Survey
* Your assessment is very important for improving the workof artificial intelligence, which forms the content of this project
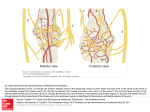
The Laryngoscope C 2016 The American Laryngological, V Rhinological and Otological Society, Inc. Are the Interarytenoid Muscles Supplied by Branches of Both the Recurrent and Superior Laryngeal Nerves? Ar an Pascual-Font*, PhD; Luis Cubillos*, MS; Teresa V azquez, PhD; Steve McHanwell, MD; Jose R. Sa~ nudo, MD; Eva Maranillo, MD Objectives/Hypothesis: It has been generally accepted that the branches of the internal branch of the superior laryngeal nerve to the interarytenoid muscle are exclusively sensory. However, some experimental studies have suggested that these branches may contain motor axons, and therefore that the interarytenoid muscle is supplied by both the superior and recurrent laryngeal nerves. The aim of this work was to determine whether motor axons to the interarytenoid muscles are present in both laryngeal nerves. Study Design: Basic research. Methods: Twelve human internal branches of the superior laryngeal nerve were dissected, and its branches to the interarytenoid muscle were removed and processed for choline-acetyltransferase immunohistochemistry, a method not used previously in studying the nerve fiber composition of the laryngeal nerves. Results: The internal branch of the superior laryngeal nerve divided into two to five branches to the interarytenoid muscle. All branches contained motor axons, with the proportion of motor axons varying from 6% to 31%. Conclusion: The present study confirms that the internal branch of the superior laryngeal nerve provides a motor innervation to the interarytenoid muscles. Key Words: Internal laryngeal nerve, internal branch of the superior laryngeal nerve, recurrent laryngeal nerve, superior laryngeal nerve, internal branch, ChAT, larynx, motor fibers. Level of Evidence: N/A. Laryngoscope, 126:1117–1122, 2016 INTRODUCTION In humans, the internal branch of the superior laryngeal nerve (ibSLN) divides into three terminal branches. The posterior branch, called the ramus communicans, runs over the interarytenoid muscle and divides into a variable number of branches that pierce it.1–3 Traditionally, it has been thought that the sole source of motor innervation to the interarytenoid muscles (oblique and transverse) is from the recurrent laryngeal nerve (RLN). However, these muscles also receive a variable number of branches from the ibSLN, which can be observed entering the muscular bellies. These branches have always been considered as being From the Department of Human Anatomy and Embryology I, School of Medicine, Complutense University of Madrid (A.P–F., L.C., T.V., J.R.S., E.M.), Madrid, Spain; and the School of Medical Education, Newcastle University (S.M.), Newcastle, United Kingdom Editor’s Note: This Manuscript was accepted for publication April 20, 2015. This work was supported by a grant FIS10–02721 (Fondo de Investigaciœn Sanitaria) of the institute of Health Carlos III (ISCIII) Institute of Health Carlos III (ISCIII) of the Spanish government and by funds obtained through postgraduate training courses by the UCM920547 Group. The authors have no other funding, financial relationships, or conflicts of interest to disclose. *Ar an Pascual-Font, and Luis Cubillos, contributed equally to this work. Send correspondence to Eva Maranillo, Departamento de Anatomia y Embriologia Humanas, Facultad de Medicina, Universidad Complutense de Madrid, Ciudad Universitaria s/n, 28040 Madrid, Spain. E-mail: [email protected] DOI: 10.1002/lary.25375 Laryngoscope 126: May 2016 exclusively sensory, either to the mucosa of the larynx or proprioceptive to the muscles themselves or to both structures.4–9 However, there have been several anatomical studies in humans that suggest the ibSLN may provide a secondary