Survey
* Your assessment is very important for improving the workof artificial intelligence, which forms the content of this project
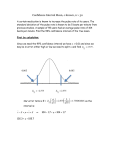
Am J Physiol Heart Circ Physiol 280: H1368–H1375, 2001. Coupling interval from slow to tachycardiac pacing decides sustained alternans pattern SHUNSUKE SUZUKI,1,2 JUNICHI ARAKI,1 YUMIKO DOI,1,2 WASO FUJINAKA,1,2 HITOSHI MINAMI,1 GENTARO IRIBE,1 SATOSHI MOHRI,1 JUICHIRO SHIMIZU,1 MASAHISA HIRAKAWA,2 AND HIROYUKI SUGA1,3 1 Department of Physiology II and 2Department of Anesthesiology and Resuscitology, Okayama University Medical School, Okayama 700-8558; and 3 National Cardiovascular Center Research Institute, Suita, Osaka 565-8565, Japan Received 27 December 1999; accepted in final form 26 October 2000 myocardium; contractility; calcium handling; arrhythmias MECHANICAL ALTERNANS OF THE HEART, either transient or sustained, consists of alternating strong and weak beats despite a regular beating rate. This phenomenon has interested investigators since it was first described in 1872 by Traube (33). The two mechanisms accounting for the phenomenon are alternations of end-diastolic volume (11, 15, 21) and cardiac contractility (5, 15, 19, 20). The former seems secondary to the latter (15, 19, 20). Cellular mechanisms, such as alternations of the action potential duration and intracellular Ca2⫹ handling via the sarcoplasmic reticulum, seem to cause the contractile alternans (15). Address for reprint requests and other correspondence: H. Suga, National Cardiovascular Center Research Institute, 5-7-1 Fujishirodai, Suita, Osaka 565-8565, Japan (E-mail: [email protected]). H1368 Various myocardial conditions are known to affect the pattern and amplitude of the mechanical alternans. Cardiac anoxia, ischemia (27, 34), tachycardia (4), and hypothermia (16) cause or intensify sustained mechanical alternans. Administration of epinephrine (4), caffeine (12, 13, 25), and ryanodine (13) diminish or eliminate the alternans. On the other hand, the same alternans pattern reappeared even after a premature or delayed beat interrupting the sustained alternans (17, 29). These results have yielded a general view that the pattern and amplitude of sustained mechanical alternans would change with cardiac contractile conditions at the same beating rate. However, we accidentally observed that the interbeat alternation pattern and amplitude of the sustained contractile alternans induced by tachycardiac pacing varied considerably with a single coupling beat interval between the same slow and tachycardiac pacing periods. Intriguingly, sustained contractile alternans even disappeared after a specific coupling interval. No literature has documented these observations. Therefore, we investigated the relationship between the coupling interval and the interbeat alternation pattern and amplitude of sustained contractile alternans in the excised, cross-circulated canine heart. We produced sustained contractile alternans by an abrupt increase in regular pacing rate in normal canine hearts. Our results showed that 1) the alternans pattern and amplitude changed markedly with the coupling interval in all of the hearts, 2) the alternans disappeared at 1–3 specific coupling intervals in each heart, and 3) the even- and odd-numbered order of strong and weak beats counted from the coupling interval reversed across these specific coupling intervals. These findings indicate that a more severe sustained contractile alternans under a given tachycardiac pacing is not always indicative of a worse cardiac contractile condition. The costs of publication of this article were defrayed in part by the payment of page charges. The article must therefore be hereby marked ‘‘advertisement’’ in accordance with 18 U.S.C. Section 1734 solely to indicate this fact. 0363-6135/01 $5.00 Copyright © 2001 the American Physiological Society http://www.ajpheart.org Downloaded from http://ajpheart.physiology.org/ by 10.220.33.3 on May 11, 2017 Suzuki, Shunsuke, Junichi Araki, Yumiko Doi, Waso Fujinaka, Hitoshi Minami, Gentaro Iribe, Satoshi Mohri, Juichiro Shimizu, Masahisa Hirakawa, and Hiroyuki Suga. Coupling interval from slow to tachycardiac pacing decides sustained alternans pattern. Am J Physiol Heart Circ Physiol 280: H1368–H1375, 2001.