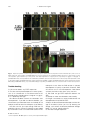
Survey
* Your assessment is very important for improving the workof artificial intelligence, which forms the content of this project
Develop. Growth Differ. (2008) 50, 459–465 doi: 10.1111/j.1440-169x.2008.01054.x Review Blackwell Publishing Asia Congenic method in the chick limb buds by electroporation Takayuki Suzuki* and Toshihiko Ogura Institute of Development, Aging and Cancer (IDAC), Tohoku University, Seiryo-cho 4-1, Aoba-ku, Sendai, 980-8575, Japan Electroporation is a powerful tool with which to study limb development. Limb development, however, remains an intricate series of events, requiring the precise dissection of developmental processes using relevant transgenes. In this review, we describe the anatomy of the limb field as the basis of targeted electroporation, and specific expression vectors are discussed. We share a useful protocol for electroporation of chick limb buds, and the expression pattern of enhanced green fluorescent protein in the limb buds is used to demonstrate relevant embryonic patterning. Finally, useful trouble-shooting techniques are described. Key words: chick embryo, electroporation, limb buds, limb field. Introduction The developing limb bud is an excellent model with which to study molecular mechanisms of embryonic patterning and tissue differentiation in vertebrates. By exploiting both classical and modern embryological methods, combined with novel techniques in molecular biology, data regarding limb development is rapidly accumulating. Classical embryological experiments, such as apical ectodermal ridge (AER) removal from the chick limb bud, revealed the presence of a key signaling pathway, and ultimately resulted in the discovery of the fibroblast growth factor (FGF) cascade. FGF interacts with the zone of polarizing activity (ZPA) (Saunders & Gasseling 1968), a region located at the posterior side of the limb bud specifying posterior limb bud identity, discovered by another classic embryological experiment, the anterior implantation of a tissue graft from the posterior chick limb bud. Thus two opposite approaches, loss-of-function and gain-of-function experiments, are necessary to dissect the molecular nature of these signal transduction pathways. In the mouse, targeted gene disruption is a powerful loss-of-function approach. For example, knocking-out FGF10 resulted in the total loss of limb buds in the *Author to whom all correspondence should be addressed. Email: [email protected] Received 14 March 2008; revised 4 April 2008; accepted 1 May 2008. © 2008 The Authors Journal compilation © 2008 Japanese Society of Developmental Biologists mouse (Sekine et al. 1999). Since the first report of gene targeting was published (Thomas & Capecchi 1987), this technique has been an invaluable tool for developmental biologists, providing essential insights into the developmental function of embryonic genes. Nonetheless, knockout mice often show lethality during early stages of development, making it difficult to study gene function during organogenesis at later stages, such as limb development. To overcome this problem, the conditional knock-out technique was developed using tissue-specific expression of the Cre recombinase; today this approach is limited only by the availability of tissue-specific promoters. In contrast to the genetic approaches developed in mice, two different approaches have been developed in chick models to introduce transgenes in a spatially and temporally controlled manner; one using retroviruses, the other, electroporation. Electroporation has several advantages over retrovirus-mediated gene transfer, including the ability to rapidly express cDNA after transient cell membrane disruption. Electroporation is also able to introduce larger constructs into cells than retrovirus-mediated techniques. However, retrovirusdependent methods feature the advantage of affording researchers the ability to misexpress transgenes stably even at later stages of limb development, making this system versatile and applicable in a wide range of scenarios in developmental biology. In this review, we will describe in detail how to choose expression vectors, as well as efficient methods for using electroporation to introduce transgenes of interest into chick limb buds. 460 T. Suzuki and T. Ogura Fig. 1. Anatomy of the limb field. (A) Forelimb field and hindlimb field are shown in orange and green, respectively, at St. 14. These are located on the lateral aspect of the somite, called the lateral plate mesoderm (LPM). (B) A horizontal section of a St. 14 forelimb field