Survey
* Your assessment is very important for improving the workof artificial intelligence, which forms the content of this project
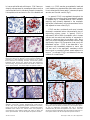
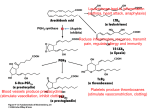
Epidemiological data and preclinical studies have generated compelling interest in the potential use of COX-2 inhibitors in chemoprevention and chemotherapy of human tumors. Michele Sassi. La Digue Island, c. 2001. Seychelles Islands, Indian Ocean. Photograph. Celecoxib: A Specific COX-2 Inhibitor With Anticancer Properties Alane T. Koki, PhD, and Jaime L. Masferrer, PhD I n addition to the well-established pathophysiological role that COX-2 plays in inflammation, recent evidence implies that this isoform may also be involved in multiple biologic events throughout the tumorigenic process. Many epidemiological studies demonstrate that nonsteroidal anti-inflammatory drugs (NSAIDs) reduce the risk of a wide range of tumors. Further, COX-2 is chronically overexpressed in many premalignant, malignant, and metastatic human cancers, and levels of overexpression have been shown to significantly correlate to invasiveness, prognosis, and survival in some cancers. Pharmacological studies consistently demonstrate that COX-2 inhibitors dose-dependently inhibit tumor growth and metastasis in various relevant animal models of cancer. Importantly, several investigators have also shown COX-2 inhibitors may act additively or synergistically with currently used cytotoxics and molecularly targeted agents. Here we present a broad overview of the growing evidence that COX-2 plays a pivotal role throughout oncogenesis and summarize the rationale to explore the use of COX-2 inhibitors for the prevention and/or treatment of cancer as a single agent or in combination with current anticancer modalities. Introduction The cyclooxygenases are responsible for the conversion of arachidonic acid to prostaglandins (PGs), and their metabolites play a pivotal role in multiple physiologic and pathophysiologic processes. Cyclooxygenase-1 (COX-1) is constitutively expressed in From the Pharmacia Corp, Chesterfield, Missouri. Address reprint requests to Alane T. Koki, PhD, Pharmacia Corporation, 700 Chesterfield Village Parkway North, Mailzone BB3K, Chesterfield, MO 63017. E-mail: [email protected] 28 Cancer Control most tissues and is responsible for maintaining physiologic processes such as gastric and renal protection and platelet function. In contrast, cyclooxygenase-2 (COX-2) is induced in response to growth factors1,2 (ie, endothelial growth factor [EGF], vascular endothelial growth factor [VEGF], fibroblast growth factor [FGF2]), cytokines (eg, tumor necrosis factor-α [TNF-α], interleukin-α [IL-α], and interleukin-1β [IL-1β]), and tumor promoters (eg, v-src, v-Ha-ras, HER-2/neu, and Wnt).3,4 COX-2 is expressed in macrophages, synoviocytes, and endothelial cells in response to inflammation and cellular activation.5-7 Conventional NSAIDs March/April 2002, Vol. 9, No.2 Supplement inhibit both COX-1 and COX-2, hence they also disrupt COX-1 dependent homeostatic functions. Therefore, molecular-based targeting strategies were employed to develop specific COX-2 inhibitors to circumvent the gastric and renal toxicities caused by mixed COX inhibitors.8-10 In addition to the well-studied role of COX-2 in acute inflammatory processes, recent work clearly suggests COX-2-derived metabolites contribute at multiple points throughout tumorigenesis, including premalignant hyperproliferation, transformation, maintenance of tumor viability, growth, invasion, and metastatic spread. Here we summarize the collective evidence that supports the potential anticancer activity of COX-2 inhibitors. Epidemiological Evidence: COX-2 and Cancer Epidemiological studies provided the first evidence that COX may be involved in the pathogenesis of cancer. Several reports indicate NSAIDs can prevent the development of various human tumors, including colon,11-18 breast,19-22 lung,19 gastric,23,24 and esophageal24,25 neoplasias. For example, in a prospective study evaluating data from 12,668 subjects over 12.4 years, the incidence of several cancers was lower in regular users of aspirin. The incidence rate for all cancers combined was 0.83 (95% CI, 0.74-0.93), lung cancer 0.68 (95% CI, 0.49-0.94), breast cancer in women 0.70 (95% CI, 0.50-0.96), and colorectal cancer in younger men 0.35 (95% CI, 0.17-0.73).19 These data collectively provide the rationale for additional clinical trials to stringently evaluate NSAID efficacy in other types of cancers as well. COX-2 Expression in Human Tumors To understand the potential importance and role of COX-2 in human cancers, we developed and utilized immunohistochemical techniques to stringently characterize COX-2 in progressing stages of tumorigenesis Head and neck Bladder Cholangiocarcinoma Hepatocellular Renal Cell Breast Mesothelioma Endometrial Pancreatic Prostate Basal Cell Gastric 1A 1B Fig 1. — COX-2 is up-regulated in various solid tumors. Fig 1a shows up-regulation in head and neck, bladder, renal cell, breast, pancreatic, and