Survey
* Your assessment is very important for improving the workof artificial intelligence, which forms the content of this project
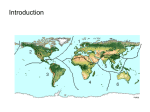
Journal of Biogeography (J. Biogeogr.) (2013) 40, 813–820 GUEST EDITORIAL 1 Department of Vertebrate Zoology, National Museum of Natural History, Smithsonian Institution, Washington, D.C. 20013-7012, USA, 2School of Biological, Earth and Environmental Sciences, University of New South Wales, NSW 2052, Australia *Correspondence: Lynne R. Parenti, Department of Vertebrate Zoology, National Museum of Natural History, Smithsonian Institution, PO Box 37012 MRC 159, Washington, D.C. 20013-7012, USA. E-mail: [email protected] Evidence and hypothesis in biogeography Lynne R. Parenti1* and Malte C. Ebach2 ABSTRACT Evidence can provide support for or against a particular biogeographical hypothesis. Treating a hypothesis as if it were evidence or an empirical observation confounds many biogeographical analyses. We focus on two recent publications that address, in part, the evolution of the biota of Sulawesi, the large Indonesian island in the centre of the Indo-Australian Archipelago. Many biogeographical explanations are hampered by invoking simple notions of mechanism or process – dispersal and vicariance – or constraints, such as dispersal from a centre of origin, and, in so doing, dismiss more complex geological phenomena such as emergent volcanoes within island chains or composite areas as irrelevant. Moreover, they do not search for, therefore never discover, biogeographical patterns that may better explain the distribution of biota through time. Keywords Biogeographical regions, co-evolution, dispersal, historical biogeography, Indo-Pacific, molecular clock, Sulawesi, vicariance, volcanoes. INTRODUCTION Historical biogeographers are like journalists. We report on the Who, What, Where, When and Why or How of biotic distributions. The Who (biota, such as bony fishes), What (particular clades, such as genera of freshwater eels) and Where (description of the distribution of each clade, in words and in maps, over time) of biotic distributions are descriptive and in practice range from the simple to the complex, from the local to the global. The When may be part empirical (during what time periods did/does the clade live) and part hypothesis [a clade is hypothesized to have split from its sister clade 45 million years ago (Ma) as inferred from a relaxed molecular clock hypothesis, the minimum age of a fossil, conformance to a geological event, or conformance to a biogeographical pattern]. The Why or How is solely a hypothesis (e.g. eggplants arose in Africa and dispersed to Asia; Weese & Bohs, 2010). Conflating these concepts – treating a hypothesis as if it were evidence or an empirical observation – introduces bias into a biogeographical analysis. The bias may come in many forms, such as favouring one explanation over another simply because no other explanation was ever considered. ALTERNATIVE EXPLANATIONS The philosophical divide in historical biogeography between those who advocate that Earth and life evolve together and ª 2013 Blackwell Publishing Ltd those who advocate centre of origin/chance dispersal explanations was enunciated by Croizat et al. (1974). In modern historical biogeographical studies the divide is between those who discover patterns versus those who generate explanations (Ebach & Humphries, 2002). As an example of the latter, the freshwater eels, family Anguillidae, were inferred to have a deep-sea origin by Inoue et al. (2010) who optimized habitats on a molecular phylogeny of anguilliform fishes: the freshwater anguillids were nested in the cladogram among a series of taxa that live in the deep sea. Freshwater eels and their close relatives live in three habitats, shallow water (SW), oceanic midwater (OM) and freshwater (FW), related as follows (summarized from Inoue et al., 2010; figure 2, node A): (SW(OM(OM, FW))). Optimization of habitats on the cladogram is presented by Inoue et al. (2010, p. 363) as the evidence for the origin of freshwater eels in the deep-sea: ‘…reconstruction of the growth habitats on the resulting tree unequivocally indicates an origination of the freshwater eels from the midwater of the deep ocean’ [italics added]. Optimization of habitats on nodes to infer an ancestral habitat is an extension of the common practice of optimizing areas on nodes to infer an ancestral area or centre of origin (e.g. Bremer, 1992; Pirie et al., 2012). Optimization to infer ancestral habitat is problematic because a deep-sea ancestor (OM) of anguillids is not ‘unequivocal’. The ancestral population