motor innervation to the interarytenoid muscles, and therefore that these muscles receive a dual motor innervation from both the RLN and the ibSLN1,10–13 (Table I). A variety of methods have been employed in attempts to determine the function of the nerve fibers in the ibSLN that enter the interarytenoid muscles. A significant number of studies have employed dissection either with or without prestaining of the nerve fibers using Sihler’s stain35–7,9–16 (Table I). Other studies have employed a variety of histological methods to determine the functional nature of these nerve fibers, which have included silver impregnation, gold chloride stain, Wrete impregnation, Romanes’ method, acetylcholinesterase stain, and horseradish peroxidase labeling7,8,10–12,17,18 (Table II). Other techniques that have been used are electromyography or studying the morphometry of the nerve.7,19–22 However, none of the studies employing the methods described above have produced results conclusive to establishing whether any of the ibSLN fibers that enter the interarytenoid muscles are motor in function. This article reports the results of a study employing an immunohistochemical method to investigate whether choline acetyltransferase (ChAT) is present in these ibSLN fibers. This technique selectively labels motor axons by using a primary antibody against ChAT, the Pascual-Font et al.: Neural Sources for the Interarytenoid Muscle 1117 TABLE I. Anatomical Studies on the Innervation of Interarytenoid Muscle by Internal Branch of the Superior Laryngeal Nerve (ibSLN) in Humans. Number of ibSLN Author (date) Number of Branches to Interarytenoid Method of Study Right Left Right Left Dilworth (1921)1 Dissection 33 33 1–2 1–2 Lemere (1932)5 Todd (1938)10 Dissection Dissection 12 96 12 96 Winckler (1948)6 Dissection Williams (1951)7 Vogel (1952)11 Dissection Dissection 30 16 30 16 Gupta et al. (1959)12 thi (1962)13 Re Dissection 25 25 Dissection Dissection Not specified 11 11 Rueger (1972)8 Not specified Not specified 3–8 2 Sensory 1 Motor Sensory Sensory 1 Motor 2 Not specified Not specified 1–2 Function Sensory Sensory Proprioception Sensory 1 Motor 1–2 Sensory 1 Motor Not specified 0–3 0–3 Sensory 1 Motor Sensory Sanders et al. (1993)14 Sihler’s stain 10 10 Mu et al. (1994)15 Sanders and Mu (1998)16 Sihler’s stain Sihler’s stain 10 5 10 5 2–4 2–3 Maranillo et al. (2005)3 Dissection 75 75 1–4 Olthoff et al. (2007)9 Dissection 30 30 Not specified 2–4 2–3 1–4 Multiple Not discernible Not discernible Not discernible Not discernible Sensory ibSLN 5 superior laryngeal nerve. enzyme responsible for synthesis of the neurotransmitter acetylcholine, which is the neurotransmitter released at the motor neuron terminals.23–25 MATERIALS AND METHODS Six human larynges (12 ibSLN) without known laryngeal pathology were obtained from necropsies (4 males and 2 females, age range from 69–93 years), from cadavers that were donated to the Human Anatomy and Embryology Department of the Faculty of Medicine, Complutense University of Madrid, Spain, in accordance with the legal procedures established for body donation in Spain. The larynges were fixed in 10% formalin and were microdissected by means of a surgical Zeiss-OPM1 microscope (Carl Zeiss AG, Oberkochen, Germany) using 43 to 63 magnification. The number and distribution of branches innervating each interarytenoid muscle were studied. The ibSLN was sampled at three sites. A segment of the posterior terminal branch of the ibSLN, the ramus communicans, and the collateral branches entering into the interarytenoid muscles were removed, embedded in paraffin, and serially sec- tioned at 10 lm. The segment of the posterior terminal branch of the ibSLN was next to its origin from the main nerve trunk. The piece of the ramus communicans was at the level of the superior third of the posterior cricoarytenoid muscle, and pieces of the