Survey
* Your assessment is very important for improving the workof artificial intelligence, which forms the content of this project
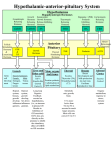
Effects of plasmid-mediated growth hormone releasing hormone supplementation in young, healthy Beagle dogs1 R. Draghia-Akli2, K. K. Cummings, A. S. Khan, P. A. Brown, and R. H. Carpenter ADViSYS, Inc., The Woodlands, TX 77381 ABSTRACT: Our study focused on the evaluation of the pharmacological and toxicological effects of plasmid-mediated GHRH supplementation with electroporation in normal adult dogs over a 180-d period. Twentyeight dogs (<2 yr of age) were randomized to four groups. Three groups (four dogs/sex for each group) were treated with ascending doses of GHRH-expressing plasmid: 0.2, 0.6, and 1 mg. One group (two dogs of each sex) served as the control. Clinical observations and body weights were recorded. Hematological, serum biochemical, and urine analyses were performed. Serum IGF-I, ACTH, and insulin were determined. Necropsies were performed on d 93 and 180; organs were weighed and tissues were fixed and processed for light microscopy. Selected tissues were used to assess plasmid biodistribution on d 93. At all doses, plasmid GHRH caused increased weight gain (P < 0.001), without organomegaly. Serum glucose and insulin in fasted dogs remained within normal ranges at all time points. Adrenocorticotropic hormone was normal in all groups. Significant increases in number of red blood cells, hematocrit, and hemoglobin (P < 0.01) were observed. In conclusion, our study shows that plasmid-mediated GHRH supplementation is safe in electroporated doses up to 1.0 mg in young healthy dogs. Key Words: Dogs, Electroportation, Insulin-Like Growth Factor, Plasmids, Somatoliberin, Somatotropin 2003 American Society of Animal Science. All rights reserved. Introduction Regulated GH secretion is essential for optimal linear growth, homeostasis of carbohydrate, protein, and fat metabolism, and for the promotion of a positive nitrogen balance (Murray and Shalet, 2000). These effects are both direct and mediated by IGF-I, the downstream effector of GH. The GH synthesis and secretion from the anterior pituitary are stimulated by GHRH, a hypothalamic peptide hormone (Muller et al., 1999). Although administration of exogenous recombinant GH (rGH) produces anabolic effects in a variety of situations, rGH therapy has disadvantages. An alternate 1 The authors would like to particularly thank D. Kern for his support of this work, C. Tone for the editorial correction of this manuscript, and to the members of our research team for their input during this study: M. Pope, L. A. Hill, and B. Malone. Our greatest appreciation for the staff at Stillmeadow, Inc., for their professionalism, and especially to A. Perez de Leon. We would also like to thank D. Hildebrandt at Pathco, Inc., for the histological analysis of the tissues. We are grateful to T. Spencer at Texas A&M University for his PCR tissue analysis. We acknowledge support for this study from ADViSYS, Inc. (The Woodlands, TX). 2 Correspondence: 2700 Research Forest Dr., Suite 180 (phone: 281296-7300, ext. 107, fax: 281-296-7333, E-mail: ruxandradraghia@ advisys.net). Received March 24, 2003. Accepted May 21, 2003. J. Anim. Sci. 2003. 81:2301–2310 method to increase GH production and release would be to administer GHRH. In a 56-d proof-of-concept study, we observed anabolic effects, correction of anemia and weight loss, and improved quality of life in dogs with cancer following injection of a GHRH-expressing plasmid (Draghia-Akli et al., 2002a). The objective of the present experiment was to determine whether a plasmid-mediated GHRH therapy would be nontoxic and would produce long-term beneficial effects at different dosages. This pilot study was conducted in young healthy male and female Beagle dogs. Follow-up evaluations demonstrated increased weight and serum IGFI concentrations as an indicator of GHRH activity, together with significantly increased red blood cell production. Necropsy and histopathology evaluation showed no adverse effects associated with plasmid delivery and GHRH expression over a period of 180 d. These results suggest that plasmid-mediated GHRH therapy can be a viable alternative to injectable recombinant protein therapies. Materials and Methods Deoxyribonucleic Acid Constructs The plasmid pSPc5-12 contains a 360-bp SacI/BamHI fragment of the SPc5-12 synthetic promoter (Li et al., 1999) in the SacI/BamHI sites of a pSK-GHRH back- 2301 2302 Draghia-Akli et al. bone (Draghia-Akli et al., 1997). The synthetic GHRH complementary DNA (cDNA), HV-GHRH, was obtained by site-directed mutagenesis (Altered Sites II in vitro Mutagenesis System, Promega, Madison, WI), and cloned into the BamHI/HindIII sites of pSK-GHRH. The GHRH cDNA is followed by the 3′ untranslated region of GH. Characterization of the vector and the long half-life analog HV-GHRH has been previously described (Draghia-Akli et al., 1999). Animals Twenty-eight dogs (Beagles; Harlan Sprague-Dawley, Inc., Houston, TX), approximately 2 yr of age, weighing 10 to 18 kg for males and 7 to 13 kg for females, were divided into one group of four animals (Group I) and three groups of eight animals each (Groups II to IV). Each group had an equal number of males and females. Normal growth, appearance, and behavior were factors used to select healthy animals for testing. Each animal received a pretest physical examination by a laboratory animal veterinarian. Animals were identified by tattoos and cage cards. Necropsies were performed on half the animals from each group on d 93 (two control animals—one male and one female, and four treated animals from each treatment group—two males and two females per group). At approximately d 180, necropsies were performed on the other half of the animals from each group (two control animals—one male and one female, and four treated animals from each treatment group—two males and two females