Survey
* Your assessment is very important for improving the workof artificial intelligence, which forms the content of this project
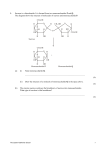
Journal of Experimental Botany, Vol. 52, No. 358, pp. 881±889, May 2001 REVIEW ARTICLE A role for `futile cycles' involving invertase and sucrose synthase in sucrose metabolism of tomato fruit Binh Nguyen-Quoc1,3 and Christine H. Foyer2 1 Centre de Recherche en Horticulture, Pavillon de l'Envirotron, FSAA, Universite Laval, QueÂbec, Canada G1K 7P4 2 Department of Biochemistry and Physiology, IACR±Rothamsted, Harpenden, Herts AL5 2JQ, UK Received 23 October 2000; Accepted 7 November 2000 Abstract Current concepts of the factors determining sink strength and the subsequent regulation of carbohydrate metabolism in tomato fruit are based upon an understanding of the relative roles of sucrose synthase, sucrose phosphate synthase and invertase, derived from studies in mutants and transformed plants. These enzymes participate in at least four futile cycles that involve sugar transport between the cytosol, vacuole and apoplast. Key reactions are (1) the continuous rapid degradation of sucrose in the cytosol by sucrose synthase (SuSy), (2) sucrose re-synthesis via either SuSy or sucrose phosphate synthase (SPS), (3) sucrose hydrolysis in the vacuole or apoplast by acid invertase, (4) subsequent transport of hexoses to the cytosol where they are once more converted into sucrose, and (5) rapid synthesis and breakdown of starch in the amyloplast. In this way futile cycles of sucroseuhexose interchange govern fruit sugar content and composition. The major function of the high and constant invertase activity in red tomato fruit is, therefore, to maintain high cellular hexose concentrations, the hydrolysis of sucrose in the vacuole and in the intercellular space allowing more efficient storage of sugar in these compartments. Vacuolar sugar storage may be important in sustaining fruit cell growth at times when less sucrose is available for the sink organs because of exhaustion of the carbohydrate pools in source leaves. Key words: Futile cycles, sucrose metabolism, tomato fruit, invertase, sucrose synthase. 3 Introduction Tomato fruit are an excellent model for the investigation of the regulation of sink activity and strength (Ho, 1996). They are simultaneously utilization-sinks, sugar-storage sinks and starch-storage sinks. Attempts to enhance crop yield through genetic manipulation of sink activity have thus far met with little success largely because of the plasticity of metabolism (for review see Koûmann et al., 1996; Sonnewald et al., 1994). Studies on the metabolism, growth and development of `fruit' models such as tomato, therefore, remain indispensable for the development of strategies for the improvement of harvest index. Tomato leaves, in common with most higher plants, use sucrose as the major form of transported carbon. Utilization of sucrose, a disaccharide composed of glucose and fructose, depends on cleavage into these hexose units. This is catalysed by one of two enzymes, SuSy or invertase. SuSy is a glycosyl transferase which, in the presence of UDP, converts sucrose into UDP-glucose and fructose. In contrast, invertase is a hydrolase cleaving sucrose into the two monosaccharides. SuSy is a cytosolic enzyme which is crucial to sucrose utilization in fruit development (Sun et al., 1992; Wang et al., 1993). There is a strong correlation between SuSy activity, the rate of growth and amount of starch accumulated in tomato fruit (Yelle et al., 1988). Extracellular and vacuolar invertases link the transport of sucrose into the apoplast and vacuole to hexose transport across the plasmalemma and tonoplast, respectively. Different extracellular and vacuolar acid invertase isoenzymes have been puri®ed and cloned from tomato and are characterized by highly differential sink tissue-speci®c To whom correspondence should be addressed. Fax: q1 418 656 7871. E-mail: [email protected] Abbreviations: ADPglc ppase, ADP-glucose pyrophosphorylase; BR, brassinosteroids; DAA, days after anthesis; F, fructose; F6P, fructose 6-phosphate; G, glucose; G1P, glucose 1-phosphate; G6P, glucose 6-phosphate; SPS, sucrose phosphate synthase; SuSy, sucrose synthase; UDPG, UDP glucose. ß Society for Experimental Biology 2001 882 Nguyen-Quoc and Foyer expression patterns (Konno et al., 1993; Sato et al., 1993; N'tchobo, 1998; Godt and Roitsch, 1997). Tomato invertases are encoded by a small gene family that respond differently to environmental and metabolic cues (Godt and Roitsch, 1997). One of the four extracellular invertase genes of tomato (Lin 6), for example, has been found to be induced by brassinosteroids (BR) in autotropic tomato cell suspension cultures linking BR-induced growth responses to localized regulation of the source± sink relationship (Prols et al., 2000). The role of each of the extracellular and vacuolar invertase isoenzyme forms in fruit development remains to be elucidated. The correlation between the activity of ADP-glucose pyrophosphorylase (ADPglc ppase) and the amount of starch in tomato fruit (Yelle et al., 1988) and in other plants (Okita, 1992; Preiss, 1988, 1991), has led to the conclusion that this enzyme controls starch accumulation. This conclusion appears to be convincing in certain cases, but not at all in the other cases. The questions as to what limits starch accumulation and which enzymes are the most important in the regulation of starch synthesis therefore remain unresolved. The aim of the present review is to discuss recent evidence that clari®es these crucial issues and to present an integrating hypothesis of the mechanisms exerting control of carbohydrate metabolism in tomato fruit. Evidence is presented in support of an hypothesis involving four futile cycles of degradation and synthesis of sucrose in the cytosol, in the vacuole and in the apoplast. The data supporting the existence of each of these cycles will now be discussed. A `futile cycle' of degradation and synthesis of sucrose in the cytosol