—We discovered that the coupling beat interval from a slow to a tachycardiac pacing period considerably affected the pattern of the beat-to-beat alternation of the tachycardia-induced sustained contractile alternans. We analyzed the relationship between the coupling interval and the pattern and amplitude of the alternans in the isovolumic left ventricle of canine blood-perfused hearts. The alternans pattern and amplitude varied transiently over the first 30–50 beats and became gradually stable over the first minute in all 12 hearts. We discovered that stable alternans, even under the same tachycardiac pacing, had three different strong-weak beat patterns depending on the coupling interval. A relatively short coupling interval produced a representative sustained alternans of the strong and weak beats. A relatively long coupling interval produced a similar sustained alternans but in a reversed order of even- and odd-numbered beats counted from the coupling interval. However, sustained alternans disappeared after 1–3 specific coupling intervals. We conclude that ventricular pacing rate does not solely determine the pattern and amplitude of sustained contractile alternans induced by tachycardia. COUPLING INTERVAL AND SUSTAINED ALTERNANS METHODS mounted on a rigid connector was fitted into the LV, and the connector was secured at the mitral annulus. LV pressure was measured with a miniature pressure gauge (model P-7, Konigsberg Instruments; Pasadena, CA) inside the apical end of the balloon and processed with a direct current strain amplifier. The balloon, primed with water without air bubbles, was connected to our custom-made volume servo pump (AR-Brown; Tokyo, Japan). LV volume was accurately controlled and precisely measured with the servo pump. LV epicardial electrocardiogram (ECG) was recorded with a pair of screw-in electrodes to trigger data acquisition and identify the onset of contraction. We monitored and maintained cardiac temperature in an acrylic box by warming the coiled portion of the arterial cross-circulation tube in a thermostat bath. In our preliminary experiment, we needed a tachycardiac pacing rate of 250–300 beats/min to produce sustained contractile alternans at 36–37°C. This suggests that the heart we prepared was reasonably physiological (34). This condition, however, rapidly deteriorated the contractility of the heart, probably because the tachycardia-induced increase in oxygen demand exhausted the coronary reserve (31). Therefore, we kept cardiac temperature slightly hypothermic (34.2 ⫾ 1.0°C) to produce stable sustained contractile alternans at a pacing rate under 250 beats/min in the present study. Mean systemic arterial blood pressure of the support dog (118 ⫾ 16 mmHg) served as coronary perfusion pressure. It was maintained stable in each experiment by slowly transfusing whole blood reserved from the heart donor dog or by infusing 6% hydroxyethyl starch solution and by continuously infusing methoxamine (5–30 mg/h) as needed. We measured repeatedly arterial pH (7.42 ⫾ 0.07), PO2 (131 ⫾ 32 mmHg), and PCO2 (31 ⫾ 8 mmHg) of the support dog and normalized them with supplemental oxygen and intravenous NaHCO3 as needed. Pacing stimuli. We programmed the pacing stimuli with the use of LabView, version 3.1 (National Instruments), on a Power Mac computer, which controlled a stimulator with an analog-to-digital converter (Lab-NB, National Instruments). Figure 1 shows two representative set of pacing stimuli in one heart. We provided a priming period of regular slow pacing stimuli at intervals of 500 ms (⬇120 beats/min) in 9 Fig. 1. Sustained contractile alternans under the same tachycardiac pacing after two different coupling intervals (CI) after the priming beats at the same slow pacing rate. Top, pacing stimuli; middle, electrocardiagram (ECG); and bottom, left ventricular pressure (LVP). A and B: intervals of the slow (L) and the tachycardiac (H) stimuli fixed at 500 and 300 ms, respectively. CI was 400 (A) and 600 (B) ms. o and e, odd- and even-numbered beats, respectively, counted after CI. Downloaded from http://ajpheart.physiology.org/ by 10.220.33.3 on May 11, 2017 Surgical preparation. We performed the present experiments in the canine excised, cross-circulated (blood perfused) heart preparation that we have been using consistently in our laboratory (3, 14, 18, 24, 28, 31). We conducted them in conformity with the “Guiding Principles for Research Involving Animals and Human Beings,” endorsed by the American Physiological Society. The surgical procedures were described in detail elsewhere (14, 18, 24). Briefly, two mongrel dogs (9–23 kg body wt) were anesthetized with pentobarbital sodium (25 mg/kg iv) after premedication with ketamine hydrochloride (20 mg/kg im). Anesthesia was maintained by fentanyl (200–250 g/h iv). The dogs were intubated and artificially ventilated and were then heparinized (10,000 units iv per dog). We used the larger dog (18 ⫾ 3 kg body wt; means ⫾ SD) as the metabolic supporter. Its common carotid arteries and right external jugular vein were cannulated and connected to the arterial and venous cross-circulation tubes, respectively. The chest of the smaller dog (12 ⫾ 2 kg body wt) was opened midsternally, and the heart was used as a donor. The arterial and venous cross-circulation tubes were cannulated into the left subclavian artery and the right ventricle through the right atrial appendage, respectively, of the donor dog. In each of the 12 experiments, the heart-lung section was isolated from the systemic and pulmonary circulation by ligating