prostate cancer. Fig 1b shows up-regulation in cholangiocarcinoma, hepatocellular (HCC), mesothelioma, endometrial, basal cell, and gastric cancers. Formalinfixed, paraffin-embedded human cancers from archival tissues were prepared by immunohistology using standard methods and stained for COX-2 with isoform-specific antisera. The red stain depicts the presence of COX-2. March/April 2002, Vol. 9, No.2 Supplement Cancer Control 29 in human epithelial and solid tumors. COX-2 was consistently overexpressed in premalignant lesions such as oral leukoplakia, actinic keratosis, prostatic intraepithelial neoplasia, and carcinoma in situ of the bladder and breast.11,22,26 COX-2 was also up-regulated in head and neck, bladder, lung, hepatocellular, pancreatic, mesothelioma, cholangiocarcinoma, gastric, cutaneous, Kaposi’s sarcoma, and cervical cancers (Fig 1). In general, COX-2 is up-regulated throughout the tumorigenic process, from early hyperplasia to metastatic disease (Fig 2). Elevated levels of COX-2 immunoreactivity were primarily detected in the neoplastic epithelium, inflammatory cells, and vasculature within and adjacent to tumor nests (Fig 3). Normal Ductal CA DCIS CA Infiltrating CA COX-2 was also consistently and more intensely observed in metastatic lesions compared with the corresponding primary tumor. In general, COX-2 is expressed in 40% to 80% of neoplastic cells in human cancers and the extent and intensity of expression is greater in cancerous than in noncancer cells. Moreover, well- and moderately-differentiated cancers have significantly higher COX-2 expression than poorly differentiated cancers. COX-2 is also detected in noncancerous cells immediately adjacent to tumor cells (<2 mm) and in the angiogenic vasculature within tumors and in pre-existing blood vessels adjacent to tumors.6 In contrast, COX-2 is not detected in the vasculature of normal tissues.27 Fig 2a. — COX-2 is expressed throughout mammary carcinogenesis. As shown here, COX-2 is not detected in normal mammary epithelial cells (normal) but is clearly expressed in ductal carcinoma in situ (DCIS), ductal carcinoma, and infiltrating carcinoma. Formalin-fixed, paraffinembedded human cancers from archival tissues were prepared by immunohistology using standard methods and stained for COX-2 with isoform-specific antisera. The red stain depicts the presence of COX-2. A Normal Primary CA Hyperplasia Metastasis to Lymph Node B Fig 2b. — COX-2 is up-regulated in colonic tumorigenesis. COX-2 is not detected in normal cells. However, enhanced levels of COX-2 are observed in early colonic hyperplasia (sporadic adenomatous carcinoma), primary colon cancer, and metastaticdisease. Formalin-fixed, paraffinembedded human cancers from archival tissues were prepared by immunohistology using standard methods and stained for COX-2 with isoform-specific antisera. The red stain depicts the presence of COX-2. 30 Cancer Control C Fig 3. — COX-2 immunoreactivity in human bladder cancer. Moderate-tostrong COX-2 immunoreactivity is detected in (a) the neoplastic epithelium (CA), (b) inflammatory (macrophage) cells, and (c) vasculature endothelial cells (EC) within and adjacent to tumor nests. Tissue was excised from patients with bladder cancer and stained for COX-2 with isoform-specific antisera. The red stain depicts the presence of COX-2. March/April 2002, Vol. 9, No.2 Supplement Our expression studies are consistent with those reported by others28-38 and collectively imply COX-2 activity may be responsible for increased prostaglandin levels in cancer tissues.39,40 Importantly, recent work demonstrates a relationship between overexpression of COX-2 and the invasiveness and survival of patients with breast,41 colon,42-44 gastric,45,46 and lung47 cancers. Association Between HER-2/neu and COX-2 Expression HER-2/neu, also known as c-erbB-2, is a receptor protein kinase whose overexpression is widely accepted as an adverse prognostic marker.48 Interestingly, COX-2 is detected in a subset of hormonally driven tumors such as breast, prostate, and ovarian cancers.6 For example, COX2 overexpression in mammary tumors appears to be correlated with HER-2/neu. In a landmark study, Subbaramaiah et al49 evaluated COX-2 expression in 29 patients with breast cancer and reported high levels of COX-2 in 14 of 15 HER-2/neu positive patients. In contrast, only 4 of 14 HER-2/neu negative patients expressed COX-2, and detected levels were much lower than in any of the HER2/neu positive patients. Recently we observed that COX2 and HER-2/neu are similarly distributed in breast cancer tissue and are often coexpressed in hyperproliferating, dysplastic, and neoplastic epithelial cells.50 In another study, we evaluated archival sections of human mammary lesions, including DCIS, ductal, lobular, infiltrating, and mucinous mammary cancers for HER-2/neu and COX-2. Of the 25 cases evaluated, 15 were HER-2/neu positive and 10 were HER-2/neu negative. Both COX-2 and HER2/neu were overexpressed in 3 of 3 cases of DCIS and 10 of 12 cases with ductal, lobular, and infiltrating carcinomas, and both were over