per group). Intramuscular Injection of Plasmid DNA The endotoxin-free plasmid (Qiagen Inc., Chatsworth, CA) preparation of pSPc5-12-HV-GHRH was diluted in water to 5 mg/mL. Animals in Groups II to IV were treated per FDA regulations in target animal safety studies with 1× (a total of 0.2 mg of plasmid), 3× (a total of 0.6 mg of plasmid), and 5× (a total of 1 mg of plasmid) the projected therapeutic dosage range of HV-GHRH plasmid by intramuscular injection followed by electroporation on d 0. Group I animals underwent the electroporation procedure but were not injected with the test article and served as untreated controls. The dogs were anesthetized with 2 mL of a 50:50 mixture of xylazine (Phoenix Scientific, Inc., St. Joseph, MO) and ketamine (Abbott Laboratories, Inc., North Chicago, IL) delivered intramuscularly. All animals received a tattoo of the projected injection area in order to properly identify and isolate the site. The plasmid was injected directly into the semitendinosus muscle with a 1-cc insulin syringe and a 21-gauge, 1.5-in needle (Becton-Dickinson, Franklin Lakes, NJ). Two minutes after injection, the injected muscle was electroporated (three pulses in one orientation, three pulses in an orientation perpendicular to the first one, 100 V/cm, 60 ms/pulse) with a two-needle electrode device, and BTX- 830 electroporator (Genetronics, San Diego, CA). Animals were observed during recovery from anesthesia and then returned to their cages. Weight, and Serum and Urine Endpoints Animal Weight. Dogs were weighed and numbered before the injection/electroporation procedure. Two preinjection blood draws (d −6 and 0), and six post-treatment draws (d 28, 56, 93, 120, 157, and 180) were performed. Average complete blood counts (CBC), biochemistry, and hormone values for d −6 and 0 served as baseline reference for each dog. Whole blood was collected in Monoject Lavender Stopper blood collection tubes with 3.0 mg of EDTA (Sherwood Medical, St. Louis, MO) and submitted for CBC analysis (Antech Diagnostics, Irvine, CA). Serum was aliquoted for radioimmunoassay and biochemical analysis (Antech Diagnostics). Serum was stored at −80°C before analysis. Biochemical analysis occurred within 48 h after serum collection. The IGF-I assay was performed within 90 d after serum collection. Insulin-like Growth Factor-I Radioimmunoassay. Dog IGF-I was measured by a heterologous human assay (Diagnostic System Lab., Webster, Texas). The sensitivity limit of the assay was 0.8 ng/mL; the intra- and interassay variations were 3.4 and 4.5%, respectively. Biochemistry, CBC, Urinalysis, and Hormone Levels. Biochemical and blood chemistry, insulin, and ACTH levels were assayed at Antech Diagnostic at the previously stated time points. During anesthesia prior to injection and on d 93 and 180 at necropsy, a urine sample was drawn from each animal and a urine analysis was performed. A laboratory animal veterinarian reviewed all values. The following parameters were evaluated: serum chemistry, hematology, and urine analysis. Serum chemistry tested for amylase, globulin, γ-glutamyltransferase, total bilirubin, blood urea nitrogen, creatinine, total protein, albumin, direct bilirubin, serum alanine aminotransferase, serum aspartate aminotransferase, alkaline phosphatase, glucose, inorganic phosphorus, chloride, calcium, sodium, potassium, triglyceride, and cholesterol. Hematology looked at erythrocyte counts, hematocrit, hemoglobin, total leukocyte count, and differential leukocyte counts (neutrophils, lymphocytes, monocytes, eosinophils, and basophils), platelet count, mean corpuscular volume, mean corpuscular hemoglobin, mean corpuscular hemoglobin concentration, and partial prothrombin time. Urine analysis analyzed color, consistency, volume, glucose, bilirubin, ketone (acetoacetic acid), specific gravity, blood, pH, protein, urobilinogen, nitrite, leukocytes, and microscopic examination of formed elements. Necropsy Gross Necropsy. All the dogs were killed with an overdose of sodium pentobarbital (Fatal Plus, Vortech Phar- 2303 Effects of plasmid GHRH in dogs maceuticals, Dearborn, MI). The gross necropsy included the following: gross observations of external surfaces, all orifices, and cranial, thoracic, abdominal, and pelvic cavities as well as accompanying organs in situ. Histopathology Analysis of Tissues. Samples of selected organs (heart, lung, liver, spleen, stomach, intestine, pancreas, gall bladder, kidney, gonads, brain, pituitary, adrenals, thyroid, parathyroid, retina and optic nerve, injected muscle, and the injection/electroporation site (skin and muscle) were examined and then assessed histologically by a licensed, independent veterinary pathologist. Polymerase Chain Reaction Analysis for Presence of Plasmid The DNA extraction from injected muscle, liver, and first lymph node was performed according to standard procedures (Sambrook et al., 1989). The PCR was performed for presence of β-actin as a control (forward primer 5′-ATGTACGTGGCCATCCA-3′ and reverse primer 5′-AGGAGGAGGGACCTCTT-3′) and origin of replication (ori) for presence of the injected HV-GHRH plasmid (forward primer 5′-CGAGCGGTATCAGCTCA3′ and reverse primer 5′-ATCGTCTCGCTCCATACAT3′). Primers for PCR amplification were synthesized by Sigma Genosys (The Woodlands, TX). The sensitivity of the conventional PCR reaction is routinely one copy of plasmid per microgram of genomic DNA (Ledwith et al., 2000). Amplification using ori primers was conducted in a GeneAmp 9700 Thermocycler (Pekin Elmer, Norwalk, CT). Polymerase chain reaction was performed in 50-L reactions with 2× PCR mix #1 (Sigma), 0.2 M forward and reverse primers, and 40 ng of dog tissue DNA or 1 ng of control plasmid DNA. Water was substituted for template DNA in negative controls. Polymerase chain reaction conditions were as follows: 1) 94°C for 4 min; 2) five cycles of 94°C for 30 s, 57°C for 30 s, 72°C for 30 s; 3) 30 cycles of 94°C for 30 s, 53°C for 30 s, 72°C for 30 s; and 4) 72°C for 