the azygos vein, descending aorta, inferior vena cava, brachiocephalic artery, superior vena cava, and bilateral pulmonary hili. The beating heart, supported by cross circulation, was then excised from the chest. Coronary perfusion of the excised heart was never interrupted during the preparation. We gave indomethacin (5–20 mg iv; donated by Banyu Pharmaceutical; Tokyo, Japan) to the support dog to prevent systemic hypotension occasionally elicited by blood cross circulation (18, 24). The left atrium was opened, and all of the left ventricular (LV) chordae tendineae were cut. Complete atrioventricular block was made by electrical ablation or by injecting 40% formaldehyde through the right atrium into the region of the His bundle (30). Electrical pacing was performed with a bipolar electrode placed on the upper interventricular septum. A thin latex balloon (unstressed volume, ⬃50 ml) H1369 H1370 COUPLING INTERVAL AND SUSTAINED ALTERNANS reasonably good (8.3 ⫾ 3.8 mmHg 䡠 ml⫺1 䡠 100 g⫺1) (14, 24, 28, 31, 32). Here, Emax was calculated as LV peak isovolumic pressure divided by LV volume minus the initial volume obtained as the LV volume at which peak isovolumic pressure was zero (31, 32). We confirmed occasionally that the sustained contractile alternans continued stably with the same pattern and amplitude of interbeat alternation over 10 min. The valley, or end-diastolic pressure, also alternated during the contractile alternans. The higher valley pressure followed the higher peak pressure and the lower valley pressure followed the lower peak pressure. Coupling interval and alternans. Figure 2 shows representative relationships between the coupling interval and the peak isovolumic pressures of the strong and weak beats in stable sustained contractile alternans. LV contractilities of the alternans beats were proportional to these peak pressures because LV volume was constant. Figure 2A shows that both peak and valley pressures of the alternans beats changed sensitively with coupling interval. However, the alternans of both peak and valley pressures completely or almost disappeared at coupling intervals of 210, 300, and 600 RESULTS Sustained alternans. Figure 1 shows simultaneous tracings of LV pacing stimuli, ECG, and isovolumic pressure in two representative runs in one heart. These two runs had the same set of the slow pacing interval (500 ms) for the priming period and the tachycardiac pacing interval (300 ms) but two different coupling beat intervals (400 and 600 ms). Under tachycardia, the contractile alternans changed transiently over the first 10–15 s or 30–50 beats. Thereafter, the strong and weak alternation pattern and amplitude of the peak isovolumic pressure alternans and hence contractile alternans became gradually stable over 1 min. The peak pressure alternated remarkably during the initial transient and successive stable periods of tachycardiac pacing in Fig. 1A, whereas it stopped to alternate after the initial transient period of tachycardiac pacing in Fig. 1B. Similar results were obtained in all of the hearts. The index of LV contractility, Emax, was Fig. 2. Relationship between CI and the alternating peak and valley pressures of the strong and weak beats of the sustained contractile alternans. F and }, Peak and valley pressures, respectively, of even-numbered beats counted after CI. E and {, Odd-numbered beats. Vertical dashed lines on the right indicate the slow pacing interval (A and B, 450 ms); those on the left indicate the tachycardiac pacing interval (A, 280; B, 330 ms). The arrows on the abscissas indicate the specific CI. A and B correspond to type I and type II of sustained contractile alternans. Downloaded from http://ajpheart.physiology.org/ by 10.220.33.3 on May 11, 2017 hearts and 450 ms (133 beats/min) in the other 3 hearts. We then provided a tachycardiac period of regular pacing stimuli at shorter intervals of 283 ⫾ 24 ms (⬇213 ⫾ 18 beats/min) to generate sustained contractile alternans. We provided a variable coupling interval between the slow priming and tachycardiac periods as a sole independent variable. We performed similar runs only by changing the coupling interval in steps of 20–50 ms from 200 to 600 ms in each heart. Experimental protocol. We used isovolumic LV contractions throughout this study. Our preliminary experiment showed that the LV with a large isovolumic volume could not maintain stable sustained contractile alternans. We eliminated this problem by keeping LV isovolumic volume at a midrange volume of 12.3 ⫾ 2.4 ml (89 ⫾ 18 g wt), where LV peak isovolumic pressure was ⬍100 mmHg. We continuously monitored LV pressure through the slow and tachycardiac pacing periods and recorded it during the last several slow beats, and both the transient and stable sustained contractile alternans beats for 1–1.5 min. We then returned to the priming period with slow pacing until LV contractility became stable. Each experiment was