expressed in the same cells. COX-2 was expressed less often in HER-2/neu negative breast cancers.51 To determine if these two oncogenes were functionally linked, HER-2/neu and COX-2 negative breast cancer cells were engineered to express HER-2/neu. COX-2 was strongly induced in HER-2/neu transfected cells but was not expressed in null transfected cells.52 Collectively, these data suggest that HER-2/neu may modulate the overexpression of COX-2 in human cancer.52 However, the upstream modulator(s) of COX-2 in ovarian and prostate cancer have yet to be elucidated. The correlation between COX-2 and HER-2/neu expression in human breast cancers may have important treatment implications. For example, patients with early disease such as DCIS are often given the option of treatment vs no treatment after lumpectomy. These data collectively imply HER-2neu/COX-2 positive patients with DCIS may represent a high-risk patient population that March/April 2002, Vol. 9, No.2 Supplement may benefit from treatment with celecoxib. Additionally, combination therapy with celecoxib and other moleculartargeted agents such as aromatase inhibitors (ie, exemestane) or agents that block HER-2/neu activation may also prove beneficial in clinical trials of breast cancer. Indeed, future research on the effect of COX-2 inhibition on HER2/neu positive vs HER-2/neu negative mammary tumors will significantly contribute to the present understanding of the potential clinical utility of COX-2 inhibitors for the prevention and/or treatment of human breast cancer. Mechanisms of COX-2–Associated Tumorigenesis COX-2 is overexpressed along the continuum of oncogenesis and is likely to be a key player in a number of biologic pathways leading to cancer. Current evidence indicates that COX-2 promotes tumor-specific angiogenesis,53-56 inhibits apoptosis,57-60 and induces proangiogenic factors such as VEGF,61,62 inducible nitrogen oxide synthetase promoter (iNOS),63 IL-6,64 IL-8,65 and TIE-2.66 COX-2-derived metabolites from infiltrating inflammatory cells undoubtedly contribute to the tumorigenic process as well. For example, enhanced prostaglandin synthesis may contribute to oncogenesis by directly stimulating mitogenesis in fibroblasts,67 osteoblasts,68,69 and mammary epithelial cells.70 Excessive local synthesis of prostaglandins has also been shown to disrupt immune surveillance that may otherwise suppress tumor growth.71,72 In addition, the direct product of COX-2, PGH2, can isomerize by both enzymatic and nonenzymatic reactions to form the potent mutagen malondialdehyde, which can induce frame shifts and base pair substitutions.73 Additional free radical damage may occur via the peroxidative activity of COX-2, which can efficiently oxidize aromatic and heterocyclic amines and dihydrodiol derivatives.74 Increased prostaglandin levels may be particularly important during the progression of breast cancer. PGE2 has recently been shown to stimulate aromatase transcription,75,76 leading to supraphysiologic local estrogen levels, which in turn leads to the subsequent release of growth factors and enhanced proliferation.77 Immunohistochemical analysis by Brueggemeier et al78 supports the association between COX-2 and aromatase. The cytochrome P450 enzyme aromatase (CYP19) and COX2 were coexpressed in all cases of human breast cancer, and a significant linear association was found between levels of CYP19 gene expression and COX-2 gene expression (R - 0.80, P<.0001).78 In addition to increasing aromatase transcription, COX-2-induced PGE2 also promotes angiogenesis,53-55 Cancer Control 31 Fig 4. — Hypothetical mechanism by which celecoxib may enhance the anticancer activity of aromatase inhibitors. COX-2-derived PGE2 potentiates estrogen biosynthesis by the cytochrome P450 enzyme aromatase and promotes angiogenesis, tumor maintenance/growth, and metastasis. The simultaneous inhibition of aromatase and COX-2 activity may therefore enhance the anticancer activity of either agent alone. which is required for tumor growth and metastasis.79,80 Taken together, these data suggest that both autocrine and paracrine mechanisms may be responsible for the development and/or progression of estrogen-dependent breast cancer either directly by stimulation of tumor cell proliferation or indirectly by enhanced prostaglandin-regulated local estrogen synthesis (Fig 4). Anticancer Activity of COX Inhibitors in Relevant Animal Models of Cancer Retrospective and/or epidemiological studies of conventional NSAID use and incidence of cancer led to the hypothesis that COX-derived metabolites play an important role in tumorigenesis. The inhibitory effect of COX inhibitors on tumor growth in relevant animal models of various epithelial cancers supports this hypothesis.81-86 However, maximum efficacy in these studies is typically dose-limited by COX-1-related toxicities. In contrast, COX-2 inhibitors have been shown to markedly inhibit tumor growth and metastasis in several animal models of colon,87-90 skin,91,92 lung,6 bladder,93 and breast cancers.94,95 For example, sporadic colorectal adenomas and