing the relatively warm and equable Tertiary(Nilsson, 1982). Within the Quaternary, glacialstageshave usuallylasted 50-1 00,000 years,with a 4 1,000 yearcycle dominant in the early Quaternaryand a 100,000 year cycle dominant after 700,000 years BP (Shackletonet al., 1988). During glacial stages largeice sheets slowly accumulatein high latitudesin an oscillatoryfashion. These glacialstagesgenerallyend with a rapid decay of the ice sheets, followed by relativelywarm interglacialsof 10-20,000 years similarto the currentHolocene Epoch,which beganabout 10,000 yearsago and may end within the next few thousand years (Emiliani, 1972; Imbrie & Imbrie, 1986). Thus glacial stages are periods of relative climatic stability followed by rapid environmental change and relatively brief interglacials. Shorter-termenvironmental changes occur within this glacial-interglacialtime frame. The alternatingwarm and cold periods of the Quaternaryare generallybelieved to result from changesin insolation caused by cyclical alterationin the earth'sorbit around the sun as set forth by the Milankovich theory (Bergeret al., 1986; Milan- IMPLICATIONSOF PALEOECOLOGY U post, < w 207 future5 E 3 zW 24.00 w ac 250 200 50 150 0 -50 100 -1OO THOUSANDSOF YEARS AGO Fig. 2. Changesin the geometry of the earth'sorbit duringthe last 250 ka and projected for 100 ka into the future.Glacial-interglacialcycles roughlycorrespondto periods when the extremesof these cycles reinforceone another.(From Imbrie & Imbrie, 1986.) kovich, 1941). This theorydescribesthreecomponentsof the geometryof the earth's orbit, each with its own long-term cycle, that combine to give an overall cyclicity. Eccentricity(the degreeto which the earth'sorbit deviates from being circular)varies from 0%to 5%or 6%on a 100,000 year cycle. Obliquity(the tilt of the earth'saxis) changes from 22 to 24.5 degrees on a 41,000 year cycle, and primarilyaffects the latitudinaldistributionof solarradiation.Precession(thetime of yearwhen the earthsun distanceis a maximum or minimum)altersapproximately10%on a 22,000 year cycle, and affectsseasonality.Long-termclimatic variationis causedby the interplay of these three cycles (Figs. 2 and 3). Climatic changesare variablebecausethe three cycles have differenteffects at various latitudes;they also amplify and/or interfere with each other at various times and cause time-transgressiveevents like glaciation and oceanic heating/cooling. In general, warm and cold stages in the Quaternary roughlycorrespondto periods when these cycles amplify one another,althoughthe effectsof orbitalforcingare modified by surfaceboundaryconditions, positive feedback, and lag effects. The occurrenceand durationof glacial cycles duringthe Quatemaryis well documented by the study of glacial geomorphologicalfeatures,fluctuationsin ocean and lake levels, stable isotope ratios in ocean sediments and ice cores, planktonicassemblages in ocean and lake sediments, plant (especially pollen) and animal fossils in lake and bog sediments, and changes in sedimentation and chemistry in peat and lake sediments (Bradley,1985). In particular,radiometricand paleomagneticmethods of dating and stable isotope analysis revolutionizedthe field of paleoecologyin the 1950's and 1960's (Cox et al., 1963; Godwin, 1980; Shackletonand Opdyke, 1973; Thompson & Oldfield, 1986). These methods helped to extend previous estimates of the number of glacial episodes duringthe Quaternaryfrom 4 to about 24 THE BOTANICALREVIEW 208 a 3~~~~~~~~0 0. 4 1" c 0 a 120 10 0 4m 10 low1 1 _~~~~~~ c 6 cr 10 15 20 YEARS AG 0 2! 3 O ~~~~~~~~~~~~0 U, 0 CO ~~~~~~~~~6 5 so 75 YEARS 1200 t ISO78 AOX10 Fig. 3. Global temperature(a-A)and ice volume (e) on differenttime scales shows that we are currentlyliving at the end of a briefwarm periodin a predominantlyglacialclimate. Note that interpretationof apparentpatternsdepends on the scale at which they are viewed. For example, the dramaticrise in temperaturefrom 1880 to 1960 (a) appearsto be a continuation of a steady warmingtrend since the end of the little ice age (b), which in turn appearsto be a briefreversalof a slow cooling trendbeginningabout 6000 yearsago when globaltemperatures duringthe currentinterglacialreachedtheir maximum. (From Webb, 1986.) IMPLICATIONSOF PALEOECOLOGY 209 (Fig. 3). They also provided a more detailed chronologyfor these glacial cycles and for shorter-termevents such as the environmentaland biotic changes since the end of the most recent glaciation. A few marine and terrestrialrecords show climatic changes at the frequencyof Milankovitchcycles for at least the past 3 million years(Hooghiemstra,1984; Shackleton et al., 1988). This suggeststhat orbitalcycles, albeit unaccompaniedby glaciations for the most part, have occurredthroughoutearth history and have gone undetected by temporallycoarse geologic evidence (Bennett, 1990). The nature of late-glacialand Holocene climate change is being refined through the use of generalcirculationmodels coupled with a global networkof paleoclimatic data (Bartleinet al., 1989; COHMAP, 1988; Kutzbach & Guetter, 1986; Webb et al., 1987). Simulationsrun on these models are in reasonablyclose agreementwith the paleocimatic data, and the integrationof these data and models are the basis for the followingbrief summaryof the climatic history of North Americafor the last 18,000 years. At the time of the last glacial maximum 18,000 years ago, solar insolation was similar to that of the present (Fig. 4), but global climate was considerablydifferent, with air and sea-surfacetemperaturesgenerally lower, and global air and ocean circulationaltered.The large3 km thickLaurentideice sheet coveringmost of Canada and the northeasternUnited States split the westerlyjet stream,which was stronger than it is presently,into northernand southernsectors. The more southerlytrackof the jet stream brought more moisture to the American southwest, while a strong anticyclone centered on the ice sheet brought drier air to the Pacific Northwest. Although the central and eastern United States were colder than at present, cold arctic air masses were trappedbehind the ice sheet in the Arctic Basin. When this air mass was able to flow down to the mid-continentplains duringwinter, it would have been warmed adiabaticallyby 200 to 300C.Yet summer temperatureswould have been cooler nearthe ice sheet. Thus the climateof the easternand centralUnited States, although cooler than at present, may have been less seasonable and more equable. From 18,000 to 11,000 years BP the earth-sun distance decreased in northern summer and the axial tilt increased,thereby increasingaveragesolar radiationand seasonalityfor northernlatitudes. Ice sheets stabilized and began to retreatas early as 16,000 years BP, althoughwidespreadmelting did not begin until 14,000 years BP. By 12,000 yearsBP increasingsolarinsolation(andperhapsfeedbackmechanisms such as bedrockrebound,increasedicebergcalving,moisturestarvationand elevated CO2concentrations)reduced the Laurentideice sheet so that it no longer deflected a somewhatweakenedwesterlyjet streamnorthward.Maximumratesof glacialdecay lagged the solar insolation maximum at 11-10,000 years BP by about 1000 years, as did maximum mid-continentaltemperatures.The transitionfrom late-glacialto Holocene climates from 14,000 to 10,000 years BP was not continuous spatiallyor temporally.Climatechangeacceleratedto a maximum towardthe end of this period, and at least one significantcool interval (possibly resulting from oceanic cooling caused by rapidly decaying ice-shelves) is recorded for many sites adjacent to the North Atlantic Ocean before a rapid warming that marked the beginning of the Holocene about 10,000 years BP (Fig. 3). From 9000 to 6000 years BP summertemperaturesreachedtheir maximum of 20 to 4?Cabove present(exceptat the boundariesof the rapidlymelting ice sheet), and thereafterdeclined to present values. During this time climatic regimes in the west THE BOTANICAL REVIEW 210 18 15 12 9 6 3 I I I I I 1 Aerosol 0 ka 1 8 c00 330 0 E N 0~~~~~~~~~~~~~~~~~. 0~~~~~~~~~' c CO) C/ 00 18 15 12 9 6 3 O ka globalclimatesimulain theCOHMAP conditionsforvariousparameters Fig.4. Boundary departure frompresentvaluesfor: tion modelforthe past 18 ka, expressedas the percentage and December-February, SJJA,SDIF,NorthernHemispheresolarradiationfor June-August Landice is expressedas the temperature. SST,globalmeanannualsea-surface respectively; fromthatof 18 ka.CO2iSpartspermillionby volume,andexcessglacialpercentdifference scale.(FromCOHMAP,1988.) ageaerosolis drawnon an arbitrary approached modemnvalues as temperaturesincreased, and precipitation patterns shifted from predominantlywinter to summer monsoonal in the southwest;in the PacificNorthwesttemperatureswerewarmerand moisturelevels lower than at present until 6000 years BP (COHMAP, 1988; Ruddiman & Wright, 1987). Although paleoclimatic data are adequate only for North America and western Europe,the reasonablyclose agreementof the data with generalcirculationmodels suggeststhe following:First, complex climatic changeshave occurredover the entire earth, driven by orbital forcing,yet modified by the interplayand feedbackof such factors as changes in ice sheet albedo, extent of sea ice, icebergcalving, jet-stream consequentoceanic deflectionby ice sheets, sea surface-landtemperaturediffierences, and atmosphericcirculation,and elementalcycling.Second,climate has variedalong a continuum of space and time, and climatic changes may have been time-transgressive with lag effects in certain regions such as at the boundaries of ice sheets. Third, there were past climatic regimesfor which there are no modern analogs.For example, thereare probablyno modernclimate analogsfor North Americanclimates from 18,000 to 12,000 BP, or for the increased seasonality in the Northern Hemisphere approximately9000 years BP (Fig. 4). IMPLICATIONSOF PALEOECOLOGY 211 B. EVIDENCE FOR BIOTIC CHANGE The evidence for long-termvegetationalchangecomes from fossil plant partssuch as pollen, seeds, leaves and twigs contained in lake muds and peats, and in drier environments,pack-ratmiddens (Berglund,1986; Betancourtet al., 1990; Birks & Birks, 1980). Most late-glacialand Holocene vegetationaldata come from studies of fossil pollen in which samples are taken from lake and peat sediments. The pollen within these sediment samplesis concentrated,sub-sampled,counted, and displayed on a pollen diagram(Fig. 5). Macrofossilssuch as seeds, leaves, and twigs offermore taxonomicand spatialresolutionthanpollen, but areless abundantin most sediments so that problemsof samplingand temporalresolutioncan arise. Informationderived from fossil pollen lacks the spatial, temporal and taxonomic resolution of living material,but offersa long-termperspectiveunavailableto ecologistswho studyliving organisms. The principlesof pollen analysis, brieflyenumeratedbelow, have been rigorously tested and refinedfor over 70 years. 1. Most pollen and spores are producedin abundance. 2. Most of these fall to the groundfrom a well-mixed pollen rain that represents the surroundingvegetation. 3. Pollen is preservedunder anaerobicconditions, for example in lake and bog sediments. 4. Smallamountsof sediment(< 1 cm3)containenoughtaxonomicallyidentifiable pollen (up to 106 grains)to representthe surroundingvegetationat the time the sediment was deposited. 5. Pollen spectraobtained from several levels of a sedimentarycolumn represent the history of the vegetation at that place. 