5 min. Products were detected by agarose electrophoresis of an aliquot (60%) of each reaction vs. a 100-bp ladder marker (Invitrogen, Carlsbad, CA). Statistical Analysis Data consisted of repeated measures in seven different time points with an unbalanced study design (Group I, n = 4; Groups II to IV, n = 8). After d 93, the sample size was reduced to half (Group I, n = 2; Groups II to IV, n = 4). Considering all these facts, we chose the MIXED model of SAS (SAS Inst., Inc., Cary, NC; analysis of simple main effects) to examine if there were any significant differences among the groups of each variable at different time points. Multiple comparisons were adjusted by the Tukey-Krammer method. Mean values were compared with Student’s t-test, ANOVA, or linear regression, with <0.05 taken as the level of statistical significance. Results Electroporation and Injection Site Erythema was the most common observation at the injection/electroporation site on d 1 and resolved in a few days. The in-life observations showed one male dog in Group II that developed an abscess on or near the injection site 3 d after injection. Healing occurred around d 19. All other dogs were normal and presented no abnormal findings throughout the study that might be test-article or procedure related. Weights During the study, weights increased in plasmidtreated dogs (Group II by 11.3%, Group III by 6.7%, and Group IV by 14.1%), and slightly decreased in control animals (Group I decreased by 0.8%; Table 1). When data were compared by analysis of covariance using the baseline weight as the regression variable in the analysis, Groups II and IV had a significant weight increase during the study (r2 = 0.59, P < 0.02 and r2 = 0.65, P < 0.001, respectively), whereas Group III had a weight increase that did not reach statistical significance due to high interanimal variability (r2 = 0.20, P < 0.20) (Figure 1). Insulin-Like Growth Factor-I Levels An increase in serum IGF-I concentration over the baseline value was taken as a measure of plasmid GHRH activity (de Boer et al., 1996; Aimaretti et al., 1998). During the study, IGF-I levels were increased in plasmid-treated dogs, and values were statistically significant for days postinjection (sum of average IGFI levels from d 0 to d 180: Group I = 121.9, Group II = 142.6, Group III = 153.4, and Group IV = 153.2, P < 0.001). A dose-related trend in IGF-I levels was observed among the treated groups, but the effect of different treatment doses was secondary (P < 0.09) once days postinjection had been considered. In this case, the variability between groups in the mean values of the various assessed parameters was not greater than the variability within each group. The data from all treated groups of animals, therefore, were combined for comparison with Group I (P < 0.001). When each dog was compared to its own baseline (IGF-I postinjection − IGF-I preinjection/IGF-I preinjection), the group treated with 1 mg of plasmid had a significant increase at all time points analyzed (Figure 2 and Table 2). Insulin-like growth factor-I levels were within normal range for all dogs. Analysis of Blood, Chemical Parameters, and Urinalysis Several variables were significantly different between controls and treated groups at different time points after treatment. Hemoglobin and hematocrit were significantly increased in all treated dogs, with 2304 Draghia-Akli et al. Table 1. Weights in dogs treated with plasmid-mediated GHRH therapya Day Baseline Day 28 Day 42 Day 56 Day 93 Day 120 Day 147 Day 180 Group I Control Group II 0.2-mg dose Group III 0.6-mg dose Group IV 1-mg dose 11.59 ± 0.70b 11.95 ± 0.47 P = 0.06 12.23 ± 0.72 P = 0.17 12.05 ± 0.66 P = 0.16 11.58 ± 0.63 P = 0.04 11.05 ± 0.11 P = 0.33 11.35 ± 0.11 P = 0.0001 11.50 ± 0.07 P = 0.33 11.36 ± 0.97 11.80 ± 1.02 P = 0.05 12.14 ± 1.19 P = 0.03 12.13 ± 1.24 P = 0.08 11.93 ± 1.15 P = 0.09 11.75 ± 1.34 P = 0.006 12.35 ± 1.56 P = 0.02 12.66 ± 1.37 P = 0.0004 11.43 ± 0.86 12.21 ± 0.79 P = 0.007 12.30 ± 0.85 P = 0.01 12.46 ± 0.82 P = 0.002 12.04 ± 0.75 P = 0.08 12.00 ± 0.63 P = 0.27 12.13 ± 0.78 P = 0.21 12.20 ± 0.69 P = 0.18 11.46 ± 0.72 12.28 ± 0.76 P = 0.0002 12.64 ± 0.81 P = 0.0003 12.53 ± 0.83 P = 0.0008 12.09 ± 0.77 P = 0.001 12.73 ± 0.63 P = 0.008 13.10 ± 0.72 P = 0.01 13.08 ± 0.79 P = 0.006 a The results are presented as means ± SEM. P-values vs. baseline weights are included for each group and time point. b Weights were recorded at least eight different times throughout the study for each individual animal on a precision balance. Group 1, n = 4; Groups II through IV, n = 8. least squares means <0.05 (Figure 3), but within normal values for dogs. Calcium:phosphorous ratio, an indication of bone remodeling (Anderson, 1996; Yilmaz et al., 1999), was significantly increased in dogs in Group IV vs. controls at d 180 postinjection (2.75 ± 0.2 vs. 2.28 ± 0.04, P < 0.045). All other values were within the normal reference laboratory range and not indicative of any abnormality. No toxicological significance can be attributed to any of these changes. Fasting glucose and insulin levels were normal for all groups throughout the study. The ACTH levels were assayed on d 120 postinjection. There were no significant differences between treated and control groups. Polymerase Chain Reaction Analysis for Presence of Plasmid Polymerase chain reactions were performed on tissues harvested at the interim necropsy on d 93. The entire injected muscle, first lymph node, and liver of all dogs, treated or control, were assayed. One dog in Group III and one dog in Group IV had a strongly positive PCR reaction on the injected muscle. One dog in Group III had a positive PCR on the injected muscle and liver. One control dog had a false positive reaction in the first lymph node, probably due to a contamination of the tissue harvest that was not reconfirmed when the assay was repeated. All other PCR reactions with dog tissues were negative. Both the positive and negative controls