completed within a few hours while the condition of the preparation was stable. In three experiments, we repeated the entire protocol to check the reproducibility of the results. Monophasic action potential. Monophasic action potential (AP) was recorded simultaneously with a contact electrode catheter (7) (Langendorff probe, Boston Scientific) on the anterolateral LV surface near its obtuse margin in three hearts. We measured the duration of the monophasic AP duration at the 90% repolarization (APD90) from the onset of the steep upstroke to the 90% repolarization level. Here, 100% was defined as the height of the monophasic AP from its diastolic baseline to the crest of its plateau (10). We measured APD90 in six consecutive beats during stable sustained contractile alternans. The six APD90 values were averaged for three strong and three weak beats separately. Data analyses. LV pressure, volume, ECG, and APD signals were digitized at 2- or 4-ms intervals with an analog-todigital converter (Lab-NB, National Instruments), displayed, and stored on the hard drive of a computer. Statistics. Data were presented as means ⫾ SD. Comparisons of the parameters between the strong and weak beats were made by Student’s paired t-test. A value of P ⬍ 0.05 was considered significant. COUPLING INTERVAL AND SUSTAINED ALTERNANS Table 1. Relationship between the number of the specific coupling intervals and the type of alternation of the peak and valley pressures of the strong and weak beats No. of Specific Coupling Intervals Type I Type II Total 0 1 2 3 Total 0 0 0 1 2 3 7 0 7 2 0 2 10 2 12 Values are means ⫾ SD for 2 dogs. Type I corresponds to Fig. 2A. Type II corresponds to Fig. 2B. Specific coupling interval means the coupling interval at which the alternation disappeared despite the tachycardiac pacing. Fig. 3. A–F: transition from the transient to sustained contractile alternans after various CI in one left ventricle. Left: LVP from slow beats to the 50th beat from the first tachycardiac beat in slow-speed recording. Right: LVP of the 201st and 202nd beats in high-speed recording. L, slow beat interval; H, tachycardiac pacing interval. Fig. 2B. Ten of the twelve hearts belonged to type I, and the other two belonged to type II. The most frequent type I case had two specific coupling intervals between 200 and 600 ms. We could not find any particular factor to determine these types. As for the valley pressures, the higher valley pressure always followed the higher peak pressure; the lower valley pressure always followed the lower peak pressure. When the peak pressure alternans disappeared at the specific coupling intervals, the valley pressure alternans also disappeared. Transition to sustained contractile alternans. Figure 3 shows the time course of the peak and valley pressure alternans from the transient to sustained phases while the coupling interval was increasing in one heart. Regardless of the coupling intervals, both peak and valley pressure alternans always started immediately after the coupling interval. The even and odd order of strong and weak beats in the transient phase of contractile alternans was retained, while the alternans became gradually and finally stable by ⬃1 min of the tachycardiac pacing. As the coupling interval increased, the strong and weak peak pressure alternans in the sustained period Downloaded from http://ajpheart.physiology.org/ by 10.220.33.3 on May 11, 2017 ms (arrows on the abscissa). Furthermore, the order of the strong and weak beats counted from the coupling interval reversed across a coupling interval of 300 ms. Above 300 ms, the odd-numbered beats were stronger and the even-numbered beats were weaker. Below 300 ms, the odd-numbered beats were weaker and the even-numbered beats were stronger. Figure 2B shows that the alternans amplitude of both peak and valley pressures suddenly diminished at a coupling interval of 375 ms (arrow) and the alternans order reversed across this coupling interval. At the other coupling intervals, the alternans amplitude of both peak and valley pressures remained little changed with the coupling interval. The alternans amplitude remained little changed, particularly between 250 and 360 ms. These results in Fig. 2 show the existence of at least one specific coupling interval in each heart, at which the amplitude of sustained contractile alternans totally or almost disappeared and across which the alternans order of strong and weak beats reversed. Such a specific coupling interval existed between the slow priming and tachycardiac pacing intervals, which are indicated by the two dashed lines in Fig. 2. We obtained similar results in all of the hearts. The order of strong and weak beats at the longer or shorter interval than a specific coupling interval was uncertain among hearts. Unless we counted carefully the beat number after the coupling interval, we could not distinguish the sustained contractile alternans of similar