adenocarcinomas can be generated by administering azoxymethane (AOM). When tested in this model, celecoxib effectively prevents the development of colon tumors in 93% of rats and inhibits tumor initiation by approximately 80%.88,96 In this model, the chemopreventive effect of celecoxib was dose-related and treatment was effective during both initiation and postinitiation phases of the disease.87 Likewise, Harris et al94 evaluated the chemopreventive potential of celecoxib and ibuprofen in the 7,12-dimethyl-benz(a)anthracene (DMBA) model of 32 Cancer Control breast cancer in female Sprague-Dawley rats and compared the results with untreated control animals. All rats were pretreated 7 days before DMBA administration and therapy was continued for 105 days. Malignant tumors developed in 100% of control animals; 95% had multiple tumors and tumor size was >1.5 cm3. Both celecoxib and ibuprofen significantly suppressed the incidence, burden, and volume of malignant tumors and prolonged the latency period of tumor induction compared with controls. However, the effects of celecoxib were significantly more pronounced than those of ibuprofen (Table).94 In a follow-up chemoprevention study in this model, Abou-Issa et al97 reported that mammary tumor incidence, burden, and volume were inhibited in a dose-dependent manner. Tumor incidence was 100% in controls vs 55%, 45%, and 25% in rats fed a diet of celecoxib 500, 1,000, or 1,500 ppm, respectively (P=.001).97 The chemotherapeutic activity of celecoxib was next evaluated on established DMBA-induced mammary tumors by Alshafie and coworkers.95 DMBA-treated rats were randomized to control or celecoxib following 6 weeks of DMBA treatment. Mammary tumors continued to grow in the control group, with an average increase in tumor size of 500% vs baseline. In the celecoxib-treated group, tumor regression was reported in approximately 90% of animals, and tumor was reduced 32% compared with baseline (P<.04). In contrast, 10 new mammary tumors were identified in the control group (30% increase). The number of palpable tumors decreased in celecoxib-treated animals from 24 to 18 (25%; P<.05); no decrease in the number of tumors was observed in the control group.95 To identify the potential points of intervention during tumorigenesis, Alshafie et al98 dosed DMBA-treated rats with celecoxib only during the initiation or promotion phases of carcinogenesis or during both initiation and promotion phases. All treatment paradigms signifiChemopreventive Effects of Celecoxib and Ibuprofen: Mammary Cancer Incidence, Tumor Burden, Tumor Volume, and Latency of Tumor Induction in a DMBA Model of Breast Cancer Effect vs Control Celecoxib Ibuprofen Reduction in tumor incidence, % 68 * † 40* Reduction in tumor burden, % 86 *† 52* Reduction in tumor volume, % 81 * † 57* Latency period of tumor induction, days‡ 95 86 * P<.001 vs controls † P<.01 vs ibuprofen ‡ 58 days in controls Data from Harris et al.94 March/April 2002, Vol. 9, No.2 Supplement cantly reduced tumor incidence and growth (P=.001), with the effect being maximal when celecoxib was fed during both the initiation and promotion phases.98 Based on the association between COX-2 and aromatase gene expression,75,78 Pesenti and colleagues99 evaluated the therapeutic potential of celecoxib (500 ppm in diet) and exemestane (50 mg/kg weekly) alone and in combination in the DMBA model of breast cancer. Female Sprague-Dawley rats with at least one tumor measuring 1 cm in size were studied. An objective response (OR) rate of 48% was achieved with combination therapy, compared with OR rates of 5% in rats treated with exemestane alone and 0 in rats treated with celecoxib alone and in control rats. The development of new tumors followed a similar pattern. These data demonstrate that the combination of exemestane and celecoxib is more effective than either agent alone in reducing tumor burden and volume, as well as the incidence of new tumors in this animal model.99 Moreover, it demonstrates that, while celecoxib inhibits the exaggerated induction of aromatase, it does not eliminate baseline or physiologic aromatase production. The addition of exemestane is required, therefore, to block the activity of aromatase enzyme whose production is not blocked by celecoxib. Antiangiogenic agents have also been shown to enhance the antitumor activity of cytotoxic drugs when given in combination in rodent models of cancer.100,101 In addition to anti-inflammatory properties, recent work convincingly demonstrates that celecoxib can also act as a potent antiangiogenic agent.54 We therefore evaluated the antitumor activity of celecoxib as a monotherapy and in combination with cytotoxic chemotherapy in the HT-29 human colon carcinoma xenograft model and in the Lewis lung carcinoma syngeneic model. The combined modality of celecoxib with cytotoxic agents was more effective in both cancer models than either agent alone. Recent work also suggests that COX-2 inhibitors may potentiate radiation therapy by increasing cellular radiosensitivity and greatly enhancing tumor response when these