Survey
* Your assessment is very important for improving the work of artificial intelligence, which forms the content of this project
* Your assessment is very important for improving the work of artificial intelligence, which forms the content of this project
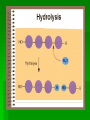
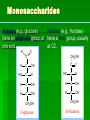
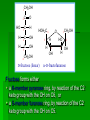
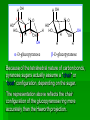
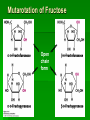
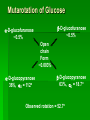
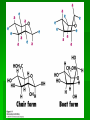
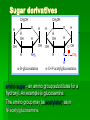
Biochemistry An Introduction to the Chemistry of Life for Students of veterinary medicine Dr Nader Vojdanifar What is Life Made of? Physical and Chemical sciences alone may not completely explain the nature of life, but they at least provide the essential framework for such an explanation. All students of VM must have a fundamental understanding of organic chemistry and biochemistry. Organic Chemistry Organic chemistry is the study of Carbon compounds. Organic compounds are compounds composed primarily of a Carbon skeleton. All living things are composed of organic compounds. Organic Chemistry What makes Carbon Special? Why is Carbon so different from all the other elements on the periodic table? The answer derives from the ability of Carbon atoms to bond together to form long chains and rings. Organic Chemistry Organic Chemistry Carbon can covalently bond with up to four other atoms. Carbon can form immensely diverse compounds, from simple to complex. Methane with 1 Carbon atom DNA with tens of Billions of Carbon atoms Biochemistry Biochemistry is a special branch of organic chemistry that deals with matter inside the living cell called Protoplasm. Protoplasm is an enormously complex mixture of organic compounds where high levels of chemical activity occur. Biochemistry How much biochemistry do you need to know for this course? 1. You need to know the structure of organic molecules important to major biological processes. 2. You will be expected to learn the basic biochemical processes of major cell functions, such as photosynthesis, respiration, and protein synthesis. Primary Organic Compounds You are expected to learn the structure and functions of these organic compounds: 1. 2. 3. 4. Carbohydrates Lipids Proteins Nucleic Acids Polymers ands Monomers Each of these types of molecules are polymers that are assembled from single units called monomers. Each type of macromolecule is an assemblage of a different type of monomer. Monomers Macromolecule Carbohydrates Monomer Monosaccharide Lipids Proteins Not always polymers; Hydrocarbon chains Amino acids Nucleic acids Nucleotides How do monomers form polymers? In condensation reactions (also called dehydration synthesis), a molecule of water is removed from two monomers as they are connected together. Hydrolysis In a reaction opposite to condensation, a water molecule can be added (along with the use of an enzyme) to split a polymer in two. Carbohydrates Carbohydrates are made of carbon, hydrogen, and oxygen atoms, always in a ratio of 1:2:1. Carbohydrates are the key source of energy used by living things. The building blocks of carbohydrates are sugars, such as glucose and fructose. Carbohydrates We may think of carbohydrates as sugar and spice and everything nice Carbohydrates What do the roots mono-, di-, oligo-, and poly mean? Each of these roots can be added to the word saccharide to describe the type of carbohydrate you have. Carbohydrates (glycans) have the following basic composition: (CH2O)n I or H - C - OH I Monosaccharides - simple sugars with multiple OH groups. Based on number of carbons (3, 4, 5, 6), a monosaccharide is a triose, tetrose, pentose or hexose. Disaccharides - 2 monosaccharides covalently linked. Oligosaccharides - a few monosaccharides covalently linked. Polysaccharides - polymers consisting of chains Monosaccharides Aldoses (e.g., glucose) Ketoses (e.g., fructose) have an aldehyde group at have a keto group, usually one end. at C2. H O CH2OH C C O HO C H OH H C OH OH H C OH H C OH HO C H