Sucrose unloading in tomato fruit Recent advances in the analysis of sucrose transporters (Gahrtz et al., 1996; Riesmeier et al., 1992, 1993; Sauer and Stolz, 1994; Sauer and Tanner, 1989; Lemoine et al., 1996; Weise et al., 2000) and of symplast connections and paths of phloem unloading in plants (Patrick 1990; Ruan and Patrick, 1995; Yin et al., 1995) have failed to resolve key issues regarding sucrose unloading in sink organs. As in other sink organs, the pattern of sucrose unloading is not constant during tomato fruit development. The unloading process consists of two phases: (i) a period of rapid fruit growth occurring from 0 to 30±35 d after anthesis (DAA), and (ii) a phase after rapid growth has ceased. This lasts from about 35 DAA (depending on the variety) to maturity and is characterized by slow growth and very little additional accumulation of dry matter. Two distinct modes of phloem unloading appear to operate in each of these two stages. In the ®rst, sucrose is unloaded intact, i.e. it is not degraded to its component sugars prior to unloading (Damon et al., 1988; Dali et al., 1992; N'tchobo et al., 1999). Similarly, experiments investigating the unloading of asymmetrically-labelled sucrose have shown that sucrose is not hydrolysed prior to unloading (Dali et al., 1992; N'tchobo et al., 1999). Despite the presence of acid invertase in the cell wall and intercellular spaces, it suggests that in the ®rst phase of fruit development sucrose is unloaded into the tomato fruit via the symplast. This conclusion is consistent with the observed continuity of cell connections, and numerous plasmodesmata, early in tomato fruit development. Later in tomato fruit development these symplastic connections are lost and sucrose is rapidly hydrolysed during unloading (Ruan and Patrick, 1995). Similarly, different patterns of glucose and sucrose uptake were observed in protoplasts, isolated from fruit at different development stages (Brown et al., 1997). The uptake of glucose (but interestingly, not sucrose) was similar to that observed in the intact fruit (Brown et al., 1997). The small amount of sucrose that is unloaded by the apoplastic pathway is hydrolysed by invertase and imported into the fruit cells by hexose transporters. Later in fruit development the apoplastic pathway becomes the predominant. The following discussion concerns the mechanisms of sucrose metabolism found principally in the ®rst phase of fruit development when SuSy is highest and its activity determines sink strength. Most of the data discussed below were obtained with whole tomato fruit including the seeds. However, since the parenchyma cells of the pericarp represent the main part of the tomato fruit, the discussion largely concerns sucrose metabolism in this tissue. Sucrose unloading in tomato fruit is a strictly controlled process The amount of sucrose unloaded into tomato differs with the age (Walker and Ho, 1977) and developmental stage of the fruit. The two tomato lines possess similar fruit SuSy activities, but have very different invertase activities and show very similar patterns of sucrose unloading (N'tchobo et al., 1999). In other experiments, in which the activities of SuSy and SPS were increased in transgenic plants, the rate of sucrose unloading into fruit was proportionally increased (Nguyen-Quoc et al., 1999). Sucrose unloading capacity was substantially decreased in 7-d-old fruit from transgenic plants where SuSy activity was decreased to 2% of control values. However, the capacity of sucrose unloading in 23 DAA fruit, a point at which SuSy activity is maximal in untransformed fruit, was not signi®cantly decreased in the antisense SuSy plants compared to controls (D'Aoust et al., 1999). These observations provide evidence that invertase exerts little or no control over sucrose unloading during the rapid growth phase. While SuSy activity is the limiting factor in the control of sucrose unloading in fruit for up to the ®rst Sucrose metabolism in tomato fruit 883 10±15 d after anthesis, these data do not preclude a role for invertase in the regulation of assimilate import into older tomato fruit (N'tchobo, 1998). The close correlation observed between the amount of sucrose unloaded from the phloem and fruit SuSy activity does not allow us to distinguish between (1) the rate of sucrose unloading and (2) SuSy activity as the major driving force determining sink strength. Moreover, sucrose unloading is maintained, albeit at a low level, during the later phase of slow growth (Walker and Ho, 1977), when SuSy activity is largely absent. Sucrose unloading may be controlled, at least in part, by the activity of the sugar transporters. These are regulated by feedback controls related to the rate of sucrose hydrolysis by SuSy. Re-synthesis of sucrose Tomato fruit contain two enzymes that are capable of catalysing sucrose synthesis. These are SuSy and sucrose phosphate synthase (SPS). The rate of sucrose synthesis varies markedly with fruit age, a good correlation being observed between sucrose synthesis and SuSy activity (N'tchobo et al., 1999). It is important to consider whether SuSy can catalyse sucrose synthesis in vivo. When hexoses are the sole carbon source in the unloading medium, SuSy has been shown to catalyse sucrose synthesis (Geigenberger and Stitt, 1993; Viola, 1996). Although SuSy preferentially functions in the direction of sucrose cleavage under in vivo conditions where sucrose is unloaded, it is suggested that SuSy is also capable of catalysing the synthetic reaction (Geigenberger and Stitt, 1993; Viola, 1996). There are good reasons for believing that SuSy does indeed catalyse sucrose synthesis in vivo, in tomato fruit. Firstly, tomato fruit accumulate high concentrations of hexoses (100 mmol g 1 FW) in vacuoles and equal amounts of glucose and fructose are present in the tissues. The vacuole represents a large and possibly important source of fructose, which could drive the reaction in the synthetic direction. Secondly, while the rate of sucrose unloading into tomato fruit varies markedly