is shown. Forelimb and hindlimb buds develop from the dorsal side of the LPM, called the somatic LPM. There is embryonic coelom between the somatic and splanchnic LPM. D, dorsal side; V, ventral side. Limb development in the chick embryo Limb development begins as a small budding of the lateral plate mesoderm (LPM) (Fig. 1A). In the chick embryo, the forelimb bud starts to develop from a restricted part of the LPM at the 15–20 somite level, whereas the hindlimb develops at the 26–32 somite level. The LPM consists of two different parts, namely the somatic LPM on the dorsal side and the splanchnic LPM on the ventral side (Fig. 1B), separated by an embryonic space (coelom) (Fig. 1B). Both forelimb and hindlimb develop from two distinct fields in the somatic LPM, in which FGF10 starts to be expressed at St. 14 to induce cell proliferation and limb bud outgrowth. Hence, one can identify the limb fields in embryos by visualizing them with an FGF10 probe. After the initiation of limb development, threedimensional axes are specified; namely, the proximal-distal (PD), dorsal-ventral (DV) and anterior-posterior (AP) axes. Differentiation of cells and subsequent pattern formation proceeds in a temporally and spatially regulated manner along these axes at later stages. For example, mesenchymal cells condense to form cartilage, followed by the migration of progenitor cells and their differentiation into muscles, tendons, and neurons, making limb buds highly complex precursors to such intricate structures as wings/hands (forelimb) and legs (hindlimb). How to choose expression vectors for electroporation We use two different expression vectors for misexpression in embryos. One is pCAGGS (Niwa et al. 1991), a transient expression vector constructed from an enhancer of the human cytomegalovirus (CMV) immediate early region and a chick beta-actin promoter. Another vector is pRCASBP, an avian replication-competent retrovirus vector (Morgan & Fekete 1996). pCAGGS drives a more robust expression of transgenes compared with pRCASBP, thus we observe misexpression of transgenes within 3 h when pCAGGS expression is monitored by enhanced green fluorescent protein (EGFP). Nonetheless, expression is transient with maximal transcription occurring 6–24 h after electroporation, followed by a gradual decrease in the level of expression, probably due to cell proliferation and concomitant dilution of the electroporated plasmids. Therefore, the pCAGGS vector is suitable to study the embryonic limb initiation stage and limb bud stage (Takeuchi et al. 2003). The utility of pRCASBP is quite distinct. Since pRCASBP is a competent viral vector, transcripts from electroporated DNA and their transcribed and translated protein products reconstitute infectious virions, which then induce secondary infection in neighboring cells when a competent chick host is used. This cycle of infection results in a wide spread of transgene expression at later stages. For this reason, initial expression of transgene is relatively slow and weak when compared with pCAGGS. However, an active expansion of transgene expression is observed in tissues where cells divide quickly, such as the progress zone of limb buds, and we usually obtain expression in the entire limb bud one day after electroporation. Therefore, the RCASBP vector is suitable for study of later stages of limb development such as skeletal patterning and tissue differentiation (Takeuchi et al. 1999). Once the RCASBP genome is integrated by the active virion, expression is stable and lasts for the entire processes of limb development. In contrast to pCAGGS, the RCASBP vector has a limited capacity for transgene inclusion (approximately 2.4 kb), since a longer insert inhibits the incorporation of the viral RNA genome into virions, and thereby prevents secondary infection and spread of the expression domain. Such limitations are also observed when virus-incompetent chick hosts are used. However, because stable expression of transgenes can be observed in subpopulations of cells even in this case, this indicates that electroporated RCASBP plasmids can be integrated into the host genome. In the typical protocol used for the pRCASBP vector, infectious virions are produced from CEF (chick embryonic fibroblast) cells after transfection, and then concentrated to a titer sufficient for effective infection after the injection of virions. Since the transgene is expressed in CEF in this step of viral propagation, it is not easy to obtain a high titer virus concentrate when the transgene is toxic to CEF. We often encounter this problem, especially when