patterns and amplitudes at longer and shorter intervals than the specific coupling interval. Table 1 summarizes the number of the specific coupling intervals and the incidence of different types of the relationship between the coupling interval and the alternation pattern in all of the hearts. We counted the number of the coupling intervals at which the alternans disappeared or the order of the strong and weak beats reversed. We classified these relationships into two types. Type I was that the alternans amplitude of both peak and valley pressures continuously changed between the specific coupling intervals as shown in Fig. 2A. Type II was that the alternans amplitude was relatively stable at all of the coupling intervals except around the specific coupling intervals as shown in H1371 H1372 COUPLING INTERVAL AND SUSTAINED ALTERNANS DISCUSSION New finding. The most important finding in this study is that the pattern and amplitude of the strong and weak contractions in sustained contractile alternans varied markedly with the coupling interval between the same set of slow priming and the tachycardiac pacing periods. Although previous investigators have produced sustained contractile alternans by abruptly increasing the pacing rate (4, 9, 25, 26, 34, 36), no one has recognized the importance of the coupling interval between the slow priming and tachycardiac pacing periods. We obtained the present finding while we maintained stable the cardiac contractile conditions that could affect the severity of sustained contractile alternans (4, 16, 27, 34). Ca2⫹ handling. The sustained contractile alternans must be the result of alternating contractility, because we fixed LV volume to exclude the Starling effect. We speculate that the contractility alternans reflects alternans of myocardial Ca2⫹ handling (6, 28, 35, 36). Even an extrasystole affects myocardial Ca2⫹ handling in successive beats as manifested by postextrasystolic restitution and potentiation (3, 6, 23, 28). These postextrasystolic effects involve two Ca2⫹ handling mechanisms. One is a change in transsarcolemmal Ca2⫹ influx in the extrasystole and its gradual recovery over successive beats (23, 28). The other is a change in Ca2⫹ recirculation (i.e., uptake and release) via the sarcoplasmic reticulum (6, 22, 28). However, these changes disappear after several postextrasystolic beats under pacing at a slow enough rate not to produce sustained contractile alternans (6, 28, 35, 36). However, sus- tained contractile alternans occurs by sufficiently tachycardiac pacing even in normal hearts (34, 36). One or both transsarcolemmal and sarcoplasmic Ca2⫹ handling mechanisms seem responsible for the sustained contractile alternans that we observed. Because there was no significant alternation of APD90, alternation of the monophasic AP, and hence transsarcolemmal Ca2⫹ influx, are not likely to be a main cause of the sustained contractile alternans in the present study. In intact hearts, a Ca2⫹ channel blockade, verapamil, suppressed APD alternans but kept LV pressure alternans (12). This suggests that sustained contractile alternans require sarcoplasmic reticulum Ca2⫹ handling, but not always transsarcolemmal Ca2⫹ influx alternans. This is compatible with the finding of no significant APD90 alternans during the sustained contractile alternans in the present study. We suspect a mechanism to exist by which even a single coupling interval preceding the tachycardiac pacing decides the pattern and amplitude of the sustained contractile alternans. Even a single coupling interval affects the excitation-contraction coupling and the contraction of the several postextrasystolic tachycardiac beats. This effect in turn may continuously affect the subsequent sustained contractile alternans. As shown in Figs. 1 and 3, transient contractile alternans began tachycardiac pacing. The alternans waxed and waned for the initial 10–15 s or 30–50 beats. The greater amplitude the transient contractile alternans had, the greater amplitude the stable contractile alternans had. At a specific coupling interval, the amplitudes of both transient and sustained contractile alternans were smaller than at other coupling intervals. This suggests that the alternans amplitude initiated on the tachycardiac pacing is carried over to the successive alternans beats without fading out. This maintenance means that a mechanism for a stronger beat follows a weaker beat, despite the regular beat intervals under tachycardia. This mechanism itself is essentially what maintains sustained mechanical alternans under tachycardia in normal hearts (34). Simulation. We examined whether any sustainedcontractile-alternans model on the basis of myocardial Ca2⫹ handling in the literature could simulate our present finding. We first used Adler et al.’s model (1, 2). This model incorporated two Ca2⫹ handling mechanisms (1, 2). One is a Ca2⫹ release to myofilaments on depolarization