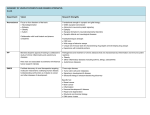
Survey
* Your assessment is very important for improving the workof artificial intelligence, which forms the content of this project
Progress in Neurobiology 80 (2006) 99–113 www.elsevier.com/locate/pneurobio ‘‘The seven sins’’ of the Hebbian synapse: Can the hypothesis of synaptic plasticity explain long-term memory consolidation? Yuri I. Arshavsky * Institute for Nonlinear Science, University of California San Diego, La Jolla, CA 92093-0402, USA Received 28 July 2006; received in revised form 25 September 2006; accepted 26 September 2006 Abstract Memorizing new facts and events means that entering information produces specific physical changes within the brain. According to the commonly accepted view, traces of memory are stored through the structural modifications of synaptic connections, which result in changes of synaptic efficiency and, therefore, in formations of new patterns of neural activity (the hypothesis of synaptic plasticity). Most of the current knowledge on learning and initial stages of memory consolidation (‘‘synaptic consolidation’’) is based on this hypothesis. However, the hypothesis of synaptic plasticity faces a number of conceptual and experimental difficulties when it deals with potentially permanent consolidation of declarative memory (‘‘system consolidation’’). These difficulties are rooted in the major intrinsic self-contradiction of the hypothesis: stable declarative memory is unlikely to be based on such a non-stable foundation as synaptic plasticity. Memory that can last throughout an entire lifespan should be ‘‘etched in stone.’’ The only ‘‘stone-like’’ molecules within living cells are DNA molecules. Therefore, I advocate an alternative, genomic hypothesis of memory, which suggests that acquired information is persistently stored within individual neurons through modifications of DNA, and that these modifications serve as the carriers of elementary memory traces. # 2006 Elsevier Ltd. All rights reserved. Keywords: Declarative memory; Synaptic plasticity; Long-term potentiation; Hebbian hypothesis; Genomic hypothesis; Memory stability; Memory localization; Reconsolidation; Adult neurogenesis; Alzheimer’s disease Contents 1. 2. 3. 4. Introduction . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . Limitations of the synaptic plasticity hypothesis . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 2.1. The synaptic plasticity hypothesis cannot explain the long-life persistence of memory . . . . . . . . . . . . . . . . . . . . . . 2.2. The synaptic plasticity hypothesis’ suggestion that the same mechanism is used for memory storage and reproduction is seriously flawed . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 2.3. Memory acquisition and storage have different localizations . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 2.4. ‘‘Synaptic’’ and ‘‘system’’ memory consolidations have different temporal characteristics. . . . . . . . . . . . . . . . . . . . 2.5. Reconsolidation of memory . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 2.6. Neurogenesis in the adult brain . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 2.7. The synaptic plasticity hypothesis does not explain the specific memory impairments present in Alzheimer’s disease Alternative (genomic) hypothesis of memory . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . Concluding remarks . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . References . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . ... ... ... 100 101 101 . . . . . . . . . 102 103 104 105 106 107 107 109 110 . . . . . . . . . . . . . . . . . . Abbreviations: aCaMKII, a-calcium-calmodulin kinase II; AD, Alzheimer’s disease; CPEB, cytoplasmic polyadenelation element binding protein; CRE, cAMP-response element; CREB, cAMP response element-binding protein; EST, electroshock therapy; LTP, long-term potentiation; MTL, medial temporal lobe; SPH, the synaptic plasticity hypothesis * Tel.: +1 858 822 2010; fax: +1 858 534 7664. E-mail address: [email protected]. 0301-0082/$ – see front matter # 2006 Elsevier Ltd. All rights reserved. doi:10.1016/j.pneurobio.2006.09.004 100 Y.I. Arshavsky / Progress in Neurobiology 80 (2006) 99–113 . . .the past, the memories and realities that are the bedrock of one’s present life, brought back suddenly by a scent, the shape of a hill, an old song – some triviality that makes one suddenly say ‘I remember. . .’ Agatha Christie. An Autobiography. 1. Introduction In this paper, I look at some of the issues related to explaining the neural mechanisms of declarative (explicit) memory, that is, conscious memory for facts, events, language, faces, music, etc. This memory is formed through life experience and, together with the genome, determines the human personality. Declarative memory includes three closely coupled processes: 1. Acquisition of input information (learning); 2. Consolidation of acquired information from initially labile to stable memories and their long-lasting (sometimes life-long) storage in the brain; 3. The recall and use of stored information (memory retrieval). The retrieval of declarative memory in humans is, by definition, the conscious recall of stored information. This is the principal impediment in discussing the memory retrieval mechanism, because understanding neural mechanisms of consciousness is beyond the scope of modern science. Therefore, I mainly concentrate on memory consolidation and storage. However, a total avoidance of memory retrieval is hardly possible because the memory storage can only be tested by its retrieval. In humans, the storage of declarative memory is usually tested through verbal accounts, whereas homologues of human’s declarative memory in animals (see Manns and Eichenbaum, 2006) are tested in behavioral experiments. Memorizing new facts and events means that entering information produces specific physical changes within the brain (the memory engram). According to the most commonly accepted viewpoint (the synaptic plasticity hypothesis, SPH) traces of memory are stored within the brain through structural modifications of synaptic connections, which result in changes of synaptic efficiency and, therefore, in formations of new patterns of neural activity. This hypothesis, suggested by Hebb in 1949, remains the only hypothesis on the cellular basis of memory that guides experimental efforts in the field (see Chklovskii et al., 2004; Kandel, 2000, 2001; Milner et al., 1998; Squire and Kandel, 1999; Sweatt, 2003 for recent reviews). As claimed by Bailey (1999), ‘‘it has seemed almost axiomatic that learning and memory must be expressed as a change in synaptic function and form.’’ The current situation in the field was vividly described by Mel (2002): ‘‘Pull the average neuroscientist off the street and ask them how learning occurs in the brain, and you’re likely to get a reflex response that includes such pat phrases as ‘activitydependent changes in synaptic strength,’ ‘LTP/LTD,’ or ‘Hebbian learning’.’’ Until recently, the central Hebbian postulate which states that ‘‘neurons that fire together wire together’’ has not been questioned by scientists. All current discussions are mainly concentrated around the details of the molecular mechanisms that determine the formation and modification synaptic connections, and maintain the stability of newly formed and modified synapses. The following quotation from a recent review paper demonstrates this widely held perspective: ‘‘Activity-dependent synaptic plasticity is induced at appropriate synapses during memory formation, and is both necessary and sufficient for the information storage underlying the type of memory mediated by the brain area in which that plasticity is observed.’’ (Martin et al., 2000) I think that the major reason why the SPH has become indisputable is rooted in the general paradigm of modern neuroscience. According to this paradigm, neural networks consisting of simple elements (whose own role is negligible) perform not only the ‘‘automatic’’ functions of the nervous system (involuntary reflexes, control of respiratory, locomotor and other rhythmic movements, maintenance of homeostasis, etc.), but also higher cognitive functions of the brain (the critical consideration of this paradigm was presented elsewhere, see Arshavsky, 2001, 2003a,c, 2006b). The paradigm goes on to say that the high complexity of cognitive functions is solely determined by the complexity of networks (see, for example, Koch and Laurent, 1999; Quartz and Sejnowski, 1997), while the function of single neurons is limited to the generation of electrical potentials and the transmission of signals to other neurons. This viewpoint was distinctly spelled out by LeDoux (2003) as follows: ‘‘In many ways, the self is synaptic.’’ It is evident that within this conceptual framework, no other mechanism for memory storage – except for modifications in synaptic connections – is possible. Another reason for the high popularity of the SPH is the discovery of the well-known synaptic phenomenon – long-term potentiation (LTP). A crucial role in forming new memories belongs to the medial temporal lobe (MTL), including the hippocampus (see Section 2.3). It was found that a brief highfrequency train of stimuli applied to intrahippocampal pathways evoked long-lasting (up to several hours) enhancement of synaptic transmission to target neurons (Bliss and Lømo, 1973). Further studies showed that several trains of stimuli evoked LTP lasted for more than 24 h, and that the same stimulation in intact animals could evoke LTP lasting for days, and even weeks (see Kandel, 2000). LTP was also found in other brain structures related to the memory formation – the neocortex, cerebellum, amygdala (Blair et al., 2001; D’Angelo et al., 2005; Linch, 2004; Teskey and Valentine, 1998). Numerous studies have shown that LTP and memory formation share similar cellular, biochemical, and molecular mechanisms (Dudai, 2004; Kandel, 2000; Lamprecht and LeDoux, 2004; Linch, 2004; Malenka and Bear, 2004; Sweatt, 2003; Wang et al., 2006). In experiments on hippocampal slices, it has been found that LTP is accompanied by structural modifications in synaptic connections (Engert and Bonhoeffer, 1999; Maletic-Savatic et al., 1999; Nagerl et al., 2004; Toni et al., 1999). Thus, Y.I. Arshavsky / Progress in Neurobiology 80 (2006) 99–113 although LTP is evoked by artificial electrical stimulation of input pathways with frequencies of up to 100 Hz, which do not occur in the nervous system, this phenomenon is usually regarded as compelling evidence for the SPH. As stated by Abraham and Williams (2003), ‘‘For three decades, long-term potentiation (LTP) has been the gold standard synaptic model for mammalian memory mechanisms.’’ 