collateral branches were immediately before the point at which they pierced the interarytenoid muscle. After treatment with sodium citrate buffer (10.2 mM, pH 6.1) for 20 minutes at 958C for antigen retrieval,26 the nerve sections were blocked with horse serum (Vector Laboratories, Peterborough, UK) (10%) and Bovine Serum Albumin (BSA) (Sigma-Aldrich, Steinheim, Germany) (3%) in Tris-phosphate buffer (TBS) with 0.3% in (Merck, Darmstadt, Germany) for 1 hour at 48C. They were incubated for 3 days with goat antiChAT primary polyclonal antibody (ChAT; 1/75; AB144P; Millipore), to identify motor axons. The primary antibody was diluted in TBS containing 0.3% Triton X-100, 5% serum. After washes with TBS containing 0.3% Triton X-100 (7 3 10 minutes), sections were incubated for 24 hours at 48C with biotin-conjugated horse anti-goat Immunoglobulin G antibody (Vector Laboratories), diluted 1:200 in TBS with 0.3% Triton X100. To visualize the presence of ChAT, the ABC-peroxidase kit TABLE II. A Summary of Results of Histological Studies on Innervation of Interarytenoid Muscle by ibSLN in Humans. Author (date) Todd (1938)10 Williams (1951)7 Vogel (1952)11 Method Number Interarytenoid Muscles End-Plates From ibSLN Function Gold chloride stain Wrete impregnation and Romanes’ method 7 5 Yes No Sensory 1 Motor Sensory Proprioception Gold chloride stain 19 Yes Sensory 1 Motor Gupta et al. (1959)12 Carey’s gold chloride method 2 Yes Sensory 1 Motor Rueger (1972)8 Freije et al. (1987)17 Gold chloride stain Acetylcholinesterase stain 6 4 No No Sensory Sensory Wustrow et al. (1988)18 Horseradish peroxidase Silver impregnation Acetylcholinesterase stain 4 No Sensory ibSLN 5 superior laryngeal nerve. Laryngoscope 126: May 2016 1118 Pascual-Font et al.: Neural Sources for the Interarytenoid Muscle TABLE III. Innervation of Interarytenoid Muscle From ibSLN (posterior branch and ramus communicans). Distribution by Sides and Sex. Larynx Sex Branches Right ibSLN Branches Left ibSLN Total Branches 1 Male 1 1 2 2 Male 2 3 5 3 4 Male Female 2 2 2 2 4 4 5 Male 2 1 3 6 Female 3 2 5 ibSLN 5 superior laryngeal nerve. Fig. 1. Posterior view of the larynx. The pharyngeal mucosa has been removed. White arrowhead: branches from ibSLN to interarytenoid muscle; black arrowhead: branches from RLN to internarytenoid muscle. ia 5 interarytenoid muscle; ibSLN 5 internal branch of the superior laryngeal nerve; pca 5 posterior cricoarytenoid muscle; pb 5 posterior branch of ibSLN; rc 5 ramus communicans (Galen’s loop); rln 5 recurrent laryngeal nerve. [Color figure can be viewed in the online issue, which is available at www.laryngoscope.com.] with Diaminobenzidine (Vector Laboratories) was used. Slides were dehydrated through a series of alcohols and mounted in DPX (Sigma Aldrich, St. Louis, MO). All of the samples were subjected to the immunohistochemical procedure in pairs, with one sample of each pair not incubated with the primary antibody to act as a negative control. The samples were photographed at 2003 to 4003, and analysis of the fibers was carried out manually with ImageJ 1.44p software (National Institutes of Health, Bethesda, MD). Each sample was quantified, taking as a reference the corresponding negative control of each pair to eliminate any nonspecific staining of nerve fibers. Nonmyelinated nerve fibers of less than 1.5 mm were excluded from the counts to ensure that type C fibers were not included in the final totals of nerve fibers counted.27 The mean and standard deviation (SD) of the number of ChAT-positive axons, as well as the total number of axons in the main trunk and in each branch, were calculated. RESULTS In humans, the ibSLN divides into three terminal branches. The posterior branch courses over the interarytenoid muscle and connects with the RLN over the surface of the posterior cricoarytenoid muscle via a branch called the ramus communicans (Fig. 1). The posterior branch and the ramus communicans on each side divded into a variable number of branches that entered the muscular belly (Fig. 1) (Table III). Laryngoscope 126: May 2016 The motor branch of the RLN to the interarytenoid muscle ascended deep to the belly of the posterior cricoarytenoid to reach the interarytenoid muscle at the level of the superior border of the cricoid cartilage (Fig. 1). ChAT-positive axons were observed in all the segments of the ibSLN that were examined (Table IV). These axons were distributed in a random way throughout all of the nerve branches that were studied (Fig. 2). The number of ChAT-positive axons that were presumptively identified as motor axons in each of the different branches of the ibSLN innervating the interarytenoid muscles can be summarized as follows: The posterior terminal branch of the ibSLN was found to contain an average of 290 6 225 (SD) ChAT-positive axons (Fig. 2A). The mean total number of axons in the nerve was 1967 6 995; therefore, these ChAT-positive axons represented 15% of the axonal population present in the nerve. The number of branches of the ibSLN varied from one to three, although it was only in a minority of specimens that three branches of the ibSLN were found to be entering the interarytenoid muscle. Consequently, the total number of axons in all three branches was calculated. The average total number of motor axons in those branches of the nerve was 342 6 159, representing 17% of the axonal population (Fig. 2C, D). The ramus communicans was found to contain an average of 127 6 101 ChAT-positive axons out of a total axonal population of 995 6 649 axons, representing 12% of the total axonal population (Fig. 2B). DISCUSSION The precise details of nerve supply of the interarytenoid muscles and how the motor supply to the muscle is distributed in humans is an aspect of the neuroanatomy of the larynx that remains unclear despite numerous attempts at resolving this issue. There are a number of experimental studies using animal models that have investigated the contributions of the recurrent and superior laryngeal nerves to the innervation of the intrinsic laryngeal muscles.28–30 However, these studies only examined the innervation of cricothyroid, thyroarytenoid, or posterior cricoarytenoid muscles. Furthermore, there are anatomical differences between the intrinsic laryngeal muscles in rat and mouse, for example, compared to humans. In the mouse, interarytenoid muscles are not present (although an Pascual-Font et al.: Neural Sources for the Interarytenoid Muscle 1119 TABLE IV. Distribution of Motor Axons in ibSLN. ibSLN PB B1 B2 B3 Total (B1 1 B2 1 B3) RC 1 right 5.90% (75/1271) 9.98% (53/531) – – 9.98% (53/531) 6.06% (20/330) 1 left 2 right 8.40% (78/929) 8.21% (168/2047) 14.43% (58/402) 8.80% (33/375) – 4.01% (16/399) – – 14.43% (58/402) 6.33% (49/774) 12.37% (24/194) 5.58% (60/1075) 9.81% (173/1763) 10.08% (286/2837) 11.42% (81/709) 10.17% (540/5309) not assessable 7.39% (180/2435) 5.37% (164/3056) 8.27% (123/1488) 12.82 (140/1092) – – 22.86% (469/2052) 31.01% (431/1390) 13.20% (334/2530) 8.33% (100/1200) 2 left 26.44% (87/329) 3 right 3 left 7.57% (207/2735) 8.37% (257/3069) 4 right 23.06% (712/3088) 25.33% (57/225) 16.20% (144/889) – 28.83% (352/1221) 4 left 5 right 23.84% (711/2982) 16.20% (353/2179) 13.53% (131/968) 17.14% (413/2409) 21.67% (91/420) 21.77% (133/611) – – 18.08% (546/3020) 7.33% (304/4148) 12.14% (75/618) 10.52% (100/951) 9.55% (127/1330) 5 left 21.61% (424/1962) 28.83% (352/731) – – 7.72% (303/3923) 20.39% (210/1030) 6 right 6 left 11.82 (121/1024) 14.64% (292/1994) 25.49% (157/616) 31.05% (227/504) 20.62% (214/1038) 30.96% (204/659) 24.62% (98/398) – 15.99% (222/1388) 18.04% (201/1114) 23.61% (265/1118) 13.56% (77/568) Mean 14.7% (290/1967) 16.4% (167/1228) 16.3% (150/1048) 18.0% (90/554)) 16.6% (342/2434) 12.3% (127/995) 