during fruit development, the turnover rate remains constant. This implies that, for a given unit of time, the proportion of unloaded sucrose that is again converted into sucrose, does not change with fruit age. This means that the absolute amount of re-synthesized sucrose ¯uctuates considerably with fruit age, by as much as a factor of 3 (N'tchobo et al., 1999). This would require considerable variations in the activities of the enzymes catalysing sucrose synthesis. SPS activity, however, remains low throughout fruit development (Dali et al., 1992; Miron and Schaffer, 1991; Klann et al., 1993). Considered together these features suggest that sucrose synthesis is catalysed by both SPS and SuSy in the rapid growth phase of tomato fruit development. Rapid turnover of sucrose unloaded in tomato fruit Once unloaded into the tomato fruit, sucrose is broken down and rapidly re-synthesized. Using sucrose labelled asymmetrically, it was found that only 5±10% of the sucrose molecules were labelled symmetrically after a 1 h pulse of sucrose unloading in vitro. After a 2 h of subsequent chase, 60±90% of the sucrose was symmetrically labelled (N'tchobo et al., 1999; Nguyen-Quoc et al., 1999). It was concluded, therefore, that almost all the sucrose unloaded was re-synthesized within 2 h. It is surprising that the turnover rate of sucrose was virtually identical in control fruit and fruit from a mutant lacking invertase (N'tchobo et al., 1999), whether measured at 10, 20 or 40 DAA. In contrast, sucrose turnover in plants overexpressing SPS was increased by 60% compared to untransformed controls. In the SPS overexpressors, fruit SuSy activity was also 25% higher than in controls. This ®nding, together with the observed absence (or very low activity) of alkaline invertase in young tomato fruits and the symplastic pathway of sucrose unloading, supports the hypothesis that the cleavage of unloaded sucrose is catalysed by SuSy in the cytosol. Acid invertase is localized in the vacuole and extracellular compartments of the fruit. Therefore, invertase can only hydrolyse sucrose when it is transported into these compartments. A small amount of the sucrose unloaded from the phloem into the fruit cells is exported rapidly from the cytosol to the apoplast where it is hydrolysed by apoplastic invertases (N'tchobo, 1998). The rate of sucrose hydrolysis in the apoplast is small compared to that in the cytosol catalysed by SuSy, which is suf®cient to explain the constant sucrose turnover rate. Similar mechanisms have been described in other sink organs, such as developing potato tubers (Geigenberger and Stitt, 1993; Viola, 1996) and Ricinum communis cotyledons (Geigenberger and Stitt, 1993) (Fig. 1). A `futile cycle' of degradation and synthesis of sucrose via the vacuole and apoplast implicating a transient storage role for acid invertase Localization of acid invertase in the tomato fruit In tomato, most of the invertase activity is attributable to soluble acid invertase, the other alkaline and insoluble invertase isoforms being either absent or present in negligible amounts (Husain, 1999; Husain et al., 1999). Tomato acid invertase isoforms have been puri®ed and cloned (Yelle et al., 1991; Klann et al., 1992; Konno et al., 1993; Sato et al., 1993) as have invertase inhibitor proteins (Greiner et al., 1998; Sander et al., 1996). The tomato apoplastic invertases are encoded by at least four 884 Nguyen-Quoc and Foyer Fig. 1. Model of the four principal `futile cycles' involved in sucrose metabolism in tomato fruit. The ®rst `futile cycle' involves degradation of sucrose in the cytosol via sucrose synthase and sucrose re-synthesis of sucrose by SPS. The second `futile cycle' involves degradation of sucrose in vacuole by invertase and re-synthesis in the cytosol catalysed by SPS or SuSy. The third futile cycle involves sucrose degradation in the apoplast by invertase followed by sucrose synthesis in the cytosol catalysed by SPS or SuSy. The fourth cycle involved simultaneous synthesis and degradation of starch on amyloplasts and connection to the cytosol via G6P. Enzymes: (1) UPD-glucose pyrophosphorylase, (2) phosphoglucomutase (3) phosphoglucose isomerase, (4) fructokinase and hexokinase, (5) ADP-glucose pyrophosphorylase, (6) starch synthase, (7) starch phosphorylase and amylase, (8) glucokinase and hexokinase. different genes that are characterized by highly speci®c tissue expression (Godt and Roitsch, 1997). Investigations to determine the activity and role of each acid invertase isoform are complicated by dif®culties in developing unambiguous techniques for the study of compartmentation and the complex regulation of invertase by inhibitors. Immunogold labelling has showed that invertase protein is present in the cell wall and vacuole compartments of tomato fruit cells (Husain et al., 2000a). The activity of invertase in each cellular compartment may be greatly decreased, however, by interaction a speci®c inhibitor protein (Greiner et al., 1998, 1999; Husain et al., 2000a; Sander et al., 1996). Correlations between fruit invertase activity and hexose accumulation suggest that most acid invertase activity is located in the vacuole, an hypothesis supported by the localization of invertase in tomato fruit protoplasts (Konno et al., 1993). Conversely, Sato and co-workers (Sato et al., 1993) have convincingly argued that activity is also found in the intercellular space. In the pericap of 20 and 50 DAA tomato fruits about 10% of acid invertase activity was found in the apoplast (N'tchobo, 1998). Synthesis of sucrose in the cytosol and degradation in the vacuole In photosynthetic cells where the rate of sucrose synthesis is high, there is only a very limited amount of futile cycling between sucrose and hexoses (Foyer 1986, 1987; Geigenberger and Stitt, 1991; Huber, 1989; Wendler et al., 1990), even though sucrose in the cytosol is transferred to the vacuole or apoplast where it is degraded. In tomato fruit, high vacuolar hexose concentrations and rapid turnover of sucrose in the cytosol support the operation of