2. Limitations of the synaptic plasticity hypothesis In this chapter I will illustrate that the SPH, which has played a critical role in understanding the cellular mechanisms of learning and initial stages of memory consolidation, faces at least seven conceptual and experimental difficulties when it seeks to explain a mechanism for long-term (potentially permanent) storage of declarative memory. These difficulties are as follows. 2.1. The synaptic plasticity hypothesis cannot explain the long-life persistence of memory The key feature of declarative memory is its permanence. Psycho-physiological studies provide evidence that human individuals possess the ability to store and retrieve large amounts of information throughout the entire lifetime, or at least during several decades, even if the information is seldom or never used. In a study on elderly individuals who had taken a Spanish course in high school, but had never used Spanish later in their lives, it was found that about 50% of the originally acquired knowledge remained accessible for over 50 years (Bahrick, 1984). A similar result was obtained in a study on memory of names and faces. The participants were able to recognize about 90% of portraits of their high school classmates 35 years after graduation (Bahrick et al., 1975).1 Our inability to recall certain facts and events does not necessarily mean that those traces of information have been deleted from the brain. Facts and events that appear to have been completely forgotten may be revived in the hypnotic state (Cheek and LeCron, 1968; Luria, 1976; Fligstein et al., 1998; Green, 1999) or in the response to electrical stimulation of the lateral temporal lobe during brain surgery (Penfield, 1958, see Box 1). Therefore, the inability to recall some facts and events may signify that these memories, which continue to be stored in the brain, are inaccessible to conscious recollection. Particularly, pertinent evidence for the remarkable brain’s ability to store memory has been shown in studies on mnemonists – individuals who demonstrate an outstanding ability to memorize and retrieve information. A mnemonist named Shereshevskii was able to reproduce useless information, such as long series of words, 15 years after this information was acquired (Luria, 1968). Another mnemonist, Peek, 55 years old, remembers by heart about 9000 books, the first of which he acquired at the age of 18 months, as they were read to him 101 Box 1. Permanency of memory (case report) Very important observations, showing that the inability to remember some facts and events does not signify that the traces of information were lost, were described by the famous Canadian neurosurgeon, Wilder Penfield. Patients had been operated upon for the relief of focal epileptic seizures at the Montreal Neurological Institute. These operations were carried out under local anesthesia and brain stimulations were applied during the operation in an attempt to localize the origin of the patient’s seizure. It was found that stimulation of the temporal lobe of the cortex could cause previous experiences to be recalled by the conscious patients. Electrically evoked recollections of previously experienced perceptions were largely auditory, or visual, or both. This recollection was much more vivid than normal memories, and was often about things that remained unrecalled under ordinary circumstances. The following is an example of one of the cases described by Penfield (1958). A 26-year-old woman, D. F., began to have seizures at age 6. In response to stimulation of the temporal lobe during the operation she said: ‘‘I hear the music. It is like the radio’’. She was asked to describe the music. When the electrode was applied again, she began to hum a tune. She was obviously humming along with orchestra at about the tempo that would be expected. After the patient had returned home, she wrote a letter that was, in part, as follows: ‘‘Now to answer your questions: I heard the song right from the beginning, and you know I could remember much more of it right in the operating room . . . There were instruments . . . It was as though it were being played by an orchestra. Definitely it was not as though I were imagining the tune to myself. I actually heard it. It is not one of my favorite songs, so I don’t know why I heard that song.’’ (Treffert and Christensen, 2005). One more case of outstanding autobiographical memory called hyperthymestic syndrome (from the Greek word thymesis meaning remembering) was recently described by Parker et al. (2006). Awoman, AJ, 41 years old, can describe with considerable accuracy what she was doing any day of her life starting from her childhood. As was highlighted by Treffert and Christensen (2005), we cannot say that we understand the mechanism of memory until we can explain these extraordinary abilities of some individuals for memory storage and retrieval. The question inevitably arises as to whether the SPH can explain the potentially permanent maintenance of declarative memory. In short, the SPH which has been described in many reviews (Kandel, 2001; Lamprecht and LeDoux, 2004; Silva, 2003; Squire and Kandel, 1999; Sweatt, 2003; Thomas and Huganir, 2004) suggests the following mechanism for synaptic modifications in the hippocampus.2 Input signals carrying new 1 Recently, I had a chance to test these observations. I found a photo of my high school class, taken in 1947. Beside me, there are 16 persons in this photo, none of whom I have seen since then. I found that I could remember 15 of them by first and last names. Therefore, this unused information has been stored in my brain for almost 60 years. 2 In addition to studies on the hippocampus, an important role in understanding the molecular mechanisms of synaptic plasticity was played by studies of post-stimulatory sensitization of the withdrawal reflex in the marine mollusk, Aplysia (Kandel, 2000; Squire and Kandel, 1999). 102 Y.I. Arshavsky / Progress in Neurobiology 80 (2006) 99–113 information act on the glutamatergic receptors of the target hippocampal neurons resulting in Ca2+ influx. Ca2+ initiates an intracellular signal transduction cascade: the activation of the Ca2+-calmodulin-dependent adenyl cyclase, the increased synthesis of cAMP, and the activation of several protein kinases. Activated kinases are translocated to the cell nucleus, where they phosphorylate the cAMP response element-binding protein (CREB). CREB binds to the cAMP-response elements (CRE) in regulatory regions of target genes and stimulates their transcription. The products of these genes may be transcription factors themselves that regulate the transcription of the downstream genes. The final step of this cascade is the transient activation of the genes involved in the synthesis of the proteins essential both for the modification of preexisting synapses and for the formation of new ones. Thousands of works using different experimental approaches have found that the mechanism described above is involved in learning and memory formation. Particularly, it has been shown that inhibitors of protein (Davis and Squire, 1984; McGaugh, 2000) and mRNA (Bailey et al., 1999; Igaz et al., 2002) synthesis administered before or shortly after learning precluded the consolidation of long-term memory (see Section 2.4). The latter fact is regarded as compelling evidence that memory formation depends on protein synthesis-dependent modifications of synaptic connections. However, the question remains, whether modifications of synaptic connections are used only at the initial stages of memory formation, or whether they are also used as the mechanism for the persistent memory storage. This is a fundamental question about memory, which must be discussed in detail. While developing the idea that ‘‘the self is synaptic,’’ LeDoux (2003) claimed that ‘‘Genes and experience . . . are . . . not different things, but different ways of doing the same thing – wiring the synapses of our brain.’’ This statement is disputable. Here, I will clarify why I do not think that genes and experience (as it is interpreted by the SPH) are the same things. The basic organization of the brain is determined by genetic mechanisms. Although the way from the genotype to the organization of the brain consisting of 1011 neurons connected through 1013 to 1015 synapses is very complex and enigmatic, it is evident that genetic programs determine not only gross connections between different brain structures, but also the organization of synaptic connections within circuitries (Benson et al., 2001). Otherwise, the existence of cytoarchitectonic atlases of cerebral neurons and their connections, as well as the reproducibility of electrophysiological results obtained in studies of synaptic connections within microcircuits, would be impossible. The stability of synaptic connections in the neocortex was recently confirmed in in vivo studies, using twophoton microscopy and transgenic mice expressing green fluorescent protein (Grutzendler et al., 2002; Zuo et al., 2005; De Paola et al., 2006). Although dendrite spines and axonal branches were continuously changing due to their elongation and retraction, the overall density of synaptic contacts did not change for a fairly long period (up to several months in mice). One can suggest that this stability of synaptic connections is maintained by the same genetic mechanisms which are responsible for the organization of intra- and internetwork connections during brain development. In contrast, the SPH suggests that experience-dependent modifications in synaptic connections are realized at the transcriptional/translational level and do not involve genomic changes. Therefore, it is assumed that structural synaptic modifications result from a transient activation of the protein synthesis (see above). Since proteins have a very short lifetime as compared with memory lifetime, additional hypotheses are required to explain how stable synaptic modifications could result from a transient activation of the protein synthesis. This difficulty of the SPH was recently highlighted by several authors in the following ways: ‘‘Maintaining an increased synaptic connection over many protein half-lives, in the face of complete breakdown and resynthesis of all the protein components that make up that synapse, requires a self-reinforcing signal’’ (Sweatt, 2003) and ‘‘The structural change once initiated is not sufficient as a maintenance mechanism for long-term memory. The structural change itself must be actively maintained’’ (Bailey et al., 2004). Several hypothetical mechanisms for the stabilization of learning-related synaptic modifications have been suggested. These hypotheses assume that the stabilization of experiencerelated synaptic modifications is maintained by local selfperpetuating translational and posttranslational mechanisms (Bailey et al., 2004; Routtenberg and Rekart, 2005; Yi and Ehlers, 2005). The most elaborate hypothesis was advanced by Kandel and coworkers (Bailey et al., 2004; Si et al., 2003a,b). They suggested that a critical role in stabilizing local synaptic changes belongs to the neuron-specific isoform of cytoplasmic polyadenelation element binding protein (CPEB). A synaptic input increases the amount of the CPEB protein in a restricted postsynaptic area and switches CPEB into a self-perpetuating prion-like form that has a great capacity to facilitate mRNA translation. Thus, conformation of CPEB into the selfperpetuating prion-like state determines both prolonged and local translation of mRNAs in the activated synapses. Essentially, all these hypotheses deal with phenomena (such as LTP in the hippocampus, or long-term sensitization of the gill-withdrawal reflex in Aplysia) that last days or weeks. Therefore, more evidence is needed to establish whether the hypothesized mechanisms (i.e., the mechanisms based on the suggestion that experience-dependent changes in the brain are realized at the transcriptional/translational level and do not involve genomic changes) can be used to form persistent declarative memories lasting years and even whole lifespan. 2.2. The synaptic plasticity hypothesis’ suggestion that the same mechanism is used for memory storage and reproduction is seriously flawed It is commonly accepted that changes in the strength of synaptic connections lead to the formation of new patterns of Y.I. Arshavsky / Progress in Neurobiology 80 (2006) 99–113 neural activity, while a reactivation of these patterns results in memory retrieval. In a recent book on neural mechanisms of memory, Maroun et al. (2001) claimed: ‘‘According to this view [the SPH], coactive and potentially interconnected neurons would, after repeated reactivity, become more strongly connected. These changes in connection strength implemented learning, its persistence comprised memory, and the reactivation of assemblies of such neurons was memory retrieval.’’ Thus, the SPH assumes that the same synaptic mechanisms are used for both memory storage and retrieval. Although this idea may initially seem appealing, it is the ‘‘Achilles heel’’ of the SPH. If we look at any artificial memory device (ranging from notebooks to video tapes and hard drives), we can see that each system requires separate mechanisms (or separate communicational channels) to save and to reproduce information. For example, recording a phone number in a notebook requires the use of our motor system and a pen, while reading the number requires the use of the visual system. Imagine what would happen if we needed to use the motor system and a pen not only to save the phone number, but to ‘‘read’’ it as well. If one had to trace the written number with a pen each time it was ‘‘read’’, the written number would soon become muddled, as our motor system has a lot of sources of noise and we cannot trace the original lines perfectly. The more it was accessed, the faster the written information would erode. Another example is the VCR. To record a movie, a VCR produces physical changes in a magnetic tape; to show a movie, it reads the tape and sends signals in ‘‘the opposite direction,’’ without affecting the stored information. Thus, the storage of information in all artificial systems is organized in such a way that information cannot be lost by being reproduced, and this allows us to use it as many times as we like. Information can solely be lost because of a physical destruction of its carrier. Unlike a VCR, in neural networks signals cannot be sent in the opposite direction through the same synaptic connections because of unilateral transmission in synapses. Consequently, the SPH assumes that the same communicational channels are used both for the memory storage and reproduction. I cannot exclude that a mechanism of this kind could be used in an ideal system lacking any noise. However, as was mentioned, the real nervous system has many sources of noise. Therefore, patterns of synaptic excitation during repeated memory retrieval will always have slight variations. If plasticity is an inherent property of synapses, each memory reactivation must lead to further modification in synaptic connections and, eventually, to memory erosion, in direct correlation to how frequently it is accessed (see the aforementioned example with the notebook). Since this is not the case, and repetitive retrievals lead to memory strengthening rather than erosion, we must conclude that memory is resistant to the synaptic activity that accompanies memory retrieval. This is not the only example that shows the resistance of memory to synaptic activity. Another example is the resistance of memory to spontaneous neuronal activity. According to the 103 SPH, we must expect that ongoing spontaneous activity will lead to incessant, uncontrolled modifications in synaptic connections and, eventually, to erosions in older memories. This glitch in the SPH was distinctly formulated by Eccles (1972) as follows: ‘‘The simple concept that disuse leads to regression of spine synapses and excess usage to hypertrophy can be criticized because . . . almost all cells . . . are discharging continuously. One can imagine therefore that there would be overall hypertrophy of all synapses under such conditions . . . Evidently frequent synaptic excitation could hardly provide a satisfactory explanation of synaptic changes involved in learning.’’ The third example of declarative memory’s resistance to synaptic activity is memory survival of epileptic fits and electroshock therapy (EST). Epileptic fits are accompanied by bursts of synchronous high-frequency discharges of cerebral neurons. In experiments on animals, it was found that the intraburst spike frequencies (80–200 Hz, Grenier et al., 2003) are comparable with the frequency of electrical stimulation used for receiving LTP (see above) and undoubtedly overcome the frequency of sensory signals leading to the formation of declarative memory. Nonetheless, although this non-controlled synaptic activity comprises widespread areas of the cerebral cortex, and is strong enough to produce substantial remodifications of all synaptic connections, it does not affect long-term memory. In most cases, seizure victims only suffer memory loss for the period during and immediately prior to the seizure (Thompson, 1991; see also Footnote 3). The same can be said of EST. Repetitive electroconvulsive treatments, which evoke synchronous excitation of most cerebral neurons and intracerebral pathways, do not affect remote declarative memory (Greenberg and Kellner, 2005; Squire et al., 1976). Together, these data, interpreted within the SPH, lead to an apparent logical paradox: declarative memory that is assumed to be based on activity-dependent modifications in synaptic connections is resistant to synaptic activity which uses the same exact synapses. The direct conclusion from this paradox is that, contrary to the SPH, the same mechanism is not used to save and retrieve memory. To save information, physical changes are produced in the neurons serving as memory carriers, and these changes must not to be affected by memory reactivations or other types of synaptic activity. 2.3. Memory acquisition and storage have different localizations Memory of new facts and events is progressively consolidated from an unstable state, vulnerable to different interventions (such as an electroshock, epileptic episode, brain trauma, hypothermia, etc.), to a stable state, resistant to these interventions (Dudai, 2004; McGaugh, 2000; Squire, 1992; Squire et al., 2001). The process of consolidation includes different aspects. One of these aspects is the role of different cerebral structures in memory consolidation. 