6. Two or more series of pollen spectra obtained from differentsites represent changesin vegetation at differentplaces. Pollen analystshave takengreatstridesin understandingdifferentialpollenproduction, dispersal, deposition and preservation (Andersen, 1974a, 1974b; Calcote & Davis, 1989; Clark, 1988a; Cushing, 1967; Davis, 1973; Davis & Brubaker,1973; Havinga, 1967; Jackson, 1990; Raynoret al., 1975). Recent advanceshave also been made in sampling,numericalanalysis and multivariatetechniques(Berglund,1986; Birks & Birks, 1980; Clark, 1988b; Gordon & Birks, 1985; Green, 1981, 1988; Jacobson & Grimm, 1986; Maher, 1972, 1981; Prentice & Webb, 1986; Schwartz, 1989; Webb et al., 1981). These studies have helped to define the taxonomic, numerical, spatial, and temporal confidence limits of pollen data, and have provided methods for reducingthe complexity of the data so that pollen assemblagescan be interpretedmore clearly with regardto stratigraphicrelationships,environmental gradients,and rates of change. Paleoecologistscan addressvarious ecologicalproblemsalong a broad continuum of spatialand temporalscales throughthe carefulselection of study sites, organisms, and field, laboratoryand analytical techniques. In general paleoecologistswant to know about the presence, abundance, location, and temporal occurrenceof taxa, communitiesand environments.Many studiescombine pollen analysiswith analyses of plant/animal macrofossils, charcoal, sediment lithology and chemistry, human artifacts,tree-rings,historical records,and modem vegetation. These studies reveal LAKE OF THE CLOUDS, MINNESOTA A. M. Swain 1973 Charcoal Influx Vorve Thickness Conifer/Sprouter RatRioo_ DEPTH DATES (cm) A.D .1 ./ TOTAL POLLEN INFLUX Pol Ion _ _ _ _ >400 000 _ _ _ _ _ _ _ _ _ _ _ lA' 49=0 -490 -4864 -40'___________- 6000 -4739 -4728 40 40 00 * *300 4300~~~~~~~~~~~ 0 5 o n4 X t462/cm0/y0or 0 o * n o mm 200 2 25 0 2 4 6*0 0 1000 +00X 40.0 0200 0* 00 mo0 Po0ll v grins 0 or IMPLICATIONSOF PALEOECOLOGY 213 patternsof climate change,naturaland anthropogenicdisturbance,soil development, erosion, community composition and diversity, migration,invasion and succession (for examples and techniquessee Berglund,1986; Birks & Birks, 1980; and Huntley & Webb, 1988). A few examples illustratethe spatial, temporaland ecologicalresolutionavailable at various scales (Fig. 1). Tree-ringanalysis, vegetationreconstructionand historical recordsprovidevery fine resolution,often comparableto data from permanentplots. Henry and Swan (1974) and Oliver and Stephens (1978) analyzedthe growthrates of live and dead stems on small plots (0.04 and 0.36 ha respectively)in conjunction with historicalrecords.They detecteddisturbancessuch as fire,windthrowand treecutting,and subsequentchangesin vegetationcomposition.Manystudies(e.g.,Foster, 1988; Heinselman, 1973; Hemstrom & Franklin, 1982; Romme, 1982) have used less intensive techniques to document disturbance and vegetation change at the landscapelevel. Tree-ringsalso provide paleoclimaticinformation at the local, regional and global level, detectingphenomenasuch as the "little ice age" 1450-1850 A.D. (Fritts, 1976). Annual laminations in lake sediments may provide yearly records of vegetation and environmental change (Saarnisto, 1986; Clark, 1990). Swain (1978) analyzed varved sediments from Hell's Kitchen Lake in north-centralWisconsin for pollen, charcoal and seeds. He found a successional sequence of aspen, birch, white pine, and hemlock following fires whose frequencyof 100 to 140 years was affectedby available moisture of the prevailingclimate. He concluded that major vegetational changeoccurredon two differenttime scales.Short-termchangesrelatedto individual fireswere superimposedon longer4termchangesresultingfrom increasesor decreases in the frequencyof these fires. Basin size (in addition to morphometry,in/outflow, and surroundingvegetation) determines to a large degree the source area of pollen falling onto the surfaceof a lake or pond (Fig. 6). Moderatesized lakes (10 to 100 ha) receive pollen mainly from within a 20 to 30 km radius.Finer spatialresolutionis availablethroughthe analysis of mor humus and sediments from small forest hollows (Bradshaw, 1988). These sites, overhungby a tree canopy that contributesabundantpollen and also filtersthe regionalpollen rain, collect pollen from a smallersource area than ponds and lakes, allowing stand-level interpretationsof vegetationalchange over thousands of years in some cases (Andersen, 1970; Jacobson& Bradshaw,1981; Mitchell, 1988). Aaby (1983) analyzedpollen and fungalhyphae from two soil profilesonly 7 m apartand found clear differences,substantiatedby historical records, between the two sites. Andersen (1984) analyzed pollen from a suite of small hollows and soil profilesin the Eldrupforest in Denmark, producinga detailed history of forest change at the stand level. Analysis of pollen from small hollows and mor humus is a well known techniquein northernEurope,but has been adopted by North Americanpaleoecologists only recently(Davidson, 1985;Frelichet al., 1989;Heide, 1984; Schoonmaker & Foster, 1989; Schwartz,unpubl.).Although mor humus and small forest hollows have been thoughtto collect most of theirpollenfrom a radiusof 20 to 30 m (Jacobson Fig. 5. Charcoalinflux, varve thickness,and pollen influx for the last 1000 years of Lake of the Clouds,Wisconsin. Sedimentin this lake is depositedin discreteannuallaminationsor varves, which allows for precisedating of events. A fire frequencyof 60 to 70 years (peaksin the charcoal influx marked by stars or dates) has maintained a relatively stable vegetation composition. (From Swain, 1973.) THE BOTANICAL REVIEW 214 (a) x I A ~ 4 ~~(b) I 9.4* A N 20kA (c) a 8~~~~~~~~~ 1m20m Fig. 6. a) Aerialview of hypothetical studyareawithpond(y) andforesthollows(x). b) Viewof pondwithpollensourcearea(diameterA-A')andc) hollowwithpollensourcearea (diameterB-B').(FromBradshaw, 1988.) & Bradshaw,1981;Prentice,1985), recentevidence suggeststhat the pollen collection propertiesof these sites vary considerably,and that extra-localand regionalpollen may contributea substantialamountof pollen input (Calcote& Davis, 1989;Jackson, 1990; Schoonmaker& Foster, 1990). Pollen analysts must be careful to test and acknowledgethe spatial and temporal limits of data from small hollows and mor humus. A combination of fine spatial scale pollen analysis and vegetation reconstruction should producea long recordof both high spatial and temporalresolution(Schoonmaker & Foster, 1990). Reconstructionof the establishment,growth and death of live, dead, and downed stems provides a spatially and temporallydetailed 100 to 1000 year history of vegetation dynamics. These data can be used to interpretthe longer but less spatially precise record available through pollen analysis. Because seeds, leaves and woody plant partsare less mobile than pollen, macrofossilanalysis should be included in studies whose aim is high taxonomic and spatial resolution (e.g., Jackson, 1989, 1991). Landscape-levelphenomenacan be investigatedby selectingsmall to medium size basins (that collect pollen from the surrounding10 to 100 km2) at distances such that pollen sourceareasdo not overlapextensively (Gaudreauet al., 1989). Brubaker (1975) analyzedpollen from lake sediments on three differentsoil types in Michigan and found that the same regionalclimate change resultedin differentcommunities on differentsoils. Gaudreau(1986) compared two sites less than 15 km apart but with an elevational differenceof 440 m in the northeasternUnited States and found that physiographicposition affectedthe arrivaldates of migratingplant species.Clark (1990) combined a stratigraphicanalysis of charcoal on petrographicthin sections IMPLICATIONSOF PALEOECOLOGY 215 from varved sediments from three small (5 ha) lakes having adjacent catchments with a study of firescars on red pine trees. This combinationof techniquesproduced a detailed picture of fire history and environmentalconditions for a 1 km2 area. Radiocarbon-datedpollen spectrafrom thousandsof basins throughoutthe world have been analyzedin the last few decades (Fig. 7). This largenumberof late-glacial and Holocene paleoecologicalstudies has enabled the detailed mapping of the distribution of taxa throughtime on a regionalor subcontinentalscale in Europeand North America (Figs. 8-11). Huntley and Birks (1983) presentedisopoll maps (contour lines connecting similar pollen percentages)of pollen sampled from 423 sites in Europe covering a period from 13,000 to 0 years BP with a resolution of 500 years. Bernabo and Webb (1977), Gaudreau(1986, 1988), and Webb et al. (1983) have drawn similar maps for the easternUnited States (Fig. 8). Davis (1981), Gaudreauand Webb (1985), and Webb et al. (1983) constructedisochronemaps (contour lines representingmigrationalfronts of taxa at differenttimes in the past) for eastern North America(Fig. 9). Delcourtand Delcourt(1981, 1985) and Davis and Jacobson (1985) mapped inferredvegetationfor easternNorth Americaduringthe late-glacial and Holocene. Most recently, Delcourt and Delcourt (1987) have produceda large set of maps based on 162 sites in easternNorth Americathat presenttree percentages at differenttimes during the Holocene as interpretedby modern pollen/vegetation analogs(Fig. 10). Gaudreau(1984) and Jacobsonet al. (1987) mapped differenttaxa simultaneouslyusing colored isopolls to reveal population centers and overlap of differenttaxa at - 500 year intervalsover the past 18,000 years (Fig. 11). This latter method may offerthe most insight into species interactions.A furtherstep might be computer enhanced interpolation providing continuous output. These and other studies carefullyselect the data for properand consistent analyticaltechnique,time control and resolution. As more sites become available, the spatial and temporal resolution of the maps will continue to improve. C. VEGETATION CHANGE DURING THE LATE-GLACIAL AND HOLOCENE The maps of changingvegetationas implied by changingpollenpercentagespresent the following picture:At the time of maximum glaciation 18,000 years ago, when ice covered much of northwesternEurope, Canada and the northeasternUnited States, the vegetation in the area immediately south of the ice sheet (the periglacial area)consisted of species drawn from a rangeof present day environmentsand life history strategies(pioneer,ruderalherbs, arctic-alpine,steppe).Some of these communities, which included sedges, grassesand taxa such as Alnus, Betula, Dryas, and Artemisia,might be recognizedas tundraor steppe (Birks, 1986). Othercommunities such as the open polardesert(dominatedby chenopods,grasses, and Artemisia)that occupied northwesternEuropebetween the ice sheet and the Alps have no modern analogs (Iversen, 1973). The absence of modern analogs for fossil assemblagesis not so surprisingwhen we consider that climatic and edaphic conditions duringthe past were not merely differentfrom those of the present,but may have combined to create unique habitats resulting in unique vegetation, for example, spruce-poplarand spruce woodland-tundraassemblages(Huntley, 1990; King, 1981; McAndrews, 1965; Prentice, 1986; Wright, 1987). South of the 50- to 100-km-wide periglacialareas, steppe and galleryforests occurredin Europewith true forestsfound south of the Alps, especiallyin southeastern 0~~~~~~~~~~~ F Cs aclveaa. 0000n Mide Daa u~~~~~~~ it~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~n a D S 6 (8kaol) Fig. 7. Location of sites in COHMAP global paleoclimate database,which does not include all rad number of sites in the NorthernHemisphere, especially in eastern North America and western Europe (From COHMAP, 1988.) IMPLICATIONSOF PALEOECOLOGY 217 Europe in locally mesic refugiasurroundedby an otherwise and environment supporting steppe and possibly sclerophyllwoodland (Bennett et al., 1991; Huntley & Webb, 1988;Van derHammen et al., 1971). In North Americasouth ofthe periglacial area, an open spruce and larch forest dominated in the central United States well into the Great Plains, with jack pine becoming more importanttoward the Appalachians and reachingdominance along the Atlantic Coastal Plain and southeastern United States. Deciduous trees may have occurredas minor elements throughout much of the conifer forest (Jacobsonet al., 1987). These deciduous species became more importantin the lower MississippiRiver Valley and had isolated refugiain the southern Appalachians(Delcourt & Delcourt, 1977; Delcourt et al., 1983; Wright, 1983). As temperaturesincreased,ice sheets beganto retreatin a spatiallyand temporally patchy mannerabout 14,000 years BP. Species on the peripheryof the glacialboundary invaded exposed substratefollowed by species from furthersouth. In Europe, boreal species such as pine and birch invaded followed by elm, beech, hornbeam and spruce,amongothers.Althoughmost taxa migratedfrom southeastto northwest, each took a somewhat differentroute, and moved at a differentand variable rate (Huntley & Birks, 1983). Spruce,a late-comer, migratedalong a double front both westwardinto centralEuropeand northwestinto Scandinavia.Birch and pine have continued to migrate southwardalong the Scandinavianpeninsula in the last few thousand years. As the ice retreatedin eastern North America, tundra species preceded spruce, jack pine and balsam fir, which moved northwardat differentrates and in different directions,creatingopen and then closed conifer forests. Furtherto the south, white pine and hemlock (Fig. 9) migratedup the Atlantic seaboard,while oak appearsto have migrated north-northwestwardalong a broad front. Following these species, elm, maple,beech and chestnutappearto have moved in that orderfromthe southern Mississippi River Valley or further west, while hickory diverged into two fronts apparentlysplit by the higher Appalachians.Not all species migratedin a general south to north direction. Duringthe late-glacial,ash appearsto have been restricted to small microhabitatssouth of the ice sheet in the midwest. In the late-glacialto early Holocene, ash expanded its range, becoming a component of a spruce-ashpoplarassemblage,and then partof an ash-ironwood-elm asemblage,beforeits range began to shrinkagainabout 10,000 years BP. Duringthis time the geographiccenter of ash's rangeremainedrelativelyfixed,just to the south of the GreatLakes(Jacobson et al., 1987) (Fig. 11). Birch appearsto have expanded to the southwest from the northeasternUnited States and then later northward(Gaudreau,1988) (Fig. 8). As in Europe,each species took a somewhat differentroute (generallynorthward)and migratedat variable rates (Davis, 198la) (Table I). When we inspect the isochrone maps (Fig. 9), we can clearly see that differenttaxa migratedat differentrates, from differentorigins and in differentdirections. Hickory invaded deciduous forests in Ohio 4000 years before beech, but beech arrived in southern New England 3000 years before hickory. Beech and hemlock arrivedin the AppalachianMountains at about the same time, but hemlock migrated more quickly thereafterand reached centralNew Englandand upper Michigan 2000 and 2500 years respectivelybefore beech. Some species such as chestnut were still migratingnorthwardsas recentlyas 2000 years BP, thousands of years after a well-developed forest had become established and climatic cooling had begun. Most species have retractedtheir ranges somewhat to the south in response to this recent cooling (Davis, 