a much more active futile cycle involving inter-conversions between sucrose and hexoses. The major function of the high and constant invertase activity in red tomato fruit is to maintain the cellular hexose concentrations. The amount of hexose found in tomato leaves and fruit is determined by the activity of vacuolar acid invertase. A decreased invertase activity would lead to increases in sucroseuhexose ratios Sucrose metabolism in tomato fruit 885 (Yelle et al., 1988; Klann et al., 1993; Scholes et al., 1996; Ohyama et al., 1995). This emphasizes not only the continuous hydrolysis of sucrose catalysed by invertase, but also the possibility of continuous sugar exchange between cytosol and vacuole (sucrose in¯ux and hexose ef¯ux). The possible physiological roles of acid invertase and the function of this futile cycle in sink organ development must now be considered. A futile cycle involving the apoplast In tomato fruits, apoplastic invertase is present at all stages of fruit development (N'tchobo, 1998; Sato et al., 1993; Godt and Roitsch, 1997). In the second stage of fruit development during which sucrose is unloaded via the apoplast, acid invertase is considered to be the main enzyme of sucrose hydrolysis (Walker et al., 1978). In young tomato fruit in which sucrose is unloaded via the symplast, the role of apoplastic invertase is not known. Only a small proportion of the sucrose unloaded from the phloem in young fruit is exported from the cytosol (N'tchobo, 1998). Hexoses produced from sucrose in the apoplast are then returned to the cytosol for sucrose re-synthesis forming a different futile cycle of sucrose degradation and synthesis. The action of apoplastic invertase probably is essentially similar to that of vacuolar invertase. However, the vacuole is a storage organ for one single cell while the apoplast is a storage organ for several cells, because the apoplast space connects cell to cell. The similarity between these futile cycles of sucrose synthesis and degradation via vacuole or apoplast could explain the similar effect of high hexose accumulation observed in the plant over-expressing yeast invertase in the apoplast or in the vacuole (Sonnewald et al., 1991; Weber et al., 1998). The concentration of hexoses in the cells could be dependent just on invertase activity, regardless of its localization because of the interconnection between these futile cycles. Physiological role of invertase The major function of invertase is to catalyse sucrose hydrolysis. Other minor catalytic activities are known, for example, invertase-mediated hydrolysis of raf®nose, hydrolysis and synthesis of fructan (Pollock, 1986), or synthesis of sucrose from raf®nose and glucose (Nadkarni et al., 1992). If the vacuole is considered to be a storage organelle, then vacuolar invertase would be expected to exercise a storage function. However, data con®rming this storage function are lacking and the physiological role of acid invertase in tomato fruit remains somewhat debatable. Tomato plants possessing low invertase activity, as a result of either antisense transformation or classical breeding (Ohyama et al., 1995; Scholes et al., 1996; Yelle et al., 1988), had similar rates of fruit growth to control plants. In this case the fruit had an altered sugar composition. Transformed tomatoes with low invertase activity accumulate more sucrose than hexose and the fruit are slightly smaller than those of control plants (Klann et al., 1996). This would suggest that sucrose only accumulates in tomato fruit in the absence of high soluble acid invertase activity. Fruit from L. pimpinellifolium have higher invertase activities, greater hexose contents and lower sucrose accumulation than L. esculentum fruit (Husain et al., 2000b). Invertase activity is present even in young green L. pimpinellifolium PI 126436 fruit and increases throughout development. Introgression of the acid invertase gene on chromosome 3 from Lycopersicon pimpinellifolium PI 126436 into L. esculentum FM 6203 modi®ed fruit sugar composition but not the amount of sugar accumulated (Husain et al., 2000b). The traits of high invertase activity and high hexoses were lost in fruit of progeny containing the L. pimpinellifolium invertase gene, suggesting that posttranscriptional controls are important in determining overall fruit invertase activity. Hexoses and not sucrose accumulate in the vacuoles of most plants (Heineke et al., 1996). It is interesting to speculate why this is the case. Considering that the vacuole is a storage organelle, it is possible that sucrose hydrolysis in the vacuole increases the storage rate (or retention rate) of sugar in this compartment. The calculated fruit hexose content during the period of rapid growth (10±25 DAA) is 100±150 mmol g 1 FW. In the low-invertase mutants, the sucrose content is about 30 mmol g 1 FW, a value similar to that of the hexose pool. Sucrose loading into the vacuole by the sucrose antiport-transporter has been demonstrated in various species (Andreev et al., 1990; Briskin et al., 1985; Getz and Klein, 1994; Getz et al., 1991; Niland and Schmitz, 1995) and there is no requirement for sucrose hydrolysis to allow vacuolar loading or unloading. Taking this into account, the following hypothesis is proposed: the hydrolysis of sucrose in the vacuole and in the intercellular space allows more ef®cient storage of sugar in these compartments. It is assumed, for example, that the storage rate (the storage rate equals the import rate minus the export rate) of sugar could be doubled when export rates of sucrose and hexose are equal. In this scenario sucrose is imported into vacuoles while hexoses are exported. Every molecule of sucrose hydrolysed in the vacuole gives two molecules of hexose. Hence, twice as many sugar molecules are exported as sucrose molecules imported and hydrolsed in the vacuole. Recent data on sugar uptake into isolated tonoplast vesicles in vitro, showed that net hexose uptake was more ef®cient than that of sucrose (Milner et al., 1995). The term `sugar uptake' in this experiment