two modalities are administered in combination.102,103 Discussion and Conclusions We have presented data to support the hypothesis that COX-2 activity modulates critical steps in the initiation, promotion, and progression of several human epithelial cancers. COX-2-derived prostaglandins may promote the development of cancer by various mechanisms, including stimulation of tumor cell growth and neovascularization.53-60,75,78,104-106 Thus, it is not surprising that COX-2 inhibitors such as celecoxib markedly inhibit March/April 2002, Vol. 9, No.2 Supplement tumor growth and metastasis in a dose-dependent manner in numerous animal models of solid tumors.87,90-94 However, the exact manner by which celecoxib reduces tumorigenesis has not been fully elucidated. The ultimate goal of cancer treatment is to specifically prevent the growth of precancerous or cancerous cells without affecting normal cells. This is particularly important in chemoprevention and treatment of early disease, which typically involves long-term treatment to healthy subjects. Therefore, NSAID selection should be based on safety as well as efficacy. Recent research has clearly established that specific COX-2 inhibitors are associated with less toxicity than the conventional mixed COX inhibitors.10 In summary, COX-2 is overexpressed in both early and late stages of carcinogenesis and has been shown to be efficacious as monotherapy and in combination with conventional chemotherapeutics in relevant animal models. Taken together, the epidemiological data and preclinical studies in animal models have generated compelling interest in the potential use of COX-2 inhibitors in chemoprevention and chemotherapy of human tumors. Clinical trials will be necessary to determine whether COX-2 inhibitors will provide clinical benefit, as well as to define the intervention points during tumor progression that will allow for optimal efficacy. References 1. Perkins DJ, Kniss DA. Rapid and transient induction of cyclooxygenase 2 by epidermal growth factor in human amnion-derived WISH cells. Biochem J. 1997;321(pt 3):677-681. 2. Diaz A, Chepenik KP, Korn JH, et al. Differential regulation of cyclooxygenases 1 and 2 by interleukin-1 beta, tumor necrosis factoralpha, and transforming growth factor-beta 1 in human lung fibroblasts Exp Cell Res. 1998;241:222-229. 3. Howe LR, Subbaramaiah K, Chung WJ, et al. Transcriptional activation of cyclooxygenase-2 in Wnt-1 transformed mouse mammary epithelial cells. Cancer Res. 1999;59:1572-1577. 4. Vadlamudi R, Mandal M,Adam L, et al. Regulation of the cyclooygenas-2 pathway by HER2 receptor. Oncogene. 1999;18:305-314. 5. Crofford LJ. COX-1 and COX-2 tissue expression: implications and predictions. J Rheumatol. 1997;24(suppl 49):15-19. 6. Koki AT, Leahy KM, Masferrer JL. Potential utility of COX-2 inhibitors in chemoprevention and chemotherapy. Expert Opin Investig Drugs. 1999;8:1623-1638. 7. Williams CS, Mann M, DuBois RN. The role of cyclooxygenases in inflammation, cancer, and development. Oncogene. 1999;18: 7908-7916. 8. Lanza FL. A guideline for the treatment and prevention of NSAID-induced ulcers. Members of the Ad Hoc Committee on Practice Parameters of the American College of Gastroenterology. Am J Gastroenterol. 1998;93:2037-2046. 9. Singh G, Ramey DR. NSAID induced gastrointestinal complications: the ARAMIS perspective - 1997. J Rheumatol. 1998;25 (suppl 51):8-16. 10. Silverstein FE, Faich G, Goldstein JL, et al. Gastrointestinal toxicity with celecoxib vs nonsteroidal anti-inflammatory drugs for osteoarthritis and rheumatoid arthritis: the CLASS study: a randomized controlled trial. Celecoxib Long-term Arthritis Safety Study. JAMA. 2000;284:1247-1255. 11. Gridley G, McLaughlin JK, Ekbom A, et al. Incidence of cancer among patients with rheumatoid arthritis. J Natl Cancer Inst. 1993;85:307-311. Cancer Control 33 12. Suh O, Mettlin C, Petrelli NJ. Aspirin use, cancer, and polyps of the large bowel. Cancer. 1993;72:1171-1177. 13. Rosenberg L, Palmer JR, Zauber GA, et al. A hypothesis: nonsteroidal anti-inflammatory drugs reduce the incidence of large bowel cancer. J Natl Cancer Inst. 1991;83:355-358. 14. Peleg II, Maibach HT, Brown SH, et al. Aspirin and nonsteroidal anti-inflammatory drug use and the risk of subsequent colorectal cancer. Arch Intern Med. 1994;154:394-399. 15. Muscat JE, Stellman SD,Wynder EL. Nonsteroidal anti-inflammatory drugs and colorectal cancer. Cancer. 1994;74:1847-1854. 16. Morgan G. Non-steroidal anti-inflammatory drugs and the chemoprevention of colorectal and oesophageal cancers. Gut. 1996; 38:646-648. 17. Thun MJ, Namboodiri MM, Calle EE, et al. Aspirin use and the risk of fatal cancer. Cancer Res. 1993;53:1322-1327. 18. Reeves MJ, Newcomb PA,Trentham-Dietz A, et al. Nonsteroidal anti-inflammatory drug use and protection against colorectal cancer in women. Cancer Epidemiol Biomarkers Prev. 1996;5:955-960. 19. Schreinemachers DM, Everson RB. Aspirin use and lung, colon, and breast cancer incidence in a prospective study. Epidemiology. 1994;5:138-146. 20. Harris RE, Nambookiri KK, Farrar WB. Nonsteroidal antiinflammatory drugs and breast cancer. Epidemiology. 