1983). The inde- 0 : 8~~~ 112 ~~~ w IMPLICATIONS OFPALEOECOLOGY L a 0B O Fm I 12 1 I 219 I oto o~~~~~~~~~~~~~~~~~~~~~~~~~~i 0 Fig. 9.ThemigraionfrontofHemlock o 400km 40k 6 6 10~~~~~~~~~~~~~~~~~~~~1 10 15 dentoto Chestnut 0 eoltonr 400 km Fig. 9. The isochrone migrationfrontof hemlock, tionsfor and chestnut depicted by coloicaltheor. Wht wesee beech, odayhickory, s meely sna-sho ofy aSynm ic maps (12 ka to present)with lines connectingarrivaldates of taxa as inferredfrom increases in pollen or presenceof macrofossilsat 1000 yearintervals.Heavy lines show presentor recent rangeof taxa. (From Davis, 1982.) pendent response of organisms to changing environments has far reaching implications for ecological theory. What we see today is merely a snap-shot of a dynamic and changing environment and biota that have come about through evolutionary and historical forces operating on several temporal and spatial scales. Fig. 8. Isopoll map for birch pollen 14 ka to presentwith contourlines connectingvalues of 5%(stippled),20%(diagonallines), and 40%(black)birchpollen. Birchappearsto have had a more northerlyrangeduringthe last glacial period than many other arborealtaxa. Ice sheet appears in the northem part of 14, 12 and 10 ka maps. Dots are fossil-pollen sites. (From Gaudreau, 1988.) THE BOTANICALREVIEW 220 * S TRU 40 A600pr L-P A.THWNTREOFTE ITI LP CMUNT The nature of the biotic community has long been a source of disagreement and controversy among ecologists (Pickett & Davis, 1990). The "organismal" concept of Clements (I1916)was criticized by ecologists who recognized the individualistic nature of species within assemblages (Cain, 1947; Gleason, 1917, 1926; Livingston &Shreve, 192 1; Ramensky, 1924) and the continuous (vs. discrete) distribution of species along environmental gradients (Curtis, 19 55, 19 59; Curtis & McIntosh, 19 51; Whittaker, 1951, 1956, 1975). It is interesting to note that Clements recognized the importance of disturbance in structuring some communities (Clements, 1910), and was well aware of the paleoecological perspective (Clements, 1924). Recent definitions of "communitiy'' have been purposely vague, ranging from "6a set of species populations that occur in some place" (Schoener, 1986) to "all orpanisms in an area" usually restricted for the convenience of the researcher "spatially, trophically, taxonomically or by life form" (Roughgarden & Diamond, 1986). A temporal restriction is not mentioned, but is often implicit (i.e., the time period during which the community is studied). IMPLICATIONSOF PALEOECOLOGY . ... * Fig. 10. t IWr --;- l,\ 221 wA . ~~~~m -TOP ~~t ::m Continued. Severallines of paleoecologicalevidence suggestthat plant communities are loose and relatively ephemeral assemblages of species populations. The individualistic response of tree species to climatic change described above, and the apparentease with which species migrate into and subsequentlyleave assemblagesdemonstrate the absence of cohesive communities. For example, chestnut invaded the mature hardwoodforest of the northeasternUnited States 2000 years ago with only modest readjustmentsin the densities of resident species (Davis, 1983). When chestnutwas eliminated as a canopy dominant in these forests in the first half of this century, other species, especially oaks and hickories,merely increasedtheir abundance(Keever, 1953; Mackey & Sivic, 1973). The sudden and temporarydecline of hemlock in North America, and of elm in Europearound 5000 years BP also resultedin the increaseddominance of remainingspecies (Allison et al., 1986; Davis, 198lb, 1983; Iversen, 1973). Both hemlock and elm recoveredwithin a few hundredto a thousand years and rapidlyregainedtheir formerabundance.Thus forests are relatively open communitiesthat may sufferapparentlylittle reorganizationor ecosystem disruption upon the invasion or decline of a dominant species. The ephemeraland transitorynatureof plant communities is demonstratedby the independentmigration of species and by historical changes in abundanceand distribution of modern species assemblages. The forests with which we are familiar 222 THE BOTANICAL REVIEW 14 ka 8 ka ka0 12 6Ska 0 k- Oska Fig. 11. Isopoll maps, 14 ka to 0.5 ka, with ice sheet to north stippled.Elm (lightgrey),ash (heavy outline), and ironwood (symbol < >) showing overlappingranges(e.g., dark grey for elm-ash). The rangesof all three taxa have decreasedthroughoutthe Holocene, to the point that their presentrangesmight be consideredrefugia.(From Jacobsonet al., 1987.) today are composed of species that arrived at differenttimes from various glacial refugia,respondingindependentlyto changingclimatic and disturbanceregimesand biotic interactions.As a particularspecies migratedin response to climatic change, it would have encounteredother species to form dynamic and constantly changing assemblages.At the height of the last glaciation, a sprucewoodland-tundraassemblagewith no modernanalogexisted in the centraland easternUnited States(Wright, 1984, 1987); several thousand years later, the hardwood(black ash-ironwood-elm) component of this community was left behind as conifers moved northward(Jacobson et al., 1987). By the time most tree species were migratingnorthward(12,00010,000 yrs BP), ephemeral communities, such as the black ash-oak-spruce-graminoid assemblage in north central Illinois (King, 1981) were not uncommon. The animal fossil record also has many examples of no-analog assemblages or what Graham (1986) calls "interminglings"of species that have separate ranges today. IMPLICATIONSOF PALEOECOLOGY 223 Table I Averagerates of late-glacialto Holocene rangeextensions in easternNorth America (modified from Delcourt & Delcourt, 1987) * Genus Rate (m/yr) Salix Populus Betula Tilia Tsuga Larix Fagus Abies Picea Juglans Pinus (Northerngroup) Ulmus Acer Quercus Fraxinus Carya Castanea Pinus (Southemgroup) Nyssa 287 263 212 209 202 189 169 159 141 140 135 134 126 126 123 119 100* 81 70 From Davis, 1982, roughlycomparableto data from Delcourtand Delcourt, 1987. This is not to say that all speciesassemblageshave been ephemeralat all times. There are examples of constancy over long periods of time (Delcourt & Delcourt, 1987; Green, 1982; Jacobson& Grimm, 1986), but in general,changeappearsto be more characteristicof the late-Quaternary. B. EVIDENCE OF DISTURBANCE IN THE PALEOECOLOGICALRECORD We define disturbancehere sensu White and Pickett (1985) as "any relatively discreteevent in time that disruptsecosystem, community or population structure, and changes resources,substrateavailability or the physical environment"and include both destructiveevents and environmentalfluctuations.Most ecologists consider disturbancein a diurnal, seasonal, decadal, or centennial time frame, yet the disturbanceregime (sensu Heinselman, 1981) of destructive events and environmental fluctuationswith profoundeffects for the vegetation of an area is best characterizedover longertime periods.The use of the long paleoecologicalrecordenables ecologists to avoid the space-for-timesubstitutionthat, althoughemployed by most successionalstudies, may be highly misleadingbecause of the difficultyof assuring between-siteuniformity(Clark, 1990; Pickett, 1988). Naturaldisturbancesof varyingsize, frequencyand severityhave beendocumented throughoutthe paleoecologicalrecord.Fires may be detectablein sediment as charcoal layersand as peaksin charredparticlescountedon slides preparedfrom sediment (Clark,1988a, 1988b;Pattersonet al., 1987) (Fig. 5). Geomorphologicalfeaturesand sediment lithology record erosional events (Campbell& Fraily, 1984), and tephra layers indicate volcanic eruptions (Sarna-Wojcickiet al., 1983). Sudden changes in pollen assemblagesoften correlatewith catastrophicevents or anthropogenicactivity 224 THE BOTANICALREVIEW I ,I T p-w1 CI 0 Woodland only 0 W O or woodland 0 ~ I ? ~~~~Prairie only tw Time period -- Fig. 12. SchematicdiWam showinga two-dimensionalcrosssectionofthe subspacesin which prairieand oak woodland will persist in an n-dimensionalhyperspacein which the axes are time period and the independentclimatic variablesinhuencingfire frequency.The curves are the thresholdsfor the changesfrom oak woodland to prairieand the reverse from prairieto oak woodland. (From Grimm, 1983.) (Brugam,1978; Green, 1981). The paleoecologicalrecordalso reveals changingpatterns of disturbanceas environmentalconditions change.Grimm (1983) documents the invasion of oak woodland in southernMinnesotaby more mesic tree species as the result of decrasing fire frequencycaused by the cooler, moister climate of the "littleice age."He presentsa model suggestingthat climate,physiography,vegetation pattern and fire interactto cause thresholdsfor vegetation change to vary in space and time (Fig. 12). Thus differentvegetationtypes can persistadjacentto one another in essentiallyidenticalenvironments,their locations bemighistoricallycontingenton past disturbances. The paleoecologicalrecord allows us to better understandthe spatial as well as temporal dynamics of disturbance.The extent and nature of discrete and periodic events may not be observable over an entire landscapeor region in a lifetime. For example, in southern New Englandthe overall frequencyof fire is relatively low. However, studies of charcoaland pollen along a transectof sites suggeststhat a very stronggradientof decreasingfire frequencyfrom the coastal oak-pine forests to the IMPLICATIONSOF PALEOECOLOGY 225 northernhardwoodforest parallelsa climate/vegetationgradient(Patterson& Backman, 1988; Winkler, 1986). The history of coastal oak-pine vgetaion repeatedly returningafterfireillustratesanotherpoint:certaintypes ofvegetation areremarkably resilient Recent and paleoecologicalevidence confirm that the regionalvegetation mosaic in communities with fire-dependentspecies appears to have been maintained by naturallyrecuning fires(Cwynar,1987; Habeck & Mutch, 1973; Heinselman, 1973; Minnich, 1988;Romme, 1982;Swain, 1978).In these communities,fireis responsible for long-termstability;the lack of fire,owing to climatic changemight be considered more of a disturbanceto the overall system, causingwidespreadspecies replacement (Grimm, 1983). Conversely,dwing a time of climatic change,a community showing some inertiamay be opened up to invasion by fire.Green (1982) documentedrapid, long-term species replacementfollowing fire in the early Holocene in Nova Scotia. Disturbancein tropical areas has been characterizedas less severe than that in temperate regions (Connell, 1978; Whitmore, 1975), but there is evidence to the contrary,such as hurricanesthat have their genesis in the Tropics of Cancer and Capricorn(Lugo et al., 1983; Wadsworth& Englerth,1954), and the 1983 El Ninio event that unleased torrential rains along the eastern Pacific rim but resulted in droughtand fire in the western Pacific (Leighton& Wirawan, 1986). Nevertheless, stand-leveldisturbancein the tropics has received more attentionin the last decade (Brokaw, 1985; Denslow, 1980; Liebermanet al., 1985; UhI et al., 1988). The paleoecological record further extends the evidence for disturbancein the tropics at several scales. Catastrophicdeluges of rain along the west coast of South America even strongerthan that in 1983 have occurredperiodicallythroughoutthe Holocene and are especiallyevident in areasof human settlement(Nialset al., 1979). Sedimentologicaland geomorphologicalevidence suggestsequallycatastrophicflooding in the Amazonbasinduringthe Holocene(Campbellet al., 1985;Liu&Colinvaux, 1988). River channel meanders revealed by satellite photographyhave inundated forestswith the resultthat 12 percentof the forestalongriversin the wester Amazon is successional(Salo et al., 1986). Another 14.6 percent lies on the floodplain and has been disturbedin the recent past. Thus a quarterof this region of the Amazon basin has been disturbedby flooding probablywithin the last few hundred years. Evidence of fire has been found in tropicalrainforestsdated back to 6000 years BP (Saldarriaga& West, 1986; Sanfordet al., 1985). Past human activities have had major impacts on the landscapeand often serve as convenient time horizons. Neolithic man most likely caused the sudden decline of elm in Europe around 5000 years BP by girdlingtrees and using the foliage for animal fodder (Birks, 1986; Iversen, 1973). North American Indians are generally believed to have had a sophisticatedunderstandingof the use of fire as a tool for hunting and clearing land (Day, 1953; Patterson & Sassaman, 1988; Pyne, 1982). Human presence drasticallyaltered vegetation in parts of central America during Mayan times (Binford& Leyden, 1987). A distinct settlement horizon of Ambrosia in most palynologicalrecords throughoutNorth America is a consequence of the clearanceof forests for agricultureby colonists from Europe. C. COMMUNITY STABILITY AND EQUILIBRIUM With the recognitionthat disturbanceis an integralpart of many ecosystems and that climate has been changingcontinuously in the past, ecologists have begun to 226 THE BOTANICALREVIEW debate the relative importance of equilibrium vs. nonequilibriumexplanations of communitystructure(Connell,1978;Hubbell& Foster, 1986;Pickett& Davis, 1990). Organismsencounterand respond to environmentalchanges throughoutthe entire spectrumof spatialand temporalscales. Indeed, we must firstdefine the taxonomic, spatialand temporalscales at which we wish to investigatequestions of stabilityand equilibrium.We might ask if