corresponds to the net sugar uptake rate (sugar imported minus sugar exported), that is equivalent to the sugar storage rate. The effect of accumulation of 886 Nguyen-Quoc and Foyer storage sugars in the vacuole through sucrose±hexose conversion would be basically analogous to the effect of conversion of sucrose to fructan or other polysaccharides. The conversion of sucrose to hexoses may have other bene®ts. For example, hexose accumulation may function to increase the osmotic potential of the tissues and allow improved nutrient uptake. Hexoses play a role in regulating gene expression and therefore, accumulation in the vacuole would permit hexose storage without adverse effects on gene expression since the hexoses would be effectively excluded from the cytosol. The hypothetical effect of vacuolar sugar storage discussed above may be important for cell growth, especially towards the end of the dark period, when less sucrose is available for unloading into sink organs owing to exhaustion of stocks in source leaves. This effect is all the more marked when plants are grown under a regime of short photoperiods or at low temperatures or under weak illumination. In these conditions, the low-invertase mutants produce relatively few fruit (Klann et al., 1996), even though fruit production is normal if they are grown with suf®cient light and at long photoperiods. `Futile cycle' of synthesis and degradation of starch in the amyloplast Starch accumulation is high during the ®rst stage of fruit development, that corresponds to the period of rapid fruit growth. The kinetics of starch accumulation can be divided into two phases. Net starch accumulation occurs in the ®rst phase (0±20 DAA) while in the second phase (20±35 DAA) net starch degradation occurs. This pattern of starch accumulation correlates well with measured changes in ADPglc ppase activity (Yelle et al., 1988). Similar relationships have often been found in starch storage organs from other plants. This implies that the quantity of accumulated starch is related to the quantity of product (ADP-glucose) supplied through the reaction catalysed by ADPglc ppase. Since all the enzymes necessary for starch synthesis and degradation are present throughout fruit development, it may be inferred that some capacity for starch synthesis is always present. In order to test this hypothesis, the amount of starch synthesized from radioactively-labelled sucrose unloaded into tomato fruit was studied in vitro (N'tchobo et al., 1999; Nguyen-Quoc et al., 1999). These studies demonstrated that synthesis of starch could be observed at all developmental stages. Surprisingly, the amount of newly synthesized starch was proportional to the amount of starch already accumulated in the fruit, while the proportion of unloaded sucrose used for synthesis of starch was always constant about 25% (N'tchobo et al., 1999; Nguyen-Quoc et al., 1999; D'Aoust et al., 1999). These results show clearly that there is continuous starch synthesis and breakdown in the tomato fruit. The extent of starch accumulation will depend on the relative rates of synthesis and degradation. In tomato fruits, the most important enzyme in starch breakdown is starch phosphorylase, which degrades starch to produce G1P. The amylase activity is very low and the activities of both starch phosphorylase and amylase change little during fruit development (Yelle et al., 1988). The rate of starch synthesis is variable whereas the rate of breakdown appears to be relatively constant. The starch amount accumulated decreases when the quantity of starch newly synthesized is lower than the starch amount degraded. The phenomenon of simultaneous starch synthesis and degradation in amyloplasts has also been described for other species such as pea (Hargreaves and ap Rees, 1988), banana (Hill and ap Rees, 1994) and potato (Geigenberger and Stitt, 1991; Viola, 1996; Sweetlove et al., 1996, 1999), constitutes an other example of `futile cycle'. The futile cycle of starch synthesis and degradation presents two potential points for rigorous control. The ®rst resides over the enzyme ADPglc ppase and the second over the point of entry (from cytosol to amyloplast) and exit (from amyloplast to cytosol) of glucose 6-P. The studies on starch synthesis in sink organs (maize, pea, Arabidopsis, potato) where ADPglc ppase activity is diminished via mutation or antisense transformation, substantiate the regulatory role of this enzyme in the direction of synthesis, i.e. limited enzyme activity limits the synthesis of starch (Tsai and Nelson, 1966; Lin et al., 1988; Neuhaus and Stitt, 1990; MuÈller-RoÈber et al., 1992; Denyer et al., 1995). Conversely, there is no evidence to suggest that augmentation of ADPglc ppase activity causes a marked enhancement of starch content. The increase of starch content in potato plants over-expressing an E. coli gene for ADPglc ppase, was disproportionate to the increase in activity (Stark et al., 1992). In a similar experiment, Sweetlove and co-workers showed no effect on starch accumulation (Sweetlove et al., 1996). All these ®ndings indicate that another control point over starch synthesis, apart from ADPglc ppase, must exist. One candidate is the concentration of hexose phosphates in the amyloplasts, or the rate of exchange of hexose phosphates between the cytosol and the amyloplast. In tomato, the principal hexose-P which crosses the amyloplast membrane is G6P (N'tchobo, 1998) which is also the case in other dicot species (Hill and Smitt, 1991; Neuhaus et al., 1993; Viola et al., 1991). All the `futile cycles' are interconnected and co-ordinated by sucrose synthase to form a regulatory cycle SuSy is strongly expressed in most plant storage organs and its activity is often found to correlate with starch accumulation or with organ growth. A recent study Sucrose metabolism in tomato fruit 887 (Zrenner et al., 1995), in which SuSy expression in potato tubers was repressed, demonstrates the crucial role of this enzyme in sink organs. Nevertheless, current understanding of how SuSy activity regulates