we use genes related to © 2008 The Authors Journal compilation © 2008 Japanese Society of Developmental Biologists Electroporation into chick limb buds apoptosis and bone morphogenetic protein (BMP) signaling. Nonetheless, by directly electroporating a toxic transgene-containing pRCASBP vector into the limb bud, we can overcome this difficulty, although we anticipate a slow expansion of transgene expression in this case. Combination of expression vectors and other useful vectors Another advantage of electroporation includes the ability to co-electroporate two or three expression plasmids simultaneously. This can be used for the visualization of an electroporated area, by mixing pCAGGS-EGFP or pCMV-EGFP vectors in a plasmid cocktail and subsequent expression of GFP fluorescent signals in limb bud. Using our techniques, we can successfully misexpress two or three transgenes in the limb bud in a single electroporation experiment. This is a significant advantage for us, since combinatorial actions of genes, for example transcription factors, are observed in vivo. In the RCASBP system, there are available subgroups A, B, D, and E (Logan & Tabin 1998). RCASBP subgroup A, B, and D could be infected in C/O (cells susceptible to all subgroups of avian leukosis viruses) egg limbs (Suzuki et al. unpubl. data, 2008), so it is possible to overexpress at least three transgenes at the same time using the RCASBP system. Protocols for electroporation into limb Egg incubation and exposure CUY21 electroporator (Nepa gene, Ichikawa, Japan), dissecting microscope (Leica MZAPO, Wetzlar, Germany), and platinum electrodes (CUY613P2, Nepa gene) are set up (Fig. 2A). Incubate eggs in a humidified incubator (IC800, Yamato, Tokyo, Japan) at 39°C until St. 13/14, kept down for easy access (Fig. 2B). One can also position eggs in a top-down manner, but this positioning might not provide enough of a window to inject plasmids and an efficient configuration of electrodes on the surface of the embryo. In addition, a flat aspect of the egg can be patched much more easily than the top of the egg using plastic tape. Complete sealant of windows is one critical factor necessary to ensure high viability of the embryos, preventing bacterial and/or fungal contamination after electroporation. After incubating eggs until St. 13/14, an 18G1/2 needle (NN-1838R, Terumo, Tokyo, Japan) or forceps (Dumont No. 5 forceps, 11 cm; No. 500341, World precision instruments) should be used to create a window at the broader edge of the eggshell. Approximately 5 mL of albumin should be gently removed from this hole with a 25-mL syringe (SS-20ESZ, Terumo). Using 461 this procedure, one can procure enough space inside the egg for electroporation. This tiny hole can then be immediately sealed with plastic tape (Fig. 2B). Subsequently, one can obtain access by cutting the eggshell with a (pair of) curved scissors. One should be sure that this window is wide enough for electroporation, while avoiding the creation of a window that is too wide, approximately bigger than 4 cm2, which could instead damage the embryo and/or lead to infection. Through this window, one can observe the living embryo. Preparation of tungsten needles and glass capillaries Chick embryos develop between the yolk sac and vitelline membrane. The membrane must be removed to expose the surface of the embryo using a tungsten needle, which has been sharpened precisely for this undertaking. For sharpening, tungsten wire tips 0.20 mm in diameter (No. 461267, Nilaco Co., Tokyo, Japan) are immersed in 1N NaOH. Electric current is then applied at 23 V. This method yields sharp tips, which can then be checked under a microscope. Glass capillaries for DNA injection are made by pulling Narishige glass capillaries (1 × 90 mm glass capillaries with filament; GPC-1, Narishige, Tokyo, Japan) with a Narishige puller (PC-10, Narishige), pre-adjusted for the desired degree of sharpness. There are three kinds of weights in the Narishige puller; two large weights (93 g) and one smaller weight (24 g), which are used to produce different pulled capillary tubes according to varying specifications. We typically use one large weight and one small weight. Using this combination, we can make glass capillaries with a natural curve after pulling. These curved capillaries are very useful for injecting DNA solution into a thinner body wall. An alternative means of producing curved capillaries involves bending the tip of glass capillaries with no. 5 forceps immediately after pulling. The curved needle tip is broken off by a no. 5 forceps under a dissecting