two populations are in equilibriumwith each other. Or whetheran entire community is at an equilibriumstate. Or whethera species is in equilibriumwith its environment.The answermay be positive at one intersection of the various scales under considerationand negative at another. A furthercomplication with pollen data is determininghow to quantify stability and change. Paleoecologists traditionallyhave recognized different pollen assemblagesor zones, and numerousmultivariatestatisticalapproacheshave been adapted to allow for more objective zonation (Gordon& Birks, 1985). Jacobsonand Grimm (1986) and othershave applieddetrendedcorrespondenceanalysis(DCA) to modem and fossil pollen data matrices. With appropriateapplication,the results show the rate and direction of change (Figs. 13 and 14). Using this method, Jacobson et al. (1987) have shownthat the highestratesof widespreadvegetationalchangein eastern North America occurredat two discrete times: 11-9000 years ago, reflectingrapid migration of taxa in response to climate change, and since European settlement, reflectinga response to pervasive anthropogenicdisturbance. An ongoingdebateamongbiogeographersand paleoecologistsconcernshow closely vegetationtracksenvironmentalchange.The consensus seems to be that the answer depends on the time scale under consideration,the rapidity and amplitude of the climate changeand the particularvegetationunderinvestigation(Prentice,1986). At a frequencyof 1000 years, vegetation usually appearsto be in dynamic equilibrium with climate (Birks, 1989; Webb, 1986). On shortertime scales, between 1 and 500 years, vegetation may often lag behind climatic change (Davis, 1986). In an environmentthat varies between long periods of glacial stasis and relatively abrupt interglacials,organisms must cope with conditions that may favor stable equilibriumcommunitiesduringglacialtimes and moredynamiccommunitiesduring interglacials.The paleoecologicalrecordprovides evidence that communities in the past and present have been both stable and unstable, and in disequilibriumand equilibriumstates(e.g.,Delcourt& Delcourt, 1987;Green, 1981;Jacobson& Grimm, 1986; McAndrews,1965; Payette & Filion, 1985; Ritchie, 1986). However, ecologically dynamic and nonequilibriumcommunities probablyhave been more characteristic throughoutthe Holocene as species migratedindependentlyin response to climatic change. The independent migration of tree species and their penetration of established deciduous forests in the northeasternU.S., the hemlock and elm declines, and the transient nature of species assemblagesin general demonstratethe disequilibrium nature of communities during the Holocene. Time lags shown by organismsin response to environmentalchange are evidence for both stability (in the sense that organismsremainwherethey are-inertia, sensu Gorham, 1957) and disequilibrium (in the sense that other organismsjoin or departfrom the assemblage).Cole (1985) has shown that some plant species growingnear the GrandCanyon at the beginning of the Holocene were slow to move northwardwith warmerclimatesand werejoined by fastermoving arrivalsfrom the south resultingin a transientspecies-richassemblage. Davis and Botkin (1985) have modeled tree species' responses to climate changein computersimulationsand predicted200 year responselags. Davis (1986) IMPLICATIONSOF PALEOECOLOGY 227 -0.2 0.0 _ Oucus-Gromho oN /D 0.2 0.4 O0.6 Gr ~~AdU1~O4K ?0 0 0u 5k * K ~~~~BeAWi-Cyperoceae BezPirws 0.8 1.0~~~~~~~~~~~~~~~~~~~~ 1.0 _ 12 1.4 AXISI Fig. 13. Directionand rateof vegetationchangein centralMinnesotaduringthe past 10,000 years. Ordinationof 100 yearsBP analogpollen assemblages(open and half-filledcircles),and a five-samplerunningaverageof fossil pollen for the last 10,000 yearsfrom Billy'sLake,central Minnesota(dots connectedby line). Points on the runningaveragecurverepresentthe position every 100 years (ratherthan the position of each sample).(From Jacobson& Grimm, 1986.) has also summarizeddata showing that various animals includingbarnacles,birds, insects and small mammalshave laggedanywherefrom a few yearsto decadesbehind environmentalchanges. Migrationmaps do show, however, that presenttemperate communities probablyhave been roughlyin equilibriumwith the environment on a regionalscale since about 2000 years BP. If instability and disequilibrium are characteristicof the Holocene, models of populationand communitydynamicsbased on equilibriumconditions,while heuristicallyuseful,may be even less realisticthanempiricalecologistsoftenclaim (Franklin et al., 1987). Most models of population growth,competition and predationignore migrationfor simplicity'ssake,but the paleoecologicalrecordsuggeststhat migration and dispersalare fundamentalto understandingthe above phenomena.Thus modelers should be exlicit about the time and space scales under consideration,and recognize that models of short time-scale phenomena may not extrapolateto longterm ecological or evolutionarytime scales. D. EVOLUTIONARY CONSIDERATIONS Paleoecologicaldata from the Quaternaryshould be of considerableinterest to those concerned with rates of evolution and speciation, and related processes includingthe evolution of diversity,coevolution,selection,competition,and extinction. Bennett (1990) has even suggestedthat Gould's (1985) three-tiertemporalhierarchy of evolutionary processes should be expanded to include glacial-interglacialcycles, thus yielding a four-tierhierarchyconsistingof: ecologicaltime (thousandsof years), 228 o 0 THEBOTANICAL REVIEW 0.20 c1 0.15S 0 o 0.10 z w o- 0.05 z. 0j 1 2 3 4 5 6 7 8 9 10O AGE(IOOO14Cyr BP) Fig. 14. Rate of vegetationchangein centralMinnesotaduringthe past 10,000 years.Graph of distance(numberof standarddeviations)moved every 100 yearsin the firstthreedimensions of the ordinationdepicted in Figure 13 vs. age. Greaterdistance indicates greaterchange in pollen spectrain 100 years. The graphshows high rates of vegetationchangefrom 10 to 8 ka, as foresttaxa were replacedby prairietaxa (see Fig. 13), relativestasis from 8 to 4 ka, and then higherrates of changeagain as differentforest taxa invaded from 4 ka to the present.Highest rates of changeare associatedwith settlement.(From Jacobson& Grimm, 1986.) orbitallyforcedclimate changes(20,000-100,000 years),geologicaltime (millions of years), and mass extinctions (approximately26 million years). Viewed over the entire Quaternary,taxa have constantlymigratedin responseto glacial-interglacialcycles, often resultingin alternationbetween widespreadpopulations in favorable times, and small, fragmentedpopulations during unfavorable periods.These oscillationsin populationsize, locationand geneticcompositionshould favor rapid evolution throughgeographicisolation and founder effects (Huntley & Webb, 1989). The rapidmigrationof tree species duringthe Holocene providesexamplesof how allopatric speciation by geographicalisolation might occur. Migration maps show taxa moving in two differentdirectionsat differentrates (e.g., Figs. 8 and 9). Rapid speciationduringthe last glacial-interglacialcycle is suggestedin the paleoecological record,e.g., the recentdivergenceof jack pine and lodgepolepine (Critchfield,1985; Cwynar& McDonald, 1987). On a longer time scale, sprucehas become rarersince the early interglacials,failing to reach Englandduringthe Holocene, while hazel has become more common. West (1980) speculatesthat these trends are likely a result of evolutionarychangesin thesetwo speciesbasedon the observationthatabundances of other species have remainedroughlysimilarduringQuaternaryinterglacials.Mengel (1964) shows how modern species of wood warblerscould have arisen from repeated migration of an ancestral species northwestwardfrom the southeastern United States during interglacialsand subsequentisolation of western populations upon glacialadvance.These examplessuggestthat environmentalchangesthat induce varyingratesand directionsof migrationand other individualisticresponsesplay an importantrole in determiningrates and directionsof speciation.Webb (1988), however, points out that the time scale of glacial-interglacialcycles may be too short for manypopulationsto develop majormorphologicaldifferencesand becomegenetically isolated. Speciesinteractionsshould be profoundlyaffectedby glacial-interglacialcycles. In contrastto a more stable environmentwhere species may have time to adapt to one IMPLICATIONSOF PALEOECOLOGY 229 another, species reacting individualisticallyto oscillating environments may form looser associations.A particularspecies migratingnorthwardin the Holocene would have competed with several species along its route, and even may have occupied various ephemeral communities in different places at different times or simultaneously. Thus while speciation may be facilitatedby repeatedisolation and contact of populations and species, the individualistic nature of species responses implies diffuseinteractionswith less time for coevolutionaryrelationshipsto develop. Furthermore, some microevolutionarychangesthat arise in a stable environmentmay be lost upon widespreadenvironmentalchangeduringmore dynamicphases of glacialinterglacialcycles (Bennett, 1990). Diffuseinteractionsand loss of microevolutionary changes may be common to both temperateand tropical regions, since widespread environmentalchange has been documented for both areas. This last point is particularlyrelevant to current thinking on the evolution of diversity in the tropics. If species responses are as individualistic in the tropics as they are in temperateregions, and species interactionsare at times diffuse,then the hypothesizedevolution of diversitythroughcompetition and niche differentiationin a stable environment becomes questionable as does the importance of long-term coevolutionaryrelationships.Instead, speciation by repeatedgeographicalisolation wouldbe the drivingevolutionaryforce.Van Devander(1986) speculatesthatbecause climatic fluctuationsare less severe towardthe equator,species with refugiaat lower latitudes are less likely to go extinct. Hence they should survive to speciate allopatrically,with intense competition after sympatryleading to niche separationand high diversity in the tropics. Alternatively,the majorityof evolutionarydifferentiation could have takenplace in stableenvironmentsbeforethe onset of the Quaternary glacial-interglacialoscillations.Morepaleoecologicaldataareneededfromthe tropics to test these hypotheses, but we must recognize the limits of the paleoecological recordin this regard.Any searchfor the ghost of competitionpast is likely to produce only circumstantialevidence. Fine resolutionpollen analysismay suggestcompetitive interactions(Bennett, 1986), and paleoecologistshave appliedcompetitionequations to fossil pollen data (Delcourt& Delcourt, 1987). But with the difficultyof defining competition and proving it has happened in extant populations, it is unlikely that most sites provide the taxonomic and spatial resolution to infer mechanisms of competitive interactionsusing pollen data as a proxy for biomass or reproductive success. We can, however, document that taxa occurredin the same locality at the same time, that their abundanceschanged,and that their rangesdiverged,separated, and coalesced. This provides us with circumstantialevidence that competition or speciation by geographicalisolation has had the chance to occur, and with a timeframe within which to place these processes. Given the fluctuatingenvironments of the Quaternary,we can probablyassume that selection pressureand direction on species have been in a state of flux. During a long stable glacialperiod competition for resourcesmay be of primaryimportance in areas where the environment is not extreme. But during a rapid environmental change,ability to adaptquicklyor disperseto more suitablehabitatsmay be favored. This should be true in the tropicsas well as in temperateregions.The variablenature of selection pressure suggests that those species (especially plants) that are most flexible in their response to environmentalchange may have been favored during the Quaternary.Some organismsthat exist today may have been able to rapidlyalter theirlife historystrategiesto adaptto changingenvironmentsand selectionpressures. This would be possible in populationswherealleles for alternatelife historystrategies 230 THE BOTANICALREVIEW exist in a small percentageof the population.However, some strategiessuch as rapid dispersalmight be at least adequateat all times, and other more stress tolerantand competitive strategies(sensu Grime, 1977) might be viable for organismsinhabiting regions where the environment has fluctuatedless rapidly or severely than others. But the recordof environmentaloscillation duringthe Quaternaryindicates that, in general,those species or populations unable to respond rapidly to a changingenvironment should be the ones most likely destined for extinction. The paleoecologicalrecord also demonstratesthe stochasticand historicallyconstrainednatureof evolution and community structure.The structureof some communities can be traced in part back to climatic fluctuationsthat began and ended hundredsof years ago (Clark, 1990; Grimm, 1983) (Fig. 12). Periodic global events with regionallycatastrophiceffects, such as the 1983 El Niino event (as yet poorly understoodon a paleoecologicalscale) may have importanteffectson the evolution of species (Gibbs & Grant, 1987). Species also may migrateinto areasthat constrain their evolutionary