1996;7:203205. 21. Harris RE, Namboodiri KK, Farrar WB. Epidemiological study of non-steroidal anti-inflammatory drugs and breast cancer. Oncol Rep. 1995;2:591-592. 22. Harris RE, Kasbari S, Farrar WB. Prospective study of nonsteroidal anti-inflammatory drugs and breast cancer. Oncol Rep. 1999;6(1):71-73. 23. Thun MJ. NSAID use and decreased risk of gastrointestinal cancers. Gastroenterol Clin North Amer. 1996;25:333-348. 24. Farrow DC,Vaughan TL, Hansten PD, et al. Use of aspirin and other nonsteroidal anti-inflammatory drugs and risk of esophageal and gastric cancer. Cancer Epidemiol Biomarkers Prev. 1998;7:97102. 25. Funkhouser EM, Sharp GB. Aspirin and reduced risk of esophageal carcinoma. Cancer. 1995;76:1116-1119. 26. Soslow RA, Dannenberg AJ, Rush D, et al. COX-2 is expressed in human pulmonary, colonic, and mammary tumors. Cancer. 2000; 89:2637-2645. 27. Buckman SY, Gresham A, Hale P, et al. COX-2 expression is induced by UVB exposure in human skin: implications for the development of skin cancer. Carcinogenesis. 1998;19:723-729. 28. Molina MA, Sitja-Arnau M, Lemoine MG, et al. Increased cyclooxygenase-2 expression in human pancreatic carcinomas and cell lines: growth inhibition by nonsteroidal anti-inflammatory drugs. Cancer Res. 1999;59:4356-4362. 29. Shirahama T, Sakakura C. Overexpression of cyclooxygenase2 in squamous cell carcinoma of the urinary bladder. Clin Cancer Res. 2001;7:558-561. 30. Okami J, Yamamoto H, Fujiwara Y, et al. Overexpression of cyclooxygenase-2 in carcinoma of the pancreas. Clin Cancer Res. 1999;5:2018-2024. 31. Zimmermann KC, Sarbia M,Weber AA, et al. Cyclooxygenase2 expression in human esophageal carcinoma. Cancer Res. 1999;59: 198-204. 32. Yoshimura R, Sano H, Masuda C, et al. Expression of cyclooxygenase-2 in prostate carcinoma. Cancer. 2000;89:589-596. 33. Karim MM, Hayashi Y, Inoue M, et al. Cox-2 expression in retinoblastoma. Am J Ophthalmol. 2000;129:398-401. 34. Shiota G, Okubo M, Noumi T, et al. Cyclooxygenase-2 expression in hepatocellular carcinoma. Hepatogastroenterology. 1999;46: 407-412. 35. Mohammed SI, Knapp DW, Bostwick DG, et al. Expression of cyclooxygenase-2 (COX-2) in human invasive transitional cell carcinoma (TCC) of the urinary bladder. Cancer Res. 1999;59:5647-5650. 36. Kulkarni S, Rader JS, Zhang F, et al. Cyclooxygenase-2 is overexpressed in human cervical cancer. Clin Cancer Res. 2001;7:429434. 37. Chan G, Boyle JO, Yang EK, et al. Cyclooxygenase-2 expression is up-regulated in squamous cell carcinoma of the head and neck. Cancer Res. 1999;59:991-994. 38. Tucker ON, Dannenberg AJ,Yang EK, et al. Cyclooxygenase-2 expression is up-regulated in human pancreatic cancer. Cancer Res. 1999;59:987-990. 39. Bennett A, Charlier EM, McDonald AM, et al. Prostaglandin and breast cancer. Lancet. 1977;2:624-626. 34 Cancer Control 40. Laekeman GM, Vergote IB, Keersmaeker GM, et al. Prostacyclin and thromboxane in benign and malignant breast tumors. Br J Cancer. 1986;54:431-437. 41. Parrett ML, Harris RE, Joarder FS, et al. Cyclooxygenase-2 gene expression in human breast cancer. Int J Oncol. 1997;10:503-507. 42. Tomozawa S, Tsuno NH, Sunami E, et al. Cyclooxygenase-2 overexpression correlates with tumour recurrence, especially haematogenous metastasis, of colorectal cancer. Br J Cancer. 2000;83:324-348. 43. Tsujii M, Kawano S, DuBois RN. Cyclooxygenase-2 expression in human colon cancer cells increases metastatic potential. Proc Natl Acad Sci U S A. 1997;94:3336-40. 44. Chen WS, Wei SJ, Liu JM, et al. Tumor invasiveness and liver metastasis of colon cancer cells correlated with cyclooxygenase-2 (COX-2) expression and inhibited by a COX-2-selective inhibitor, etodolac. Int J Cancer. 2001;91:894-899. 45. Murata H, Kawano S,Tsuji S, et al. Cyclooxygenase-2 overexpression enhances lymphatic invasion and metastasis in human gastric carcinoma. Am J Gastroenterol. 1999;94:451-455. 46. Chen CN, Sung CT, Lin MT, et al. Clinicopathologic association of cyclooxygenase 1 and cyclooxygenase 2 expression in gastric adenocarcinoma. Ann Surg. 2001;233:183-188. 47. Achiwa H, Yatabe Y, Hida T, et al. Prognostic significance of elevated cyclooxygenase 2 expression in primary, resected lung adenocarcinomas. Clin Cancer Res. 1999;5:1001-1005. 48. Slamon DJ, Clark GM, Wong SG, et al. Human breast cancer: correlation of relapse and survival with amplification of the HER2/neu oncogene. Science. 1987;235:177-182. 49. Subbaramaiah K, Norton L, Gerald W, et al. Increased expression of cyclooxygenase-2 in HER-2/neu overexpressing human breast cancer cells. National Cancer Institute 7th Spore Investigators Workshop,. Rockville, Md; 1999. 50. Koki AT, Harmon J, Woerner BM, et al. Characterization of cyclooxygenase-2 (COX-2) expression and Her2/neu in human breast, colon, and prostate cancer (abstract 101). Clin Cancer Res. 2000;6(suppl):4486s. 51. Harmon JM, Tsai CC, Dannenberg AJ, Koki AT. Cyclooxygenase-2 (COX-2) is co-localized with HER-2/neu in human breast cancer (abstract 4804). Proc Am Assoc Cancer Res. 2001;42:895. 