could have lived in both freshwater and the deep sea, or in neither: it could have lived in shallow water, epicontihttp://wileyonlinelibrary.com/journal/jbi doi:10.1111/jbi.12069 813 L. R. Parenti and M. C. Ebach nental seas. Repetition or geographical paralogy of the OM habitat is given as the evidence that the shift in habitat was from deep-sea (OM) to freshwater (FW), but it means only that the deep-sea taxa are more common or widespread (Ebach, 1999). Other biogeographical ideas are so well-worn that they have in practice crossed the line from supporting a hypothesis to appearing to present evidence. We address two such proposals here with respect to the biota of the Indo-Pacific, in particular the Indonesian islands of Sulawesi: 1. Volcanoes (and other oceanic islands) must have been colonized via dispersal. 2. Estimates of divergence time trump biogeographical patterns. (a) widespread biota sea (b) VOLCANOES (AND OTHER OCEANIC ISLANDS) Today, many of the Philippine and Wallacean islands are generally regarded as oceanic islands because they have had no terrestrial connection to any surrounding land since their emergence…. Consequently, their biota arose predominantly via dispersal and not vicariance. (c) 1 2 Lohman et al. (2011, p. 209) Although dispersal may be ‘less parsimonious’ than vicariance… sometimes it is still the only sensible explanation of a given pattern of distribution. For inhabitants of oceanic islands, this would seem to be the case regardless of the age of the taxon. Brower & Vane-Wright (2011, p. 602) The two recent quotes above sum up what some consider a rule of thumb: endemic taxa on oceanic islands must have dispersed from terrestrial source areas. We reject this ‘rule’ because it assumes that the islands, and much of the sea surrounding them, were abiotic until life made the overseas journey from the nearest continent. Formation of a volcano or volcanic chain may be interpreted as a vicariance event. We illustrate a hypothetical example in Fig. 1. A biota may range throughout a broad area in the sea (Fig. 1a). Formation of a volcanic chain may bisect the biota (Fig. 1b) and lead to its subsequent divergence into two sister biotas (Fig. 1c). The marine biota evolves along with the volcanoes and, as the emerging volcanic islands become high enough to have established freshwater streams, taxa that once lived exclusively in the sea may become part of the terrestrial island biota. This is a gradual, co-evolutionary process. Once any land appears above sea level, it will have a biota in its tidal pools and other saline water bodies. The volcano supports a biota even before freshwater streams are established. As an example, the Hawaiian archipelago has five endemic freshwater fishes, Eleotris sandwicensis, Awaous stamineus, Lentipes concolor, Stenogobius hawaiiensis and Sicyopterus stimpsoni, all gobiiforms. The taxa (genera and their relatives) are widespread throughout the tropics and inferred to be older than the Hawaiian Islands (cf. Heads, 2011). The endemic species may be the same age as or older than the archipelago; there 814 Figure 1 Simplified representation of the formation of a volcanic chain bisecting a biota. (a) The range of a widespread biota throughout the sea. (b) Formation of a volcanic chain (arrows) in the biotic range. (c) Subsequent divergence of the biota into two sister biotas, 1 and 2, and growth of the volcanic chain. is little genetic structure of species populations among the islands, which may be interpreted as an indication that the species are endemic to the archipelago, but not to individual Hawaiian islands (Zink et al., 1996). The species have diverged from close relatives: Awaous stamineus from Hawaii and Awaous guamensis from Guam, once considered conspecific (Watson, 1992), have been shown to be genetically and morphologically distinct (Lindstrom et al., 2012). To say that the gobiiform fishes must have colonized the Hawaiian Islands after they were formed because the islands are volcanic is to reject, a priori, the study and discovery of co-evolution of the islands and the fishes (see also Nelson, 2006; Heads, 2012a). We hypothesize that Hawaii’s endemic freshwater fish lineages ranged throughout the region where the Hawaiian Islands formed and, that the lineages of part of Hawaii’s terrestrial biota ranged throughout that region, not necessarily as marine organisms, but on once emergent lands (Cain, 1944, p. 222; Heads, 2012a). Whether distributed via dispersal or vicariance, an insular terrestrial organism had to be able to survive across a vast sea. Terrestrial Hawaiian relicts include the endemic plant Hillebrandia sandwicensis, inferred