future. An example of this is the cloud forest of Amazonian affinitiesthat is isolated in pocketsalong the westernfoothills of the Andes (Koepcke & Koepcke, 1958). Tree species diversity in Europe is much lower than in eastern North America, quite possibly because the east-west trendingAlps trappedspecies retreatingsouthwardfrom advancingglaciersand causedtheir extinction,while small relict southern populations also became extinct (Bennett, 1991; Davis, 1983; Reid, 1935). Slowmigratingand late-arrivingspecies were probablyunable to spread throughoutthe British Isles at the beginning of the most recent interglacialbecause the English Channeland IrishSea were floodedabout 8000 yearsBP by the risingAtlanticOcean (Birks, 1989; Godwin, 1975). Becauseof the complexnatureof past climatechangeand biotic response,historical constraintshave probablycompounded in many cases so that it is highly unlikely that species responses to past glacial-interglacialcycles have been identical to responsesduringthe most recentWisconsinglacial-Holoceneinterglacial.Speciesprobably encounteredvarious competitors, predators,mutualists and pathogensduring differentglacial stages that have influenced their course of evolution. West (1970) presentssome evidence for differentcommunity composition duringeach of the last 7 interglacials(TableII). Thus, on an evolutionarytime scale, we might think of past climaticallydynamicinterglacialsas times when the biotic cardsarereshuffled,setting the stage for glacial periods when species in more stable assemblageshave a better chanceof playingan extendedgame. If Milankovitchcycles do indeed extendbeyond the Quaternary,this scenariomight be characteristicof biotic interactionsthroughout earth history. E. REFUGIA IN THE PALEOECOLOGICALRECORD The individual nature of species as revealed by the paleoecologicalrecord has importantimplications for refugetheory, which has gained more attention with the acceptanceof wide-spreadclimatic changein the tropicsduringthe Pleistocene(Binford & Leyden, 1987; Colinvaux, 1987; Van der Hammen, 1988; Walker& Chen, 1987). The refuge hypothesis states that climatic oscillations have caused species rangesto contractand expand, and has focused on tropicalrainforestspecies whose ranges are thought to have contractedand fragmentedduring the cool, dry glacial periodsand expandedand agglomeratedduringwarm,moist interglacials(Whitmore 231 IMPLICATIONS OF PALEOECOLOGY Table II Occurrencesof generain fossil deposits from seven interglacialstages,includingthe Holocene, in England(modified from West, 1970) stage Interglacial Genus Quercus Ulmus Tilia Carpinus Picea Abies Tsuga Ludhamian Antian Pastonian Cromerian + + + + + + - - - + + + + + + + + + + + + - - - + + + IpHoxnian swichian Holocene + + + + + + + + + + + + + & Prance, 1987). Thus forest communities would have been constantly changing, and the opportunityfor speciation by geographicisolation would have arisen many times duringthe Pleistocene.Various areashave been identifiedas probablerefugia in the Amazon, based on present diversity, endemism, contact zones and climatic conditions (Prance, 1982). This approachis probablytoo simplistic in light of what we know of long-term forest dynamics in temperate zones and the complexity of climatic change. Global climate simulations of full glacial climate suggest, for example, that much of the tropics was cooler, but that rainfallwas similar or slightly less than at present,with decreasedseasonalityand less intense summerrains in the southerntropics (COHMAP, 1988). These climate simulationsdo not disprove the refugehypothesis, but neither do they presentcompelling evidence for locating specific refugialareas in the tropics. The paleoecologicalevidencefor rainforestcontractionand expansionin the tropics is scanty. Walker and Chen (1987) compiled a list of 118 sites that have been investigatedby pollen analysis in the tropics (Table III). Only 24 of these sites are from lowlandeverwetrainforests,and of these sites, 13 contain less desirablerecords fromswampsand riverinelakes.None of these containevidence of full-glacialrefugia. In the Amazon, for example, there is no complete palynologicalrecord back to full-glacialtimes for lowland rainforest.However, Liu and Colinvaux (1985) producedradiocarbon-datedevidencefor a 700-meterloweringofthe lowlandrainforestlower montane rainforestboundarybetween 33,000 and 26,000 years BP. Absy and Van der Hammen's (1976) pollen diagramfor Rond6nia shows that land currently supportingseasonal rainforestwas occupied by savanna sometime in the early Holocene. Hence, the palynologicalevidence for lowlandrainforestrefugiain Amazonia is circumstantial,suggestingonly that refugia probably occurred. Paleoecological evidence does suggest,however,intermediatedisturbanceprocessessuch as flooding, erosion,and fireas possiblemechanismsforgeneratingdiversity(Campbell& Frailey, 1984; Colinvaux et al., 1985; Salo et al., 1986; Sanfordet al., 1985; Saldarriaga& West, 1986). Elsewhere,some lowland rainforestin Africa occupies what was savanna or dry woodland only 10,000 years BP (Livingstone, 1975), and an apparentPleistocene refugehas been documentedfor montane rainforestin east Africa (Hamilton, 1982). In northeasternAustralia, savanna woodland changed to seasonal rainforestfrom 10,000 to 5000 years BP (Chen, 1986; Kershaw, 1983). Clearevidence exists for the 232 THE BOTANICALREVIEW Table III Numberof radiocarbondatedpollen recordsfor non-aridenvironmentsin the tropics (modified from Walker& Chen, 1987) Lowlandeverwet rainforest Lowlandeverwet swamps and riverinelakes Lowlandseasonal forest Otherlow to mid altitude forestsand woodlands Savanna,shruband grassland,some secondaryforest Lowermontane rainforest Upper montane and subalpineforest Subalpineand alpine nonforest Total 11 13 7 7 6 28 17 29 118 loweringof forest zones in the upperAmazon, centralAmerica,tropicalAfrica and southeastAsia duringglacialtimes (Bushand Colinvaux, 1990; Flenley, 1979, 1984; Hamilton, 1982; Liu and Colinvaux, 1985). The tropics,especially,requireextensivepalynologicalandgeologicalinvestigation, such as the ongoingwork of Colinvaux(1991) and co-workersin the Amazon basin, before any hypothesis of refugia composition, size, location, and history can be confidentlyaccepted (Campbellet al., 1985; Colinvaux, 1987; Connor, 1986). Examples of testing the locations of proposed refugiaare provided by Campbellet al. (1985) who found one proposed refugium overlain by Holocene alluvial deposits, and by Bush and Colinvaux (1990) who producedpalynologicalevidence suggesting climatic cooling inconsistent with proposed refugialareas in Panama. In summary,no tropicalrainforestrefugiahave been documented,and some postulatedrefugialareashave been disproven.Manypollen studieshave found,however, that non-rainforestvegetationwas more widespreadduringthe late-glacial,implying that rainforestrangeswere restricted,perhapsto the point of functioningas refugia. Althoughmost interestin the refugehypothesishas been in connectionwith tropical rainforestspecies, refugia (or at least sites where sparse populations occurred)are best documented by paleoecologicalevidence in temperateregions. The proposed Mixed Mesophytic Forest refugium,believed by Braun (1950) and others to have included most tree species presentlygrowingin eastern North America, has never been documented in the paleoecologicalrecord. Instead it appears that taxa had various refugiaindependentof each other (Davis, 1983; Delcourt& Delcourt, 1987). Refugia may have occurredin some surprisingplaces. For example, white pine and hemlock appearto have had a refugiumlocated partiallyon the continentalshelf of the Carolinaswhich was above sea level duringglacial times (Davis, 1983). Many hardwoodsmay have occurredalong the bluffs of the southernMississippi Valley, and in isolated pockets of suitable habitat in the southernAppalachians(Delcourt & Delcourt, 1977; Delcourt et al., 1983). The higherdiversityof tree species in easternNorth Americacomparedto Europe generally has been attributed to tree species being able to migrate independently down the Appalachianmountain chain and find appropriaterefugiathroughoutthe south. Europeantrees on the other hand, would have had to migratesouthwardover or aroundthe Alps duringclimatic deterioration(Campbell,1982; Davis, 1983). The difficultyof reachingproposedrefugianear the Mediterraneanand Asia Minor may have resultedin a significantloss of species. Bennettet al. (1991), however, maintain that species repeatedlybecame extinct in their northern ranges upon initiation of IMPLICATIONS OF PALEOECOLOGY 233 glacial stages, while relict populationssurvived in the south. Thus the crucialfactor in the elimination of a species would be the extinction of its relict populationsin the Balkansand southernEurope.There is evidence that the biogeographicbarnierpresented by high mountain ranges can be breached (Davis et al., 1988), but rapid climate change may precludethis for some species. Refuge theory should take into account the dynamic nature of species behavior and the complexity and interdependenceof past climates, edaphic factors and disturbanceregimes.It also shouldrecognizethatrefugiafor variousspeciesareprobably constantlyformingand expandingand contracting:species that are limited to refugia today (e.g., plant and animal species isolated in mountainrangesand surroundedby desert in the Great Basin, Fraserfir in the centralAppalachianmountains)may be widespreadduringglacialstagesand vice versa.Finally,given the independentnature of species' migrationand life history traits (fecundity,wind vs. animal pollination, seed dispersalmechanisms, environmentaltolerances,etc.) the concept of a 'refuge' may not be appropriatefor many species. The response of species to unfavorable conditions probablyspans a continuum from migrationtowards refugiahaving no overlapwith the formerrange,to rangecontractions,to survivalof sparsepopulations in suitable microhabitatsthroughoutthe former range. Many other scenarios are possible and indeed can be observed in contemporarybiota. V. Prescriptionsand Applications The greatestadvantageof a paleoecologicalperspectiveis that it extendsknowledge of spatial patterns into the very useful fourth dimension of time. Patterns can be observedover long periodsof time and processescan be inferredfrom these patterns. These patterns and inferred processes can be compared with currentlyoccurring phenomena to interpretand perhapspredict environmentaland biotic change. Most ecologistsare awareof the geologyand recenthistoryof their study sites, but few take full advantage of the available paleoecologicaldata to better understand disturbanceregimes and other historical constraints that may operate at several spatialand temporalscales (Hamburg& Sanford,1986). For example,Hamburgand Cogbill (1988) present long-termdata that suggestclimatic warming(in addition to recent pollution) as the causal factor for red sprucedecline in New Hampshire. Paleoecological data also can provide tests for hypotheses that infer temporal processesfrom spatialpatterns.Jacksonet al. (1988) cored a dune pond on the shore of Lake Michigan and showed that Cowles' (1899, 1901) classical hydroserewas a product of human disturbancecommencing 150 years ago. Campbell et al. (1985) demonstratedthat the diverse biota of a proposedrefugiumin the westernAmazon must have invaded the site after several meters of Holocene-agealluvium were laid down. The comparisonof paleoecologicaland currentdatanot only allows the community ecologist and evolutionarybiologist to place currentphenomenainto the context of a longertime-frame,it also can be helpfulin the areaof appliedecology.For example, globalclimate circulationmodels that relateorbitalgeometryof the earthand climate can be tested with paleoecologicaldata to refine the models and help predict future climate and biota (Davis, 1990; Davis & Zabinski, 1991). We also know from geomorphologicaldata, and from increasesin paludificationand retrogressionof certain forested areas that soils have become increasingly leached and nutrient poor in northerntemperateareasduringthe latterhalf of the Holocene (Engstrom& Hansen, 234 THE BOTANICAL REVIEW 1985; Godwin, 1975; Iversen, 1964). Long-termecosystem acidificationdriven by climate and vegetation change during interglacialperiods has been shown to occur especiallyundercertainconditions (Ford, 1990). Vegetationallymediatedsoil change is an ongoing process and should have important implications for foresters and agronomists(Birks, 1986). On a shortertime scale, tree-ringchronologiesand historical recordsmay be helpful in characterizingcurrentclimate in relationto recent climatic changes,and in determiningwhat changesconstitutea long-termtrend(such as globalwarming)vs. a recentcycle (such as the 20-yearprecipitationpatternacross much of SaharanAfrica) vs. random short-termvariation (a two-year drought in parts of North America). Informationon species responsesto past environmentalchangecan help us predict futurebehavior of species. One area of much concernis the possible warmingof the earth'satmosphereowing to increasedemissions from the burningof fossil fuels. In theory, it should be possible to suggestfuture scenariosof community change and species migration by integratingpast vegetation changes with a global circulation model that includes anthropogenicallygeneratedglobalwarming(Davis & Zabinski, 1991). In practicethis may