104 Y.I. Arshavsky / Progress in Neurobiology 80 (2006) 99–113 The MTL (which includes the hippocampus, as well as adjacent entorhinal, perirhinal, and parahippocampal cortices) plays a critical role in acquiring new information about facts and events. About 50 years ago, the patient H.M. underwent a bilateral resection of most of his MTL, which resulted in persisted anterograde amnesia (Scoville and Milner, 1957; see also Milner et al., 1998; Squire and Kandel, 1999). Although his perceptual and cognitive abilities were not destroyed, and H.M. was able to hold immediate perceptions in his mind, all these perceptions were lost as soon as his attention was turned to some other object. For example, he could not remember what he had eaten for breakfast, recognize members of the hospital staff, or acquire the meaning of unknown words. The inability of patients with bilateral lesions of the MTL to acquire new semantic knowledge was confirmed in further clinical observations (Bayley and Squire, 2005; Rosenbaum et al., 2005). Although the MTL plays an enabling role in learning and memory consolidation, it has a time-limited significance in memory storage (Bayley et al., 2005; Milner et al., 1998; Squire, 1992; Squire and Kandel, 1999; Squire et al., 2001). In patients with damage to the MTL, recent memory suffers, whereas remote memory is spared. For example, H.M. cannot acquire the meaning of new words, but remembers his native language, and has no evident problems in language comprehension, production, and reading. Although he did not recognize members of the hospital staff, he could recognize Dr. Scoville (see Scoville and Milner, 1957) whom he had met before his operation.3 In a recent study with an improved quantitative method for memory scoring, it has been shown that patients with large MTL lesions can still recall autobiographical memories from early in their lives, at the same rate as control individuals (Bayley et al., 2005). In addition to clinical observations, numerous experimental studies have shown the critical role of the MTL in acquisition of new information and its transient role in memory storage (see Frankland and Bontempi, 2005; Wiltgen et al., 2004; Wang et al., 2006 for recent reviews). I will describe four examples for illustration: (i) Inhibitors of protein synthesis administered before or shortly after training on rats prevented memory formation both when they were infused systemically and when they were locally applied to the hippocampus (Taubenfeld et al., 2001). 3 There is one more reason why the H.M. case is essential to understanding the mechanism of memory consolidation. H.M. was surgically treated to help his epilepsy, which originated within the MTL. He had at least one major seizure nearly each week. According to the SPH, which assumes that plasticity is an inherent property of synapses, each seizure had to radically rewire the existing patterns of synaptic connections and, therefore, to destroy memories consolidated before the beginning of each epileptic attack. Evidently, this is not the case, as H.M. remembers fact and events acquired before the surgery. This is one of the examples (see Section 2.2) that disprove the idea that the memory storage is based on activity-dependent structural modifications of synaptic connections. (ii) Lesions in the hippocampus abolished contextual fear memory in rats trained for 1 day before the lesion, but not in those trained for 28 days before the lesion (Anagnostaras et al., 1999). In contrast, lidocaine-induced inactivation of the prefrontal cortex disrupted the retrieval of remote (18 or 36 days after training), but not recent (1 or 3 days) contextual fear memories (Frankland et al., 2004). Similarly, lidocaine-induced inactivation of the hippocampus or the anterior cingulate cortex blocked recent or remote spatial memory, respectively (Teixeira et al., 2006; but see Broadbent et al., 2006 who found that lidocaineinduced inactivation of the hippocampus disrupted both recent and remote spatial memory). (iii) Monkeys with hippocampal lesions were severely impaired in discriminating recently learned objects. However, they remembered objects they had learned long ago as well as normal monkeys did (Zola-Morgan and Squire, 1990; compare with results of clinical observations by Bayley et al., 2005). (iv) Results of imaging studies on rodents showed that memory retrieval 1–5 days after learning was associated with increased activity in the hippocampus and related structures, and low activity in the neocortex. In contrast, the same tests taken 3 weeks later were associated with low hippocampal and increased neocortical activity (Bontempi et al., 1999; Ross and Eichenbaum, 2006). Thus, although declarative memory is primarily dependent on the MTL, it is eventually consolidated in the neocortex and probably in some other structures of the brain, which are the ultimate repositories for persistent MTL-independent memories. The role of the MTL in cortical memory consolidation is still poorly understood. However, regardless of the MTL’s role in cortical memory consolidation, the fact that different cerebral structures are used for temporary and permanent storage of information goes against the SPH, which states that ‘‘changes in the connection strength implement learning,’’ while memory retrieval is ‘‘the reactivation of assemblies of such neurons’’ (Maroun et al., 2001, see above). It is evident that the mode of memory representation in the neocortex cannot be a direct replica of the mode in which the information is originally acquired in the MTL because of different cytoarchitecture of the MTL and neocortex. This means that the nature of memory representation is different in these two structures and that the mechanisms used by the neocortex for the persistent storage of declarative memory is not equivalent to the mechanisms used by the MTL for learning and temporary maintenance of memory traces. In any case, memory retrieval cannot occur through the reactivation of the same neural assemblies that are involved in initial learning, because these two processes have different localizations. 2.4. ‘‘Synaptic’’ and ‘‘system’’ memory consolidations have different temporal characteristics The process of memory consolidation from the unstable to stable state takes a long time and occurs in several stages Y.I. Arshavsky / Progress in Neurobiology 80 (2006) 99–113 (Eichenbaum, 1997; McGaugh, 2000; Squire, 1992; Squire and Kandel, 1999). Recently, the terms ‘‘synaptic’’ and ‘‘system’’ consolidation have been suggested to emphasize certain functional aspects of memory consolidation (Dudai, 2004; Frankland and Bontempi, 2005; Wang et al., 2006). Synaptic consolidation includes the signaling cascade outlined above (see Section 2.1), which is initiated by hippocampal inputs and lead to protein synthesis-dependent modifications of synaptic connections. The duration of synaptic consolidation is measured by the period of time when memory is sensitive to the inhibition of protein synthesis (Dudai, 2004). Synaptic consolidation is completed several hours after learning, and there is no evidence for the involvement of protein synthesis in further memory consolidation. System consolidation leading to the formation of persistent memory occurs in the neocortex (see Section 2.3). This process of memory conservation takes weeks, months, and even years (in humans) to be completed. The duration of synaptic consolidation is measured by the time it takes for memory to become independent on the MTL (Dudai, 2004). Between synaptic and system consolidation, there is a prolonged transient period when the memory storage still depends on the MTL. Little is known about the physiological mechanisms underlying system consolidation and mode of the information storage within the neocortex. The terms ‘‘synaptic’’ and ‘‘system’’ consolidation seem to suggest different mechanisms for the transient memory storage in the MTL and the persistent storage in the neocortex. However, due to a lack in any alternative to the SPH, it is automatically assumed that system consolidation of memory, like synaptic consolidation, is also based on structural modifications in synaptic connections within the neocortex (Wang et al., 2006; Wiltgen et al., 2004). As claimed by Wiltgen et al. (2004), ‘‘at least some of the molecules involved in hippocampal plasticity (e.g., PKA, PKC, MAPK, CREB) may also be required for remote memory storage in cortical networks.’’ This suggestion is based on the following two findings: (i) It is well known that triggering the intracellular signaling cascade underlying synaptic consolidation in the MTL requires the activation of NMDA receptors (Nakazawa et al., 2004; Wittenberg and Tsien, 2002). To study the role of the NMDA receptors in system consolidation, Cui et al. (2004) used transgenic mice in which the NMDA receptors can be temporarily switched off in the forebrain. It was found that 30-day-switching-off the NMDA receptors (6 months after initial training and at least 2 months prior to memory retrieval) resulted in severe disruption of fear memories. (ii) One of the enzymes essential for hippocampus-dependent learning is a-calcium-calmodulin kinase II, aCaMKII (Lisman et al., 2002; Wiltgen et al., 2004; Wang et al., 2006). The role of this kinase in system memory consolidation was studied on heterozygous aCaMKII+/ mice (Elgersma et al., 2004; Frankland et al., 2001). These mice were found to have an impaired spatial and contextual 105 memory when tested 10–50 days after training. It was concluded that aCaMKII is involved in system memory consolidation.4 Neither of these two findings can be regarded as evidence for the use of the same mechanisms for both synaptic consolidation in the MTL and system consolidation in the neocortex. (i) Glutamate is the principal excitatory neurotransmitter in the cortex. Therefore, there are many reasons why inactivation of NMDA receptors can affect consolidation and/or retrieval of system memory. (ii) aCaMKII is a multifunctional enzyme and, even a confirmation of its role in system consolidation cannot be regarded as compelling evidence that long-term memory storage in the neocortex is based on protein synthesisdependent modifications in synaptic connections. On the other hand, different durations for the two types of memory consolidation, as well as the absence of evidence demonstrating the requirement of protein synthesis for system consolidation, support the aforementioned suggestion (see Section 2.3) that primary synaptic consolidation in the MTL and persistent system consolidation in the neocortex may have radically different mechanisms. 