consistently yielded the respective predicted reactions. Histopathological Evaluation of Tissues Figure 1. Weights in dogs treated with plasmid-mediated GHRH therapy. The linear regression curves are presented. For Group I (control), n = 4; for Groups II (0.2 mg of plasmid GHRH), III (0.6 mg of plasmid GHRH), and IV (1 mg of plasmid GHRH), n = 8 per group to d 93. For Group I, n = 2, and for Groups II to IV, n = 4 per group from d 93 to the end of the study. Group II, *P < 0.02, Group IV, **P < 0.001. Samples of the selected organs listed in the Materials and Methods section were examined and then histologically assessed by a board-certified veterinary pathologist. Not all samples could be collected from all dogs due to time constraints during the necropsy process. For some organ/tissue samples, extensive dissection procedures were necessary, and the quality/quantity of tissue obtained was inadequate for complete histological evaluation. As presented in Table 3, there were no test article-related lesions identified by histopathologic examination. Occasional observations, such as pitu- 2305 Effects of plasmid GHRH in dogs Figure 2. Serum IGF-I levels in dogs treated with plasmid-mediated GHRH therapy. The linear regression curves are presented. For Group I (control), n = 4; for Groups II (0.2 mg of plasmid GHRH), III (0.6 mg of plasmid GHRH), and IV (1 mg of plasmid GHRH), n = 8 per group to d 93. For Group I, n = 2, and for Groups II to IV, n = 4 per group from d 93 to the end of the study. Two-way ANOVA, *P < 0.001 for days after treatment for treated vs. control animals. itary cysts, are often seen in young dogs, and the frequency was not different among groups. Discussion Preclinical studies have suggested that anabolic hormones, such as GH, IGF-I, and IGFBP-3, may reverse the catabolic state associated with cachexia, and thus result in increases in weight, lean body mass, and work output in cancer patients (Colao et al., 1999; Kotler, 2000) or patients with kidney failure (Ramirez et al., 1990; Pasqualini et al., 1996). For instance, malnutrition and wasting are important determinants of morbidity and mortality in companion animals with chronic renal failure on dialysis. Even pets with a relatively modest degree of chronic renal insufficiency are characterized by reduced lean body mass, bone mineral content, and basal energy expenditure (O’Sullivan et al., 2002). Growth hormone synthesis and secretion from the anterior pituitary are stimulated by GHRH, a hypothalamic peptide hormone (Muller et al., 1999). Low levels of GHRH (about 100 pg/mL) are sufficient to induce GH secretion (Esch et al., 1982; Corpas et al., 1993). Hence, an alternate method to increase GH production and release would be to administer GHRH. Unfortunately, the short half-life of the GHRH peptide in plasma (<12 min) mandates frequent i.v. or s.c. injections of the hormone to sustain its activity (Evans et al., 1985; Thorner et al., 1986b), making recombinant GHRH therapy impractical for chronic conditions. Both GH and GHRH are currently administered therapeutically as recombinant proteins. Current knowledge about the interaction between GH and its receptor suggests that the molecular heterogeneity of circulating GH may have important implications in growth and homeostasis. Although administration of exogenous rGH produces anabolic effects in a variety of situations (Lieberman et al., 1994; Molon-Noblot et al., 1998; Pichard et al., 1999), rGH therapy has disadvantages. Growth hormone must be administered s.c. or i.m. as frequently as once a day over the entire treatment period, and the nonphysiological hormonal peaks and Table 2. Serum IGF-I concentrations in dogs treated with plasmid-mediated GHRH therapya Day Baseline Day 28 Day 56 Day 93 Day 120 Day 157 Day 180 Median Mean Group I Control Group II 0.2-mg dose Group III 0.6-mg dose Group IV 1-mg dose 13.51 ± 4.81b 16.30 ± 4.03 P = 0.35 14.86 ± 3.47 P = 0.49 14.20 ± 1.50 P = 0.38 17.33 ± 4.02 P = 0.44 22.31 ± 8.82 P = 0.08 23.41 ± 3.06 P = 0.24 16.3 17.42 ± 1.49 14.81 ± 2.28 17.21 ± 3.26 P = 0.26 21.83 ± 4.83 P = 0.04 17.18 ± 5.40 P = 0.33 26.38 ± 4.51 P = 0.02 22.11 ± 4.28 P = 0.06 23.03 ± 4.72 P = 0.07 21.83 20.37 ± 1.54 16.55 ± 4.58 26.22 ± 6.56 P = 0.08 18.77 ± 5.09 P = 0.11 19.34 ± 6.05 P = 0.33 25.03 ± 4.84 P = 0.04 26.09 ± 8.57 P = 0.12 21.38 ± 3.76 P = 0.06 21.38 21.91 ± 1.47 12.31 ± 3.04 17.98 ± 2.90 P = 0.0004 14.34 ± 1.30 P = 0.01 19.37 ± 3.80 P = 0.006 34.47 ± 4.52 P = 0.01 29.91 ± 6.58 P = 0.06 24.77 ± 6.50 P = 0.04 19.37 21.88 ± 3.1 a The results are presented as means ± SEM. P-values vs. baseline weights are included for each group and time point. b The IGF-I concentrations were measured at least eight different times throughout the study for each individual animal; each sample was assayed in duplicate; assays were twice repeated. Group 1, n = 4; for Groups II through IV, n = 8. 2306 Draghia-Akli et al. Figure 3. Hemoglobin and PCV in dogs treated with plasmid-mediated GHRH therapy. The results are presented as means ± SEM. A) Least squares means analysis of hemoglobin for the dogs treated with 0.2 mg, Group II, **P < 0.002; dogs treated with 0.6 mg, Group III, ***P < 0.0001; and dogs treated with 1 mg, Group IV, vs. Group I, controls, *P < 0.05. B) Least squares means analysis of PCV for the dogs treated in Group II, **P < 0.01, dogs treated in Group III, ***P < 0.0001, and dogs treated in Group IV, vs. Group I, *P < 0.025. For Group I (control), n = 4; for Groups II (0.2 mg of plasmid GHRH), III (0.6 mg of plasmid GHRH), and IV (1 mg of plasmid GHRH), n = 8 per group to d 93. For Group I, n = 2, and for Groups II to IV, n = 4 per group from d 93 to the end of the study. troughs that follow such injections often result in impaired glucose tolerance and insulin resistance (Angelopoulos et al., 1998). Moreover, it is clear that biological responses to exogenous rGH are not similar to physiological responses to the naturally occurring isoforms of this protein through