to have had broadly distributed boreotropic or Journal of Biogeography 40, 813–820 ª 2013 Blackwell Publishing Ltd Evidence and hypothesis in biogeography Malesian–Pacific close relatives (Clement et al., 2004; see also Cowie & Holland, 2008). TESTING BIOGEOGRAPHICAL PATTERNS Sulawesi, in the heart of the Indo-Australian Archipelago, is a group of islands that has a special place in biogeography because of its high degree of endemism and complex geological history (Wallace, 1863, 1876; Hall, 2002). Sulawesi lies at the junction of the Asian, Australian and Pacific plates (Hall, 2002; Villeneuve et al., 2002; Spakman & Hall, 2010). Biogeographically it has been recognized as an area of overlap or interdigitation or a ‘… hinge between two worlds of life…’ (Croizat, 1958, p. 1196), Asian and Australian (Wallace, 1876). Wallacea, named formally by Dickerson et al. (1928, p. 101), is a broad region in the centre of the Indo-Australian Archipelago that includes Sulawesi. Wallacea is an undiagnosable area in a comparative, systematic biogeographical analysis (Parenti & Ebach, 2010): some areas within Wallacea are more closely related to areas outside of Wallacea than they are to each other. Part of the two worlds that collided and overlap today were identified by Parenti & Ebach (2010) as Indo-Malayan and Pandora – biogeographical subregions that were disjunct in the Oligocene, 30 Ma (Fig. 2). Recognizing and naming Pandora as a separate subregion highlights the close relationship among portions of the modern biotas of Madagascar, Australia and Sulawesi, and their surrounding seas, as demonstrated, for example, by species groups of the teleost genera Trimmatom and Hippocampus, and the gastropod genus Strombus (see Parenti & Ebach, 2009, pp. 227–234), as well as by atherinomorph rainbowfishes in the family Melanotaeniidae (Sparks & Smith, 2004) and terapontid perches of the genus Mesopristes (Vari, 1992). Once proposed, Pandora is open to test. The disjunct distribution of Madagascan/Melanesian sister groups of boas of the genera Candoia (Melanesian) and Sanzinia (Madagascar) was concluded to be (Austin, 2000, p. 348) ‘…not the result of a recent dispersal event’. More recently, Chakrabarty et al. (2012) hypothesized that the south-western Madagascan endemic gobiiform genus Typhleotris is the sister group of the north-western Australian endemic genus Milyeringa. These are blind, obligate cave fishes that live in similar subterranean karst habitats. Chakrabarty et al. (2012, p. 1) describe this as an ‘… extraordinary case of Gondwanan vicariance’. We consider the boa and blind gobiiform distributions as more support for Pandora. The modern overlap of the western (eastern Indo-Malayan) and eastern (eastern Pandora) segments of the IndoAustralian biota in eastern Borneo, southern Philippines and south-western Sulawesi – the ‘hinge’ of Croizat – is seen in the atheriniform fish family Phallostethidae. Its two subfamilies, Phallostethinae and Denatherininae (Parenti & Louie, 1998), together range broadly from Thailand to Fiji (Fig. 3). There are no known fossil phallostethids. As sister taxa, phallostethines and dentatherines are logically of the same age. We hypothesize that the ancestral population was widespread, perhaps throughout the Meso-Tethys Sea (Hall, 2011), then was disrupted by reconstruction and rearrangement of land and changes in sea level that may have facilitated expansion of one taxon into the range of the other. Asking whether they came from the west or the east is moot as it does not address the biogeographical question of how they diverged and now overlap in the centre of their distribution ranges (see also Ladiges et al., 2012). As it has since the time of Wallace, understanding the evolution of such biogeographical distributions centres on Sulawesi. Indo-Malayan Pandora * Figure 2 Biogeographical subregions Pandora and Indo-Malayan depicted on an early Oligocene (30 Ma) reconstruction of global tectonic plate arrangement (map from University of Texas Institute for Geophysics). The asterisk (*) approximates the location of the South Pandora Ridge in North Fiji (Parenti & Ebach, 2010, figure 15.6). Copyright 2010 by the Regents of the University of California. Journal of Biogeography 40, 813–820 ª 2013 Blackwell Publishing Ltd 815 L. R. Parenti and M. C. Ebach 110 E 130 E 150 E 170 E Pacific Ocean 20 N Dentatherininae 0 0 Phallostethinae 10 S Indian Ocean 90 E 110 E 130 E Figure 3 Distribution limits of the freshwater and coastal Phallostethinae and its sister taxon, the marine shorefish Denatherininae (following Parenti & Louie, 1998). The western (eastern Indo-Malayan) and