microscope, and placed at the end of an aspirator tube. DNA solution preparation The pCAGGS or RCASBP vector is next diluted with distilled water to a final concentration of 3–5 μg/μL and mixed with pCAGGS-EGFP expression vector (0.8 –1.0 μg/μL) for electroporation. This concentration of the pCAGGS-EGFP vector is enough to visualize the entire electroporated area (Momose et al. 1999). Viscous DNA in solution (a concentration greater than 5 μg/μL) decreases the efficiency of electroporation and the subsequent expression of the transgene of interest. A cocktail of expression plasmids can then be placed on the cap of a PCR tube (0.2 mL), and mixed © 2008 The Authors Journal compilation © 2008 Japanese Society of Developmental Biologists 462 T. Suzuki and T. Ogura Fig. 2. Electroporation into the limb field. (A) CUY21 electroporator, dissecting microscope, and platinum electrodes are set up. (B) Anode electrode and cathode electrode are shown on the right and left, respectively. The egg is arranged lying down. (C) After injection of Rotring-phosphate-buffered saline (PBS) solution, a St. 14 chick embryo is highlighted on the black background. (D) Forelimb and hindlimb fields are shown by a yellow circle. (E) Expression pattern of cfgf10 at St. 14 in a horizontal section of the forelimb field. cfgf10 is expressed at the somatic lateral plate mesoderm (LPM) and nephric primordium. (F) A cathode electrode is inserted under the embryo. (G) A cathode electrode is placed under the forelimb field. (H) The vitelline artery is labeled by India ink injection. (I) Fast Green-DNA solution is injected into the forelimb field. (J) An anode electrode is prepared on the forelimb field. The anode electrode is placed in the center of the forelimb field (K), anterior (L), posterior (M), or parallel to cathode electrode (N). (O) The hindlimb field is shown by a yellow circle. (P) Fast Green-DNA solution is injected into the hindlimb field. (Q) An anode electrode is placed in the center of the hindlimb field. with a solution of 1% Fast Green (diluted in PBS; No. 061-00031, Wako, Osaka, Japan) before electroporation. Coloring the DNA solution is important to obtain high electroporation efficiency. However, a concentration of Fast Green that is too high, greater than 1%, might decrease the efficiency of transfection. This electroporation cocktail is then vacuum-extracted by a curved glass capillary attached to an aspirator tube. Injection of DNA solution into limb fields After opening an eggshell, one can observe the chick embryo under the vitelline membrane. At this moment, a small amount of a Rotring-phosphate-buffered saline (PBS) solution (PBS with 1:40 Rotring; Art-R 591017, Sanford Co., Tokyo, Japan) (approximately 100–200 μL) is injected from outside of the sinus terminalis located near the tail bud, using a syringe equipped with a 26G1/2 needle (NN-2613S, Terumo). This produces a black background, which provides a higher degree of contrast with which to visualize the embryo (Fig. 2C). After bathing the embryo in 1 mL of sterilized PBS, vitelline membrane near either the forelimb or hindlimb field is gently shorn by a sharpened tungsten needle (Fig. 2D). At St. 14 (the best stage for electroporation into limb buds) 22 pairs of somites © 2008 The Authors Journal compilation © 2008 Japanese Society of Developmental Biologists Electroporation into chick limb buds are present. DNA solution is injected into an embryonic space located between the somatic LPM and the splanchnic LPM (Fig. 2E). An L-shaped platinum cathode is inserted from the hole that was created for injection of the Rotring-PBS solution (Fig. 2F), and placed under either the forelimb (Fig. 2G) or hindlimb field (Fig. 2O). As described above, the forelimb field forms at the 15–20 somite level of the LPM, spanning from an extended edge of the anterior LPM to a location corresponding to the position of the vitelline artery (Fig. 2H). At this stage, it is not easy to locate the hindlimb field by counting somites. Instead, one can gauge its position by checking the expression domain of cfgf10, which spans approximately six somite lengths from the posterior edge of the LPM, in the boundary between the body wall and amnion. To inject DNA solution into the forelimb field, the tip of a curved glass capillary tube is inserted from the anterior side of the forelimb field by pricking the thin embryonic tissue. For injection into the hindlimb field, a glass capillary tip is inserted from the posterior side of the hindlimb field. Care is required to avoid any damage to the vitelline artery, which typically results in the death of an embryo. After insertion of the needle, DNA solution can be injected. After successful injection, one can observe green pigment in the Fast Green-filling forelimb (Fig. 2I) or hindlimb field (Fig. 2P). Electroporation into limb fields We developed three different methods by modifying our original protocol (Ogura 2002), typically using a platinum anode (Fig. 2J). 