not be so easy. First, the predictedclimatic changeswill take place over a time span of decades to a few hundredyears instead of hundreds to thousandsof years.Second,the predictedglobalwarmingwill be causedby changes in atmosphericchemistry affectingradiationtransmittance,reflectanceand absorption ratherthan by orbitallyinduced changesas in the past. Third, the effects of an alteredglobal climate will be confoundedby the physiologicaleffectsof greenhouse gases, such as CO2,on plants. Fourth, the earth's predictedfuture orbital geometry indicates global cooling, a trend reflectedin global climate and vegetation over the last 2000 years up to the 19th century.Furthermore,generalcirculationmodels and paleoecologicalevidence suggest that disturbanceregimes will likely change with changingclimate and vegetation (Grimm, 1983; Rind et al., 1989). Predictions of futuregreenhouseeffectson the earth'sbiota must take into accountall these factors, and recognize that future climate may not simply be warmer or drier but entirely differentacross a continuum of spatial scales. In light of the difficultiesof predictingclimate change, what generalizationscan we make about biotic responses?First, the type and rate of response will vary taxonomically,even phenotypically.Second, responsewill vary locally with the severity of climatic change. Third, where climatic change is severe response rates will be at or near their maximum. Fourth,even at a maximum responserate, the earth'sbiota will likely lag behind climate change. Fifth, in generalmost species will move polewardor upwardin altitude.Sixth, populationsat polewardextremitiesof landmasses and near mountaintopswill be the most likely to become extinct. Seventh, species that are more phenotypicallyand genotypicallyflexible in their responses will be favored over less flexible ones. Eighth, as species migrateat differentrates, assemblages of species will change. The prediction of species responses to impending environmentalchanges is particularlyimportantwith regardto wildernesspreservesand refugesthat protect endangeredspecies (Graham, 1988; Hunter et al., 1988). Rapid migration of species in response to environmental change and the variable nature of past Quaternary refugiasuggestthat planning refugesis a more complicatedtask than most conservationists realize. Communities as we know and classify them will add and lose species, perhapsproducingassemblagesthat have never occurredin the earth'shistory. Simply markingoff an area may not be enough. Instead, nature preservation IMPLICATIONSOF PALEOECOLOGY 235 will have to take on a fourth, temporal dimension. Hunter et al. (1988) emphasize continental scale migration corridorsand large reserves encompassinga variety of physicalenvironments.This contrastswith the presentstrategyof preservingspecific communities or ecosystems. One practicalapplicationof paleoecologicaltechniques that should be employed more widely is the documentationof an area'shistory before a decision is made to preserveit in its assumed naturalstate. Backmanand Patterson(1984) found that a scrub oak-pitch pine forest in southeasternMassachusetts,which was managedto maintainthat uniquehabitat,had been a white pine-hardwoodsforestin pre-colonial times, and was likelyalteredby land-clearingassociatedwith settlementand extensive fire. Such informationmay not deterthe effortto preservean area,but might greatly alter the managementprocedures. Fire histories can be reconstructedto help the forest managerunderstandnatural firecycles and plan accordingly(Wright,1974). A largebody of such literatureexists for manyregionsin NorthAmerica(e.g.,Harmon, 1983;Heinselman, 1973;Morrison & Swanson, 1990; Romme, 1982), and has been employed in the implementation of National Parkmanagementstrategies(e.g., Hemstrom & Franklin,1982; Kilgore, 1973; Kilgore& Taylor, 1979; Romme, 1982). Anotherapplied use of paleoecologicaldata is as baseline informationwith which to compare environmentalchangescaused by humans. One may want to know, for example, the various characteristicsof the previous natural community before attemptingto restorea naturalecosystem(Engstrom& Wright,1984). Past abundances of organismssensitive to acidity, toxic compounds and eutrophicationcan give us an idea of naturalconditions in presentlypolluted lakes (Battarbee,1984; Battarbee et al., 1989; Davis, 1987). Brugam(1978) documentedpast limnologicalconditions in LinsleyPond usingpollen, diatoms, otheralgae,chironomids,cladocerans,rotifers, and chemical analysis of the sediments. By studyingdiatom assemblagesburied in lake muds, Stocknerand Benson (1967) detected changesin eutrophicationin Lake Washingtonassociated with the development of and sewage input from the city of Seattle. After sewage was diverted from the lake, Shapiro et al. (1971) recorded improvementsin waterquality by comparingthe chemistryof pre- and post-sewage sediment cores. Davis (1987) describesan extensive effort to document changes in lake acidificationby analyzingpast diatom populations from a network of lakes in the northeasternUnited States. Paleolimnologicalstudiesin CentralAmericaand the Caribbeansuggestthat widespreadenvironmentaldestructionin some over-exploitedareas occurredin Mayan times (Binford & Leyden, 1987). Contemporarypatterns similar to those in early Mayan times should help to alert us to areas prone to incipient environmental destruction.In summarypaleoecologicaldata can help answer the question, "what is a 'natural'ecosystem?"in addition to such questions as, "how did the ecosystem arise, what is its history, and which processesare implied by past patterns?" VI. Summary Most ecologistsstudyorganismson a scaleof less than 100 years,but many patterns in naturecan be better understoodon a scale of hundredsto thousands of years or more. The paleoecologicalrecordcan help contemporaryecologistsput currentphenomena into the context of a longer time-frame,allowing us to see processestaking longer than a lifetime, and to gaugethe importanceof past cyclic or unusualevents, disturbanceregimes,andhistoricallyconstrainedphenomena.The Holocenehas been 236 THE BOTANICAL REVIEW a time of rapid environmental change following a more stable glacial. The biotic responseto this changehas been complex and probablydifferentin detail in each of the precedingglacial-interglacialcycles. Species react in an individualistic manner to long-term environmental change; when considered on a paleoecological scale, communities are transient assemblagesof species. Species populations and ranges have respondedto and sometimes laggedbehind environmentalchangesin the Quaternary;these populations and rangescan vary between states of stasis and equilibrium vs. instability and disequilibrium.Paleoecologicalstudies indicate that disturbances of various sizes and intensities play an important role in structuring communities. The environmenthas been highly cyclical duringthe Quaternary,and perhaps throughoutearth history. Such cyclicity should: 1) select for flexibility in species responses to environmental changes and biotic interactions;2) allow for allopatricspeciationby geographicisolation;3) cause some loss of microevolutionary change as biotic assemblages are reorganized.Paleoecology can be helpful in explaining present processes in light of past patterns,making comparisons,and predicting futurepatterns. VII. Acknowledgments We thankD. Gaudreau,H. D. Hammond, A. Knoll, D. Perlman,T. Sipe, V. Viana and G. Whitney for critically reading the manuscript,B. Flye for assistance with typing, and M. Fluet for assistancewith the figures.Schoonmakerwas supportedby fellowships from the National Science Foundation and the R. T. Fisher Fund of Harvard University. This work is a contribution from the Long Term Ecological ResearchProgramat the HarvardForest (National Science FoundationGrant BSR 8811764). VIII. LiteratureCited Aaby, B. 1983. Forestdevelopment,soil genesis and humanactivityillustratedby pollen and hypha analysisof two neighboringpodzols in Draved Forest, Denmark.Danm. geol. Unders. Ser. II, 114, 114 pp. Absy,M. L. &T. vanderHammen. 1976. Some paleoecologicaldatafromRondonia,southern part of the Amazon basin. Acta Amazonica6: 293-299. Allison, T. D., R. E. Moeller & M. B. Davis. 1986. Pollen in laminatedsedimentsprovides evidence for a mid-Holoceneforest pathogenoutbreak.Ecology67: 1101-1105. Andersen,S. Th. 1970. The relative pollen productivityand pollen representationof north Europeantrees, and correctionfactorsfor tree pollen spectra.Danm. geol. Unders. Ser. II, 96, 99 p. 1974a. Wind conditions and pollen deposition in a mixed deciduousforest. I. Wind conditions and pollen dispersal.Grana 14: 57-63. 1974b. Wind conditions and pollen deposition in a mixed deciduousforest. II. Seasonal and annualpollen deposition 1967-1972. Grana 14: 64-77. 1984. Forests at Lovenholm, Djursland,Denmark,at presentand in the past. Biol. Skr. 24: 1. 208 pp. Backman,A. E. & W. A. PattersonIII. 1984. Vegetationand fire history of Miles Standish State Forest interpretedfrom sedimentarypollen and charcoal analyses. University of MassachusettsCooperativeForestryResearchUnit, ReportNo. 2. 21 pp. Barbour,M. G. & W. D. Billings (eds.). 1988. North Americanterrestrialvegetation.CambridgeUniversity Press, Cambridge. Bartlein,P. J., T. Webb Ill & S. Hostetler. 1989. Climatology.Pages 53-88 in R. L. Hunter IMPLICATIONSOF PALEOECOLOGY 237 & C. J. Mann (eds.), Techniques for determiningprobabilitiesof events and processes affectingthe performanceof geologicdepositories-Literaturereview.U.S. NuclearRegulatory Commission, Washington,D.C. Battarbee,R. W. 1984. Diatom analysisand the acidificationof lakes. Philos. Trans.,Ser. B 305: 451477. A. C. Stevenson,B. Rippey,C. Fletcher,J. Natkamski,M. Wik & R. J. Flower. 1989. Causesof lake acidificationin Galloway,south-westScotland;a paleoecologicalevaluation of the relativeroles of atmosphericcontaminationand catchmentchangefor two acidified sites with non-afforestedcatchments.J. Ecol. 77: 651-672. Bennett, K. D. 1986. Competitive interactionsamong forest tree populations in Norfolk, England,duringthe last 10,000 years. New Phytol. 103: 603-620. 1990. Milankovitchcycles and their effectson speciesin ecologicaland evolutionary time. Paleobiology16: 11-21. 1988. Post-glacialvegetationhistory:Ecologicalconsiderations.Pages699-724 in B. Huntley & T. Webb III (eds.), VegetationHistory. Kluwer,The Hague. 9P. C. Tzedakis & K. J. Willis. 1991. Quatemaryrefugiaof north Europeantrees. J. Biogeogr.18: 103-115. Berger, A., J. Imbrie, J. Hayes, G. Kukla & B. Saltzman (eds.). 1986. Milankovitchand climate. Reidel, Dordrecht. Berglund,B. E. (ed.). 1986. Handbookof Holocene paleoecologyand paleohydrology.John Wiley & Sons, New York. Bernabo,J. C. & T. Webb III. 1977. Changingpatternsin the Holocene pollen record in northeasternNorth America:A mapped summary.Quat. Res. 8: 64-96. Betancourt,J. L., T. R. Van Devander& P. S. Martin (eds.). 1990. Packratmiddens:The last 40,000 years of biotic change.University of Arizona Press, Tucson. Binford, M. W. & B. Leyden. 1987. Ecosystems, paleoecologyand human disturbancein subtropicaland tropicalAmerica. QuaternarySci. Rev. 6: 115-128. Birks,H. J. B. 1986. Late-Quaternarybiotic changesin terrestrialand lacustrineenvironments with particularreferenceto nort-westEurope.Pages3-65 in B. E. Berglund(ed.),Handbook of Holocene paleoecologyand paleohydrology.John Wiley & Sons, New York. 1989. Holocene isochronemaps and patternsof tree-spreadingin the BritishIsles. J. Biogeogr.16: 503-540. & H. H. Birks. 1980. Quaternarypaleoecology.EdwardArnold, London. Bradley,R. S. 1985. Quaternarypaleoclimatology.Allen and Unwin, Boston. Bradshaw,R. H. W. 1981. Quantitativereconstructionof local woodland vegetationusing pollen analysis from a small basin in Norfolk, England.J. Ecol. 69: 941-955. 1988. Spatially-precisestudies of forest dynamics. Pages 725-751 in B. Huntley & T. Webb III (eds.), Vegetationhistory.Kluwer,The Hague. & N. G. Miller. 1987. Recent successionalprocessesinvestigatedby pollen analysis of closed-canopyforest sites. Vegetatio76: 45-54. Braun,E. L. 1950. Deciduousforestsof easternNorth America.HafnerPubl. Co., New York. Brokaw,N. L. V. 1985. Gap phase regenerationin a tropicalforest. Ecology66: 682-687. Brubaker,L. B. 1975. Postglacialforest patternsassociatedwith till and outwash in northcentralupperMichigan.Quat. Res. 5: 499-527. Brugam,R. B. 1978. Pollen indicatorsof land-usechangein southernConnecticut.Quat.Res. 9: 349-362. Bush, M. B. & P. A. Colinvaux. 1990. A pollen recordof a completeglacialcycle fromlowland Panama.J. Veg. Sci. 1: 105-118. Cain, S. A. 1947. Characteristicsof naturalareas and factors in their development. Ecol. Monogr.17: 185-200. Calcote,R. R. & M. B. Davis. 1989. Comparisonof pollen in surfacesamplesof foresthollows with surroundingforests. Bull. Ecol. Soc. Amer. 704(2): 75. Campbell,J. J. N. 1982. Pearsand persimmons:A comparisonof temperateforestsof Europe and easternNorth America. Vegetatio49: 85-101. 238 THE BOTANICALREVIEW Campbell,K.E., Jr., & D. Frailey. 1984. Holocene flooding and species diversity in southwesternAmazonia. Quat. Res. 21: 369-375. , C. D. Frailey & J. Arellano. 1985. The geologyof the Rio Beni:Furtherevidence for Holocenefloodingin Amazonia.Contr.in Sci., No. 364, pp. 1-18. NaturalHistoryMuseum of Los Angeles County. Chen, Y. 1986. EarlyHolocene vegetationdynamicof LakeBarrinebasin, northeastQueensland, Australia.Ph.D. Thesis. AustralianNational University, Canberra. Clark,J. S. 1988a. Particlemotion and the theoryof charcoalanalysis:Sourcearea,transport, deposition, and sampling.Quat. Res. 30: 67-80. 