as a function of Ca2⫹ in a releasable terminal to affect only the subsequent beat. The other is a strong Ca2⫹ buffering capability of the sarcoplasmic reticulum to affect several subsequent beats. This model also incorporated transmembrane Ca2⫹ influx and efflux. The Adler model successfully simulated sustained contractile alternans with an abrupt increase in the beating rate, as shown in Fig. 4. Although a step increase in heart rate from 120 to 240 beats/min generated sustained contractile alternans, the pattern and amplitude of the contractile alternans remained identical despite varied coupling intervals (200–400 ms). Although we recognized that the order of strong Downloaded from http://ajpheart.physiology.org/ by 10.220.33.3 on May 11, 2017 first increased from Fig. 3, A–C, then decreased from Fig. 3, C and D, and increased again from Fig. 3, D–F. The alternans almost disappeared, as shown in Fig. 3 D, namely, at a specific coupling interval. Between Fig. 3, D and E, the order of the strong and weak beats reversed as exemplified in the odd- (o, 201st) and evennumbered (e, 202nd) beats. We obtained similar results in all of the hearts. The valley pressure alternans was largely proportional to the peak pressure alternans in the transient phase of alternans. However, as the peak pressure alternans became stable, the valley pressure alternans almost or completely disappeared, as shown in Fig. 3, A, D, and E, but remained a little in Fig. 3, B, C, and F. Therefore, a higher valley pressure did not always precede a proportionally weaker peak pressure in the next beat. Action potential duration. We found that monophasic AP showed little alternans during the sustained contractile alternans. The APD90 of the strong and weak beats recorded in stable sustained contractile alternans were 211 ⫾ 3 versus 212 ⫾ 3 ms (12 runs) in one heart, 210 ⫾ 6 versus 209 ⫾ 4 ms (8 runs) in another heart, and 192 ⫾ 5 versus 193 ⫾ 5 ms (13 runs) in a third heart. The difference in APD90 between strong and weak beats was not significant in these hearts (Student’s paired t-test, P ⬎ 0.05). COUPLING INTERVAL AND SUSTAINED ALTERNANS Fig. 4. Simulation in the model proposed by Adler et al. (Ref. 1, Eqs. 1–12). After the priming period of 500 ms pacing intervals, pacing intervals were shortened to 250 ms for the tachycardiac pacing with a variable CI (arrow). CI was 200 (top), 300 (middle), and 400 (bottom) ms. E and F, Odd-numbered and even-numbered beats, respectively, counted after the CI. Pattern and amplitude of alternation were unchanged for all CI. Fig. 5. Simulation in the model proposed by Freeman et al. (Ref. 8, 2⫹ APPENDIX). Each panel shows the beat-to-beat amounts of Ca in three compartments: sarcoplasm, uptake pool, and releasable pool. A: extrasystole (ES; 250 ms, top, and 300 ms, bottom) during regular beat intervals (400 ms). B: step increase from slow pacing intervals (400 ms) to tachycardiac intervals (300 ms) with a CI (250 ms, top, and 500 ms, bottom). The right-most beats show sarcoplasmic Ca2⫹ in the 201st and 202nd beats from the first beat after CI. to the assumed constancy of the total intracellular Ca2⫹ throughout the transient and steady-state phases of tachycardia. A new Ca2⫹ handling model, including an appropriate combination of the Adler et al. (1) and Freeman et al. (8) models, may account for the present finding. Development of such a model is beyond the scope of the present study, although such an effort is warranted. The Adler and Freeman models do not include alternating changes in end-diastolic (valley) pressure and Ca2⫹. Additional components like the end-diastolic pressure and Ca2⫹ alternans to these models may help simulate our observation of the valley pressure alternans accompanying the peak pressure (contractile) alternans. The Adler and Freeman models included neither alternating changes in Ca2⫹ sensitivity nor responsiveness of contractility. A recent study (37) suggests that there is an interbeat change in the relation between Ca2⫹ handling and contractility after an extrasystole. The addition of such a mechanism to the Adler and Freeman models might lead to a successful simulation, although we have not attempted this. Downloaded from http://ajpheart.physiology.org/ by 10.220.33.3 on May 11, 2017 and weak beats reversed between coupling intervals of 300 and 400 ms, no coupling interval existed to attenuate or eliminate the alternans. The Adler model could not simulate any diastolic pressure alternans. Freeman et al. (8) presented a different model to simulate sustained contractile alternans. They used three time constants to explain the Ca2⫹ movement between the three compartments, i.e., the sarcoplasm, a Ca2⫹ uptake pool, and a Ca2⫹ releasable pool (38). They assumed the Ca2⫹ uptake time constant to be inversely proportional to sarcoplasmic Ca2⫹ (35). However, they assumed no contribution of transsarcolemmal Ca2⫹ transport to the alternans, namely, constancy