rapid negative feedback regulation on pituitary release (Satozawa et al., 2000; Leung et al., 2002). Numerous studies in humans, sheep, and pigs showed that continuous infusion with recombinant GHRH protein restores the normal GH pattern without desensitizing GHRH receptors or depleting GH supplies since this system is capable of feed-back regulation, which is abolished in the GH therapies (Vance et al., 1985; Dubreuil et al., 1990; Vance, 1990), thus avoiding adverse effects (Thorner et al., 1986a; Abribat et al., 1989). Preliminary studies in small and large animal models suggested that a single injection of a GHRH plasmid into skeletal muscle will ensure physiologic GHRH expression for many months (Draghia-Akli et al., 1999; Draghia-Akli et al., 2002b). In a phase I trial in dogs with spontaneous malignancies, we administered a GHRH-expressing plasmid once intramuscularly to severely debilitated, anemic dogs with naturally occurring tumors (Draghia-Akli et al., 2002a). Most (75%) of the animals showed a physiological increase in serum IGF-I concentration, which was associated with the correction of anemia and a significant increase in circulating lymphocytes. There were no discernible adverse effects associated with this therapy. Taken together, these observations demonstrated the feasibility of GHRH-plasmid mediated therapy to stimulate GH synthesis and release in large animals, leading to the correction of anemia and other catabolic processes associated with cachexia. Similar benefits may be achievable in renal failure or geriatric companion animals. Nevertheless, because dogs from the previous study had numerous preexisting conditions and spontaneous malignancies, a clear conclusion about the possible adverse effects of the therapy cannot be made. The present study was designed to address the specific issue of longterm effects of GHRH plasmid-mediated therapy on healthy dogs. This new treatment modality is now practical in vivo because of the significant enhancement of plasmid delivery obtained using electroporation. Electroporation has been used successfully to enhance plasmid uptake by tumor cells after injection (Matsubara et al., 2001; Lucas et al., 2002) and in various organs to deliver therapeutic genes that encode for a variety of hormones, cytokines, or enzymes in mice (Lucas et al., 2001; Vilquin et al., 2001; Lesbordes et al., 2002), rats (Terada et al., 2001; Yasui et al., 2001), and dogs (Fewell et al., 2001) without major adverse effects linked to the procedure itself. Intramuscular injection of plasmid followed by electroporation has been used successfully in ruminants for vaccination purposes (Babiuk et al., 2003; Tollefsen et al., 2003). The dogs that received the plasmid injection followed by electroporation had minimal and transitory erythema at the injection point, but no permanent damage to skin or muscle. By PCR analysis, we were unable to demonstrate the presence of plasmid in the muscles of all treated animals, even when its biological activity was obvious. The entire injected muscle was collected and processed, and the plasmid dose was relatively low. The limit of detection of the assay is such that we could detect the presence of approximately 5,000 molecules or 0.2 pg of plasmid per gram of tissue (Manam et al., 2000). It is thus possible we were not able to detect it under the assay parameters. Nevertheless, previous biodistribution studies using PCR showed similar results. In mini pigs, the plasmid was detected in the treatment sites and also in the inguinal lymph nodes on d 2. At d 57, plasmid was present in the treatment sites only, and by d 141, it seemed to have cleared, even if the transgene product was still present (Haworth and Pilling, 2000; Pilling et al., 2002). Effects of plasmid GHRH in dogs 2307 Table 3. The combined results (from both d 93 and 180), from males (M) and females (F) from the microscopic examination of all available samples Brain Peripheral nerve Hypothalamus Pituitary Cyst Retina Optic nerve Adrenal cortex Adrenal medulla Kidney Pelvis-lymphocytic infiltrate Lymphocytic infiltrate Chronic inflammation Mineralization Testis Prostate Chronic inflammation Uterus/cervix Ovaries Lung Chronic inflammation Stomach Small intestine Cystic mucosal gland Large intestine Spleen Pancreas Liver Central cytoplasmid vacuolization Acute inflammation Gall bladder Bone Bone marrow Heart Skin Chronic inflammation First lymph node Pigmented Hemorrhage Skeletal muscle Injection site The weight gain associated with increased production of GHRH is in agreement with other studies that utilized recombinant porcine GH in dogs. In one of these studies, recombinant porcine GH was administered for 14 wk in dogs (Prahalada et al., 1998). Porcine GH caused increased BW gain in mid- and high-dose groups (2.8 and 4.7 kg, respectively) compared with 0.4 and 0.8 kg in the control and low-dose groups, respectively. During our 6-mo study, GHRH-treated animal weights increased by up to 1.6 kg (controls had a slight weight loss). At necropsy, the body composition was similar among groups. Insulin-like growth factor-I levels were significantly increased in treated dogs, but the increase was not dose dependent, and levels were identical between the groups that received 0.6 and 1 mg of plasmid. As the system maintains feedback and regulation, this is not an unexpected finding. In a study on male Beagles that received GHRH[1-29]NH2 (25 g/kg, subcutaneously, twice daily) for 57 d, it was shown that GHRH injections Normal (M/F) Normal (M/F) Normal (M/F) 1 M in Groups I, II, III and IV 1 F in Group I, 2 F in Groups III and IV Normal (M/F) Normal (M/F) Normal (M/F) Normal (M/F) 2 M in Group III, 1 F in Group II 1 F in Group III 1 F in Group II 1 F in Group IV Normal (M) 1 M in Groups I, II, and IV Normal (F) Normal (F) 1 F in Group III Normal (M/F) 1 M in Group II Normal (M/F) Normal (M/F) Normal (M/F) 1M in Group II and IV, 1 F in Group III 1 F in Group III Normal (M/F) Normal (M/F) Normal (M/F) Normal (M/F) 1F in Group III 3 M in Group III, 1 M in Group IV, 1 F in Group II 1 F in Group IV Normal (M/F) Normal (M/F) produced a significant increase in GH release following each injection, and an increase in GH response over time. The concentration of IGF-I increased for 5 wk and then reached a plateau (Dubreuil et al., 1996) and was constant for the remainder of the study. The short stature of some dog breeds is associated with low serum levels of IGF-I (Guler et al., 1989). When these animals are given recombinant IGF-I, serum levels of IGF-I increase to a certain value and stay constant thereafter. Also, radial length and BW are not increased to a greater extent in the IGF-I-infused dogs than in controls. These facts are indicators of a tight regulation in the GHRH axis in dogs. In our current study, glucose and insulin levels were monitored and found to be normal in all circumstances. Dogs were healthy and playful and did not show signs of lameness. It has been suggested that adverse effects, including insulin resistance, may result from the fact that the basal GH levels are raised and the natural GH episodic pulses are abolished with exogenous GH. 2308 Draghia-Akli et al. Numerous studies have shown that continuous infusion with GHRH restores normal GH pulsatile pattern, with no desensitization of GHRH receptors or depletion of GH supplies. At the same time, this system is capable of feedback. Virtually no side effects have been reported for GHRH therapies (Thorner et al., 1986b). Thus, GHRH therapy may be more physiological than GH therapy. An entire class of GH-releasing peptides is also used in clinics to stimulate GH and IGF-I in humans and dogs. Hexarelin, a potent and well-studied GH-releasing peptide, is capable of causing profound GH release in normal individuals. The GH response to hexarelin in humans becomes appreciably attenuated following longterm administration (attenuation that is partial and reversible) and could seriously limit the potential longterm therapeutic use of hexarelin and similar agents (Rahim and Shalet, 1998). Growth hormonereleasing peptides similar to corticotropin-releasing hormone also possess acute ACTH- and cortisol-releasing activity, although the mechanisms underlying the stimulatory effect of GH-releasing peptides on the hypothalamo-pituitary-adrenal axis are still unclear (Rigamonti et al., 2002). More recent studies in humans have demonstrated acute increases in ACTH (Ghigo et al., 1999), cortisol, and prolactin (Svensson and Bengtsson, 1999) after GH-releasing peptide administration (hexarelin, MK-0677) (Jacks et al., 1996; Schleim et al., 1999). The potential adverse effects of repeated episodes of transient (even minor) hyperprolactinemia and hypercortisolemia during long-term therapy with GHreleasing peptides and similar agents have raised concern and require further study (Rahim et al., 1999). In our study, ACTH levels were normal throughout and not significantly different among groups. The beneficial effect on hemoglobin level and red cell number was not anticipated, because GH administration to normal Beagle dogs over 14 wk of age produced a dose-related normochromic, normocytic, and nonregenerative anemia (Prahalada et al., 1998). Growth hormone-releasing hormone stimulates IGF-I, and there are data to suggest that IGF-I, rather than erythropoietin, is the primary mediator of erythropoiesis during catabolic states and in uremic patients (Urena et al., 1992) and can induce a proportional increase in body mass and oxygen transport capacity (Kurtz et al., 1990). The increase in hemoglobin and hematocrit confirms the in vivo erythropoietic growth-promoting effects of GH that are also observed during GH treatment in GH-deficient children or adults (Christ et al., 1997; Valerio et al., 1997). No dogs showed evidence of polycythemia following plasmid administration, indicating that this approach to stimulating the GHRH/GH/IGF-I axis respects normal physiological levels of hemoglobin and red blood cell synthesis. Implications The ability of plasmid-mediated growth hormone-releasing hormone supplementation to induce growth hormone production and release through natural physiological mechanisms may be a useful single-dose alternative to multiple growth hormone injections. The absence of adverse effects as shown in this study indicates that this treatment strategy can produce increased levels of growth hormone and associated hormones without adversely affecting homeostasis. This treatment methodology represents a significant advance in the ability of veterinarians to treat different conditions in companion or production animals. Literature Cited Abribat, T., A. Regnier, and M. Morre. 1989. Growth hormone response induced by synthetic human growth hormone-releasing factor (1–44) in healthy dogs. Zentbl. Veterinarmed. Reihe A 36:367–373. Aimaretti, G., G. Corneli, P. Razzore, S. Bellone, C. Baffoni, J. Bellone, F. Camanni, and E. Ghigo. 1998. Usefulness of IGF-I assay for the diagnosis of GH deficiency in adults. J. Endocrinol. Invest. 21:506–511. Anderson, J. J. 1996. Calcium, phosphorus and human bone development. J. Nutr. 126:1153S–1158S. Angelopoulos, T. J., R. L. Seip, and T. G. Cole. 1998. Effect of shortterm recombinant growth hormone administration on plasma lipoproteins in elderly adults. Gerontology 44:228–231. Babiuk, L. A., R. Pontarollo, S. Babiuk, B. Loehr, and S. van Drunen Littel-van den Hurk. 2003. Induction of immune responses by DNA vaccines in large animals. Vaccine 21:649–658. Christ, E. R., M. H. Cummings, N. B. Westwood, B. M. Sawyer, T. C. Pearson, P. H. Sonksen, and D. L. Russell-Jones. 1997. The importance of growth hormone in the regulation of erythropoiesis, red cell mass, and plasma volume in adults with growth hormone deficiency. J. Clin. Endocrinol. Metab. 82:2985–2990. Colao, A., A. Cuocolo, C. Di Somma, G. Cerbone, A. M. Della Morte, E. Nicolai, R. Lucci, M. Salvatore, and G. Lombardi. 