H C H C CH2OH CH2OH D-glucose D-fructose How do two monosaccharides combine to make a polysaccharide? Polysaccharides homopolysaccharide heteropolysaccharide Numbers, Groups and Names “OSE” is the suffix denoting a sugar and “ULOSE” denotes a keto sugar. The monosaccharides you will encounter in biochemistry have 3, 4, 5, 6, and 7 carbons. Prefixes such as tri, tetra, penta, hexa, and hepta alert you to the number. Know these prefixes (click 1). You will also see “aldo” and “keto” to denote the type of functional group (click 1). The term aldopentose denotes two structural features, chain length and functional group. Aldo refers to aldehyde and keto to ketone. The figures show an aldopentose and a ketohexose. C C C C C C C C C C C C C C C Tri- C C C C Tetra- C C C C C Penta- Hexa- C O C H C=O C C C C C Aldopentose C C C C Ketohexose Hepta- An aldo sugar always has the “carbonyl” group on C-1, a keto sugar has it on C-2. This is good to remember. –OH groups are not indicated by this terminology. More terms are need, therefore, to describe a specific sugar. Click to go on. Balls and Sticks: All sugars have one carbonyl group and at least two –OH groups. All have at least one –CH2OH group. What distinguishes one from another is the right-left orientations of internal –OH groups. Stereochemistry is best learned by using balls and sticks. For example, D-glucose, an aldohexose, is show as (click 1). The ball represents the –CHO group, the sticks –OH groups. Because a – CH2OH group is common to all sugars, it is not necessary to draw this group each time. D-glucose L-glucose D-fructose D-mannose L-mannose D-ribose The red line shows the –OH group whose left/right orientation determines if its a D- or L-sugar. L-glucose is the mirror image of D-glucose (click 1). Focus on chiral center orientations to learn the sugars. Keto sugars can only be represented by sticks, with a = to show the keto group (click 1). Ball and stick representations are a quick way to help you see differences between sugars and imprint these in you memory (click 1) Rules for Configurations What you saw on the previous slide was the importance of –OH group orientation to pinpoint a specific sugar’s name. Now you will see that internal configurations have their own prefixes, such as “gluco, manno, galacto, fructo, etc. Configuration prefixes help you compare sugars. Here are examples of D sugar configurations (click 1). Note that glucose and galactose differ by orientation around C-4 (click 1) and glucose and mannose differ at C-2 (click 1). The other aldohexoses are allose with all –OH groups on the same side (click 1) to idose (the idiot sugar) that can decide which side to put its – OH groups (click 1). gluco ribo galacto arabino manno xylo lyxo gulo allo erythro ido threo altro talo fructo In the pentoses, one sees that ribose has all –OH groups on the same side (click 1). The tetroses differ by orientation around C-2. Fructose is set apart because of the keto group on C-2 (click 1). But, note that C-3 to C-6 of fructose have the same configuration as glucose (click 1). Click to go on. D vs L Designation CHO CHO D & L designations are based on the configuration about the single asymmetric C in glyceraldehyde. H OH HO D-glyceraldehyde OH CH2OH D-glyceraldehyde H L-glyceraldehyde CHO C C CH2OH CH2OH H The lower representations are Fischer Projections. C CHO HO C H CH2OH L-glyceraldehyde Sugar Nomenclature For sugars with more than one chiral center, D or L refers to the asymmetric C farthest from the aldehyde or keto group. Most naturally occurring sugars are D isomers. O H C H – C – OH HO – C – H H – C – OH H – C – OH CH2OH D-glucose O H C HO – C – H H – C – OH HO – C – H HO – C – H CH2OH L-glucose D & L sugars are mirror images of one another. They have the same name, e.g., D-glucose & L-glucose. Other stereoisomers have unique names, e.g., glucose, mannose, galactose, etc. O H C H – C – OH HO – C – H H – C – OH H – C – OH CH2OH D-glucose O H C HO – C – H H – C – OH HO – C – H HO – C – H CH2OH L-glucose The number of stereoisomers is 2n, where n is the number of asymmetric