eastern (eastern Pandora) clades overlap in the centre of the Indo-Australian Archipelago. Where and How did the unique biota of Sulawesi evolve? Stelbrink et al. (2012) concentrate on the hypothetical How and apply a popular approach. Time of divergence between a Sulawesi taxon and its extra-Sulawesi sister group is estimated using a relaxed molecular clock. That time is compared with temporal estimates of presumed relevant vicariance events imposed by a geological model. If the taxon divergence time is the same as or older than the inferred vicariance event, vicariance is supported. If the taxon divergence time is younger than the inferred vicariance event, dispersal is supported. Despite the logical appeal of this method (e.g. de Queiroz, 2005; Sanmartın et al., 2008; Crisp et al., 2009), its application has been seriously flawed (Nelson & Ladiges, 2009; Brower & Vane-Wright, 2011; Heads, 2012b). Molecular sequence differences constitute empirical evidence. An estimate of divergence time requires calibration of a molecular clock and is a hypothesis, not empirical evidence. Further, minimum ages of taxa as estimated from oldest fossils have been treated illogically as maximum ages in selection of prior probability curves in Bayesian estimates of divergence times in biogeographical analyses rendering their results spurious (see especially Heads, 2012b). After analysing 27 datasets representing 20 taxa, Stelbrink et al. (2012, p. 2267) conclude: ‘Dispersal seems the primary mechanism of bringing taxa to the island and a standardized molecular clock approach has [led] to the falsification of vicariance hypotheses for some Sulawesi taxa of Asian origin.’ Historical biogeography is limited in what it can say about explicit processes such as vicariance and dispersal. The analyses and conclusions of Lohman et al. (2011) and Stelbrink et al. (2012) are problematic for several reasons. First, they assume that vicariance is old and dispersal is new. Yet cyclical changes in sea level are hypothesized to have facilitated the spread of the freshwater biota throughout Sundaland sensu Metcalfe (2011) and the subsequent fragmentation of the land and its biota (Molengraaff & Weber, 1921). We hypothesize dispersion of a biota (sensu Platnick, 1976), followed by vicariance. This is likely to be followed by more 816 dispersion and more vicariance. Here, we mean range expansion or normal ecological dispersal sensu Heads (2012a), not dispersal from a centre of origin. Second, Lohman et al. (2011) and Stelbrink et al. (2012) focus on the timing of a particular geological event and invoke dispersal or vicariance as an explanation where lineage divergence time is younger or older, respectively, than the event. But that method can reject only particular vicariance events, not identify what other vicariance event may be invoked as an explanation. Lastly, and most importantly, Stelbrink et al. (2012, p. 2265) also remark: Not only is there no obvious respective pattern in the distribution of lineages that have colonized Sulawesi independently, there is in our opinion also little hard evidence for any Sulawesi clade being confined to part of the island only, which might suggest a link of its origin on Sulawesi and tectonic processes. To address the second assertion first, restricted endemism in Sulawesi is well documented (Michaux, 2010; Thomas et al., 2011; Heads, 2012a). The vastly different, disjunct floras of the Latimodjong Mountains and Mount Lompobatang (Bonthain) in south-western Sulawesi were described by Croizat (1964, p. 228) as ‘…ancient blocks of life [that] face each other in age-long immobilism’ [italics in the original]. Ricefishes, a taxon in the Stelbrink et al. (2012) analysis, include the genus Adrianichthys comprising four species, all endemic to Lake Poso, central Sulawesi (Parenti, 2008). To return to the first assertion, Stelbrink et al. (2012) claim that there is no pattern in the distribution of lineages. But they did not look for a biogeographical pattern. Instead, they assume, a priori, independent dispersal or vicariant origins of Sulawesi taxa in line with the philosophy of Crisp et al. (2011), who argue that process, not pattern, is primary in biogeography. We note that by ‘pattern’ Crisp et al. (2011) mean, for example, a single phylogenetic hypothesis of a single taxon used as the basis for biogeographical inference. In contrast, by ‘pattern’ we mean the comparison of more than one statement of area relationship across a range of taxa (Parenti & Ebach, 2009). Journal of Biogeography 40, 813–820 ª 2013 Blackwell Publishing Ltd Evidence and hypothesis in biogeography Table 1 Informative