1999. Impaired cardiac performance in elderly patients with growth hormone deficiency. J. Clin. Endocrinol. Metab. 84:3950–3955. Corpas, E., S. M. Harman, M. A. Pineyro, R. Roberson, and M. R. Blackman. 1993. Continuous subcutaneous infusions of growth hormone (GH) releasing hormone 1–44 for 14 days increase GH and insulin-like growth factor-I levels in old men. J. Clin. Endocrinol. Metab. 76:134–138. de Boer, H., G. J. Blok, C. Popp-Snijders, L. Stuurman, R. C. Baxter, and E. van der Veen. 1996. Monitoring of growth hormone replacement therapy in adults, based on measurement of serum markers. J. Clin. Endocrinol. Metab. 81:1371–1377. Draghia-Akli, R., M. L. Fiorotto, L. A. Hill, P. B. Malone, D. R. Deaver, and R. J. Schwartz. 1999. Myogenic expression of an injectable protease-resistant growth hormone-releasing hormone augments long-term growth in pigs. Nat. Biotechnol. 17:1179–1183. Draghia-Akli, R., K. A. Hahn, G. K. King, K. Cummings, and R. H. Carpenter. 2002a. Effects of plasmid mediated growth hormone releasing hormone in severely debilitated dogs with cancer. Mol. Ther. 6:830–836. Draghia-Akli, R., X. G. Li, and R. J. Schwartz. 1997. Enhanced growth by ectopic expression of growth hormone releasing hormone using an injectable myogenic vector. Nat. Biotechnol. 15:1285– 1289. Draghia-Akli, R., P. B. Malone, L. A. Hill, K. M. Ellis, R. J. Schwartz, and J. L. Nordstrom. 2002b. Enhanced animal growth via ligandregulated GHRH myogenic-injectable vectors. FASEB J. 16:426–428. Dubreuil, P., T. Abribat, B. Broxup, and P. Brazeau. 1996. Longterm growth hormone-releasing factor administration on growth hormone, insulin-like growth factor-I concentrations, and bone healing in the Beagle. Can. J. Vet. Res. 60:7–13. Dubreuil, P., D. Petitclerc, G. Pelletier, P. Gaudreau, C. Farmer, T. F. Mowles, and P. Brazeau. 1990. Effect of dose and frequency Effects of plasmid GHRH in dogs of administration of a potent analog of human growth hormonereleasing factor on hormone secretion and growth in pigs. J. Anim. Sci. 68:1254–1268. Esch, F. S., P. Bohlen, N. C. Ling, P. E. Brazeau, W. B. Wehrenberg, M. O. Thorner, M. J. Cronin, and R. Guillemin. 1982. Characterization of a 40 residue peptide from a human pancreatic tumor with growth hormone releasing activity. Biophys. Biochem. Res. Commun. 109:152–158. Evans, W. S., M. L. Vance, D. L. Kaiser, R. P. Sellers, J. L. Borges, T. R. Downs, L. A. Frohman, J. Rivier, W. Vale, and M. O. Thorner. 1985. Effects of intravenous, subcutaneous, and intranasal administration of growth hormone (GH)-releasing hormone-40 on serum GH concentrations in normal men. J. Clin. Endocrinol. Metab. 61:846–850. Fewell, J. G., F. MacLaughlin, V. Mehta, M. Gondo, F. Nicol, E. Wilson, and L. C. Smith. 2001. Gene therapy for the treatment of hemophilia B using PINC-formulated plasmid delivered to muscle with electroporation. Mol. Ther. 3:574–583. Ghigo, E., E. Arvat, F. Broglio, R. Giordano, L. Gianotti, G. Muccioli, M. Papotti, A. Graziani, G. Bisi, R. Deghenghi, and F. Camanni. 1999. Endocrine and non-endocrine activities of growth hormone secretagogues in humans. Horm. Res. 51 Suppl 3:9–15. Guler, H. P., K. Binz, E. Eigenmann, S. Jaggi, D. Zimmermann, J. Zapf, and E. R. Froesch. 1989. Small stature and insulin-like growth factors: Prolonged treatment of mini-poodles with recombinant human insulin-like growth factor I. Acta Endocrinol. (Copenh.) 121:456–464. Haworth, R., and A. M. Pilling. 2000. The PCR assay in the preclinical safety evaluation of nucleic acid medicines. Hum. Exp. Toxicol. 19:267–276. Jacks, T., R. Smith, F. Judith, K. Schleim, E. Frazier, H. Chen, D. Krupa, D. Hora Jr., R. Nargund, A. Patchett, and G. Hickey. 1996. MK-0677, a potent, novel, orally active growth hormone (GH) secretagogue: GH, insulin-like growth factor I, and other hormonal responses in beagles. Endocrinology 137:5284–5289. Kotler, D. P. 2000. Cachexia. Ann. Intern. Med. 133:622–634. Kurtz, A., R. Matter, K. U. Eckardt, and J. Zapf. 1990. Erythropoiesis, serum erythropoietin, and serum IGF-I in rats during accelerated growth. Acta Endocrinol. (Copenh.) 122:323–328. Ledwith, B. J., S. Manam, P. J. Troilo, A. B. Barnum, C. J. Pauley, T. G. Griffiths, L. B. Harper, H. B. Schock, H. Zhang, J. E. Faris, P. A. Way, C. M. Beare, W. J. Bagdon, and W. W. Nichols. 2000. Plasmid DNA vaccines: Assay for integration into host genomic DNA. Dev. Biol. (Basel) 104:33–43.. Lesbordes, J. C., T. Bordet, G. Haase, L. Castelnau-Ptakhine, S. Rouhani, H. Gilgenkrantz, and A. Kahn. 2002. In vivo electrotransfer of the cardiotrophin-1 gene into skeletal muscle slows down progression of motor neuron degeneration in pmn mice. Hum. Mol. Genet. 11:1615–1625. Leung, K. C., C. Howe, L. Y. Gui, G. Trout, J. D. Veldhuis, and K. K. Ho. 2002. Physiological and pharmacological regulation of 20-kDa growth hormone. Am. J. Physiol. Endocrinol. Metab. 283:E836–E843. Li, X., E. M. Eastman, R. J. Schwartz, and R. Draghia-Akli. 1999. Synthetic muscle promoters: activities exceeding naturally occurring regulatory sequences. Nat. Biotechnol. 17:241–245. Lieberman, S. A., G. E. Butterfield, D. Harrison, and A. R. Hoffman. 1994. Anabolic effects of recombinant insulin-like growth factorI in cachectic patients with the acquired immunodeficiency syndrome. J Clin. Endocrinol. Metab. 78:404–410. Lucas, M. L., L. Heller, D. Coppola, and R. Heller. 2002. IL-12 plasmid delivery by in vivo electroporation for the successful treatment of established subcutaneous B16.F10 melanoma. Mol. Ther. 5:668–675. Lucas, M. L., M. J. Jaroszeski, R. Gilbert, and R. Heller. 2001. In vivo electroporation using an exponentially enhanced pulse: a new waveform. DNA Cell Biol. 20:183–188. Manam, S., B. J. Ledwith, A. B. Barnum, P. J. Troilo, C. J. Pauley, L. B. Harper, T. G. Griffiths, Z. Niu, L. Denisova, T. T. Follmer, S. J. Pacchione, Z. Wang, C. M. Beare, W. J. Bagdon, and W. W. Nichols. 