of the total intracellular Ca2⫹ among alternans beats. Figure 5 shows our simulation using this Freeman model. Figure 5A shows sustained contractile alternans generated even by a single extrasystole interrupting the continuous regular beats (400 ms). These extrasystoles after coupling intervals of 250 (upper) and 300 ms (lower) generated different but stable patterns of alternation of sarcoplasmic Ca2⫹. However, we were not able to produce this type of nontachycardiac sustained alternans in our heart preparation. Figure 5B shows our simulation of sustained contractile alternans during tachycardia in a protocol similar to our experiment. These coupling intervals of 250 and 500 ms generated sustained contractile alternans and the pattern and amplitude of sarcoplasmic Ca2⫹ alternans depend on the coupling interval. However, the alternans waxed gradually after both coupling intervals until the strong beat increased twofold of the regular beat and the weak beat faded out completely. Thus the Freeman model as it is cannot simulate our present observation. This model can neither yield any obvious end-diastolic Ca2⫹ alternans. This may be due H1373 H1374 COUPLING INTERVAL AND SUSTAINED ALTERNANS We thank Kimikazu Hosokawa for continuous animal care and Banyu Pharmaceutical for donating indomethacin. We also thank Dr. Terumasa Morita, Department of Cardiovascular Surgery, for surgical assistance. This study was supported by Grants-in-Aid for Scientific Research 07508003, 09470009, 10558136, 10770307, 10877006, and 12680832 from the Ministry of Education, Science, Sports and Culture, Cardiovascular Diseases Research Grant 11C-1 from the Ministry of Health and Welfare, a Cardiac Physiome grant from the Science and Technology Agency, and research grants from the Suzuken Memorial Foundation and the Vehicle Racing Commemorative Foundation, all of Japan. REFERENCES 1. Adler D, Wong AYK, Mahler Y, and Klassen GA. Model of calcium movements in the mammalian myocardium: intervalstrength relationship. J Theor Biol 113: 379–394, 1985. 2. Adler D, Wong AYK, and Mahler Y. Model of mechanical alternans in the mammalian myocardium. J Theor Biol 117: 563–577, 1985. 3. Araki J, Takaki M, Matsushita T, Matsubara H, and Suga H. Postextrasystolic transient contractile alternans in canine hearts. Heart Vessels 9: 241–248, 1994. 4. Badeer HS, Ryo UY, Gassner WF, Kass EJ, Cavaluzzi J, Gilbert JL, and Brooks CM. Factors affecting pulsus alternans in the rapidly driven heart and papillary muscle. Am J Physiol 213: 1095–1101, 1967. 5. Braveny P. The relation of alternating contractility of the heart to the inotropic effects of rhythm. Arch Int Physiol Biochem 72: 553–566, 1964. 6. Burkhoff D, Yue DT, Franz MR, Hunter WC, and Sagawa K. Mechanical restitution of isolated perfused canine left ventricles. Am J Physiol Heart Circ Physiol 246: H8–H16, 1984. 7. Franz MR, Burkhoff D, Spurgeon H, Weisfeldt ML, and Lakatta EG. In vitro validation of a new cardiac catheter technique for recording monophasic action potentials. Eur Heart J 7: 34–41, 1986. 8. Freeman GL, Widman LE, Campbell JM, and Colston JT. An evaluation of pulsus alternans in closed-chest dogs. Am J Physiol Heart Circ Physiol 262: H278–H284, 1992. 9. Gilbert JL, Janse MJ, Lu HH, Pinkston JO, and Brooks CM. Production and abolition of alternation in mechanical action of the ventricle. Am J Physiol 209: 945–950, 1965. 10. Gillis AM, Kulisz E, and Mathison HJ. Cardiac electrophysiological variables in blood-perfused and buffer-perfused, isolated, working rabbit heart. Am J Physiol Heart Circ Physiol 271: H784–H789, 1996. 11. Gleason WL and Braunwald E. Studies on Starling’s law of the heart. VI. Relationships between left ventricular end-diastolic volume and stroke volume in man with observations on the mechanism of pulsus alternans. Circulation 25: 841–848, 1962. 12. Hirayama Y, Saitoh H, Atarashi H, and Hayakawa H. Electrical and mechanical alternans in canine myocardium in vivo. Dependence on intracellular calcium cycling. Circulation 88: 2894–2902, 1993. 13. Kihara Y and Morgan JP. Abnormal Cai2⫹ handling is the primary cause of mechanical alternans: study in ferret ventricular muscles. Am J Physiol Heart Circ Physiol 261: H1746– H1755, 1991. 14. Kohno K, Takaki M, Ishioka K, Nakayama Y, Suzuki S, Araki J, Namba T, and Suga H. Effects of intracoronary fentanyl on left ventricular mechanoenergetics in the excised cross-circulated canine heart. Anesthesiology 87: 658–666, 1997. 15. Lab MJ and Seed WA. Pulsus alternans. Cardiovasc Res 27: 1407–1412, 1993. 16. Lu HH, Lange G, and Brooks CM. Comparative studies of electrical and mechanical alternation in heart cells. J Electrocardiol 1: 7–17, 1968. 17. Mahler Y and Rogel S. Interrelation between restitution timeconstant and alternating myocardial contractility in dogs. Clin Sci (Colch) 39: 625–639, 1970. 