52. Howe LR, Subbaramaiah K, Brown AM, et al. Cyclooxygenase2: a target for the prevention and treatment of breast cancer. Endocr Relat Cancer. 2001;8:97-114. 53. Leahy KM, Ornberg RL,Want Y, et al. Cyclooxygenase-2 inhibition by celecoxib reduces proliferation and induces apoptosis in angiogenic endothelial cells in vivo. Cancer Res. 2002;62:625-631. 54. Masferrer JL, Leahy KM, Koki AT, et al. Antiangiogenic and antitumor activities of cyclooxygenase-2 inhibitors. Cancer Res. 2000;60:1306-1311. 55. Leahy KM, Koki AT, Masferrer JL. Role of cyclooxygenases in angiogenesis. Curr Med Chem. 2000;7:1163-1170. 56. Liu XH, Kirschenbaum A,Yao S, et al. Inhibition of cyclooxygenase-2 suppresses angiogenesis and the growth of prostate cancer in vivo. J Urol. 2000;164:820-825. 57. Souza RF, Shewmake K, Beer DG, et al. Selective inhibition of cyclooxygenase-2 suppresses growth and induces apoptosis in human esophageal adenocarcinoma cells. Cancer Res. 2000;60: 5767-5772. 58. Tsujii M, DuBois RN. Alterations in cellular adhesion and apoptosis in epithelial cells overexpressing prostaglandin endoperoxide synthase 2. Cell. 1995;83:493-501. 59. Ding XZ,Tong WG,Adrian TE. Blockade of cyclooxygenase-2 inhibits proliferation and induces apoptosis in human pancreatic cancer cells. Anticancer Res. 2000;20:2625-2631. 60. Hsu AL, Ching TT, Wang DS, et al. The cyclooxygenase-2 inhibitor celecoxib induces apoptosis by blocking Akt activation in human prostate cancer cells independently of Bcl-2. J Biol Chem. 2000;275:11397-11403. 61. Shweiki D, Itin A, Soffer D, et al. Vascular endothelial growth factor induced by hypoxia may mediate hypoxia-initiated angiogenesis. Nature. 1992;359:843-845. 62. Liu Y, Cox SR, Morita T, et al. Hypoxia regulates vascular endothelial growth factor gene expression in endothelial cells. Circ Res. 1995;77:638-645. 63. Mellilo G, Musso T, Sica A, et al. A hypoxia-responsive element mediates a novel pathway of activation of the inducible nitric oxide synthase promoter. J Exp Med. 1995;182:1683-1693. 64. Yan SF,Tritto I, Pinsky D, et al. Induction of interleukin 6 (IL6) by hypoxia in vascular cells. Central role of the binding site for March/April 2002, Vol. 9, No.2 Supplement nuclear factor-IL-6. J Biol Chem. 1995;270:11463-11471. 65. Karakurum M, Shreeniwas R, Chen J, et al. Hypoxic induction of interleukin-8 gene expression in human endothelial cells. J Clin Invest. 1994;93:1564-1570. 66. Tian H, McKnight SL, Russel DW. Endothelial PAS domain protein I (EPAS1), a transcription factor selectively expressed in endothelial cells. Genes Dev. 1997;11:72-82. 67. Nolan RD, Danilowicz RM, Eling TE. Role of arachidonic acid metabolism in the mitogenic response of BALB/c 3T3 fibroblasts to epidermal growth factor. Mol Pharmacol. 1988;33:650-656. 68. Goin M, Pignataro O, Jimenez de Asua L. Early cell cycle diacylglycerol (DAG) content and protein kinase C (PKC) activity enhancement potentiates prostaglandin F2 alpha (PGF2 alpha) induced mitogenesis in Swiss 3T3 cells. FEBS Lett. 1993;316:68-72. 69. Quarles LD, Haupt DM, Davidai G, et al. Prostaglandin F2 alpha-induced mitogenesis in MC3T3-E1 osteoblasts: role of protein kinase-C-mediated tyrosine phophorylation. Endocrinology. 1993; 132:1505-1513. 70. Bandyopadhyay GK, Imagawa W, Wallace D, et al. Linoleate metabolites enhance the in vitro proliferative response of mouse mammary epithelial cells to epidermal growth factor. J Biol Chem. 1987;262:2750-2756. 71. Stolina M, Sharma S, Zhu L, et al. Lung cancer cyclooxygenase-2 dependent inhibition of dendritic cell maturation and function. Proc Am Assoc Cancer Res. 2000;41:619. 72. Huang M, Sharma S, Mao JT, et al. Non-small cell lung cancerderived soluble mediators and prostaglandin E2 enhance peripheral blood lymphocyte IL-10 transcription and protein production. J Immunol. 1996;157:5512-5520. 73. Marnett LJ. Aspirin and the potential role of prostaglandins in colon cancer. Cancer Res. 1992;52:5575-5589. 74. Wiese FW, Thompson PA, Kadlubar FF. Carcinogen substrate specificity of human COX-1 and COX-2. Carcinogenesis. 2001;21:5-10. 75. Zhao Y,Agarwal VR, Mendelson CR, et al. Estrogen biosynthesis proximal to a breast tumor is stimulated by PGE2 via cyclic AMP, leading to activation of promoter II of the CYP 19 (aromatase) gene. Endocrinology. 1996;137:5739-5742. 76. Purohit A, Singh A, Ghilchik MW, et al. Inhibition of tumor necrosis factor alpha-stimulated aromatase activity by microtubulestabilizing agents, paclitaxel and 2-methoxy-estradiol. Biochem Biophys Res Commun. 1999;261:214-217. 77. Harris RE, Robertson FM,Abou-Issa HM, et al. Genetic induction and upregulation of cyclooxygenase (COX) and aromatase (CYP19): an extension of the dietary fat hypothesis of breast cancer. Med Hypotheses. 1999;52:291-292. 78. Brueggemeier RW, Quinn AL, Parrett ML, et al. Correlation of aromatase and cyclooxygenase gene expression in human breast cancer specimens. Cancer Lett. 1999;140:27-35. 79. Folkman J. Tumor angiogenesis: therapeutic implications. N Engl J Med. 1971;285:1182-1186. 