2.5. Reconsolidation of memory As mentioned earlier, newly learned memories go through an initial fragile stage before being consolidated into stable, long-term memories. According to the SPH, memory stability is based on irreversible structural modifications of synaptic connections. However, in the 1960s and 1970s, a puzzling phenomenon, seemingly incompatible with the SPH, was found. In rodents, different amnestic factors (electroconvulsive shock, hypothermia, head injury, etc.) could destroy wellconsolidated memory when applied directly after memory retrieval (Mactutus et al., 1979; Misanin et al., 1968; see Sara, 2000 for review). Particularly, Judge and Quartermain (1982) found that the protein synthesis inhibitor, anisomycin produced amnesia for well-consolidated avoidance reactions, provided that it was injected immediately after conditional stimulus. These findings suggest that retrieval can render consolidated memory into the initial fragile state and that de novo protein synthesis is required for memory reconsolidation. The phenomenon of memory reconsolidation was recently confirmed in a study on fear memory. The site for fear memory acquisition is the amygdala, and infusion of anisomycin into the amygdala directly after training prevents the consolidation of fear conditioning. The same effect was produced by the infusion of anisomycin into the amygdala after memory reactivation, regardless of whether retrieval was performed 1 or 14 days after conditioning (Nader et al., 2000). The authors concluded that this phenomenon of memory reconsolidation after retrieval is not ‘‘predicted by traditional theories of memory consolidation,’’ since it seems unlikely that ‘‘retrieval 4 This conclusion should be regarded with some caution, as it was recently found that CaMKII contributes to short-term memory, but is not necessary for long-term memory (Irvine et al., 2005, 2006). 106 Y.I. Arshavsky / Progress in Neurobiology 80 (2006) 99–113 reverses structural modifications of synaptic connections induced by learning.’’ The work by Nader et al. (2000) initiated a lot of new studies in this field. Most of these studies confirmed that traces of memory, including system memories (Debiec et al., 2002; Duvarci and Nader, 2004; Frankland et al., 2006; but see Milekic and Alberini, 2002 for the opposite result), undergo postretrieval reconsolidation (reviewed by Dudai, 2004; Nader, 2003). But in spite of the discovery of postretrieval reconsolidation, which seemed to ‘‘overturn the old dogma’’ (Nader, 2003), the SPH was not overturned. Very complex logical constructions were suggested to bring together the SPH and memory reconsolidation (see Dudai, 2004 and Millin et al., 2001, for example). However, these logical constructions can hardly explain the phenomenon of postretrieval memory reconsolidation within the bounds of the SPH. Rather, they demonstrate authors’ unwillingness to go beyond the traditional hypothesis. Meanwhile, the viewpoint that persistent declarative memory is stored in the brain through structural modifications in synaptic connections is incompatible with the phenomenon of memory reconsolidation after retrieval. It does seem unlikely that each memory reactivation produces at least a transient reversal of structural modifications in the synaptic connections formed during memory consolidation. The existence of the phenomenon of postretrieval reconsolidation of consolidated memory (if this phenomenon does really exist, see Alberini, 2005; Alberini et al., 2006; Biedenkapp and Rudy, 2004; Cammarota et al., 2004; Milekic and Alberini, 2002 who suppose that the phenomenon of reconsolidation is related to recent memory only) requires a radical revision of the existing concept on the mechanism of the memory storage. 2.6. Neurogenesis in the adult brain For many years, it was commonly accepted that new neurons are not produced in the adult mammalian brain. Neuron stability in the brain was considered to contribute to the stability of new patterns of neural activity formed over the course of memory consolidation. As stated by Rakic (1985), ‘‘A stable population of neurons in primates . . . may be important for the continuity of learning and memory over a lifetime.’’ The traditional view on the absence of neurogenesis in the adult brain has been called into question by recent studies demonstrating neurogenesis in adult rodents and primates, including humans (Eriksson et al., 1998; Gould et al., 1999a; Gross, 2000; Kempermann et al., 2004; Kornack and Rakic, 1999; Lledo et al., 2006; Ming and Song, 2005; Schinder and Gage, 2004). Adult neurogenesis takes place mainly in the hippocampus. The presence of neurogenesis in the neocortex, however, is disputable (cf. Gould et al., 1999b and Rakic, 2002). In rats, about 9000 new granule cells are produced each day in the dentate gyrus of the hippocampus; the number of new neurons generated each month is about 6% of the total size of the cell population (Cameron and McKay, 2001). The adultgenerated cells form synaptic connections with other neurons and, therefore, are integrated in hippocampal circuits (Gross, 2000; Liu et al., 2003; Lledo et al., 2006; Ming and Song, 2005; Schinder and Gage, 2004). Gross (2000) concluded that the existence of adult-generated neurons may lead to ‘‘the development of entirely new circuits with new and previously unused elements as well as the modulation of older circuits and connections.’’ The continual incorporation of new-born neurons into existing hippocampal circuits may be a source of the instability in neural interconnections established in the process of learning and hippocampus-dependent memory consolidation (see Sections 2.3 and 2.4). The contradiction between the SPH and the continuous incorporation of new neurons into the hippocampal circuits was highlighted by Nottebohm (2002) as follows: ‘‘A theory of learning based on synapses did not predict a need for replacing healthy adult neurons by other neurons of the same kind, and so something basic must have been missing. If synaptic plasticity did not predict neuronal replacement, how must we reconstitute our logic of brain function so that both phenomena, synaptic plasticity and spontaneous neuronal replacement, can coexist, conceptually, in a harmonious manner?’’ In contrast to the prediction of the SPH that adult neurogenesis may cause the erosion of hippocampus-dependent memory, results obtained by some authors suggest that adult neurogenesis improves an acquisition and initial consolidation of new information (Gould et al., 1999c; Gross, 2000; Schinder and Gage, 2004). This notion was supported by the following facts. The stimulating effect of environmental enrichment and physical activity on adult hippocampal neurogenesis improved hippocampus-dependent learning (Kempermann et al., 1997; Nilsson et al., 1999; van Praag et al., 1999). On the other hand, the suppression of cell proliferation in the hippocampus by Xor gamma-irradiation and by chronic treatment with an antimitotic agent, methylazoxymethanol, impaired hippocampusdependent learning (Bruel-Jungerman et al., 2005; Madsen et al., 2003; Shors et al., 2001; Snyder et al., 2005). The functional role of hippocampal adult neurogenesis in learning and memory remains to be established (see Aimone et al., 2006; Lledo et al., 2006). The most accepted viewpoint is that adult neurogenesis increases network capacities for the acquisition of new information (Kempermann, 2002; Schinder and Gage, 2004; Gross, 2000; Tashiro et al., 2006; Wang et al., 2006). For example, according to Gross (2000), after completing memory consolidation in the neocortex, old hippocampal neurons may die, ‘‘making room for new naive neurons that would then function similarly in consolidating new memories.’’ However, it is important to be aware that the SPH does not predict the necessity of replacing healthy neurons to create more room for new memories (Nottebohm, 2002; see above). According to the SPH, acquisition of new information requires a re-modification of the patterns of synaptic connections – that is, deleting the synapses related to ‘‘old memories’’ and creating new synaptic connections, but not the generation of new neurons. Adult neurogenesis can create more room for learning new tasks only if memory consolidation occurs not at the synaptic level, but within individual neurons. Y.I. Arshavsky / Progress in Neurobiology 80 (2006) 99–113 2.7. The synaptic plasticity hypothesis does not explain the specific memory impairments present in Alzheimer’s disease Alzheimer’s disease (AD) is a neurodegenerative disease characterized by the formation of amyloid plaques and neurofibrillary tangles. AD starts as the specific disease of memory, while other neurological functions are intact. As emphasized in a recent review, ‘‘Alzheimer’s disease characteristically produces a remarkably pure impairment of declarative memory in its earliest stages’’ (Walsh and Selkoe, 2004). Correspondingly, the degeneration of neurons leading to AD is area-specific. It occurs primarily in the cerebral regions related to the acquisition and storage of declarative memory (the hippocampus and entorhinal cortex, amygdala, and association cortices), but not in the primary sensory and motor cortices (Price, 2000; Selkoe, 2001). Within the memoryrelated regions, specific neuronal populations are particularly vulnerable, whereas other neurons are spared. It appears that degeneration selectively affects those neurons which serve as carriers of memory traces. Since AD starts as the specific disease of memory, any hypothesis on the mechanism of learning and memory consolidation has to include an explanation of this specificity (see Small et al., 2001). Why are the neurons involved in learning and memory storage more vulnerable, versus other neurons, to the genetic and cardiovascular risk factors for