18. Matsubara H, Takaki M, Yasuhara S, Araki J, and Suga H. Logistic time constant of isovolumic relaxation pressure-time curve in the canine left ventricle. Better alternative to exponential time constant. Circulation 92: 2318–2326, 1995. 19. McGaughey MD, Maughan WL, Sunagawa K, and Sagawa K. Alternating contractility in pulsus alternans studied in the isolated canine heart. Circulation 71: 357–362, 1985. 20. Miller WP, Liedtke AJ, and Nellis SH. End-systolic pressurediameter relationships during pulsus alternans in intact pig hearts. Am J Physiol Heart Circ Physiol 250: H606–H611, 1986. 21. Mitchell JH, Sarnoff SJ, and Sonnenblick EH. The dynamics of pulsus alternans: alternating end-diastolic fiber length as a causative factor. J Clin Invest 42: 55–63, 1963. 22. Mohri S, Araki J, Imaoka T, Iribe G, Maesako M, Mizuno J, Shimizu J, Matsubara H, Ohe T, Hirakawa M, and Suga H. Myocardial mechanical restitution and potentiation partly underlie alternans decay of postextrasystolic potentiation: simulation. Heart Vessels 14: 82–89, 1999. 23. Morad M and Goldman Y. Excitation-contraction coupling in heart muscle: membrane control of development of tension. Prog Biophys Mol Biol 27: 257–313, 1973. 24. Nakayama Y, Takaki M, Kohno K, Araki J, and Suga H. Mechanoenergetics of the negative inotropism of isoflurane in the canine left ventricle. Anesthesiology 87: 82–93, 1997. 25. Narayan P, McCune SA, Robitaille PML, Hohl CM, and Altschuld RA. Mechanical alternans and the force-frequency relationship in failing rat hearts. J Mol Cell Cardiol 27: 523– 530, 1995. 26. Noble RJ and Nutter DO. The demonstration of alternating contractile state in pulsus alternans. J Clin Invest 49: 1166– 1177, 1970. 27. Parmley WW, Tomoda H, Fujimura S, and Matloff JM. Relation between pulsus alternans and transient occlusion of the left anterior descending coronary artery. Cardiovasc Res 6: 709– 715, 1972. 28. Shimizu J, Araki J, Mizuno J, Lee S, Syuu Y, Hosogi S, Mohri S, Mikane T, Takaki M, Taylor TW, and Suga H. A new integrative method to quantify total Ca2⫹ handling and futile Ca2⫹ cycling in failing hearts. Am J Physiol Heart Circ Physiol 275: H2325–H2333, 1998. 29. Spear JF and Moore EN. A comparison of alternation in myocardial action potentials and contractility. Am J Physiol 220: 1708–1716, 1971. 30. Steiner C and Kovalik ATW. A simple technique for production of chronic complete heart block in dogs. J Appl Physiol 25: 631–632, 1968. 31. Suga H, Hisano R, Hirata S, Hayashi T, Yamada O, and Ninomiya I. Heart rate-independent energetics and systolic Downloaded from http://ajpheart.physiology.org/ by 10.220.33.3 on May 11, 2017 Limitations. One may expect that inserting an appropriately coupled extrasystole could abolish sustained alternans. If this method works, it could be used to abolish alternans. Although this possibility remains to be studied, the disappearance of alternans per se may not be beneficial as long as tachycardia remains. Besides, because we studied isovolumic contractions in the excised heart, it remains unknown whether the same phenomenon could occur in ejecting contractions in in situ hearts. Because we used normal hearts, the same phenomenon remains to be examined in failing hearts. In summary, our new finding is the coupling interval-dependent variation of the pattern and amplitude of the sustained contractile alternans. We were not able to simulate this observation with the representative Ca2⫹ handling models reported in the literature. The cellular mechanism of the phenomenon remains to be elucidated for better understanding of cardiac sustained contractile alternans and Ca2⫹ handling. COUPLING INTERVAL AND SUSTAINED ALTERNANS pressure-volume area in dog heart. Am J Physiol Heart Circ Physiol 244: H206–H214, 1983. 32. Suga H and Sagawa K. Instantaneous pressure-volume relationships and their ratio in the excised, supported canine left ventricle. Circ Res 35: 117–126, 1974. 33. Traube L. Ein Fall von Pulsus bigeminus nebst Bemerkungen über die Leberschwellungen bei Klappenfehlern und über acute Leberatrophie. Berl Klin Wschr 9: 185–188, 1872. 34. Weber KT and Janicki JS. Interdependence of cardiac function, coronary flow, and oxygen extraction. Am J Physiol Heart Circ Physiol 235: H784–H793, 1978. H1375 35. Wier WG and Yue DT. Intracellular calcium transients underlying the short-term force-interval relationship in ferret ventricular myocardium. J Physiol (Lond) 376: 507–530, 1986. 36. Wohlfart B. Analysis of mechanical alternans in rabbit papillary muscle. Acta Physiol Scand 115: 405–414, 1982. 37. Wohlfart B. [Ca2⫹]i following extrasystoles in guinea-pig trabeculae microinjected with fluo-3. A comparison with frog skeletal muscle fibres. Acta Physiol Scand 169: 1–11, 2000. 38. Yue DT, Burkhoff D, Franz MR, Hunter WC, and Sagawa K. Postextrasystolic potentiation of the isolated canine left ventricle. Relationship to mechanical restitution. Circ Res 56: 340–350, 1985. Downloaded from http://ajpheart.physiology.org/ by 10.220.33.3 on May 11, 2017