centers. The 6-C aldoses have 4 asymmetric centers. Thus there are 16 stereoisomers (8 D-sugars and 8 L-sugars). D vs L Classification Simplifies Names Recall, aldohexoses have 4 chiral centers, or 16 (24) stereoisomers possible. Does that mean 16 individual sugar names? No. When we denote aldohexoses as D or L, only 8 (23) D-steroisomers are possible. This is because the D, L designation fixes one of the centers. Therefore, of the 16, 8 D and 8 L will have the same name. Common aldohexoses you will encounter in your studies are D-glucose, D-mannose, and D-galactose. Know these and Dfructose (click 1). D-glucose L-glucose D-mannose D-galactose D-fructose D-ribose D-xylose D-erythrose D-threose Applying the same rule, there are 4, D-aldopentoses and 2, D-aldotetroses. The common Daldopentoses are D-ribose and D-xylose (pronounced zy-lose) (click 1). If you use your imagination you should see an X in the structure of xylose (click 1). Perhaps calling xylose the “idiot sugar of pentoses” will help you remember the structure. The aldotetroses are represented by D-erythrose and D threose (click 1). D-erythrose, like D-ribose has all –OH groups on the right. D-threose has one right and one left (another idiot sugar?). Click to go on. Terminology We end the lesson by considering the terminology that describes the properties of sugars. Understand that terminology is needed to draw comparisons between structures. So, the question you must ask is “how does this term tell me how two sugars differ? Remember isomers must have something in common as well as different. 1. Glucose and galactose are epimers (click 1). The word epimer is used when comparing sugars with multiple chiral centers. It literally says only one center is different. 2. L-glucose is the enantiomer of D-glucose (click 1) This means that one is the mirror image of the other. 3. Alpha D-glucose is the anomer of beta D-glucose (click 1) Anomers differ in the stereochemistry around the ring-forming carbon. Since alpha and beta differ in only one chiral center, anomers can also be considered epimers 4. Glucose and galactose are diastereoisomers (click 1) Diastereoisomers have different physical properties. Generally optical isomers with one chiral center differ only in the direction they rotate plane-polarized light. Diastereoisomers differ both in rotation and physical properties. D-galactose and D-glucose, for example have the same chemical formulas (C6H12O6), the same straight carbon chain and the same number of –OH groups. But, besides rotation, they also differ in melting point, solubility, heat of vaporization, etc. That is why they are considered “dia” (lit., opposed to being simple) “stereoisomers”. D-Stereochemistry of Glucose Convention for the Fischer projection: CHO H C OH HO C H Carbonyl (#1C) is at the top. OH to right = "D" OH to left = "L" H C OH H C OH Penultimate C CH2 -OH Glucose is assigned the D stereochemistry because the penultimate carbon atom is D. Hemiacetal & hemiketal formation H An aldehyde can react with an alcohol to form a hemiacetal. C H O + R' OH R' O R aldehyde C OH R alcohol hemiacetal R A ketone can react with an alcohol to form a hemiketal. C R O + "R OH R' ketone "R O C R' alcohol hemiketal OH Pentoses and hexoses can cyclize as the ketone or aldehyde reacts with a distal OH. Glucose forms an intra-molecular hemiacetal, as the C1 aldehyde & C5 OH react, to form a 6-member pyranose ring, named after pyran. 1 H HO H H 2 3 4 5 6 CHO C OH C H D-glucose C OH (linear form) C OH CH2OH 6 CH2OH 6 CH2OH 5 H 4 OH H OH 3 H O H H 1 2 OH -D-glucose OH 5 H 4 OH H OH 3 H O OH H 1 2 OH -D-glucose These representations of the cyclic sugars are called Haworth projections. H Rings Only 5-, 6-, and 7-carbon sugars form rings. The ring can either have 6 atoms (pyranose) or 5 atoms (furanose). The –OH group on a hexose will attack C-1 to form a ring (click 1). When the ring forms, a new asymmetric carbon is introduced into the molecule (click 1). The –OH on the new asymmetric carbon can be drawn so as to appear on the same side of the ring-forming oxygen (alpha sugar) (click one), or