6 SD 7.4% (225/995) 9.0% (118/1016) 8.4% (77/779) 9.3% (12/244) 8.9% (159/1566) 5.5% (101/649) B1–3 5 branches to interarytenoid muscle; ibSLN 5 superior laryngeal nerve; PB 5 posterior branch; RC 5 ramus communicans; SD 5 standard deviation. anatomical counterpart is present), whereas in the rat, an alar cricoarytenoid muscle and superior cricoarytenoid muscle can be found that are absent in humans.31,32 Also, there are always uncertainties in relation to precise muscle homologies between species, meaning that any direct comparisons between apparently similar muscles always have to be made with caution. There are also significant differences in the Fig. 2. Immunohistochemical labelling for the enzyme ChAT in axial sections of the nerves. The stain revealed the presence of motor axons in all the samples. (A) Posterior branch of the internal branch of the superior laryngeal nerve. (B) Ramus communicans (Galen’s loop). (C, D) Branches to the interarytenoid muscle. Arrowhead: an example of ChAT-positive axon (motor). Scale bars: 100 mm. ChAT 5choline acetyltransferase. [Color figure can be viewed in the online issue, which is available at www.laryngoscope.com.] Laryngoscope 126: May 2016 1120 Pascual-Font et al.: Neural Sources for the Interarytenoid Muscle composition of the laryngeal nerves between rat and human because afferent axons are absent from the RLN in the rat.33 The total number of branches that enter the interarytenoid muscle has been reported as varying from four to eight, with two branches contributed by the RLN and two to six branches contributed by the ibSLN or by the ramus communicans that joins the two nerves.3 Branches from RLN have been considered as exclusively motor, whereas branches from the ibSLN have been thought by some to be purely sensory4 or proprioceptive7,9 or by others to be mixed.1,10–13 A variety of methodologies have been employed in attempting to resolve this issue (Table I). Dissection, sometimes in combination with Sihler’s stain, has been extensively used by many authors, but we consider these methods to simply lack any degree of selectivity that would allow one to determine the functional character of axons that reach the interarytenoid muscle.14–16 The interpretation of the results of any dissection studies is further complicated by the fact of the existence of a complex neural plexus within the interarytenoid muscle, which receives contributions from both the RLN and the ibSLN. This means that it is simply not possible to be certain that any axons observed within the muscle itself have originated from the ibSLN or from the RLN.2 Studies using standard histological techniques,7,8,10–12 anterograde tracers,18 and histochemical methods17,18 also fail to provide conclusive evidence in favor of one or other hypothesis. Again, this is because of the above-mentioned existence of this complex neural plexus, which prevents the precise source of the axons within the muscle and ending as motor end plates to be identified as originating from one laryngeal nerve or the other (Table II). In the present study, axons within the branches of the ibSLN were stained using an immunohistochemical method to determine whether they contained ChATpositive axons. Significant numbers of ChAT-positive axons were seen in all segments of the ibSLN that entered the interarytenoid muscles. Somatic motor axons express ChAT, but ChAT expression is not confined to this functional category of axons. ChAT expression can also be seen in the preganglionic axons of the autonomic nervous system, both sympathetic and parasympathetic divisions; in all the postganglionic parasympathetic fibers; and in the postganglionic axons of the sympathetic nervous system innervating the sweat glands and blood vessels that provide a vascular supply to skeletal muscles.34–36 However, only large myelinated ChAT-positive axons within the