1. When an anode is placed at the center of either the forelimb (Fig. 2K) or the hindlimb field (Fig. 2Q) before electric pulses (8 V, 60 ms pulse-on, 50 ms pulse-off, three repetitions), one can express transgene along the entire limb bud, with strong expression along the middle part of the limb. By positioning an anode at either the anterior side (Fig. 2L) or the posterior side of the limb field (Fig. 2M), transgene expression becomes restricted to the anterior or posterior portions, respectively. 2. One can expand the domain of transgene expression by moving an anode serially. In this case, the electroporator is set for three pulses (8 V, 60 ms pulse-on, 1 second pulse-off, for three repetitions). For the first pulse, an anode is placed in the anterior limb field (Fig. 2L). During a 1 second pulse-off pause, this anode is moved to the central part of the limb field (Fig. 2K), and then a second 463 pulse is applied. The anode is moved further to the posterior limb field (Fig. 2M) for the last electric pulse. By applying three electric pulses serially in different parts of the limb field, one can overexpress transgene strongly and uniformly in the whole limb bud. 3. An anode can be placed over the limb field, making a parallel configuration with a cathode (Fig. 2N) for electric pulses (5 V, 60 ms pulse-on, 50 ms pulse-off, three repetitions). In this setting, one can misexpress transgene throughout the entire limb bud. However, lower voltage pulses must be used, since the electric field formed between two parallel electrodes is wider and a higher voltage in this setting may damage cells. During electroporation, two electrodes must be kept in solution, not touching the surface of the embryos or the vitelline artery in order to avoid tissue damage. These methods are efficient at somatic LPM from St. 12 to St. 16. At later stages, from St. 17, it is difficult to keep DNA solution inside of the limb bud. One may be able to electroporate at later stages using these methods, but expression of the electroporated plasmid would be weaker. When truncation or shortening of the limb buds is observed, even after electroporation with non-toxic pCAGGS-EGFP, this finding must be replicated in order to confirm the electrodes were free from the embryonic tissues, and not affecting the experimental results. After electroporation, the cathode is gently withdrawn from the amnion, and 30 μL of a penicillin-streptomycin solution (PBS with 1:100 penicillin-streptomycin; No. 15140-122, Invitrogen, Carlsbad, CA) (for 1 L 1 x PBS: NaCl 5.8 g, NaH2PO42H2O 0.36 g, Na2HPO412H2O 2.76 g) is used to bathe the embryo. The eggshell window is then sealed firmly with plastic tape. Embryos should be incubated again immediately after sealing. When pCAGGS-EGFP (5 μg/μL) is electroporated into the forelimb field, EGFP expression can be detected after 1–3 h (Fig. 3A–D). EGFP expression becomes stronger at 6–12 h after electroporation (Fig. 3E–H), and expands until 24–48 h (Fig. 3I–L), probably due to proliferation and migration of cells. When a lower concentration of pCAGGS-EGFP (0.1 μg/μL) is used, weak expression of EGFP can be detected at 12 h, whereas electroporation of 1 μg/μL of pCAGGS-EGFP results in an intermediate level of expression (Fig. 3M–P). Robust EGFP expression can be observed in the hindlimb field at 12 h after electroporation of 5 μg/μL pCAGGS-EGFP (Fig. 3Q,R). By changing the positions of an anode as shown in Figure 2K–M, one can target expression of the transgene of interest to either the anterior (Fig. 3S), middle (Fig. 3T) or posterior side (Fig. 3U) of the limb bud. © 2008 The Authors Journal compilation © 2008 Japanese Society of Developmental Biologists 464 T. Suzuki and T. Ogura Fig. 3. Expression pattern of enhanced green fluorescent protein (EGFP) after electroporation into the limb field. (A–L) Time course of EGFP expression after 5 μg/μL of pCAGGS-EGFP was electroporated into the forelimb field. EGFP expression is detected at 3 h after electroporation (yellow arrow in C). Lower panels show high magnification of upper panels, respectively. 0.1 μg/μL or 1.0 μg/μL of pCAGGS-EGFP was electroporated into the forelimb field, and EGFP expression was detected 12 h after electroporation (M and N, O and P). (Q and R) 5 μg/μL of pCAGGS-EGFP was electroporated into the hindlimb. 