it can be drawn to be on the side away from the ring-forming oxygen (beta sugar) (click 1). O C H C-OH HO-C H OH HO *C C-OH H C C-OH HO-C HO-C C-OH C-OH C-OH C-OH C CH2OH CH2OH CH2OH alpha D-glucopyranose beta D-glucopyranose O C O You should now be able to see how all the nomenclature discussed thus far is needed to pin down a specific monosaccharide. To help you see this, consider the alternatives to alpha D-glucose. Its alpha (not beta), D (not L), gluco (not galacto, manno, fructo, etc.) pyranose (not furanose). The nomenclature is precise for just one sugar. Click one to go on. CH2OH 1 HO H H 2C O C H C OH C OH 3 4 5 6 HOH2C 6 CH2OH D-fructose (linear) H 5 H 1 CH2OH O 4 OH HO 2 3 OH H -D-fructofuranose Fructose forms either a 6-member pyranose ring, by reaction of the C2 keto group with the OH on C6, or a 5-member furanose ring, by reaction of the C2 keto group with the OH on C5. 6 CH2OH 6 CH2OH 5 H 4 OH H OH 3 H O H H 1 2 OH -D-glucose OH 5 H 4 OH H OH 3 H O OH H 1 2 H OH -D-glucose Cyclization of glucose produces a new asymmetric center at C1. The 2 stereoisomers are called anomers, & . Haworth projections represent the cyclic sugars as having essentially planar rings, with the OH at the anomeric C1: (OH below the ring) (OH above the ring). H OH H OH 4 6 H O HO HO H O HO H HO 5 3 H H 2 H OH 1 OH -D-glucopyranose H OH OH H -D-glucopyranose Because of the tetrahedral nature of carbon bonds, pyranose sugars actually assume a "chair" or "boat" configuration, depending on the sugar. The representation above reflects the chair configuration of the glucopyranose ring more accurately than the Haworth projection. Pyranose and Furanose forms Hemiacetal, Haworth structure Hemiketal, Haworth structure Haworth Structure of Glucose Draw Fischer, rotate 90o, then form the hemiacetal -anomer -anomer Haworth Structure of Fructose Ribose and Deoxyribse Haworth structures Mutarotation of Fructose Open chain form Mutarotation of Glucose -D-glucofuranose ~0.5% -D-glucofuranose ~0.5% Open chain Form ~0.003% -D-glucopyranose 36%, D = 112o -D-glucopyranose 63%, D = 18.7o Observed rotation = 52.7o Test and Extend Your Understanding Q: Don’t make the mistake of thinking that left/right orientation of the critical –OH group changes a D into an L sugar. To show this, what sugar would you form if C-5 on D-glucose was oriented to the left instead of the right? A: L-idose Q: Are D-glucose and D-ribose isomers? If so, what term describes the relationship? A: D-glucose and D-ribose are not isomers of one another because they have different chemical formulas. To be considered a structural or stereoisomer, the two molecules must have the same empirical formula but differ only in the positioning of the atoms. Q: What is the relationship between D-glucose and D-fructose? A: This is a tough call. Both have the same formula and both have a carbonyl functional group. Dglucose has an aldehyde as its functional group and D-fructose has a ketone. The two must, therefore, be considered “structural isomers” and not stereoisomers. Q: How many epimers are there of D-glucose? Of -D-glucose? A: Two, D-mannose and D-galactose. -D-glucose has 3: -D-glucose, -D-mannose, -D-galactose. Q: How many stereoisomers of a heptulose are possible? How many are D and how many are L sugars? How many names will be needed for all the isomers? (hint: the name tells you the structure of this sugar). A: A heptulose is a 7 carbon keto sugar. Therefore, it has 4 chiral centers, which means the straight chain form has 16 isomers; 8 are D and 8 are L, just like glucose. There will be 8 names needed. Sugar derivatives CH2OH CH2OH O H H OH H H OH H OH OH H NH2 -D-glucosamine O H H H O OH OH H N C CH3 H -D-N-acetylglucosamine amino sugar - an amino group substitutes for a hydroxyl. An example is glucosamine. The amino group may be acetylated, as in N-acetylglucosamine. H O H3C C O NH R H COO H R= OH H HC OH HC OH CH2OH OH H N-acetylneuraminate (sialic acid) N-acetylneuraminate (N-acetylneuraminic acid, also called sialic acid) is often found as a terminal residue of oligosaccharide chains of glycoproteins. Sialic