branches of the ibSLN, which were characterized as Aa motor axons, were included in the counts. The smaller type C nonmyelinated fibers of the autonomic nervous system were excluded. Furthermore, it was considered highly unlikely that any postganglionic sympathetic fibers to sweat glands would be included in the counts because these are confined to skin, are not found in the mucosa of the larynx, and are also nonmyelinated. Thus, it seems possible to conclude that, using the criteria outlined above, only somatic motor axons were included in the counts of ChAT-positive axons. Laryngoscope 126: May 2016 The finding in this study of motor axons in branches of the ibSLN could also provide an explanation for a number of previously difficult-to-explain findings in cases of RLN palsies. In these cases, the reports of the presence of motor-unit potentials in denervated laryngeal muscles,37,38 the findings on the position of the vocal folds by neurotization of the adjacent muscles,39 and the phenomenon of synkinesis40,41 could all be attributed to the presence of motor axons in the ibSLN. CONCLUSION Based on our results and the considerations described above, it can be concluded that the ibSLN does contain a small but significant number of motor fibers distributed to the interarytenoid muscles. This dual innervation, previously described by other authors and confirmed in the present study, needs be taken into account for reinnervation and denervation procedures and to understand the consequences of recurrent laryngeal nerve damage. BIBLIOGRAPHY 1. Dilworth TF. The Nerves of the Human Larynx. J Anat 1921;56:48–52. 2. Sanudo JR, Maranillo E, Leon X, Mirapeix RM, Orus C, Quer M. An anatomical study of anastomoses between the laryngeal nerves. Laryngoscope 1999;109:983–987. 3. Maranillo E, Leon X, Orus C, Quer M, Sanudo JR. Variability in nerve patterns of the adductor muscle group supplied by the recurrent laryngeal nerve. Laryngoscope 2005;115:358–362. 4. Onodi A. Die Anatomie und Physiologie der Kehlkopfnerven: Mit erganzenden pathologischen Beitragen. Berlin, Germany: Verlag von Oscar Coblentz; 1902. (Cited by Rueger, 1972). 5. Lemere F. Innervation of the larynx. I. Innervation of the laryngeal muscles. Am J Anat 1932;51:417–437. 6. Winckler, G. Remarques sur l’innervation motrice et sensitive des muscles du larynx. Compt Rend Assn Anat 1948;55:424–428. 7. Williams AF. The nerve supply of the laryngeal muscles. J Laryngol Otol 1951;65:343–348. 8. Rueger RS. The superior laryngeal nerve and the interarytenoid muscle inhumans: an anatomical study. Laryngoscope 1972;82:2008–2031. 9. Olthoff A, Schiel R, Kruse E. The supraglottic nerve supply: an anatomic study with clinical implications. Laryngoscope 2007;117:1930–1933. 10. Todd HH Jr. Innervation of the Larynx. Unpublished master’s thesis. Chicago, IL: Loyola University; 1938: 1–27. 11. Vogel PH. The innervation of the larynx of man and the dog. Am J Anat 1952;90:427–447. 12. Gupta BC, Misra RN, Narayan D. Innervation of the human larynx. Indian J Otolaryngol 1959;11:199–212. 13. Rethi A. L’innervation du larynx. Acta Otorinolaryngol Iber Am 1962;13: 585–595. 14. Sanders I, Wu BL, Mu L, Li Y, Biller HF. The innervation of the human larynx. Arch Otolaryngol Head Neck Surg 1993;119:934–939. 15. Mu L, Sanders I, Wu BL, Biller HF. The intramuscular innervation of the human interarytenoid muscle. Laryngoscope 1994;104:33–39. 16. Sanders I, Mu L. Anatomy of the human internal superior laryngeal nerve. Anat Rec 1998;252:646–656. 17. Freije J, Malmgren LT, Gacek RR. Motor end-plate distribution in the human interarytenoid muscle. Arch Otolaryngol Head Neck Surg 1987; 113:63–68. 18. Wustrow TP, Martin F, Hollander H. [Auxiliary motor innervation of the laryngeal muscles via the internal branch of the superior laryngeal nerve]. Untersuchungen zur zusatzlichen motorischen Innervationder Kehlkopfmuskulatur iiber den Ramus internusdes Nervus laryngeus superior. [Article in German]. Laryngol Rhinol Otol (Stuttg) 1988;67: 275–281. 