1988b. Stratigraphiccharcoalanalysis on petrographicthin sections:Applicationto fire history in northwesternMinnesota.Quat. Res. 30: 81-91. 1990. Fireandclimateduringthe last 750 yrin northwesternMinnesota.Ecol.Monogr. 60: 135-159. Clements,F. E. 1910. The life historyof lodgepolepine bum forests.U.S. ForestServiceBull. 79. 1916. Plant succession.An analysisof the developmentof vegetation.CarnegieInst. WashingtonPubl. 242. Washington,D.C. 1924. Methods and principles of paleo-ecology.Yearbook 32. Washington,D.C.: CarnegieInstitutionof Washington. COHMAP Members. 1988. Climatechangesof the last 18,000 years:Observatonsand model simulations.Science 241: 1043-1052. Cole, K. 1985. Past rates of change, species richness,and a model of vegetationalinertia in the GrandCanyon,Arizona.Amer. Nat. 125: 289-303. Colinvaux,P. 1987. Amazon diversityin light of the paleoecologicalrecord.Quat. Sci. Rev. 6: 93-114. 1991. Song of the Livingstonein Amazonia. Bull. Ecol. Soc. Amer. 72(1): 14-15. M. C. Miller, K-b.Liu, M. Steinitz-Kannan& I. Frost. 1985. Discoveryof permanent Amazon lakes and hydraulicdisturbancein the upperAmazon Basin. Nature313: 42-45. Connell,J. H. 1978. Diversityin tropicalrainforestsand coralreefs.Science199: 1302-1310. Connor,J. H. 1986. The role of Pleistoceneforest refugiain the evolution and biogeography of tropicalbiotas. Trends in Ecol. and Evol. 1(6): 165-168. Cowles, H. C. 1899. The ecological relations of the vegetation on the sand dunes of Lake Michigan.Bot. Gaz. 27: 95-117, 167-202, 281-308, 361-391. 1901. The physiographicecologyof Chicagoand vicinity. Bot. Gaz.31: 73-108,145182. Cox, A., R. R. Doell & G. R. Dalrymple. 1963. Geomagneticpolarityepochs and Pleistocene geochronometry.Nature 198: 1049-1051. Critchfield,W. B. 1985. The late Quaternaryhistory of lodgepoleand jack pines. Canad.J. For. Res. 15: 749-772. Curtis,J. T. 1955. A prairiecontinuumin Wisconsin.Ecology36: 558-566. 1959. Vegetationof Wisconsin:An ordinationof plant communities.University of Wisconsin,Madison. & R. P. McIntosh. 1951. An upland forest continuum in the prairie-forestborder regionof Wisconsin. Ecology32: 476-496. Cushing,E. J. 1967. Evidencefordifferentialpollenpreservationin late-Quaternary sediments in Minnesota.Rev. Paleobot. Palynol. 4: 87-10 1. Cwynar,L. C. 1987. Fire and the foresthistoryof the northCascadeRange.Ecology68: 791802. & G. M. MacDonald. 1987. Geographicalvariation of lodgepole pine in relationto populationhistory. Amer. Nat. 129: 463-469. Davidson,J. L. 1983. Paleoecologicalanalysis of Holocene vegetation,Lake in the Woods, Cades Cove, Great Smokey MountainsNational Park. M.S. Thesis. University of Tennessee, Knoxville. Davis, M. B. 1973. Redepositionof pollen grainsin lake sediment.Limnol.Ocean.18: 44-52. IMPLICATIONSOF PALEOECOLOGY 239 * 1981a. Quaternaryhistoryand the stabilityofforest communities.Pages 132-153 in D. C. West, H. H. Shugart& D. B. Botkin (eds.), Forest succession.Springer-Verlag, New York. 198lb. Outbreaksofforest pathogensin Quaternaryhistory.Proceedingsofthe Fourth InternationalPalynologicalConference,Lucknow3: 216-227. 1983. Quaternaryhistoryof deciduousforestsof easternNorth Americaand Europe. Ann. MissouriBot. Gard. 70: 550-563. 1986. Climaticinstability,time lags, and communitydisequilibrium.Pages269-284 in J. Diamond & T. J. Case (eds.), Communityecology. Harper& Row Publ., New York. 1990. Insightsfrom paleoecologyon globalchange.Bull. Ecol. Soc. Amer. 70(4): 222228. & D. B. Botkin. 1985. Sensitivityof cool temperateforestsand their pollen recordto rapidtemperaturechange.Quat. Res. 23: 327-340. -~ & L. B. Brubaker. 1973. Differentialsedimentationof pollen grainsin lake sediment. Limnol. Ocean. 18: 635-646. & C. Zabinski. 1991. Changesin geographicalrangeresultingfrom greenhousewarming-effectson biodiversityin forests.In R. L. Peters& T. Lovejoy(eds.), Consequencesof C02-inducedclimatic warmingon biodiversity.Yale University Press,New Haven, Connecticut.In press. , K. D. Woods,S. L. Webb& R. P. Futyma. 1986. Dispersalversusclimate:Expansion of Fagus and Tsuga into the Upper Great Lakesregion.Vegetatio67: 93-103. Davis, O. K., R. S. Anderson& K. L. Cole. 1988. Westwarddisplacementof the SierraNevada biogeographicboundaryduringthe last glacialtermination.Bull. Ecol. Soc. Amer. 69(2): 115. Davis, R. B. 1987. Paleolimnologicaldiatom studiesof acidificationof lakes by acid rain:An applicationof Quaternaryscience. Quat. Sci. Rev. 6: 147-164. & G. L. Jacobson,Jr. 1985. Late-glacialand early Holocene landscapesin northern New Englandand adjacentareasof Canada.Quat. Res. 23: 341-368. Day, G. M. 1953. The Indianas an ecologicalfactorin the northeastforest.Ecology34: 329346. Delcourt,H. R. & P. A. Delcourt. 1985. Quaternarypalynologyand vegetationhistoryof the southeasternUnited States. Pages 1-37 in V. M. Bryant,Jr. & R. G. Holloway (eds.), Pollenrecordsof Late-Quaternary North Americansediments.Amer.Assoc. Strat.Palynol. Foundation,Dallas. & E. C. Spiker. 1983. A 12,000 year record of forest history from Cahaba 1) Pond, St. ClairCounty,Alabama.Ecology64: 874-887. & T. Webb III. 1983. Dynamic plant ecology: The spectrumof vegetation 1) changein space and time. Quat. Sci. Rev. 1: 153-175. Delcourt,P. A. & H. R. Delcourt. 1977. The TunicaHills, Louisiana-Mississippi:Lateglacial locality for spruceand deciduousforest species. Quat. Res. 7: 218-237. & . 1981. Vegetationmaps for easternNorth America:40,000 yr BP to the present.GeobotanyII (R. C. Romans, ed.). Plenum Publishing,New York. & . 1987. Long-termforest dynamics of the temperatezone. Springer-Verlag, New York. Denslow,J. S. 1980. Gap partitioningamong tropicalrainforesttrees. Biotropica12(Suppl.): 47-55. Emiliani, C. 1972. Quaternaryhypsithermals.Quat. Res. 2: 270-273. Engstrom,D. R. &B.C. S. Hansen. 1985. Postglacialvegetationalchangeandsoil development in southeasternLabradoras inferredfrom pollen and chemicalstratigraphy.Canad.J. Bot. 63: 543-561. & H. E. Wright. 1984. Chemicalstratigraphyof lake sediments as a recordof environmentalhistory. Pages 11-67 in E. Y. Haward & W. G. Lund (eds.), Lake sediments and environmentalhistory. University of MinnesotaPress, Minneapolis. Flenley,J. R. 1979. The EquatorialRain Forest:A geologicalhistory.Butterworths,London. 240 THE BOTANICALREVIEW * 1984. LateQuaternarychangesof vegetationand climatein the Malesianmountains. Erdwiss.Forsch. 18: 261-267. Ford, M. S. 1990. A 10,000-yrhistory of naturalecosystem acidification.Ecol. Monogr.60: 57-89. Foster, D. R. 1988. Disturbancehistory, community organizationand vegetationdynamics of the old-growthPisgah Forest, southwesternNew Hampshire,U.S.A. J. Ecol. 76: 105134. ,P. K. Schoonmaker& S. T. A. Pickett. 1990. Insightsfrompaleoecologyto community ecology. Trendsin Ecol. and Evol. 5: 119-122. Franklin,J. F., H. H. Shugart& M. E. Harmon. 1987. Tree death as an ecologicalprocess. BioScience37: 550-556. Frelich, L. E., M. B. Davis & S. R. Hill. 1989. Persistenceof hardwood stands in Upper Michigan.Bull. Ecol. Soc. Amer. 70(2): 115. Fritts, H. C. 1976. Tree ringsand climate. Academic Press, New York. Gaudreau,D. C. 1984. Mapped pollen history of northeasternNorth America for the past 14,000 years. Geol. Soc. America, Northeast Section, 19th Annual Meeting Abstracts 16: 42. 1986. Late-Quaternaryhistory of the Northeast:Paleoecologicalimplicationsof topographicpatternsin pollen distributions.Ph.D. Thesis. Yale University, New Haven, Connecticut. & T. Webb III. 1985. Late-Quaternarypollen stratigraphyand isochrone maps for the northeasternUnited States. Pages 247-280 in V. M. Bryant& R. G. Holloway (eds.), Pollenrecordsof Late-Quaternary North Americansediments.AmericanAssociationStrat. Palynol. Foundation,Dallas. 1988. Paleoecologicalinterpretationof geographicpatternsin pollen data:Spruceand birch in northeasternNorth America. Pages 13-29 in R. S. Lamb, N. G. Miller & D. W. Steadman(eds.), Late Pleistoceneand EarlyHolocene paleoecologyand archeologyof the EasternGreat Lakesregion. Bull. BuffaloSoc. Nat. Sci., Vol. 33. , S. T. Jackson & T. Webb III. 1989. Spatial scale and sampling strategyin paleoecologicalstudiesof vegetationpatternsin mountainousterrain.Acta Bot. Neerl. 38: 369390. Gibbs,H. L. & P. R. Grant. 1987. Ecologicalconsequencesof an exceptionallystrongEl Niiio event on Darwin'sFinches. Ecology68: 1735-1746. Gleason, H. A. 1917. The structureand development of the plant association. Bull. Torrey Bot. Club. 44: 463-481. 1926. The individualisticconcept of the plant association.Bull. TorreyBot. Club53: 7-26. Godwin,H. 1975. The history of the Britishflora.CambridgeUniversity Press, Cambridge. 1980. The archivesof the peat bogs. CambridgeUniversity Press, Cambridge. Gordon,A. D. & H. J. B. Birks. 1985. Numericalmethodsin paleoecology.John Wiley, New York. Gorham,E. 1957. Development of peatlands.Quart.Rev. Biol. 32: 145-166. Gould,S. J. 1985. The paradoxofthe firsttier:An agendafor paleobiology.Paleobiology11: 2-12. Graham,R. W. 1986. Responseof mammaliancommunitiesto environmentalchangesduring the late Quaternary.Pages 300-313 in J. Diamond and T. J. Case (eds.), Community ecology. Harper& Row Publ., New York. 1988. The role of climate changein the design of biologicalreserves:The paleoecological perspectivefor conservationbiology. ConservationBiol. 2: 391-394. Green, D. G. 1981. Time series and post-glacialforest ecology. Quat. Res. 15: 265-277. 1982. Fire and stabilityin the post-glacialforests of southwestNova Scotia. J. Biogeogr.9: 29-40. 1988. Fine resolutionpollen analysis.J. Biogeogr.15: 685-701. IMPLICATIONS OF PALEOECOLOGY 241 Grime, J. P. 1977. Evidence for the existence of three primarystrategiesin plants and its relevanceto ecologicaland evolutionarytheory. Amer. Nat. 111: 1169-1194. Grimm,E. C. 1983. Chronologyand dynamicsof vegetationchangein the prairie-woodland region of southernMinnesota,U.S.A. New Phytol. 93: 311-335. Hamburg,S. P. & C. V. Cogbill. 1988. Historical decline of red spruce populations and climatic warming.Nature 331: 428-431. & R. L. Sanford,Jr. 1986. Disturbance,Homo sapiens, and ecology. Bull. Ecol. Soc. Amer. 67: 169-171. Hamilton,A. C. 1982. Environmentalhistory of East Africa.Academic Press, New York. Harmon, M. 1983. Fire history of the westernmostportion of Great Smokey Mountains National Park. Bull. TorreyBot. Club 109: 74-79. Habeck,J. R. & R. W. Mutch. 1973. Firedependentforestsin the northernRockyMountains. Quat. Res. 3: 408-424. Havinga,A. J. 1967. Palynologyand pollen preservation.Rev. Paleobot.Palynol. 2: 81-98. Heide, K. 1984. Holocene pollen stratigraphyfrom a lake and small hollow in north-central Wisconsin, USA. Palynology8: 3-20. Heinselman, M. L. 1973. Fire in the virgin forests of the BoundaryWaters Canoe Area, Minnesota.Quat. Res. 3: 329-382. 1981. Fire and succession in the conifer forests of northernNorth America. Pages 374-405 in D. C. West, H. H. Shugart& D. B. Botkin (eds.), Forest succession.SpringerVerlag,New York. Hemstrom,M. A. & J. F. Franklin. 1982. Fire and otherdisturbancesof the forestsin Mount RainierNational Park.Quat. Res. 18: 32-51. Henry,J. D. & J. M. A. Swan. 1974. Reconstructingforest historyfrom live and dead plant material-An approachto the study of forest succession in southwest New Hampshire. Ecology55: 772-783. Hooghiemstra,H. 1984. Vegetationaland climatic history of the High Plains of Bogota, Colombia:A continuousrecordof the last 3.5 million years. DissertationesBotanica79. J. CramerVerlag,Vaduz. Hubbell,S. P. & R. B. Foster. 1986. Biology,chanceand historyand the structureof tropical rain forest tree communities. Pages 314-329 in J. Diamond and T. J. Case (eds.), Community ecology. Harper& Row Publ., New York. Hunter, Jr., M. L., G. L. Jacobson,Jr. & T. Webb III. 1988. Paleoecologyand the coarse filterapproachto maintainingbiologicaldiversity.ConservationBiol. 2: 375-385. Huntley, B. 1990. Dissimilaritymappingbetweenfossil and contemporarypollen spectrain Europefor the past 13,000 years. Quat. Res. 33: 360-376. & H. J. B. Birks. 1983. An atlasof past and presentpollen maps for Europe:0-13,000 years ago. CambridgeUniversity Press, Cambridge. & T. Webb III. 1988. Vegetationhistory. KluwerAcademic Publ., The Hague. & . 1989. Migration:Species'responseto climaticvariationscausedby changes in the earth'sorbit. J. Biogeogr.16: 5-19. Imbrie,J. & K. P. Imbrie. 1986. Ice ages. HarvardUniversity Press, Cambridge,Massachusetts. Iversen,J. 1964. Retrogressivevegetationalsuccessionin the post-glacial.J. Ecol. 52(Suppl.): 59-70. 1973. The developmentof Denmarksnaturesincethe last glacial.Danks.Geol. Under. V. 7-C, 126 pp. Jackson, S. T. 1989. Postglacialvegetationalchanges along an elevational gradientin the AdirondackMountains(New York):A study of plant macrofossils.New York State Museum and Science Service Bull. 465, Albany, New York. 1990. Pollen sourceareaand representationin small lakes of the northeasternUnited States. Rev. Paleobot. Palynol. 63: 53-76. , R. P. Futyma& D. A. Wilcox. 1988. A paleoecologicaltest of a classical hydrosere in the Lake Michigandunes. Ecology69: 928-936. 242 -~ THE BOTANICALREVIEW & D. R. Whitehead. 1991. Holocenevegetationpatternsin the AdirondackMountains. Ecology72: 641-653. Jacobson, G. L. & R. H. W. Bradshaw. 1981. The selection of sites for paleovegetational studies. Quat. Res. 16: 80-96. & E. C. Grimm. 