acid imparts negative charge to glycoproteins, because its carboxyl group tends to dissociate a proton at physiological pH, as shown here. Glycosidic Bonds The anomeric hydroxyl and a hydroxyl of another sugar or some other compound can join together, splitting out water to form a glycosidic bond: R-OH + HO-R' R-O-R' + H2O E.g., methanol reacts with the anomeric OH on glucose to form methyl glucoside (methylglucopyranose). H OH H OH H2O H O HO HO H H H + CH3-OH H O HO HO H OH H OH -D-glucopyranose methanol H OH OCH3 methyl--D-glucopyranose Disaccharides: Maltose, a cleavage product of starch (e.g., amylose), is a disaccharide with an (1 4) glycosidic link between C1 - C4 OH of 2 glucoses. It is the anomer (C1 O points down). 6 CH2OH 6 CH2OH H 4 5 O H OH OH 3 H H H H 1 4 4 OH 5 H OH H OH maltose H H 1 OH OH 6 CH2OH H H 1 O 4 5 O H OH H H 3 H 2 3 O H OH O O 2 6 CH2OH H 5 2 OH 3 cellobiose H 2 OH 1 H OH Cellobiose, a product of cellulose breakdown, is the otherwise equivalent anomer (O on C1 points up). The (1 4) glycosidic linkage is represented as a zigzag, but one glucose is actually flipped over relative to the other. Other disaccharides include: Sucrose, common table sugar, has a glycosidic bond linking the anomeric hydroxyls of glucose & fructose. Because the configuration at the anomeric C of glucose is (O points down from ring), the linkage is (12). The full name of sucrose is -D-glucopyranosyl(12)--D-fructopyranose.) Lactose, milk sugar, is composed of galactose & glucose, with (14) linkage from the anomeric OH of galactose. Its full name is -D- CH2OH H O H OH H H H 1 O OH 6CH OH 2 5 O H 4 OH 3 H OH H H H H 1 O H OH CH2OH CH2OH CH2OH H H H O H OH H O O H H O H OH H H O OH 2 OH H OH H OH H OH amylose Polysaccharides: Plants store glucose as amylose or amylopectin, glucose polymers collectively called starch. Glucose storage in polymeric form minimizes osmotic effects. Amylose is a glucose polymer with (14) linkages. The end of the polysaccharide with an anomeric C1 not involved in a glycosidic bond is called the reducing end. CH2OH CH2OH O H H OH H H OH H O OH CH2OH H H OH H H OH H H OH CH2OH O H OH O H OH H H O O H OH H H OH H H O 4 amylopectin H 1 O 6 CH2 5 H OH 3 H CH2OH O H 2 OH H H 1 O CH2OH O H 4 OH H H H H O OH O H OH H H OH H OH Amylopectin is a glucose polymer with mainly (14) linkages, but it also has branches formed by (16) linkages. Branches are generally longer than shown above. The branches produce a compact structure & provide multiple chain ends at which enzymatic cleavage can CH2OH CH2OH O H H OH H H OH H O OH CH2OH H H OH H H OH H H OH CH2OH O H OH O H OH H H O O H OH H H OH H H O 4 glycogen H 1 O 6 CH2 5 H OH 3 H CH2OH O H 2 OH H H 1 O CH2OH O H 4 OH H H H H O OH O H OH H H OH H OH Glycogen, the glucose storage polymer in animals, is similar in structure to amylopectin. But glycogen has more (16) branches. The highly branched structure permits rapid glucose release from glycogen stores, e.g., in muscle during exercise. The ability to rapidly mobilize glucose is more essential to animals than to plants. CH2OH H O H OH H OH H 1 O H H OH 6CH OH 2 5 O H 4 OH 3 H H H 1 2 OH O O H OH CH2OH CH2OH CH2OH H H O O H OH H OH O H O H OH H OH H H OH cellulose Cellulose, a major constituent of plant cell walls, consists of long linear chains of glucose with (14) linkages. Every other glucose is flipped over, due to linkages. This promotes intra-chain and inter-chain H-bonds and van der Waals interactions, that cause cellulose chains to be straight & rigid, and pack with a crystalline arrangement in thick bundles - microfibrils. See: Botany online website; website at Georgia Tech. OH H H H H H Schematic of arrangement of cellulose chains in a microfibril. CH2OH H O H OH H OH H 1 O H H OH 6CH OH 2 5 O H 4 OH 3 H H H 1 2 OH O O H OH CH2OH CH2OH CH2OH H H O O H OH H OH O H O H OH H OH OH H H H H H H H OH cellulose Multisubunit Cellulose Synthase complexes in the plasma membrane spin out from the cell surface microfibrils consisting of 36 parallel, interacting cellulose chains. These microfibrils are very strong. The role of cellulose