starch synthesis, sucrose accumulation, or growth remains far from complete. From a consideration of other hypotheses (Dancer et al., 1990; Geigenberger and Stitt, 1991, 1993; Hill and ap Rees, 1994; Huber, 1989; Viola, 1996), as well as the authors' own results, an explanation is offered here which, it is hoped, will prove fruitful in provoking further discussion and experimental work. Sucrose unloading in sink organs takes place primarily via the symplast and the quantity unloaded is determined by SuSy activity. The concept that sink strength is determined by SuSy activity is, thus, widely accepted. In the main futile cycle, sucrose degradation is catalysed by SuSy while sucrose synthesis is catalysed by SPS or SuSy. This reaction sequence effectively controls the concentration of several major metabolites including UDPG, G1P, G6P, F6P, F, G, and sucrose itself. These metabolites and their products regulate cell wall synthesis, the respiration rate, starch synthesis, storage in the vacuoles and other product synthesis. For example, the amount of accumulated or newly-synthesized starch will depend on the cytosolic concentration of G6P. In the same way, the rate of G6P utilization will be related to the concentration of F6P, which will in turn depend on rates of synthesis and breakdown of sucrose. An increase in alkaline invertase and SPS activities would be capable, in theory, of compensating for a decrease in SuSy activity. Conclusions The key features of the regulation of sugar metabolism in tomato fruit involves four principal futile cycles that are all strictly connected. These are (i) rapid, continuous degradation and re-synthesis of sucrose in the cytosol. In this cycle, sucrose degradation is catalysed by SuSy and sucrose synthesis by SuSy or SPS. It is proposed that SuSy has the major regulatory role in this cycle. (ii) Hydrolysis of sucrose in the vacuole, catalysed by acid invertase. Most of the hexose is sequestered in the vacuole but some is re-converted into sucrose in the cytosol. These two opposing reactions form the second `futile cycle', which functions to increase the ef®ciency of sugar storage in these compartments. It is suggested that the physiological role of acid invertase is to provide a store for sucrose equivalents (hexoses). (iii) Hydrolysis of unloaded sucrose in the apoplast catalysed by apoplastic invertase. Most of the hexoses formed in the apoplast is re-converted to sucrose in the cytosol forming the third futile cycle. (iv) Rapid ongoing synthesis and breakdown of starch in the amyloplast. This constitutes the fourth `futile cycle'. The relative rates of synthesis and breakdown determine the quantity of starch accumulated. The overall rate of starch synthesis is controlled by ADPglc ppase, however, the relative rates of synthesis and degradation are controlled by the concentration of G6P. This, in turn, depends on the relative ¯ux of this metabolite across the amyloplast membrane. The G6P concentration in the amyloplast will hence also depend, to some degree, on the SuSy activity in the cytosol. The possibility cannot be excluded that differential activity of the tonoplast-bound sugar transporters (hexose and sucrose-transporters) plays a regulatory role in determining the direction of the SuSy reaction by control of the cytosolic sucrose and hexoses pools (Getz and Klein, 1995). Acknowledgements We are thankful to Dr HP Getz, Dr S Huber and Dr A Lecharny for their discussions on the preparation of this paper. References Andreev IM, Korenkov VD, Orlova MS. 1990. SucroseuHq antiport across the tonoplast and the possible role of vacuolar Hq ATPase in turgor control of sucrose transport in cells of the sugar beet root. Soviet Plant Physiology 37, 324±330. Briskin DP, Thornley WR, Wyse RE. 1985. Membrane transport in isolated vesicles from sugar beet taproot: evidence for a sucroseuHq-antiport. Plant Physiology 78, 871±875. Brown MM, Hall LJ, Ho LC. 1997. Sugar uptake by protoplasts isolated from tomato fruit tissues during various stages of fruit growth. Physiologia Plantarum 101, 533±539. Dali N, Michaud D, Yelle S. 1992. Evidence for the involvement of sucrose phosphate synthase in the pathway of sucrose accumulation in sucrose accumulating tomato fruits. Plant Physiology 99, 434±438. Damon S, Hewitt J, Nieder M, Bennett AB. 1988. Sink metabolism in tomato fruit. II. Phloem unloading and sugar intake. Plant Physiology 87, 731±736. Dancer J, Hatzfeld WD, Stitt M. 1990. Cytosolic cycles regulate the turnover of sucrose in heterotrophic cell-suspension cultures of Chenopodium rubrum L. Planta 182, 223±231. D'Aoust MA, Yelle S, Nguyen-Quoc B. 1999. Antisense inhibition of tomato fruit sucrose synthase decreases fruit setting and sucrose unloading capacity of young fruit. The Plant Cell 11, 2407±2418. Denyer K, Foster J, Smith AM. 1995. The contributions of adenosine 59-diphosphoglucose pyrophosphorylase and starch-branching enzyme to the control of starch synthesis in developing pea embryos. Planta 197, 57±62. Foyer C. 1986. The regulation of carbon regulation in photosynthesis. Chemistry in Britain 22, 723±726. Foyer CH. 1987. The basis for source±sink interaction in leaves. Plant Physiology and Biochemistry 25, 649±657. Gahrtz M, Schmelzer E, Stolz J, Sauer N. 1996. Expression of the PmSUC1 sucrose carrier gene from Plantago major L. is induced during seed development. The Plant Journal 9, 93±100. Geigenberger P, Stitt M. 1991. A `futile cycle' of sucrose synthesis and degradation is involved in regulating partitioning between sucrose, starch and respiration in cotyledons of 888 Nguyen-Quoc and Foyer germinatitng Ricinus communis L. seedlings when phloem transport is inhibited. Planta 185, 81±90. Geigenberger P, Stitt M. 1993. Sucrose synthase catalyses a readily reversible reaction in vivo in developing potato tuber and other plant tissues. Planta 189, 329±339. Getz HP, Klein M. 1994. Further evidence for sucroseuHq antiport across the tonoplast of red beet root vacuoles. Biologia Plantarum 36, S240. Getz HP, Klein M. 1995. A preliminary turnover