1986. A numericalanalysisof Holocene forestand prairievegetation in centralMinnesota.Ecology67: 958-966. , T. Webb & E. C. Grimm. 1987. Patternsand rates of vegetationchangeduringthe deglaciationof easternNorth America.Pages277-288 in W. F. Ruddiman& H. E. Wright (eds.),The geologyof North America.Vol. K-3. GeologicalSoc. Amer., Boulder,Colorado. Keever,C. 1953. Present composition of some stands of former oak-chestnutforest in the southernBlue Ridge Mountains.Ecology34: 44-55. Kershaw,A. P. 1983. A Holocene pollen diagramfrom Lynch'sCrater,nort-easternQueensland, Australia.New Phytol. 94: 669-682. Kilgore,B. M. 1973. The ecological role of fire in Sierranconifer forests:Its applicationto National Parkmanagement.Quat. Res. 3: 496-513. & D. Taylor. 1979. Fire history of a Sequoia-mixedconifer forest. Ecology60: 129142. King, J. E. 1981. Late Quaternaryhistory of Illinois. Ecol. Monogr.51: 43-62. Koepcke,H. W. & M. Koepcke. 1958. Los restos de bosques en las vertientesoccidentales de los andes peruanos.Bol. Comite Nat. Proteci6nNaturaleza16: 22-30. Lima. Kutzbach,J. E. & P. J. Guetter. 1986. The influence of changingorbital parametersand surfaceboundaryconditions on climate simulationsfor the past 18,000 years. J. Atmos. Sci. 43: 1726-1759. Leighton,M. & N. Wirawan. 1986. Catastrophicdroughtand fire in Borneantropicalrain forestsassociatedwith the 1982-83 ENSO event. Pages 75-102 in G. Prance(ed.), AAAS Symposiumon TropicalRain Forests and the World Atmosphere. Lieberman,D., M. Lieberman,R. Peralta & G. S. Hartshorn. 1985. Mortalitypatternsand stand turnoverrates in a tropicalforest in Costa Rica. J. Ecol. 73: 915-924. Liu, K-B. & P. Colinvaux. 1985. Forest changesin the Amazon Basin duringthe last glacial maximum. Nature 318: 556-557. 1988. A 5200 year history of Amazon rain forest. J. Biogeogr.15: 231-248. Livingston,B. E. & F. Shreve. 1921. The distributionof vegetationin the United States as relatedto climatic conditions. CarnegieInstitutionPubl. 284, Washington,D.C. Livingstone,D. A. 1975. Late Quaternaryclimate changein Africa.Annual Rev. Ecol. Syst. 6: 249-280. Lugo, A. E., M. Applefield,D. J. Pool & R. B. MacDonald. 1983. The impact of Hurricane David on the forests of Dominica. Canad.J. For. Res. 13: 201-211. Mackey, H. E. & N. Sivic. 1973. Presentcomposition of formeroak-chestnutforest in the AlleghenyMountainsof westernPennsylvania.Ecology54: 915-919. Maher, L. J. 1972. Nomograms for computing0.95 confidencelimits of pollen data. Rev. Paleobot. Palynology13: 85-93. McAndrews,J. H. 1965. Postglacialhistory of prairie,savanna, and forest in northwestern Minnesota.Mem. Torrey.Bot. Club 22: 1-72. Mengel, R. M. 1964. The probable history of species formation in some northernwood warblers(Parulidae).Living Bird 3: 9-43. Milankovitch,M. 1941. Kanon der Erdbestrahlungund seine Andwendungauf das Eiszeitenproblem.Royal Serb.Acad. Spec. Publ. 133. Belgrade.Englishtranslationpublishedin 1969 by IsraelProgramfor ScientificTranslationsavailablefrom U.S. Dept. Comm. Minnich,R. A. 1988. The biogeographyof firein the SanBernardinoMountainsof California. Universityof CaliforniaPublicationsin Geography.Vol. 28. Universityof CaliforniaPress, Berkeley,California. Mitchell, F. J. G. 1988. The vegetationalhistory of the KillarneyOakwoods,SW Ireland: Evidence from fine spatial resolutionpollen analysis.J. Ecol. 76: 415-536. IMPLICATIONSOF PALEOECOLOGY 243 Morrison,P. H. & F. J. Swanson. 1990. Firehistoryand patternin a CascadeRangelandscape. USDA Forest Serv. Gen. Tech. Rep. PNW-GTR-254. Nials, F. L., E. E. Deeds, M. E. Moseley, S. G. Pozorski, T. G. Pozorski & R. A. Feldman. 1979. El Ninio:The catastrophicfloodingof coastalPeru.Bull.Field Mus. Nat. Hist. 50(7): 4-14; 50(8): 4-10. Nilsson, T. 1982. The Pleistocene.D. Reidel Publ. Co., Dordrecht,Holland. Ogden,J. 1985. Past, presentand future:Studieson the populationdynamicsof some longlived trees. Pages 3-16 in J. White (ed.), Studies on plant demography:A Festschriftfor John L. Harper.Academic Press, London. Oliver, C. D. & E. P. Stephens. 1978. Reconstructionof a mixed-speciesforest in central New England.Ecology58: 562-572. PattersonIII, W. A. & A. E. Backman. 1988. Fire and disease history of forests.Pages 603632 in B. Huntley & T. Webb III (eds.), Vegetationhistory. Kluwer,The Hague. , W. A., K. J. Edwards& D. Maguire. 1987. Microscopiccharcoalas a fossil indicator of fire. QuaternarySci. Rev. 6: 3-23. , W. A. & K. E. Sassaman. 1988. Indian firesin the prehistoryof New England.Pages 107-135 in G. P. Nichols (ed.), Holocene human ecology in northeasternNorth America. Plenum Publ., New York. Payette, S. & L. Filion. 1985. White spruce expansion at the treeline and climatic change. Canad.J. For. Res. 15: 241-251. Peglar, S. M., S. C. Fritz, T. Alapieti, M. Saarnisto& H. J. B. Birks. 1984. Composition and formationof laminatedsedimentsin Diss Mere,Norfolk, England.Boreas13: 13-28. Pickett, S. T. A. 1988. Space-for-timesubstitutionas an alternativeto long-termecological studies. Pages 110-135 in G. E. Likens (ed.), Long-termstudies in ecology:Approaches and alternatives.Springer-Verlag, New York. & M. B. Davis (organizers). 1990. 75th AnniversarySymposium-The shift from an equilibriumto a nonequilibriumparadigmin ecology. Ecol. Soc. Amer. Bull. 71(2): 32. Prance,G. T. (ed.). 1982. Biologicaldiversificationin the tropics.ColumbiaUniversityPress, New York. Prentice, I. C. 1985. Pollen representation,source area, and basin size: Toward a unified theory of pollen analysis.Quat. Res. 23: 76-86. 1986. Vegetationresponsesto past climatic variation.Vegetatio67: 131-141. 1988. Paleoecologyand plantpopulationdynamics.Trendsin Ecol.and Evol. 3: 343345. & T. Webb III. 1986. Pollen percentages,tree abundancesand the Fagerlindeffect.J. Quat. Sci. 1: 35-43. Pyne, S. J. 1982. Fire in America.PrincetonUniversity Press, Princeton. Ramensky,L. G. 1924. Die Grundgestzmassigkeitten im Aufbauder Vegetationsdecke(Russian). VestnikOpytn.dela Sredne-Chernoz.Obl., Voronezh.(Bot. Centralbl.,N.F. 7: 453455, 1926). Raynor,G. S., J. V. Hayes & E. C. Ogden. 1975. Particulatedispersionfrom sourceswithin a forest. BoundaryLayerMeteorology9: 257-277. Reid, E. M. 1935. Britishflorasantecedentto the GreatIce Age. Proc. Royal Soc. 118: 197202. Ricklefs, R. E. 1987. Community diversity:Relative roles of local and regionalprocesses. Science 235: 167-171. Rind, O., R. Goldberg& R. Reudy. 1989. Changein climatic variabilityin the 21st century. ClimaticChange 13. Ritchie,J. C. 1987. Postglacialvegetationof Canada.CambridgeUniversityPress,New York. Romme,W. H. 1982. Fireandlandscapediversityin subalpineforestsof YellowstoneNational Park.Ecol. Monogr.52: 199-221. Roughgarden,J. &J. Diamond. 1986. Overview:The roleof speciesinteractionsin community 244 THE BOTANICALREVIEW ecology. Pages 337-343 in J. Diamond & T. J. Case (eds.), Communityecology. Harper & Row Publ., New York. Ruddiman,W.F. & H.E. Wright,Jr.(eds.), 1987. North Americaand adjacentoceans during the last deglaciation.DNAG Volume K-3. Geol. Soc. Amer., Boulder,Colorado. Russell, E. W. 1983. Indian-setfiresin the forestsof the northeastemUnited States.Ecology 64: 78-88. Saarnisto,M. 1986. Anmnuallylaminatedlake sediments.Pages 343-370 in B. E. Berglund (ed.), Handbookof Holocene paleoecologyand paleohydrology.John Wiley & Sons, New York. Saldarriaga,J. G. & D. C. West. 1986. Holocene firesin the northem Amazon basin. Quat. Res. 26: 358--366. Salo, J., R. Kalliola, I. Hakkinen, Y. Makinen, P. Niemela, M. Pahukka & P. Coley. 1986. River dynamics and the diversityof Amazon lowland forest. Nature322: 254-258. Sanford,R. L., Jr., J. Saldarriaga,K. E. Clark, C. Uhl & R. Herrera. 1985. Amazon rainforest fires. Science 227: 53-55. Sarna-Wojcicki,A. M., D. E. Champion& J. 0. Davis. 1983. Holocene volcanism in the coterminalUnited Statesand the role of silicic volcanic ash layersin correlationof latestPleistoceneand Holocenedeposits.Pages 52-77 in H. E. Wright,Jr. (ed.), LateQuatemary environmentsof the United States. Vol. 2. University MinnesotaPress, Minneapolis. Schoener,T. W. 1986. Overview:Kinds of ecologicalcommunities-Ecology becomespluralistic. Pages 467-479 in J. Diamond & T. J. Case (eds.), Communityecology. Harper& Row Publ., New York. Schoonmaker,P. K. & D. R. Foster. 1989. Stand-levelforest historyin centralNew England investigatedby pollen analysis.Bull. Ecol. Soc. Amer. 70(2): 256. & . 1990. Long-termforesthistoryfrom pollen in soils and hollows versusstem reconstruction:A comparisonof techniquesand results.Bull. Ecol. Soc. Amer. 71(2): 317. Schwartz,M. W. 1989. Predictingtree frequenciesfrom pollen frequency:An attempt to validate the R value method. New Phytol. 112: 112-143. Shackleton,N. J., J. Imbrie& N. G. Pisias. 1988. The evolution of oceanic oxygen-isotope variabilityin the North Atlantic over the past three million years. Philos. Trans. Ser. B, 318: 679-688. & N. D. Opdyke. 1973. Oxygenisotope and paleomagneticstratigraphyof equatorial Pacificcore V28-238: Oxygenisotope temperaturesand ice volumes on a 105and 106 year scale. Quat. Res. 3:39-55. Shapiro,J., W. T. Edmondson& D. E. Allison. 1971. Chjangesin the chemicalcomposition of Lake Washington,1958-70. Limnol. Ocean. 16: 437-452. Stockner,J. G. & W. W. Benson. 1967. The successionof diatom assemblagesin the recent sediments of Lake Washington.Limnol. & Oceanogr.12: 513-532. Swain, A. M. 1973. A history of fire and vegetationin northeasternMinnesotaas recorded in lake sediments. Quat. Res. 3: 383-396. 1978. Environmentalchangesduringthe past 2000 yearsin north-centralWisconsin: Analysis of pollen, charcoal,and seeds from varved lake sediments.Quat. Res. 10: 55-68. Thompson,R. & F. Oldfield. 1986. Environmentalmagnetism.Allen and Unwin, London. Uhl, C., K. Clark,N. Dezzeo & P. Maquirino. 1988. Vegetationdynamicsin Amazon treefall gaps. Ecology69: 751-763. Urban, D. L., R. V. O'Niell & H. H. Shugart,Jr. 1987. Landscapeecology. BioScience37: 119-127. Van der Hammen,T. 1988. Late-Tertiaryand Pleistoceneenvironmentalhistory-20 My20 Ky: South America. Pages 307-340 in J. Huntley & T. Webb III (eds.), Vegetation history. Kluwer,The Hague. T., T. A. Wijmstra& W. H. Zagwijn. 1971. The floralrecordof the late Cenozoic of Europe.Pages391-424 in K. K. Turekian(ed.), LateCenozoicglacialages.Yale University Press, New Haven. IMPLICATIONSOF PALEOECOLOGY 245 Van Devander,T. R. 1986. Climatic cadences and the composition of Chihuahuandesert communities:The late Pleistocenepackratmidden record.Pages 285-299 in J. Diamond & T. J. Case (eds.), Communityecology. Harper& Row Publ., New York. van Donk, J. 1976. 018 of the Atlantic Ocean for the entire PleistoceneEpoch. Mem. Geol. Soc. Amer. 145: 147-164. Wadsworth,F. H. & G. H. Englerth. 1959. Effectsof the 1956 hurricaneon forestsin Puerto Rico. CaribbeanForest. 1959: 38-42. Walker,D. A. 1982. Vegetation'sfourthdimension. New Phytol. 90: 419-429. & Y. Chen. 1987. Palynologicallight on tropicalrainforestdynamics.QuaternarySci. Rev. 6: 77-92. Watts, W. A. 1970. Rates of change and stability in vegetation in the perspectiveof long periods of time. Pages 195-206 in H. J. B. Birks & R. G. West (eds.), Quaternaryplant ecology. Blackwell,London. Webb, T., III. 1986. Is vegetationin equilibriumwith climate?How to interpretlate-Quaternarypollen data. Vegetatio67: 75-91. 1988. The appearanceand disappearanceof major vegetationalassemblages:Longterm vegetationaldynamicsin easternNorth America.Vegetatio69: 177-187. , P. J. Bartlein & J. E. Kutzbach. 1987. Climatic change in easternNorth America duringthe past 18,000 years;comparisonsof pollen data with model results.Pages 447462 in W. F. Ruddiman& H. E. Wright,Jr. (eds.), North America and adjacentoceans duringthe last deglaciation.DNAG Volume K-3. Geol. Soc. Amer., Boulder,Colorado. , S. E. Howe, R. H. W. Bradshaw& K. M. Heide. 1981. Estimatingplant abundances from pollen percentages:The use of regressionanalyses.Rev. Paleobot.Palynol.34: 269300. , P. J. H. Richard& R. J. Mott. 1983. A mapped history of Holocene vegetationin southernQuebec. Syllogeus49: 273-336. of ecologyand Quaternarypaleobotany.J. Ecology52(Suppl.): West,R. G. 1964. Inter-relations 47-57. 1970. Pleistocenehistoryof the Britishflora.Pages 1-11 in D. Walker& R. G. West (eds.), Studies in the vegetationhistory of the British Isles. CambridgeUniversity Press, London. 1980. Pleistoceneforest history in East Anglia. New Phytol. 85: 571-622. Whitmore,T. C. 1975. Tropicalrainforestsof the Far East. ClarendonPress, Oxford. & G. T. Prance(eds.). 1987. Biogeographyand Quaternaryhistoryin tropicalAmerica. ClarendonPress, Oxford. Whittaker,R. H. 1951. A criticism of the plant association and climatic climax concepts. NorthwestScience 25: 17-31. 1956. Vegetationof the Great Smokey Mountains.Ecol. Monogr.26: 1-80. 1975. Communitiesand ecosystems.MacmillanPubl. Co., New York. Winkler, M. G. 1985. A 12,000 year history of vegetationand climate for Cape Cod, Massachusetts.Quat. Res. 23: 301-312. Wright, H. E., Jr. 1974. Landscapedevelopment,forest fires and wildernessmanagement. Science 186: 487-495. (ed.). 1983. Late-Quaternaryenvironmentsof the United States. University of Minnesota Press, Minneapolis. . 1984. Sensitivityand responsetime of naturalsystems to climaticchangein the Late Quaternary.QuaternarySci. Rev. 3: 91-132. 1987. Synthesis;the land south of the ice sheets. Pages479-488 in W. F. Ruddiman & H. E. Wright,Jr. (eds.), North Americaand adjacentoceans duringthe last deglaciation. DNAG Volume K-3. Geol. Soc. Amer., Boulder,Colorado.