is to impart strength and rigidity to plant cell walls, which can withstand high hydrostatic pressure gradients. Osmotic swelling is prevented. Explore and compare structures of amylose & cellulose using Chime. CH 2OH D-glucuronate 6COO H 4 6 5 H OH 3 H H 2 OH 1 H H OH O O H 4 O H 5 3 H 2 1 O H NHCOCH 3 N-acetyl-D-glucosamine hyaluronate Glycosaminoglycans (mucopolysaccharides) are linear polymers of repeating disaccharides. The constituent monosaccharides tend to be modified, with acidic groups, amino groups, sulfated hydroxyl and amino groups, etc. Glycosaminoglycans tend to be negatively charged, because of the prevalence of acidic groups. CH 2OH D-glucuronate 6 6COO H 4 5 H OH 3 H H 2 OH 1 H H OH O O H 4 O H 5 3 H 2 1 O H NHCOCH 3 N-acetyl-D-glucosamine hyaluronate Hyaluronate (hyaluronan) is a glycosaminoglycan with a repeating disaccharide consisting of 2 glucose derivatives, glucuronate (glucuronic acid) & N-acetylglucosamine. The glycosidic linkages are (13) & (14). core protein heparan sulfate glycosaminoglycan transmembrane -helix cytosol Proteoglycans are glycosaminoglycans that are covalently linked to serine residues of specific core proteins. The glycosaminoglycan chain is synthesized by sequential addition of sugar residues to the core protein. Some proteoglycans of the extracellular matrix bind non-covalently to hyaluronate via protein domains called link modules. E.g.: • Multiple copies of the aggrecan proteoglycan associate with hyaluronate in cartilage to form large complexes. • Versican, another proteoglycan, binds hyaluronate in the extracellular matrix of loose connective tissues. Websites on: Aggrecan Aggrecan & versican. CH 2OH D-glucuronate 6 6COO H 4 5 H OH 3 H hyaluronate H 2 OH 1 H H OH O O H 4 O H 5 3 H 2 1 O H NHCOCH 3 N-acetyl-D-glucosamine N-sulfo-glucosamine-6-sulfate iduronate-2-sulfate CH2OSO3 H H COO OH O O H O H H OH H H H H OSO3 O H NHSO3 heparin or heparan sulfate - examples of residues Heparan sulfate is initially synthesized on a membrane-embedded core protein as a polymer of alternating N-acetylglucosamine and glucuronate residues. Later, in segments of the polymer, glucuronate residues may be converted to the sulfated sugar iduronic acid, while N-acetylglucosamine residues Heparin, a soluble glycosaminoglycan found in granules of mast cells, has a structure similar to that of heparan sulfates, but is more highly sulfated. PDB 1RID When released into the blood, it inhibits clot formation by interacting with the protein antithrombin. heparin: (IDS-SGN)5 Heparin has an extended helical conformation. C O N S Charge repulsion by the many negatively charged groups may contribute to this conformation. Heparin shown has 10 residues, alternating IDS (iduronate-2-sulfate) & SGN (N-sulfo-glucosamine-6sulfate). Some cell surface heparan sulfate glycosaminoglycans remain covalently linked to core proteins embedded in the plasma membrane. core protein heparan sulfate glycosaminoglycan transmembrane -helix cytosol The core protein of a syndecan heparan sulfate proteoglycan includes a single transmembrane helix, as in the simplified diagram above. The core protein of a glypican heparan sulfate proteoglycan is attached to the outer surface of the plasma membrane via covalent linkage to a modified phosphatidylinositol lipid. Proteins involved in signaling & adhesion at the cell surface recognize & bind heparan sulfate chains. E.g., binding of some growth factors (small proteins) to cell surface receptors is enhanced by their binding also to heparan sulfates. Regulated cell surface Sulf enzymes may remove sulfate groups at particular locations on heparan sulfate chains to alter affinity for signal proteins, e.g., N-sulfo-glucosamine-6-sulfate iduronate-2-sulfate CH OSO H growth factors. O O 2 H Diagram by Kirkpatrick & Selleck. H H COO OH 3 O H H OH H H H OSO3 O H NHSO3 heparin or heparan sulfate - examples of residues Oligosaccharides that are covalently attached to proteins or to membrane lipids may be linear or branched chains. C CH2OH O H H OH O CH2 CH NH H O serine residue O H OH H HN C CH3 -D-N-acetylglucosamine