AD? Guided by the SPH, many authors concentrated on studying LTP in transgenic mice that develop an AD-like phenotype due to expression of human mutant tau and/or mutant amyloid b protein precursor proteins. However, the results of these studies are extremely inconsistent. Although some studies reported age-related deficits (although usually not radical) in the induction and/or maintenance of LTP in transgenic versus wild-type mice (Chapman et al., 1999; Gureviciene et al., 2004; Jacobsen et al., 2006; Oddo et al., 2003; Trinchese et al., 2004), others found no change or even a robust increase of LTP tested on the hippocampus and prefrontal cortex (Fitzjohn et al., 2001; Hsia et al., 1999; Jolas et al., 2002; Larson et al., 1999; Roder et al., 2003). Thus, results obtained in transgenic mice can hardly be regarded as evidence for a link between decreased synaptic plasticity tested by changes in LTP and AD-like phenotype. Several authors, who did not find any changes of LTP, reported deficits of basal synaptic functions in amyloid b protein precursor transgenic mice (Hsia et al., 1999; Fitzjohn et al., 2001). Correspondingly, biochemical and morphological analyses of brain tissue taken from AD patients led to conclusion that synapses in the hippocampus and association cortices are the initial targets in AD (Davies et al., 1987; Terry et al., 1991; see Small et al., 2001 and Selkoe, 2002 for reviews). However, deficits in basal synaptic transmission can hardly explain why AD starts with specific impairments of learning and memory because not only memory, but all brain functions equally depend on synaptic transmission. One can conclude that the specific impairment of memory in the earliest stage of AD can hardly be interpreted within the 107 context of the SPH. The specificity of synapses is mainly determined by the transmitter released from presynaptic terminals and accepted by receptors of target neurons. Since synaptic connections newly formed during learning and memory consolidation are unlikely to use specific ‘‘memory’’ transmitters, it is plausible to suggest that memory consolidation involves not only synaptic, but also intracellular modifications. In this case, the selective vulnerability of the memory-related neurons to risk factors for AD can be regarded as the result of specific intraneural modifications that arise in the process of memory consolidation (see below). 3. Alternative (genomic) hypothesis of memory Here I have tried to show that all the ‘‘sins’’ of the SPH are rooted in its major intrinsic self-contradiction. It is hardly possible to build stable declarative memory on such a nonstable foundation as synaptic plasticity. The phenomenon of synaptic plasticity may be used for learning and transient memory saving. However, persistent memory which lasts throughout the entire lifespan must be ‘‘etched in stone.’’ The only ‘‘stone-like’’ molecules in the living cells are the molecules of DNA. Therefore, it is plausible to hypothesize that acquired information is persistently stored within individual neurons through modifications of DNA, and that these modifications serve as the carriers of elementary memory traces. This idea (the genomic hypothesis of memory5) was distinctly proclaimed by Davis and Squire (1984) and Crick (1984) in their attempt to explain the persistence of memory (see also Arshavsky, 2003a; Been and Dietrich, 2004; De Fonzo et al., 2000; Dietrich and Been, 2001; Nottebohm, 2002; Peña de Ortiz and Arshavsky, 2001; Peña de Ortiz et al., 2004): ‘‘Some long-term memories may last days, weeks, or even many years. . . . This suggests that the maintenance of longterm memories cannot be based solely on either posttranslational protein modification or the synthesis of new proteins at or near the time of new learning. This suggestion is based on observation that brain proteins have a half-life on the order of days to weeks . . . It is possible that neurons have genetic commands to maintain those molecular or morphological alterations that occur when memory is initially established . . .’’ (Davis and Squire, 1984). ‘‘The time span of human memory (without obvious rehearsal) is often a matter of years, sometimes even tens of years. Yet . . . almost all the molecules in our bodies, with the exception of DNA, turn over in a matter of days, weeks or at the most a few months . . . Several possible solutions of the problem suggest themselves. For example, memory might be coded in alterations to particular stretches of chromosomal DNA’’ (Crick, 1984). 5 In previous publications, the term ‘‘chemical hypothesis of memory’’ was used instead of ‘‘genomic hypothesis’’ (Arshavsky, 2003a,b; Peña de Ortiz and Arshavsky, 2001). 108 Y.I. Arshavsky / Progress in Neurobiology 80 (2006) 99–113 There are two attempts to specify the genomic hypothesis of memory. One of them suggests that persistent memory storage occurs either via the mechanism of somatic DNA recombination similar (although not necessary identical) to V(D)J recombination in cells of the immune system (Arshavsky, 2003a,b, 2006a; Peña de Ortiz and Arshavsky, 2001; Peña de Ortiz et al., 2004) or via the meiotic mechanism of DNA recombination acting in an early stage of postnatal development (Dietrich and Been, 2001; Been and Dietrich, 2004). Some experimental data supporting the hypothesis on using somatic V(D)J-like DNA recombination for the memory storage have been obtained (Colón-Cesario et al., 2006; McGowan et al., 2004; Peña de Ortiz et al., 2003; Ren and Peña de Ortiz, 2002; Wang et al., 2003). In particular, it was found that a blockade of enzymes participating in the process of somatic DNA recombination impairs learning and memory consolidation. Although these results cannot be regarded as compelling evidence for the genomic mechanism of memory, they may stimulate further investigation in this direction. The other version of the genomic hypothesis suggests that the persistence of memory is based on epigenetic modifications (Holliday, 1999; Levenson and Sweatt, 2005, 2006; Levenson et al., 2006). It is known that DNA molecules associate with proteins (histones) to form chromatin. Epigenetic modifications alter the structure of the chromatin due to chemical changes to the DNA or histones without changing the DNA nucleotide sequence. Changes to the chromatin structure exert a profound effect on the cell-specific pattern of gene expression and, as a result play an important role in cell differentiation, including neuron differentiation (Hsieh and Gage, 2005; Levenson and Sweatt, 2006). Epigenetic modifications produced by a brief signaling event can be very stable and maintained throughout the entire lifespan of a cell. Therefore, it was hypothesized that epigenetics may be used as a mechanism for persistent memory consolidation. Experimental support for this hypothesis came from studies by Sweatt and coworkers, who found increased histone acetylation and phosphorylation in hippocampal neurons of rats during consolidation of contextual fear memory (Chwang et al., 2006; Levenson et al., 2004). Although the idea that traces of declarative memory are stored at the level of modified DNA molecules has obtained little recognition, the genomic hypothesis of memory seems to be promising. Here, I will illustrate that the aforementioned problems faced by the SPH can be overcome within the genomic hypothesis: (1) The modified genes would provide a stable basis for the storage of memory, as presumed DNA changes arising in the process of memory consolidation cannot be reversed or altered. When describing the consolidation of memory from a labile to stable state, Dudai (2004) used the term which he named Lady Macbeth’s Principle: ‘‘What’s done cannot be undone.’’ This principle can hardly be applied to synaptic rewiring, which can be altered and undone, but is completely applicable to DNA modifications. Memory traces coded in modified DNA can be only deleted from the brain upon the death of the neurons storing those memory traces. (2) The genomic hypothesis of memory suggests that different mechanisms are used for memory storage and retrieval. This is the reason why memory engrams are resistant to synaptic activity. Neuron excitation (regardless of how strong it is) does not affect once-produced DNA modifications. (3) and (4) The genomic hypothesis does not suggest the same mechanism (i.e., rewiring of synaptic connections) for learning and for permanent memory storage. Therefore, it does not assume that memory retrieval results from reactivating the same neural assemblies involved in the primary encoding of the information. This means that the differences between ‘‘synaptic memory’’ and ‘‘system memory’’ (their different localizations, the different temporal characteristics of their consolidation, and the fact that the former depends on protein synthesis – whereas the latter does not) do not contradict the genomic hypothesis (unlike the SPH, which is contradicted on all accounts). (5) As highlighted above, the reconsolidation of system memory (if this phenomenon does exist) contradicts the SPH and requires its radical revision. In contrast, the phenomenon of memory reconsolidation is not crucial to the genomic hypothesis. One can suggest that the proteins produced by modified genes, which serve as carriers of memory traces, may be depleted during retrieval, necessitating the production of new proteins for memory restoration. This notion, while speculative, may help to make sense of such puzzling phenomena as postretrieval memory reconsolidation. A direct prediction following from this notion is that inhibitors of protein synthesis (which completely block memory consolidation when administered after learning) should cause only a temporary amnesia when applied after memory reactivation. Indeed, some studies have found that inhibitors of protein synthesis applied directly after retrieval do produce a temporary loss of memory, which then recovers spontaneously (Anokhin et al., 2002; Judge and