80. Liotta LA, Kleinerman J, Saidel GM. Quantitative relationships of intravascular tumor cells, tumor vessels, and pulmonary metastases following tumor implantation. Cancer Res. 1974;34:997-1004. 81. Robertson FM, Parrett ML, Joarder FS, et al. Ibuprofen-induced inhibition of cyclooxygenase isoform gene expression and regression of rat mammary carcinomas. Cancer Lett. 1998;122:165-175. 82. McCormick DL, Madigan MJ, Moon RC. Modulation of rat mammary carcinogenesis by indomethacin. Cancer Res. 1985;45: 1803-1808. 83. Alshafie GA, Harris RE, Robertson FM, et al. Comparative chemopreventive activity of ibuprofen and N-(4-hydroxyphenyl) retinamide against the development and growth of rat mammary adenocarcinomas. Anticancer Res. 1999;19:3031-3036. 84. Sheng H, Shao J, Kirkland SC, et al. Inhibition of human colon cancer cell growth by selective inhibition of cylcooxygenase-2. J Clin Invest. 1997;99:2254-2259. 85. Sawaoka H, Kawano S, Tsuji S, et al. Cyclooxygenase-2 inhibitors suppress the growth of gastric cancer xenografts via induction of apoptosis in nude mice. Am J Physiol. 1998;274:G1061G1067. 86. Oshima M, Dinchuk JE, Kargman SL, et al. Suppression of intestinal polyposis in Apc delta716 knockout mice by inhibition of cyclooxygenase 2 (COX-2). Cell. 1996;87:803-809. 87. Reddy BS, Hirose Y, Lubet R, et al. Chemoprevention of colon cancer by specific cyclooxygenase-2 inhibitor, celecoxib, administered during different stages of carcinogenesis. Cancer Res. 2000;60: 293-297. 88. Kawamori T, Rao CV, Seibert K, et al. Chemopreventive activMarch/April 2002, Vol. 9, No.2 Supplement ity of celecoxib, a specific cyclooxygenase-2 inhibitor, against colon carcinogenesis. Cancer Res. 1998;58:409-412. 89. Williams CS, Watson AJ, Sheng H, et al. Celecoxib prevents tumor growth in vivo without toxicity to normal gut: lack of correlation between in vitro and in vivo models. Cancer Res. 2000; 60:6045-6051. 90. Jacoby RF, Seibert K, Cole CE, et al. The cyclooxygenase-2 inhibitor celecoxib is a potent preventive and therapeutic agent in the min mouse model of adenomatous polyposis. Cancer Res. 2000;60:5040-5044. 91. Wilgus TA, Ross MS, Parrett ML, et al. Topical application of a selective cyclooxygenase inhibitor, celecoxib, suppresses UVB-mediated cutaneous inflammation. Postaglandins Other Lipid Mediat. 2000;62:367-384. 92. Pentland AP, Schoggins JW, Scott GA, et al. Reduction of UVinduced skin tumors in hairless mice by selective COX-2 inhibition. Carcinogenesis. 1999;20:1939-1944. 93. Grubbs CJ, Lubet RA, Koki AT, et al. Celecoxib inhibits Nbutyl-N-(4-hydroxybutyl)-nitrosamine-induced urinary bladder cancers in male B6D2F1 mice and female Fischer-344 rats. Cancer Res. 2000;60:5599-5602. 94. Harris RE, Alshafie GA, Abou-Issa H, et al. Chemoprevention of breast cancer in rats by celecoxib, a cyclooxygenase 2 inhibitor. Cancer Res. 2000;60:2101-2103. 95. Alshafie GA, Abou-Issa HM, Seibert K, et al. Chemotherapeutic evaluation of Celecoxib, a cyclooxygenase-2 inhibitor, in a rat mammary tumor model. Oncol Rep. 2000;7:1377-1381. 96. Reddy BS, Rao CV, Seibert K. Evaluation of cyclooxygenase-2 inhibitor for potential chemopreventive properties in colon carcinogenesis. Cancer Res. 1996;56:4566-4569. 97. Abou-Issa HM, Alshafie GA, Seibert K, et al. Dose-response effects of the COX-2 inhibitor, celecoxib, on the chemoprevention of mammary carcinogenesis (abstract 1655). Proc Am Assoc Cancer Res. 2001;42:307. 98. Alshafie GA, Abou-Issa HM, Seibert k. et al. Relative chemopreventive efficacy of the COX-2 inhibitor, celecoxib, on the initiation and promotion phases of rat mammary carcinogenesis (abstract). Proc Am Assoc Cancer Res. 2001;42:307. 99. Pesenti E, Masferrer JL, di Salle E. Effect of exemestane and celecoxib alone or in combination on DMBA-induced mammary carcinoma in rats (abstract 445). Breast Cancer Res. 2001;69:288. 100. Klement G, Baruchel S, Rak J, et al. Continuous low-dose therapy with vinblastine and VEGF receptor-2 antibody induces sustained tumor regression without overt toxicity. J Clin Invest. 2000;105:R15-R24. 101. Kakeji Y,Teicher BA. Preclinical studies of the combination of angiogenic inhibitors with cytotoxic agents. Invest New Drugs. 1997;15:39-48. 102. Peterson S, Petersen C, Kishi K, et al. COX-2 inhibitor enhances tumor radioresponse in vitro and in vivo. Eicosanoids & Other Bioactive Lipids in Cancer, Inflammation, and Related Diseases. 7th International Conference. 1999;A11:85. 103. Milas L, Kishi K, Hunter N, et al. Enhancement of tumor response to gamma-radiation by an inhibitor of cyclooxygenase-2 enzyme. J Natl Cancer Inst. 1999;91:1501-1504. 104. Attiga FA, Fernandez PM, Weeraratna AT, et al. Inhibitors of prostaglandin synthesis inhibit human prostate tumor cell invasiveness and reduce the release of matrix metalloproteinases. Cancer Res. 2000;60:4629-4637. 105. Marnett L. Aspirin and the potential role of prostaglandins in colon cancer. Cancer Res. 1992;52:5575-5589. 106. Masferrer JL, Leahy K, Zweifel BS, et al. Celecoxib: a specific cyclooxygenas-2 (COX-2) inhibitor with antiangiogenic and anticancer activities (abstract 102). Clin Cancer Res. 2000;6(suppl): 4486s. Cancer Control 35