areagrams or relationships among the four areas – Sulawesi, Asia, Southeast Asia (Philippines, Moluccas, Lesser Sunda Islands East of Bali) and Australia (including New Guinea), as specified by the cladograms in Stelbrink et al. (2012) for each of their 27 molecular datasets. Dataset no. Taxon Dataset type Informative area relationship 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 Mite harvestmen Water beetles Freshwater snails Freshwater crabs Shrews Shrews Macaques Phalangerids Water beetles Sailfin silversides Cockroaches Squirrels Squirrels Megapodes Fanged frogs Fanged frogs Tarsiers Grasshoppers Ricefishes Bovids Toads Water beetles Ricefishes Macaques Water snakes Freshwater bivalves Pigs concatenated Cyt b 16S 16S Cyt b Cyt b NADH 16S COI ND5 COII 16S 12S ND2 16S 12S Cyt b COI 16S Cyt b 12S 16S 12S 12S Cyt b COI control region none Sulawesi (Asia, Australia) Asia (Sulawesi, Australia) none Sulawesi (Asia, Southeast Asia) Asia (Sulawesi, Southeast Asia) none none Australia (Sulawesi, Asia) none Sulawesi (Asia, Australia) none none none none none none none none Sulawesi (Asia, Southeast Asia) none Asia (Sulawesi, Australia) Sulawesi (Asia, Southeast Asia) none (Australia, Southeast Asia) (Sulawesi, Asia) Australia (Sulawesi, Asia) Australia (Sulawesi, Asia) Is there a biogeographical pattern expressed by the distribution of the 20 Sulawesi taxa examined by Stelbrink et al. (2012)? We extracted informative three-area statements of area relationship (Nelson & Ladiges, 1996; Parenti & Ebach, 2009) from the 27 cladograms of Stelbrink et al. (2012). A hypothetical informative three-area relationship is X(Y,Z). An uninformative area relationship is X(X,Z), in which X is paralogous or repeated. The data may be analysed by hand or by implementing a computer algorithm, as described in Parenti & Ebach (2009). The 20 taxa live in four general areas recognized by Stelbrink et al. (2012): Sulawesi, Asia, Southeast Asia (Philippines, Moluccas, and Lesser Sunda Islands East of Bali) and Australia (including New Guinea). Relationships in this example are simple enough that we can extract informative statements of area relationship manually (Table 1). Of the 27 areagrams, 15 have no informative area relationships. Among the remaining 12, there are eight informative area relationships (A–H; Table 2), distributed as in Table 3. Three of the three-area statements, A, C and D, summarize the relationships among Asia, Sulawesi and Australia. Of these, C (Australia (Sulawesi, Asia)), is shared by four clades – water beetles, water snakes, freshwater bivalves and pigs – and may be considered a general pattern. Alternative patterns, A (Sulawesi (Asia, Australia)) and D (Asia (Sulawesi, Australia)), are each supported by two clades. Another informative area relationship, G (Sulawesi Journal of Biogeography 40, 813–820 ª 2013 Blackwell Publishing Ltd (Australia, Southeast Asia)), is unique to clade 25, water snakes, hence not a general pattern. Three other informative area relationships, B, E and F, are less useful in this analysis because they do not include any areas east of Sulawesi. Informative area relationship H is of no value in this analysis because it does not include Sulawesi. Why the conflict or seeming lack of resolution? Modern Sulawesi is a biogeographical and geological composite area that harbours several different biotic areas (e.g. Thomas et al., 2011, figure 4; Heads, 2012a, figures 5–16). We have identified a general pattern, C (Australia (Sulawesi, Asia)). Yet any or all of the three broadly informative area relationships – A, C and D – may be supported by other taxa and indicate alternative general patterns. This is the type of conflict or incongruence we would expect when trying to infer the biotic relationships of a geological composite such as Sulawesi, especially from datasets that treat Sulawesi as one area. Further, the conflict reflects the overlap of Asian and Australian biotas in the centre of the Indo-Australian Archipelago, as discussed above and illustrated for phallostethid fishes (Fig. 3). To discover a robust general biogeographical pattern, we need to recognize the separate biotic areas of Sulawesi and other composites based on taxic distributions that span the Indo-Australian Archipelago. Yet, Stelbrink et al. (2012) provide no distribution maps or detailed descriptions of taxon distributions. Do shrews live throughout Sulawesi or are they 817 L. R. Parenti and M. C. Ebach Table 2 The eight informative three-area statements (A–H) specified by the 12 areagrams of Table 1. A: Sulawesi (Asia, Australia) B: Sulawesi (Asia, Southeast Asia) C: Australia (Sulawesi, Asia) D: Asia (Sulawesi, Australia) E: Asia (Sulawesi, Southeast Asia) F: Southeast Asia (Sulawesi, Asia) G: Sulawesi (Australia, Southeast Asia) H: Asia (Australia, Southeast Asia) CONCLUSIONS Table 3 Distribution of the eight (A–H) informative three-area statements (Table 2) among the 12 areagrams, numbered as in Table 1. Informative area Statement Dataset no. Taxon A A B B B C C C C D D E F G H 2 11 5 20 23 9 25 26 27 3 22 6 25 25 25 Water beetles Cockroaches Shrews Bovids Ricefishes Water beetles Water snakes Freshwater bivalves Pigs Freshwater snails Water beetles Shrews Water snakes Water snakes Water snakes restricted to particular regions? Mapping is critical to biogeographical inference. The overlap of the sister clades of phallostethid fishes simply illustrate one part of the complexity of the Sulawesi biota, for example (Fig. 3). Detailed description of areas of endemism in Sulawesi is still inchoate. About one half of the approximately 60 endemic species of Sulawesi freshwater fishes have been described since 1989 (Parenti, 2011). Phallostethids as a taxon were unknown from Sulawesi until collected in 1995 and described in 1998 (Parenti & Louie, 1998). No ricefishes were known to science from south-eastern Sulawesi until their collection in 2007 and description three years later (Parenti & Hadiaty, 2010). Interestingly, the ricefish species described in 2010 is endemic to Muna Island, a terrane hypothesized to have been submerged until it started to emerge some 15 Ma (Spakman & Hall, 2010). For the purposes of biogeography, we view the emergence of once submerged landmasses as analogous to the formation of a volcano (see above). Is Sulawesi a single biotic area with a unique history or a composite biotic area (the problem posed by Wallace in 1876)? Lohman et al. (2011) and Stelbrink et al. (2012) do not ask this important biogeographical question. Instead, they generate a series of hypotheses or explanations without considering biogeographical patterns in the form of biotic 818 area relationships. Despite their claims, Lohman et al. (2011, p. 214) have not illustrated ‘…patterns of dispersal and diversification illuminated by molecular phylogenetic and phylogeographic evidence’ [italics added]. Evidence and hypothesis are not always distinguished in historical biogeographical studies. Inferences that result from application of methods, such as optimization of areas on a cladogram to infer an ancestral area, or assumptions about mechanism, such as ‘volcanoes must always be colonized by dispersal’, add an unnecessary bias or constraint. We may speculate about the process or mechanism that may have caused a particular distribution, but always as a hypothesis, not as evidence. Rejecting particular vicariance events to explain a distribution does not demonstrate dispersal as it ignores other vicariance events that may be invoked as an explanation. We offer an alternative line of investigation that extracts and summarizes information from distributions and phylogenetic relationships: biotic area relationships, namely pattern. Rather than trying to explain individual distributional histories, we are able to reconstruct the history of biotic areas, their relationships to each other and may consider the abiotic factors (such as climate, tectonics or orogeny) that have shaped them. When we change our focus from individual taxon histories to shared biotic area histories, we move historical biogeography beyond the dispersal versus vicariance debate to a period of discovery. ACKNOWLEDGEMENTS Plate tectonic reconstruction maps were obtained through the courtesy of Lisa Gahagan and Lawrence Lawver, University of Texas Institute for Geophysics, The Plates Project (http://www.ig.utexas.edu/research/projects/plates/). Three referees provided thoughtful comments that improved the text. Figure 2 was published originally as Figure 15.6 in Parenti & Ebach (2010); it is reproduced here with permission of the University of California Press. REFERENCES Austin, C.C. (2000) Molecular phylogeny and historical biogeography of Pacific island boas (Candoia). Copeia, 2000, 341–352. Bremer, K. (1992) Ancestral areas: a cladistic reinterpretation of the center of origin concept. Systematic Biology, 41, 436–445. Brower, A.V.Z. & Vane-Wright, R.I. (2011) Anomalous areas and awkward ages: concerns about over-reliance on model-based biogeographical and temporal inferences. Systematic Entomology, 36, 601–603. Cain, S.A. (1944) Foundations of plant geography. Harper & Brothers, New York. Journal of Biogeography 40, 813–820 ª 2013 Blackwell Publishing Ltd Evidence and hypothesis in biogeography Chakrabarty, P., Davis, M.P. & Sparks, J.S. (2012) The