O-linked oligosaccharide chains of glycoproteins vary in complexity. They link to a protein via a glycosidic bond between a sugar residue & a serine or threonine OH. O-linked oligosaccharides have roles in recognition, interaction, and enzyme regulation. C CH2OH O H H OH O CH2 CH NH H O serine residue O H OH H HN C CH3 -D-N-acetylglucosamine N-acetylglucosamine (GlcNAc) is a common O-linked glycosylation of protein serine or threonine residues. Many cellular proteins, including enzymes & transcription factors, are regulated by reversible GlcNAc attachment. Often attachment of GlcNAc to a protein OH alternates with phosphorylation, with these 2 modifications having opposite regulatory effects (stimulation or inhibition). CH2OH O O H H OH HN C HN CH2 C H H OH H HN C CH3 O N-acetylglucosamine Initial sugar in N-linked glycoprotein oligosaccharide Asn CH O HN HC R C O X HN HC R C O Ser or Thr N-linked oligosaccharides of glycoproteins tend to be complex and branched. First N-acetylglucosamine is linked to a protein via the side-chain N of an asparagine residue in a particular 3-amino acid sequence. NAN NAN NAN Gal Gal Gal NAG NAG NAG Man Man Man Key: NAG NAG Asn N-linked oligosaccharide Fuc NAN = N-acetylneuraminate Gal = galactose NAG = N-acetylglucosamine Man = mannose Fuc = fucose Additional monosaccharides are added, and the Nlinked oligosaccharide chain is modified by removal and addition of residues, to yield a characteristic branched structure. Many proteins secreted by cells have attached N-linked oligosaccharide chains. Genetic diseases have been attributed to deficiency of particular enzymes involved in synthesizing or modifying oligosaccharide chains of these glycoproteins. Such diseases, and gene knockout studies in mice, have been used to define pathways of modification of oligosaccharide chains of glycoproteins and glycolipids. Carbohydrate chains of plasma membrane glycoproteins and glycolipids usually face the outside of the cell. They have roles in cell-cell interaction and signaling, and in forming a protective layer on the surface of some cells. Lectins are glycoproteins that recognize and bind to specific oligosaccharides. Concanavalin A & wheat germ agglutinin are plant lectins that have been useful research tools. The C-type lectin-like domain is a Ca++-binding carbohydrate recognition domain in many animal lectins. Recognition/binding of CHO moieties of glycoproteins, glycolipids & proteoglycans by animal lectins is a factor in: • cell-cell recognition • adhesion of cells to the extracellular matrix • interaction of cells with chemokines and growth factors • recognition of disease-causing microorganisms • initiation and control of inflammation. Examples of animal lectins: Mannan-binding lectin (MBL) is a glycoprotein found in blood plasma. It binds cell surface carbohydrates of diseasecausing microorganisms & promotes phagocytosis of these organisms as part of the immune response. Selectins are integral proteins of mammalian cell plasma membranes with roles in cell-cell recognition & binding. selectin lectin domain outside The C-type lectin-like domain transmembrane is at the end of a multi-domain -helix cytosol extracellular segment cytoskeleton binding domain extending out from the cell surface. A cleavage site just outside the transmembrane -helix provides a mechanism for regulated release of some lectins from the cell surface. A cytosolic domain participates in regulated interaction with the actin cytoskeleton. Lipids Lipids are molecules that consist of long hydrocarbon chains. Attaching the three chains together is usually a glycerol molecule. Lipids are NONpolar. Saturated vs. Unsaturated Fat Proteins Proteins are building blocks of structures called amino acids. Proteins are what your DNA codes to make (we will talk about this in great detail in a month or so). A peptide bond forms between amino acids by dehydration synthesis. Levels of Protein Structure Protein Structure Level Primary Secondary Description The amino acid sequence Helices and Sheets Disulfide bridges Tertiary Quaternary Multiple polypeptides connect