rate for sucroseuHq-antiport on the tonoplast of red beet storage tissue. In: Pontis HG, Salerno GL, Echeverria EJ, eds. Current topics in plant physiology, Vol. 14. Maryland USA: American Society of Plant Physiologists, 225±236. Getz HP, Thom M, Maretzki A. 1991. Proton and sucrose transport in isolated tonoplast vesicles from sugarcane stalk tissue. Physiologia Plantarum 83, 404±410. Godt DE, Roitsch T. 1997. Regulation and tissue-speci®c distribution of mRNAs for three extracellular invertase isoenzymes of tomato suggests an important function in the establishing and maintaining sink metabolism. Plant Physiology 115, 273±282. Greiner S, Krausgrill S, Rausch T. 1998. Cloning of a tobacco apoplasmic invertase inhibitor. Proof of function of the recombinant protein and expression analysis during plant development. Plant Physiology 116, 733±742. Greiner S, Rausch T, Sonnewald U, Herbers K. 1999. Ectopic expression of a tobacco invertase inhibitor homolog prevents cold-induced sweetening of potato tubers. Nature Biotechnology 17, 708±711. Hargreaves JA, ap Rees T. 1988. Turnover of starch and sucrose in root of Pisum sativum. Phytochemistry 27, 1627±1629. Heineke D, Lolhaus G, Winter H. 1996. Compartmentation of CuN metabolism. In: Foyer CH, Quick WP, eds. A molecular approach to primary metabolism in higher plants. London: Taylor and Francis Publishers, 219±236. Hill LM, Smitt AL. 1991. Evidence that glucose 6-phosphate is imported as substrate for starch synthesis by the plastids of developing pea embryos. Planta 185, 91±96. Hill SA, ap Rees T. 1994. Fluxes of carbohydrate metabolism in ripening bananas. Planta 192, 52±60. Ho LC. 1996. Tomato. In: Zamski E, Shaffer AA, eds. Photoassimilate distribution in plants and crops, source±sink relationships. New York: Marcel Dekker, Inc. 709±728. Huber SC. 1989. Biochemical mechanism for regulation of sucrose accumulation in leaves during photosynthesis. Plant Physiology 91, 656±662. Husain SE. 1999. The role of invertase in the carbohydrate metabolism of tomato fruit. PhD thesis, University of Cambridge, UK. Husain SE, Harrison J, ap Rees T, Shields R, Foyer CH. 1999. The role of invertase in the carbohydrate metabolism of tomato fruit. Acta Horticulturae 487, 77±84. Husain SE, Harrison J, Shields R, Foyer CH. 2000a. Invertase protein but not activity is present throughout development of Lycopersicon esculentum and L. pimpinellifolium fruit. New Phytologist (in press). Husain SE, James C, Sheilds R, Foyer CH. 2000b. Manipulation of fruit sugar composition but not content in Lycopersicon esculentum fruit by introgression of an acid invertase gene from L. pimpinellifolium. New Phytologist (in press). Klann E, Yelle S, Bennet AB. 1992. Tomato fruit acid invertase complementary DNA. Plant Physiology 99, 351±353. Klann HM, Chetelat RT, Bennet AB. 1993. Expression of acid invertase gene controls sugar composition in tomato (Lycopersicon) fruit. Plant Physiology 103, 863±870. Klann HM, Hall B, Bennet AB. 1996. Antisense acid invertase (TIV1) gene alters soluble sugar composition and size in transgenic tomato fruit. Plant Physiology 112, 1321±1330. Koûmann J, MuÈller-RoÈber B, Riesmeier JW, Willmitzer L. 1996. Potential for modifying source±sink interactions through the genetic manipulation of carbohydrate metabolism. In: Zamski E, Schaffer AA, eds. Photoassimilate distribution in plants and crops: source±sink relationships. New York: Marcel Dekker Inc., 369±387. Konno Y, Vedvick T, Fitzmaurice L, Mirkov TE. 1993. Puri®cation, characterization and subcellular localization of soluble invertase from tomato fruit. Journal of Plant Physiology 141, 385±392. Lemoine R, KuÈhn C, Thiele N, Delrot S, Frommer WB. 1996. Antisense inhibition of the sucrose transporter in potato: effects on amount and activity. Plant, Cell and Environment 19, 1124±1131. Lin TP, Caspar T, Somerville C, Preiss J. 1988. A starchde®cient mutant of Arabidopsis thaliana with low ADPglucose pyrophosphorylase activity lacks one of two subunits of the enzyme. Plant Physiology 88, 1175±1181. Milner ID, Ho LC, Hall JL. 1995. Properties of proton and sugar-transport at the tonoplast of tomato (Lycopersicum esculemtum) fruit. Physiologia Plantarum 94, 399±410. Miron C, Schaffer AA. 1991. Sucrose phosphate synthase, sucrose synthase and invertase activities in developing fruits of Lycopersicon esculentum Mill. Plant Physiology 95, 623±627. MuÈller-RoÈber B, Sonnewald U, Willmitzer L. 1992. Inhibition of the ADP-glucose pyrophosphorylase in transgenic potatoes leads to sugar-storing tubers and in¯uences tuber formation and expression of tuber storage proteins. EMBO Journal 11, 1229±1238. Nadkarni MA, Nair CKK, Pandey VN, Pradhan DS. 1992. Raf®nose metabolism and novel mode of sucrose synthesis in Corynebacterium murisepticum. Journal of General and Applied Microbiology 38, 533±539. Neuhaus HE, Henrichs G, Scheibe R. 1993. Characterization of glucose-6-phosphate incorporation into starch by isolated intact cauli¯ower-bud plastids. Plant Physiology 101, 573±578. Neuhaus HE, Stitt M. 1990. Control analysis of photosynthate partitioning: impact of reduced activity of ADPG-glucose pyrophosphorylase or plastid phosphoglucomutase on the ¯uxes to starch and sucrose in Arabidopsis thaliana (L.) Heynh. Planta 182, 445±454. Nguyen-Quoc B, N'tchobo H, Foyer CH, Yelle S. 1999. Overexpression of sucrose phosphate synthase increase sucrose unloading in transformed tomato fruit. Journal of Experimental Botany 50, 785±791. Niland S, Schmitz K. 1995. Sugar transport in the storage tubers of Starchys sieboldii Miq.: evidence for symplastic unloading and starchyos uptake into storage vacuoles by an Hq antoport mechanism. Botanica Acta 108, 24±33. N'tchobo H. 1998. Sucrose unloading in tomato fruits. II. Subcellular distribution of acid invertase and possible roles in sucrose turnover and hexose storage in tomato fruit. PhD thesis, Laval University, 87±118. N'tchobo H, Dali N, Nguyen-Quoc B, Foyer CH, Yelle S. 1999. Starch synthesis in tomato remains constant throughout fruit development and is dependent on