5 μg/μL of pCAGGS-EGFP was electroporated into the anterior limb field (S), middle limb field (T), or posterior limb field (U), respectively, and EGFP fluorescence was detected 24 h after electroporation. Trouble shooting Q: We cannot detect any EGFP expression. A: First, be sure the plasmid expresses in other systems, such as in cultured cells. Is the concentration of the plasmid correct and adjusted to a range of 3–5 μg/μL, or is it potentially too high? We observe bubbles around the electrodes after electroporation. If you did not observe bubbles, you should move your electrode closer to the body of the embryo, but do not touch the embryonic tissues directly. Also, be sure that you inject enough DNA into the embryonic column. Successful injection can be judged by visualization of the Fast Green-DNA cocktail in the embryonic tissues. After the DNA solution is injected, electroporate as quickly as possible. Otherwise, DNA will diffuse out of the electroporation field before sufficient plasmid can be taken up by the cells. Q: We could not get EGFP expression beneath the AER. A: Attempt to move the electrodes more laterally. Q: We got a truncated or a shorter limb bud when we electroporated with pCAGGS-EGFP. A: Adjust the distance between electrodes and the voltage of the electric pulses you use. By lowering the voltage, such artifacts will often be prevented. Voltages below 5 V are not typically effective. Q: We need stronger expression. © 2008 The Authors Journal compilation © 2008 Japanese Society of Developmental Biologists Electroporation into chick limb buds A: Move the electrodes closer to the embryos and/or increase the electroporation voltage. If the concentration of plasmid is less than 5 μg/μL, concentrate it. Since the pCAGGS vector shows the most effective transgene transcription in the limb, use this vector. Q: Malformations developed outside of the limb. A: Your electrodes are likely too close to the midline tissue of the embryo. Move your electrodes to a more lateral position. Arrange two electrodes closer together to create a narrower pulse field and apply lower voltages. Q: Our embryos do not survive well after electroporation. A: It is best to check viability of the embryos 24 h after electroporation. If you see malformation even at this early time point, your electroporation setting is probably too strong. Check your electroporation conditions using the points described above. If no malformation develops, there might instead be a problem(s) with the egg, incubation or sealant of the eggshell. Make sure that you use fresh eggs, re-incubate them immediately after electroporation and seal the eggshell window completely. Acknowledgments Special thanks to Minoru Omi for helpful discussions. This work was funded by Japan Society for the Promotion of Science to T.S., and Takeda Science Foundation to T.O. 465 References Logan, M. & Tabin, C. 1998. Targeted gene misexpression in chick limb buds using avian replication-competent retroviruses. Methods 14, 407– 420. Momose, T., Tonegawa, A., Takeuchi, J., Ogawa, H., Umesono, K. & Yasuda, K. 1999. Efficient targeting of gene expression in chick embryos by microelectroporation. Dev. Growth. Differ. 41, 335 –344. Morgan, B. A. & Fekete, D. M. 1996. Manipulating gene expression with replication-competent retroviruses. Methods Cell Biol. 51, 185 –218. Niwa, H., Yamamura, K. & Miyazaki, J. 1991. Efficient selection for high-expression transfectants with a novel eukaryotic vector. Gene 108, 193 –199. Ogura, T. 2002. In vivo electroporation: a new frontier for gene delivery and embryology. Differentiation 70, 163 –171. Saunders, J. W. Jr & Gasseling, M. T. 1968. ‘Epitherial-Mesenchymal Interractions’ (eds R. Fleischmajer & R. F. Billingham), pp. 78 –97. Williams & Wilkins, Baltimore. Sekine, K., Ohuchi, H., Fujiwara, M. et al. 1999. Fgf10 is essential for limb and lung formation. Nat. Genet. 21, 138 –141. Takeuchi, J. K., Koshiba-Takeuchi, K., Matsumoto, K. et al. 1999. Tbx5 and Tbx4 genes determine the wing/leg identity of limb buds. Nature 398, 810 – 814. Takeuchi, J. K., Koshiba-Takeuchi, K., Suzuki, T., Kamimura, M., Ogura, K. & Ogura, T. 2003. Tbx5 and Tbx4 trigger limb initiation through activation of the Wnt/Fgf signaling cascade. Development 130, 2729 –2739. Thomas, K. R. & Capecchi, M. R. 1987. Site-directed mutagenesis by gene targeting in mouse embryo-derived stem cells. Cell 51, 503 –512. © 2008 The Authors Journal compilation © 2008 Japanese Society of Developmental Biologists