19. Ogura JH, Lam RL. Anatomical and physiological correlations on stimulating the human superior laryngeal nerve. Laryngoscope 1953;63:947– 959. 20. Tomasch HJ, Britton WA. A fibre analysis of the laryngeal nerve-supply in man. Acta Anat (Basel) 1955;23:386–398. 21. Dedo HH. The paralyzed larynx: an electromyographic study in dogs and humans. Laryngoscope 1970;80:1455–1517. 22. Martin-Oviedo C, Maranillo E, Lowy-Benoliel A, et al. Functional role of human laryngeal nerve connections. Laryngoscope 2011;121: 2338–2343. Pascual-Font et al.: Neural Sources for the Interarytenoid Muscle 1121 23. Aquilonius SM, Eckernas SA, Gillberg PG. Topographical localization of choline acetyltransferase within the human spinal cord and a comparison with some other species. Brain Res 1981;211:329–340. 24. Oda Y, Imai S, Nakanishi I, Ichikawa T, Deguchi T. Immunohistochemical study on choline acetyltransferase in the spinal cord of patients with amyotrophic lateral sclerosis. Pathol Int 1995;45:933–939. 25. Oda Y. Choline acetyltransferase: the structure, distribution and pathologic changes in the central nervous system. Pathol Int 1999;49:921–937. 26. Badia J, Boretius T, Pascual-Font A, Udina E, Stieglitz T, Navarro X. Biocompatibility of chronically implanted transverse intrafascicular multichannel electrode (TIME) in the rat sciatic nerve. IEEE Trans Biomed Eng 2011;58:192–201. 27. Gasser HS. The classification of nerve fibers. Ohio J Sci 1941;41:145–159. 28. Roubeau B, Lefaucheur JP, Moine A, Lacau St Guily J. Asymmetry of the laryngeal reflex responses to superior laryngeal nerve stimulation unrelated to the length of the recurrent nerves in the porcine model. Acta Otolaryngol 1998;118:882–886. 29. Hydman J, Mattsson P. Collateral reinnervation by the superior laryngeal nerve after recurrent laryngeal nerve injury. Muscle Nerve 2008;38:1280–1289. 30. Bjorck G, Margolin G, Maback GM, Persson JK, Mattsson P, Hydman J. New animal model for assessment of functional laryngeal motor innervation. Ann Otol Rhinol Laryngol 2012;121:695–699. 31. Inagi K, Schultz E, Ford CN. An anatomic study of the rat larynx: establishing the rat model for neuromuscular function. Otolaryngol Head Neck Surg 1998;118:74–81. Laryngoscope 126: May 2016 1122 32. Thomas LB, Stemple JC, Andreatta RD, Andrade FH. Establishing a new animal model for the study of laryngeal biology and disease: an anatomic study of the mouse larynx. J Speech Lang Hear Res 2009;52: 802–811. 33. Pascual-Font A, Hernandez-Morato I, McHanwell S, et al. The central projections of the laryngeal nerves in the rat. J Anat 2011;219:217–228. 34. Bolme P, Novotny J, Uvnas B, Peter G. Wright PG. Species distribution of sympathetic cholinergic vasodilator nerves in skeletal muscle. Acta Physiol Scand 1970;78:60–64. 35. Crossman AR, Neary D. Neuroanatomy. 2nd Ed. New York, NY: Churchill Livingstone; 2000: 47. 36. Kandel ER, Schwartz JH, Jessell TM. Principles of Neural Science. 4th Ed. New York, NY: McGraw-Hill; 2000: 282. 37. Hirano M, Nozoe I, Shin T, Maeyama T. Electromyography for laryngeal paralysis. In: Hirano M, Kirchner JA, Bless DM eds. Neurolaryngology: Recent Advances. Boston, MA: College-Hill Press; 1987: 232–248. 38. Wood GE. Configuration of the glottis in laryngeal paralysis. I: clinical study. Laryngoscope 1993;103:1227–1234. 39. Lewis WS, Crumley RL, Pitcock JK, Blanks RHI. Does intralaryngeal motor nerve sprouting occur following unilateral recurrent laryngeal nerve paralysis? Laryngoscope 1991;101: 1259–1263. 40. Crumley RL. Experiments in laryngeal reinnervation. Laryngoscope 1982; 92(suppl 30):1–27. 41. Crumley RL. Laryngeal synkinesis revisited. Ann Oto Rhinol Laryngol 2000;109:365–371. Pascual-Font et al.: Neural Sources for the Interarytenoid Muscle