sucrose supply and sucrose activity. Journal of Experimental Botany 50, 1457±1463. Okita T. 1992. It there an alternative for starch synthesis? Plant Physiology 100, 560±564. Ohyama A, Ito H, Sato T, Nishimura S, Imai T, Hirai M. 1995. Supression of acid invertase activity by antisense RNA Sucrose metabolism in tomato fruit 889 modi®es the sugar composition of tomato fruits. Plant and Cell Physiology 36, 369±376. Patrick JW. 1990. Sieve element unloading: cellular pathway, mechanism and control. Physiologia Plantarum 78, 298±308. Pollock CJ. 1986. Fructan and the metabolism of sucrose in vascular plants. New Phytologist 104, 1±24. Preiss J. 1988. Biosynthesis of starch and its degradation. In: Preiss J, ed. The biochemistry of plants, Vol. 14. New York: Academic Press, 181±254. Preiss J. 1991. Starch biosynthesis and its regulation. In: Mi¯in J, ed. Oxford surveys of plant molecular and cell biology, Vol. 7. New York: Academic Press, 59±114. Prols R, Goetz M. Roitsch T. 2000. Differential expression of extracellular invertases in sink tissues and regulation by brassinosteroids. Botanikertagung Abstract P22-15, 339. Riesmeier JW, Hirner B, Frommer WB. 1993. Potato sucrose transporter expression in minor veins indicates a role in phloem loading. The Plant Cell 5, 1591±1598. Riesmeier JW, Willmitzer L, Frommer WB. 1992. Isolation and characterization of a sucrose carrier cDNA from spinach by function expression in yeast. EMBO Journal 11, 4705±4713. Ruan Y-L, Patrick JW. 1995. The cellular pathway of postphloem sugar transport in developing tomato fruit. Planta 196, 434±444. Sander A, Krausgrill S, Greiner S, Weil M, Rausch T. 1996. Sucrose protects cell wall invertase but not vacuolar invertase against proteinaceous inhibitors. FEBS Letters 385, 171±175. Sato T, Iwatsubo T, Tkahashi M, Nakagawa H, Ogura N, Mori H. 1993. Intercellular localization of acid invertase in tomato fruit and molecular cloning of cDNA for the enzyme. Plant and Cell Physiology 34, 263±269. Sauer N, Stolz J. 1994. SUC1 and SUC2: two sucrose transporters from Arabidopsis thaliana: expression and characterization in baker's yeast and identi®cation of the histidine tagged protein. The Plant Journal 6, 67±77. Sauer N, Tanner W. 1989. The hexose carrier from Chlorella cDNA cloning of a eucaryotic Hq-cotransporter. FEBS Letters 259, 43±46. Scholes JD, Bundock N, Wilde R, Rolfe SA. 1996. The impact of reduced vacuolar invertase activity on the photosynthetic and carbohydrate metabolism of tomato. Planta 200, 265±272. Sonnewald U, Brauer M, von Schaewen, Stitt M, Willmitzer L. 1991. Transgenic tobacco plants expressing yeast-derived invertase in either the cytosol, vacuole or apoplast: a powerful tool for studying sucrose metabolism and sinkusource interactions. The Plant Journal 17, 649±658. Sonnewald U, Lerchl J, Zrenner R, Fromer W. 1994. Manipulation of sink±source relation in transgenic plant. Plant, Cell and Environment 17, 649±658. Stark DM, Timerman KP, Barry GF, Preiss J, Kishore GM. 1992. Regulation of the amount of starch in plant tissues by ADP glucose pyrophosphorylase. Science 258, 287±292. Sun J, Loboda T, Sung S-JS, Black CCJ. 1992. Sucrose synthase in wild tomato, Lycopersicon chmielewskii, and tomato fruit sink strength. Plant Physiology 98, 1163±1169. Sweetlove LJ, Burrell MM, ap Rees T. 1996. Starch synthesis in tubers of transgenic potato (Solanum tuberosum) with increased ADPglucose pyrophosphorylase. Biochemistry Journal 320, 493±498. Sweetlove LJ, Muller-Rober B, Willmitzer L, Hill SA. 1999. The contribution of adenosine 59-diphosphoglucose pyrophosphorylase to the control of starch synthesis in potato tubers. Planta 209, 330±337. Tsai CY, Nelson OE. 1966. Starch-de®cient maize mutant lacking adenosine diphosphate glucose pyrophosphorylase activity. Science 151, 341±343. Viola R. 1996. Hexose metabolism in discs excised from developing potato (Solanum tuberosum L.) tubers: estimation of ¯uxes in vivo and evidence that fructokinase catalises a near rate-limiting reaction. Planta 198, 186±196. Viola R, Davis HV, Chudeck AR. 1991. Pathway of starch and sucrose biosynthesis in developing tuber of potato (Solanum tuberosum L.) and seed of faba bean (Vicia faba L.): elucidation by 13C-nuclear-magnetic-resonance spectroscopy. Planta 183, 202±208. Walker AJ, Ho LC. 1977. Carbon translocation in the tomato: carbon import and fruit growth. Annals of Botany 41, 813±823. Walker AJ, Ho LC, Baker DA. 1978. Carbon translocation in the tomato: pathway and the rate of translocation. Annals of Botany 42, 901±909. Wang F, Sanz A, Brenner ML, Smith A. 1993. Sucrose synthase, starch accumulation and tomato fruit sink strength. Plant Physiology 101, 321±327. Weise A, Barker L, Kuhn C, Lalonde S, Buscchmann H, Frommer WB, Ward JM. 2000. A new subfamily of sucrose transporters, SUT4, with low af®nityuhigh capacity localized in enucleate sieve elements of plants. The Plant Cell 12, 1345±1355. Wendler R, Veith R, Dancer J, Stitt M, Komor E. 1990. Sucrose storage in cell suspension culture of Sacharum sp. (sugarcane) is regulated by a cycle of synthesis and degradation. Planta 183, 31±39. Weber H, Heim U, Golombek S, Borisjuk L, Manteuffel R, Wobus U. 1998. Expression of a yeast-derived invertase in developing cotyledons of Vicia narbonensis alters the carbohydrate state and affects storage function. The Plant Journal 16, 163±172. Yelle S, Chetelat RT, Dorais M, DeVerna JW, Bennet AB. 1991. Sink metabolism in tomato fruit. IV. Genetic and biochemical analysis of sucrose accumulation. Plant Physiology 95, 1026±1035. Yelle S, Hewitt JD, Robinson NL, Damon S, Bennet AB. 1988. Sink metabolism in tomato fruit. III. Analysis of carbohydrate assimilation in a wild species. Plant Physiology 87, 737±740. Yin H, Wang MD, Svoboda K, Landick R, Block SM, Gelles J. 1995. Transcription against an applied force. Science 270, 1653±1657. Zrenner R, Salanoubat M, Willmitzer L, Sonnewald U. 1995. Evidence of the crucial role of sucrose synthase for sink strength using transgenic potato plants (Solanum tuberosum L.). The Plant Journal 7, 97±107.