Quartermain, 1982; Lattal and Abel, 2004; Power et al., 2006; but see Debiec et al., 2002; Duvarci and Nader, 2004 for the opposite result). These findings suggest that, while the inhibition of protein synthesis after learning interferes with memory consolidation, the inhibition of protein synthesis after memory reactivation results in a transient retrieval deficit and does not interfere with the engram. (6) It is difficult to discuss the role of adult neurogenesis in learning and memory, since the literature on this issue is full of both conceptual and experimental contradictions (see Leuner et al., 2006). The prevalent viewpoint is that incessant renewal of adult hippocampal neurons is necessary to delete old memories after their permanent consolidation within the neocortex, to make room for the acquisition of new memories. As emphasized above, the SPH does not predict the necessity of replacing healthy neurons to create more room for new memories. According to this hypothesis, the acquisition of new information requires a re-modification of the patterns of synaptic connections – deleting the synapses related to ‘‘old memories’’ and creating new synaptic connections. In Y.I. Arshavsky / Progress in Neurobiology 80 (2006) 99–113 contrast, adult neurogenesis can create more room for acquisition of new information if memory is consolidated at the intracellular – genomic – level, but not at the synaptic level. (7) The genomic hypothesis provokes a new approach to understanding the etiology of AD as specific disease of memory. The idea that traces of declarative memory are stored at the genomic level suggests that modified pattern of gene expression can produce new proteins that were not expressed during embryogenesis and early postnatal development. If these proteins migrate to the neuron surface, they must be recognized by the immune system as ‘‘non-self’’ antigens. It was suggested that the immune reaction against one’s own neurons serving as carriers of memory traces are prevented by the blood–brain barrier, which protects the brain from the entry of immunoglobulins and strongly restricts the penetration of T lymphocytes (Arshavsky, 2003a, 2006a; Peña de Ortiz and Arshavsky, 2001; Peña de Ortiz et al., 2004). Results of numerous studies have shown that all risk factors for AD – both cardiovascular (atherosclerosis, hypertension, stroke, etc.) and genetic (mutations of the amyloid precursor protein and presenilin 1 and 2 genes, as well as the inheritance of the E4 allele of the apolipoprotein E gene) – lead to the destruction of the blood–brain barrier (de la Torre, 2004; Iadecola, 2004; Jellinger and Attems, 2005; Jeynes and Provias, 2006; Sadowski et al., 2004; Zlokovic, 2005). In experiments on transgenic mice developing an AD-like phenotype, it was found that impairments of the blood–brain barrier emerged prior any signs of neuron degeneration and memory deficits (Ujiie et al., 2003). Within the outline of the genomic hypothesis, AD can be interpreted as an autoimmune disease associated with blood–brain barrier impairments (see Arshavsky, 2006a for detail).6 This interpretation explains why the process of neuron degeneration during AD, at least in its earliest stages, specifically affects the neurons implicated in the consolidation and storage of memory traces. As to a loss of mental functions and motor disability that develop in later stages of AD, they can result from secondary non-specific inflammation (Zipp and Aktas, 2006). 4. Concluding remarks At present, the Hebbian hypothesis of synaptic plasticity is practically the only hypothesis on cellular mechanisms of memory that is widely recognized. All current knowledge on learning and the initial stage of memory consolidation (synaptic consolidation) is based on this hypothesis. However, due to the lack of rival hypotheses, the SPH has been used not only to explain learning and initial memory consolidation, but also for 6 Recently the idea on the autoimmune origin of AD was advocated by D’Andrea who studied hippocampal and entorhinal tissues from AD patients by using polyclonal anti-Ig antibodies. As compared to the age-matched controls, a significant increase in Ig staining was detected in both parenchyma and individual neurons of AD tissue (D’Andrea, 2003, 2005). 109 permanent (system) consolidation. The major objective of this paper is to illustrate that this approach to understanding longterm declarative memory faces a series of conceptual and experimental problems that are unlikely to be resolved within the context of the SPH. All these problems are rooted in the attempt to use the intrinsically unstable phenomenon of synaptic plasticity to explain the mechanism of stable memory, which can be stored in the brain throughout decades and even an entire lifespan. Potentially permanent declarative memory requires a different mechanism – one that can provide more stability than synaptic plasticity, which is based on transient changes in gene transcription and protein synthesis. Here, I advocate the alternative, genomic hypothesis, which suggests that memory traces are stored within individual neurons through genomic modifications. The advantage of this hypothesis is that genomic modifications forming in the process of memory consolidation cannot be altered or undone. In other words, this hypothesis suggests that memory consolidation is akin to the irreversible steps that occur in the process of cell differentiation (see Nottebohm, 2002; Levenson et al., 2006; Levenson and Sweatt, 2006). The genomic hypothesis of declarative memory is part of a more general hypothesis on the role of cellular and network mechanisms in performing complex cognitive functions closely linked to consciousness (Arshavsky, 2001, 2003a,c, 2006b). This hypothesis suggests that cognitive functions are performed not by neural networks consisting of simple neurons (see Introduction), but by networks consisting of ‘‘complex’’ neurons that are carriers of ‘‘elementary cognition’’ – in particular, carriers of elementary traces of declarative memory. The main criticism of the genomic hypothesis (particularly, of the version developed by Peña de Ortiz and Arshavsky, see above) is that it does not offer any mechanistic explanation for memory retrieval. In fact, this criticism implies that the commonly accepted SPH offers such a mechanism – namely, the reactivation of neuronal assemblies which are formed in the process of learning. I think this argument is not fear. As shown above, memory retrieval cannot occur through the reactivation of the same neuronal assemblies which are involved in initial learning, because of the different localization of these two processes. Here, I would like to point out the principle impediment to understanding a mechanism of memory retrieval – an impediment that is related to any hypothesis of declarative memory. By definition, the retrieval of declarative memory in humans is a conscious recollection of facts and events. This means that the question about the mechanism of memory retrieval is tightly linked to the much more fundamental question about the cellular mechanisms of consciousness. It is futile to hope that understanding memory retrieval can be possible without addressing the nature of consciousness, which remains beyond the scope of modern neuroscience. Therefore, this paper does not pretend to discuss the mechanism for retrieving declarative memory. Rather, its objective is: (i) to emphasize the monopoly of the pure connectionist approach on understanding memory consolidation and storage, (ii) to show the main limitations of this approach, and (iii) to stimulate alternative, cellular-oriented approaches to the problem. 110 Y.I. Arshavsky / Progress in Neurobiology 80 (2006) 99–113 References Abraham, W.C., Williams, J.M., 2003. Properties and mechanisms of LTP maintenance. Neuroscience 9, 463–474. Aimone, J.B., Wiles, J., Gage, F.G., 2006. Potential role for adult neurogenesis in the encoding of time in new memories. Nat. Neurosci. 9, 723–727. Alberini, C.M., 2005. Mechanisms of memory stabilization: are consolidation and reconsolidation similar or distinct processes? Trends Neurosci. 28, 51– 56. Alberini, C.M., Milekic, M.H., Tronel, S., 2006. Mechanisms of memory stabilization and de-stabilization. Cell. Mol. Life Sci. 63, 999–1008. Anagnostaras, S.G., Maren, S., Fanselow, M.S., 1999. Temporally graded retrograde amnesia of contextual fear after hippocampal damage in rats: within-subjects examination. J. Neurosci. 19, 1106–1114. Anokhin, K.V., Tiunova, A.A., Rose, S.P., 2002. Reminder effects – reconsolidation or retrieval deficit? Eur. J. Neurosci. 15, 1759–1765. Arshavsky, Y.I., 2001. Role of individual neurons and neural networks in cognitive functioning of the brain: a new insight. Brain Cogn. 46, 414–428. Arshavsky, Y.I., 2003a. Cellular and networks properties in the functioning of the nervous system: from central pattern generators to cognition. Brain Res. Rev. 41, 229–267. Arshavsky, Y.I., 2003b. Long-term memory: does it have a structural or chemical basis? Trends Neurosci. 26, 465–466. Arshavsky, Y.I., 2003c. When did Mozart become a Mozart? Neurophysiological insight into behavioral genetics. Brain Mind 4, 327–339. Arshavsky, Y.I., 2006a. Alzheimer’s disease, brain immune privilege and memory: a hypothesis. J. Neural. Transm. 113, 1697–1707. Arshavsky, Y.I., 2006b. ‘‘Scientific roots’’ of dualism in neuroscience. Prog. Neurobiol. 79, 190–204. Bahrick, H.P., 1984. Semantic memory content in permastore: fifty years of memory for Spanish learned in school. J. Exp. Psychol. Gen. 113, 1–29. Bahrick, H.P., Bahrick, P.O., Wittlinger, R.P., 1975. Fifty years of memories for names and faces. J. Exp. Psychol. Gen. 104, 54–75. Bailey, C.H., 1999. Structural changes and the storage of long-term memory in Aplysia. Can. J. Physiol. Pharmacol. 