first record of a trans-oceanic sister-group relationship between obligate vertebrate troglobites. PLoS ONE, 7, e44083. Clement, W.L., Tebbitt, M.C., Forrest, L.L., Blair, J.E., Brouillet, L., Eriksson, T. & Swensen, S. (2004) Phylogenetic position and biogeography of Hillebrandia sandwicensis (Begoniaceae): a rare Hawaiian relict. American Journal of Botany, 91, 905–917. Cowie, R. H. & Holland, B. S. (2008) Molecular biogeography and diversification of the endemic terrestrial fauna of the Hawaiian Islands. Philosophical Transactions of the Royal Society B, 363, 3363–3376. Crisp, M.D., Arroyo, M.T.K., Cook, L.G., Gandolfo, M.A., Jordan, G.J., McGlone, M.S., Weston, P.H., Westoby, M., Wilf, P. & Linder, H.P. (2009) Phylogenetic biome conservatism on a global scale. Nature, 458, 754–756. Crisp, M.D., Trewick, S.A. & Cook, L.G. (2011) Hypothesis testing in biogeography. Trends in Ecology and Evolution, 26, 66–72. Croizat, L. (1958) Panbiogeography or an introductory synthesis of zoogeography, phytogeography, and geology; with notes on evolution, systematics, ecology, anthropology, etc. Published by the author, Caracas. Croizat, L. (1964) Space, time, form: the biological synthesis. Published by the author, Caracas. Croizat, L., Nelson, G. & Rosen, D.E. (1974) Centers of origin and related concepts. Systematic Zoology, 23, 265–287. Dickerson, R.E., Merrill, E.D., McGregor, R.C., Schultze, W., Taylor, E.H. & Herre, A.W. (1928) Distribution of life in the Philippines. Monograph of the Bureau of Science, Manila, 21, 1–322. Ebach, M.C. (1999) Paralogy and the centre of origin concept. Cladistics, 15, 387–391. Ebach, M.C. & Humphries, C.J. (2002) Cladistic biogeography and the art of discovery. Journal of Biogeography, 20, 427–444. Hall, R. (2002) Cenozoic geological and plate tectonic evolution of SE Asia and the SW Pacific: computer-based reconstructions, model and animations. Journal of Asian Earth Science, 20, 353–431. Hall, R. (2011) Australia–SE Asia collision: plate tectonics and crustal flow. The SE Asian Gateway: history and tectonics (ed. by R. Hall, M.A. Cottam and M.E.J. Wilson). Geological Society, London, Special Publication, 355, 75–109. Heads, M. (2011) Old taxa on young islands: a critique of the use of island age to date island-endemic clades and calibrate phylogenies. Systematic Biology, 60, 204–218. Heads, M. (2012a) Molecular panbiogeography of the tropics. University of California Press, Berkeley, CA. Heads, M. (2012b) Bayesian transmogrification of clade divergence dates: a critique. Journal of Biogeography, 39, 1749–1756. Inoue, J.G., Miya, M., Miller, M.J., Sado, T., Hanel, R., Hatooka, K., Aoyama, J., Minegishi, Y., Nishida, M. & Tsukamoto, K. (2010) Deep-ocean origin of the freshwater eels. Biology Letters, 6, 363–366. Journal of Biogeography 40, 813–820 ª 2013 Blackwell Publishing Ltd Ladiges, P.Y., Bayly, M.J. & Nelson, G. (2012) Searching for ancestral areas and artifactual centers of origin in biogeography: with comment on east–west patterns across southern Australia. Systematic Biology, 61, 703–708. Lindstrom, D.P., Blum, M.J., Walter, R.P., Gagne, R.B. & Gilliam, J.F. (2012) Molecular and morphological evidence of distinct evolutionary lineages of Awaous guamensis in Hawai’i and Guam. Copeia, 2012, 293–300. Lohman, D.J., de Bruyn, M., Page, T., von Rintelen, K., Hall, R., Ng, P.K.L., Shih, H.-T., Carvalho, G.R. & von Rintelen, T. (2011) Biogeography of the Indo-Australian Archipelago. Annual Review of Ecology, Evolution, and Systematics, 42, 205–226. Metcalfe, I. (2011) Tectonic framework and Phanerozoic evolution of Sundaland. Gondwana Research, 19, 3–21. Michaux, B. (2010) Biogeology of Wallacea: geotectonic models, areas of endemism, and natural biogeographical units. Biological Journal of the Linnean Society, 101, 193–212. Molengraaff, G.A.F. & Weber, M. (1921) On the relation between the Pleistocene glacial period and the origin of the Sunda Sea (Java and South China-Sea), and its influence on the distribution of coral reefs and on the land- and freshwater fauna. Koninklijke Akademie van Wetenschappen, Amsterdam, 23, 395–447. Nelson, G. (2006) Hawaiian vicariance. Journal of Biogeography, 33, 2154–2155. Nelson, G. & Ladiges, P.Y. (1996) Paralogy in cladistic biogeography and analysis of paralogy-free subtrees. American Museum Novitates, 3167, 1–58. Nelson, G. & Ladiges, P.Y. (2009) Biogeography and the molecular dating game: a futile revival of phenetics? Bulletin de la Societe Geologique de France, 180, 39–43. Parenti, L.R. (2008) A phylogenetic analysis and taxonomic revision of ricefishes, Oryzias and relatives (Beloniformes, Adrianichthyidae). Zoological Journal of the Linnean Society, 154, 494–610. Parenti, L. R. (2011) Endemism and conservation of the native freshwater fish fauna of Sulawesi, Indonesia. Proceedings of the VIth National Seminar and 3rd Congress of the Indonesian Society of Ichthyologists (ed. by C. P. H. Simanjuntak, A. Zahid, M. F. Rahardjo, R. K. Hadiaty, Krismono, Haryono and A. H. Tjakrawidjaja), pp. 1–10. Masyarakat Iktiologi Indonesia [Indonesian Society of Ichthyologists], Cibinong. Parenti, L.R. & Ebach, M.C. (2009) Comparative biogeography: discovering and classifying biogeographical patterns of a dynamic Earth. University of California Press, Berkeley, CA. Parenti, L. R. & Ebach, M.C. (2010) Wallacea deconstructed. Beyond cladistics: the branching of a paradigm (ed. by D. M. Williams and S. Knapp), pp. 303–318. University of California Press, Berkeley, CA. Parenti, L.R. & Hadiaty, R.K. (2010) A new, remarkably colorful, small ricefish of the genus Oryzias (Beloniformes, Adrianichthyidae) from Sulawesi, Indonesia. Copeia, 2010, 269–274. Parenti, L.R. & Louie, K.D. (1998) Neostethus djajaorum, new species from Sulawesi, Indonesia, the first phallostethid fish 819 L. R. Parenti and M. C. Ebach (Teleostei, Atherinomorpha) known from east of Wallace’s Line. Raffles Bulletin of Zoology, 46, 139–150. Pirie, M.D., Humphreys, A.M., Antonelli, A. & Linder, H.P. (2012) Model uncertainty in ancestral area reconstruction: a parsimonious solution? Taxon, 61, 652–664. Platnick, N.I. (1976) Concepts of dispersal in historical biogeography. Systematic Zoology, 25, 294–295. de Queiroz, A. (2005) The resurrection of oceanic dispersal in historical biogeography. Trends in Ecology and Evolution, 20, 68–73. Sanmartın, I., van der Mark, P. & Ronquist, F. (2008) Inferring dispersal: a Bayesian approach to phylogeny-based island biogeography, with special reference to the Canary Islands. Journal of Biogeography, 35, 428–449. Spakman, W. & Hall, R. (2010) Surface deformation and slab-mantle interaction during Banda arc subduction rollback. Natural Geosciences, 3, 562–566. Sparks, J.S. & Smith, W.L. (2004) Phylogeny and biogeography of the Malagasy and Australasian rainbowfishes (Teleostei: Melanotaeniidae): Gondwana vicariance and evolution in freshwater. Molecular Phylogenetics and Evolution, 33, 719–734. Stelbrink, B., Albrecht, C., Hall, R. & von Rintelen, T. (2012) The biogeography of Sulawesi revisited: is there evidence for a vicariant origin of taxa on Wallace’s “anomalous island”? Evolution, 66–67, 2252–2271. Thomas, D.C., Hughes, M., Phutthai, T., Ardi, W.H., Rajbhandary, S., Rubite, R., Twyford, A.D. & Richardson, J. (2011) West to east dispersal and subsequent rapid diversification of the mega-diverse genus Begonia (Begoniaceae) in the Malesian archipelago. Journal of Biogeography, 39, 98–113. Vari, R.P. (1992) Redescription of Mesopristes elongatus (Guinchenot, 1866), an endemic Malagasy fish species (Pisces, Terapontidae). American Museum Novitates, 3039, 1–7. Villeneuve, M., Gunawan, W., Cornee, J.-J. & Vidal, O. (2002) Geology of the central Sulawesi belt (eastern Indonesia): 820 constraints for geodynamic models. International Journal of Earth Sciences, 91, 524–537. Wallace, A.R. (1863) On the physical geography of the Malay Archipelago. Journal of the Royal Geographical Society, 33, 217–234. Wallace, A.R. (1876) The geographical distribution of animals; with a study of the relations of living and extinct faunas as elucidating the past changes of the Earth’s surface. Macmillan, London. Watson, R.E. (1992) A review of the gobiid fish genus Awaous from insular streams of the Pacific Plate. Ichthyological Exploration of Freshwaters, 3, 161–176. Weese, T.L. & Bohs, L. (2010) Eggplant origins: out of Africa, into the Orient. Taxon, 59, 49–56. Zink, R.M., Fitzsimons, J.M., Dittmann, D.L., Reynolds, D.R. & Nishimoto, R.T. (1996) Evolutionary genetics of Hawaiian freshwater fish. Copeia, 1996, 330–335. BIOSKETCHES Lynne R. Parenti is a Curator of Fishes and Research Scientist at the National Museum of Natural History, Smithsonian Institution, Washington, D.C. She studies the systematics, biogeography and reproductive biology of bony fishes, collections-based comparative biology, and the theory and methods of biogeography. Malte C. Ebach is an ARC Future Fellow and Senior Lecturer in the School of Biological, Earth and Environmental Science (BEES) at the University of New South Wales (UNSW). Malte’s research focuses on Australasian biogeography, the historical development of phyto- and zoogeography, and classification. Editor: Liliana Katinas Journal of Biogeography 40, 813–820 ª 2013 Blackwell Publishing Ltd