2000. Plasmid DNA vaccines: Tissue distribution 2309 and effects of DNA sequence, adjuvants and delivery method on integration into host DNA. Intervirology 43:273–281. Matsubara, H., Y. Gunji, T. Maeda, K. Tasaki, Y. Koide, T. Asano, T. Ochiai, S. Sakiyama, and M. Tagawa. 2001. Electroporationmediated transfer of cytokine genes into human esophageal tumors produces anti-tumor effects in mice. Anticancer Res. 21:2501–2503. Molon-Noblot, S., P. Laroque, S. Prahalada, L. G. Stabinski, C. M. Hoe, C. P. Peter, P. Duprat, and M. J. van Zwieten. 1998. Effect of chronic growth hormone administration on skeletal muscle in dogs. Toxicol. Pathol. 26:207–212. Muller, E. E., V. Locatelli, and D. Cocchi. 1999. Neuroendocrine control of growth hormone secretion. Physiol Rev. 79:511–607. Murray, R. D., and S. M. Shalet. 2000. Growth hormone: current and future therapeutic applications. Expert. Opin. Pharmacother. 1:975–990. O’Sullivan, A. J., J. A. Lawson, M. Chan, and J. J. Kelly. 2002. Body composition and energy metabolism in chronic renal insufficiency. Am. J Kidney Dis. 39:369–375. Pasqualini, T., J. Ferraris, P. Fainstein-Day, A. A. Eymann, C. S. Moyano, S. Ruiz, J. Ramirez, and R. Gutman. 1996. Growth acceleration in children with chronic renal failure treated with growth-hormone-releasing hormone (GHRH). Medicina (B. Aires) 56:241–246. Pichard, C., U. G. Kyle, P. Jolliet, D. O. Slosman, T. Rochat, L. Nicod, J. Romand, N. Mensi, and J. C. Chevrolet. 1999. Treatment of cachexia with recombinant growth hormone in a patient before lung transplantation: a case report. Crit. Care Med. 27:1639– 1642. Pilling, A. M., R. M. Harman, S. A. Jones, N. A. McCormack, D. Lavender, and R. Haworth. 2002. The assessment of local tolerance, acute toxicity, and DNA biodistribution following particlemediated delivery of a DNA vaccine to minipigs. Toxicol. Pathol. 30:298–305. Prahalada, S., L. G. Stabinski, H. Y. Chen, R. E. Morrissey, G. De Burlet, D. Holder, D. H. Patrick, C. P. Peter, and M. J. van Zwieten. 1998. Pharmacological and toxicological effects of chronic porcine growth hormone administration in dogs. Toxicol. Pathol. 26:185–200. Rahim, A., P. A. O’Neill, and S. M. Shalet. 1999. The effect of chronic hexarelin administration on the pituitary-adrenal axis and prolactin. Clin. Endocrinol. (Oxford) 50:77–84. Rahim, A. and S. M. Shalet. 1998. Does desensitization to hexarelin occur? Growth Horm. IGF Res. 8(Suppl. B):141–143. Ramirez, G., B. B. Bercu, P. A. Bittle, C. W. Ayers, and A. Ganguly. 1990. Response to growth hormone-releasing hormone in adult renal failure patients on hemodialysis. Metabolism 39:764–768. Rigamonti, A. E., S. M. Bonomo, S. G. Cella, and E. E. Muller. 2002. GH and cortisol rebound rise during and following a somatostatin infusion: Studies in dogs with the use of a GH-releasing peptide. J. Endocrinol. 174:387–394. Sambrook, J., E. F. Fritsch, and T. Maniatis. 1989. Molecular Cloning—A Laboratory Manual. Cold Spring Harbor Laboratory Press, Cold Spring Harbor, NY. Satozawa, N., K. Takezawa, T. Miwa, S. Takahashi, M. Hayakawa, and H. Ooka. 2000. Differences in the effects of 20 K- and 22 K-hGH on water retention in rats. Growth Horm. IGF Res. 10:187–192. Schleim, K. D., T. Jacks, P. Cunningham, W. Feeney, E. G. Frazier, G. W. Niebauer, D. Zhang, H. Chen, R. G. Smith, and G. Hickey. 1999. Increases in circulating insulin-like growth factor I levels by the oral growth hormone secretagogue MK-0677 in the beagle are dependent upon pituitary mediation. Endocrinology 140:1552–1558. Svensson, J. A., and B. Bengtsson. 1999. Clinical and experimental effects of growth hormone secretagogues on various organ systems. Horm. Res. 51(Suppl. 3):16–20. Terada, Y., H. Tanaka, T. Okado, S. Inoshita, M. Kuwahara, T. Akiba, S. Sasaki, and F. Marumo. 2001. Efficient and ligand-dependent regulated erythropoietin production by naked DNA injection and in vivo electroporation. Am. J. Kidney Dis. 38:S50–S53. 2310 Draghia-Akli et al. Thorner, M. O., M. L. Vance, W. S. Evans, R. M. Blizzard, A. D. Rogol, K. Ho, D. A. Leong, J. L. Borges, M. J. Cronin, R. M. MacLeod, et al. 1986a. Physiological and clinical studies of GRF and GH. Rec. Prog. Horm. Res. 42:589–640. Thorner, M. O., M. L. Vance, W. S. Evans, A. D. Rogol, J. Rivier, W. Vale, Blizzard, and RM. 1986b. Clinical studies with GHRH in man. Horm. Res. 24:91–98. Tollefsen, S., M. Vordermeier, I. Olsen, A. K. Storset, L. J. Reitan, D. Clifford, D. B. Lowrie, H. G. Wiker, K. Huygen, G. Hewinson, I. Mathiesen, and T. E. Tjelle. 2003. DNA injection in combination with electroporation: a novel method for vaccination of farmed ruminants. Scand. J. Immunol. 57:229–238. Urena, P., A. Bonnardeaux, K. U. Eckardt, A. Kurtz, and T. B. Drueke. 1992. Insulin-like growth factor I: a modulator of erythropoiesis in uraemic patients? Nephrol. Dial. Transplant 7:40–44. Valerio, G., S. Di Maio, M. Salerno, A. Argenziano, R. Badolato, and A. Tenore. 1997. Assessment of red blood cell indices in growthhormone-treated children. Horm. Res. 47:62–66. Vance, M. L. 1990. Growth-hormone-releasing hormone. Clin. Chem. 36:415–420. Vance, M. L., D. L. Kaiser, W. S. Evans, R. Furlanetto, W. Vale, J. Rivier, and M. O. Thorner. 1985. Pulsatile growth hormone secretion in normal man during a continuous 24-hour infusion of human growth hormone releasing factor (1–40). Evidence for intermittent somatostatin secretion. J. Clin. Invest. 75:1584– 1590. Vilquin, J. T., P. F. Kennel, M. Paturneau-Jouas, P. Chapdelaine, N. Boissel, P. Delaere, J. P. Tremblay, D. Scherman, M. Y. Fiszman, and K. Schwartz. 2001. Electrotransfer of naked DNA in the skeletal muscles of animal models of muscular dystrophies. Gene Ther. 8:1097–1107. Yasui, A., K. Oda, H. Usunomiya, K. Kakudo, T. Suzuki, T. Yoshida, H. M. Park, K. Fukazawa, and T. Muramatsu. 2001. Elevated gastrin secretion by in vivo gene electroporation in skeletal muscle. Int. J. Mol. Med. 8:489–494. Yilmaz, N., M. Bayram, A. B. Erbagci, and M. S. Kilincer. 1999. Diagnostic value of biochemical markers of bone turnover and postmenopausal osteoporosis. Clin. Chem. Lab Med. 37:137–143.