77, 738–747. Bailey, C.H., Kandel, E.R., Si, K., 2004. The persistence of long-term memory; a molecular approach to self-sustaining changes in learning-induced synaptic growth. Neuron 44, 49–57. Bailey, D.J., Kim, J.J., Sun, W., Thompson, R.F., Helmstetter, F.J., 1999. Acquisition of fear conditioning in rats requires the synthesis of mRNA in the amygdala. Behav. Neurosci. 113, 276–282. Bayley, P.J., Gold, J.J., Hopkins, R.O., Squire, L.R., 2005. The neuroanatomy of remote memory. Neuron 46, 799–810. Bayley, P.J., Squire, L.R., 2005. Failure to acquire new semantic knowledge in patients with large medial temporal lobe lesions. Hippocampus 15, 273– 280. Been, W., Dietrich, A., 2004. DNA recombination, memory storage and learning. In: Parisi, V., De Fonzo, V., Aluffi-Pentini, F. (Eds.), Dynamical Genetics. Research Signpost, India, pp. 365–380. Benson, D.L., Colman, D.R., Huntley, G.W., 2001. Molecules, maps and synapse specificity. Nat. Rev. Neurosci. 2, 899–909. Biedenkapp, J.C., Rudy, J.W., 2004. Context memories and reactivation: constraints on the reconsolidation hypothesis. Behav. Neurosci. 118, 956–964. Blair, T.H., Schafe, G.E., Bauer, E.P., Rodrigues, S.M., LeDoux, J.E., 2001. Synaptic plasticity in the lateral amygdala: a cellular hypothesis of fear conditioning. Learn. Mem. 8, 229–242. Bliss, T.V., Lømo, T., 1973. Long-lasting potentiation of synaptic transmission in the dentate area of the anaesthetized rabbit following stimulation of the perforant path. J. Physiol. (Lond.) 232, 331–356. Bontempi, B., Laurent-Demir, C., Destrade, C., Jaffard, R., 1999. Timedependent reorganization of brain circuitry underlying long-term memory storage. Nature 400, 671–675. Broadbent, N.J., Squire, L.R., Clark, R.E., 2006. Reversible hippocampal lesions disrupt water maze performance during both recent and remote memory tests. Learn. Mem. 13, 187–191. Bruel-Jungerman, E., Laroche, S., Rampon, C., 2005. New neurons in the dentate gyrus are involved in the expression of enhanced long-term memory following environmental enrichment. Eur. J. Neurosci. 21, 513–521. Cameron, H.A., McKay, R.D., 2001. Adult neurogenesis produces a large pool of new granule cells in the dentate gyrus. J. Comp. Neurol. 435, 406– 417. Cammarota, M., Bevilaqua, L.R., Medina, J.H., Izquierdo, I., 2004. Retrieval does not induce reconsolidation of inhibitory avoidance memory. Learn. Mem. 11, 572–578. Chapman, P.F., White, G.L., Jones, M.W., Cooper-Blacketer, D., Marshall, V.J., Irizarry, M., Younkin, L., Good, M.A., Bliss, T.V., Hyman, B.T., Younkin, S.G., Hsiao, K.K., 1999. Impaired synaptic plasticity and learning in aged amyloid precursor protein transgenic mice. Nat. Neurosci. 2, 271–276. Cheek, D.B., LeCron, L.M., 1968. Clinical Hypnotherapy. Grune a. Stratton, New York. Chklovskii, D.B., Mel, B.W., Svoboda, K., 2004. Cortical rewiring and information storage. Nature 431, 782–788. Chwang, W.B., O’Riordan, K.J., Levenson, J.M., Sweatt, J.D., 2006. ERK/ MAPK regulates hippocampal histone phosphorylation following contextual fear conditioning. Learn. Mem. 13, 322–328. Colón-Cesario, M., Wang, J., Ramos, X., Garcı́a, H.G., Dávila, J.J., Laguna, J., Rosado, G., Peña de Ortiz, S., 2006. An inhibitor of DNA recombination blocks memory consolidation, but not reconsolidation, in context fear conditioning. J. Neurosci. 26, 5524–5533. Crick, F., 1984. Memory and molecular turnover. Nature 312, 101. Cui, Z., Wang, H., Tan, Y., Zaia, K.A., Zhang, S., Tsien, J.Z., 2004. Inducible and reversible NR1 knockout reveals crucial role of the NMDA receptor in preserving remote memories in the brain. Neuron 41, 781–793. D’Andrea, M.R., 2003. Evidence linking neuronal cell death to autoimmunity in Alzheimer’s disease. Brain Res. 982, 19–30. D’Andrea, M.R., 2005. Add Alzheimer’s disease to the list of autoimmune diseases. Med. Hypoth. 64, 458–463. D’Angelo, E., Rossi, E., Gall, D., Prestory, F., Nieus, T., Maffei, A., Sola, E., 2005. Long-term potentiation of synaptic transmission at the mossy fibergranule cell relay of cerebellum. Prog. Brain Res. 148, 69–80. Davies, C.A., Mann, D.M., Sumpter, P.Q., Yates, P.O., 1987. A quantitative morphometric analysis of the neuronal and synaptic content of the frontal and temporal cortex in patients with Alzheimer’s disease. J. Neurol. Sci. 78, 151–164. Davis, H.P., Squire, L.R., 1984. Protein synthesis and memory: a review. Psychol. Bull. 96, 518–559. Debiec, J., LeDoux, J.E., Nader, K., 2002. Cellular and systems reconsolidation in the hippocampus. Neuron 36, 527–538. De Fonzo, V., Bersani, E., Aluffi-Pentini, F., Parisi, V., 2000. A new look at the challenging world of tandem repeats. Med. Hypoth. 54, 750–760. de la Torre, J.C., 2004. Alzheimer’s disease is a vasocognopathy: a new term to describe its nature. Neurol. Res. 26, 517–524. De Paola, V., Holtmaat, A., Knott, G., Song, S., Wilbrecht, L., Caroni, P., Svoboda, K., 2006. Cell type-specific structural plasticity of axonal branches and boutons in the adult neocortex. Neuron 49, 861–875. Dietrich, A., Been, W., 2001. Memory and DNA. J. Theor. Biol. 208, 145–149. Dudai, Y., 2004. The neurobiology of consolidations, or, how stable is the engram? Annu. Rev. Psychol. 55, 51–86. Duvarci, S., Nader, K., 2004. Characterization of fear memory reconsolidation. J. Neurosci. 24, 9269–9275. Eccles, J.C., 1972. Possible synaptic mechanism subserving learning. In: Karcmar, A.G., Eccles, J.C. (Eds.), Brain and Human Behavior. Springer-Verlag, New York, pp. 39–61. Eichenbaum, H., 1997. Declarative memory: insights from cognitive neurobiology. Annu. Rev. Psychol. 48, 547–572. Elgersma, Y., Sweatt, J.D., Giese, K.P., 2004. Mouse genetic approaches to investigating calcium/calmodulin-dependent protein kinase II function in plasticity and cognition. J. Neurosci. 24, 8410–8415. Engert, F., Bonhoeffer, T., 1999. Dendritic spine changes associated with hippocampal long-term synaptic plasticity. Nature 399, 66–70. Eriksson, P.S., Perfilieva, E., Bjork-Eriksson, T., Alborn, A.M., Nordborg, C., Peterson, D.A., Gage, F.H., 1998. Neurogenesis in the adult human hippocampus. Nat. Med. 4, 1313–1317. Y.I. Arshavsky / Progress in Neurobiology 80 (2006) 99–113 Fitzjohn, S.M., Morton, R.A., Kuenzi, F., Rosahl, T.W., Shearman, M., Lewis, H., Smith, D., Reynolds, D.S., Davies, C.H., Collingridge, G.L., Seabrook, G.R., 2001. Age-related impairment of synaptic transmission but normal long-term potentiation in transgenic mice that overexpress the human APP695SWE mutant form of amyloid precursor protein. J. Neurosci. 21, 4691–4698. Fligstein, D., Barabasz, A., Barabasz, M., Trevisan, M.S., Warner, D., 1998. Hypnosis enhances recall memory: a test of forced and non-forced conditions. Am. J. Clin. Hypn. 40, 297–305. Frankland, P.W., Bontempi, B., 2005. The organization of recent and remote memories. Nat. Rev. Neurosci. 6, 119–130. Frankland, P.W., Bontempi, B., Talon, L.E., Kaczmarek, L., Silva, A.J., 2004. The involvement of the anterior cingulate cortex in remote contextual fear memory. Science 304, 881–883. Frankland, P.W., Ding, H.K., Takahashi, E., Suzuki, A., Kida, S., Silva, A.J., 2006. Stability of recent and remote contextual fear memory. Learn. Mem. 13, 451–457. Frankland, P.W., O’Brien, C., Ohno, M., Kirkwood, A., Silva, A.J., 2001. Alpha-CaMKII-dependent plasticity in the cortex is required for permanent memory. Nature 411, 309–313. Gould, E., Reeves, A.J., Fallah, M., Tanapat, P., Gross, C.G., Fuchs, E., 1999a. Hippocampal neurogenesis in adult Old World primates. Proc. Natl. Acad. Sci. U.S.A. 96, 5263–5267. Gould, E., Reeves, A.J., Graziano, M.S., Gross, C.G., 1999b. Neurogenesis in the neocortex of adult primates. Science 286, 548–552. Gould, E., Tanapat, P., Hastings, N.B., Shors, T.J., 1999c. Neurogenesis in adulthood: a possible role in learning. Trends Cogn. Sci. 3, 186–192. Green, J.P., 1999. Hypnosis, context effects, and the recall of early autobiographical memories. Int. J. Clin. Exp. Hypn. 47, 284–300. Greenberg, R.M., Kellner, C.H., 2005. Electroconvulsive therapy: a selected review. Am. J. Geriatr. Psychiatry 13, 268–281. Grenier, F., Timofeev, I., Steriade, M., 2003. Neocortical very fast oscillations (ripples, 80–200 Hz) during seizures: intracellular correlates. J. Neurophysiol. 89, 841–852. Gross, C.G., 2000. Neurogenesis in the adult brain: death of a dogma. Nat. Rev. Neurosci. 1, 67–73. Grutzendler, J., Kasthuri, N., Gan, W.B., 2002. Long-term dendritic spine stability in the adult cortex. Nature 420, 812–816. Gureviciene, I., Ikonen, S., Gurevicius, K., Sarkaki, A., van Groen, T., Pussinen, R., Ylinen, A., Tanila, H., 2004. Normal induction but accelerated decay of LTP in APP + PS1 transgenic mice. Neurobiol. Dis. 15, 188–195. Hebb, D.O., 1949. The Organization of Behavior. Wiley, New York. Holliday, R., 1999. Is there an epigenetic component in long-term memory? J. Theor. Biol. 200, 339–341. Hsia, A.Y., Masliah, E., McConlogue, L., Yu, G.Q., Tatsuno, G., Hu, K., Kholodenko, D., Malenka, R.C., Nicoll, R.A., Mucke, L., 1999. Plaqueindependent disruption of neural circuits in Alzheimer’s disease mouse models. Proc. Natl. Acad. Sci. U.S.A. 96, 3228–3233. Hsieh, J., Gage, F.H., 2005. Chromatin remodeling in neural development and plasticity. Curr. Opin. Cell Biol. 17, 664–671. Iadecola, C., 2004. Neurovascular regulation in the normal brain and in Alzheimer’s disease. Nat. Rev. Neurosci. 5, 347–360. Igaz, L.M., Vianna, M.R., Medina, J.H., Izquierdo, I., 2002. Two time periods of hippocampal mRNA synthesis are required for memory consolidation of fear-motivated learning. J. Neurosci. 22, 6781–6789. Irvine, E.E., Vernon, J., Giese, K.P., 2005. AlphaCaMKII autophosphorylation contributes to rapid learning but is not necessary for memory. Nat. Neurosci. 8, 411–412. Irvine, E.E., von Hertzen, L.S., Plattner, F., Giese, K.P., 2006. aCaMKII autophosphorylation: a fast track to memory. Trends Neurosci. 29, 459–465. Jacobsen, J.S., Wu, C.C., Redwine, J.M., Comery, T.A., Arias, R., Bowlby, M., Martone, R., Morrison, J.H., Pangalos, M.N., Reinhart, P.H., Bloom, F.E., 2006. Early-onset behavioral and synaptic deficits in a mouse model of Alzheimer’s disease. Proc. Natl. Acad. Sci. U.S.A. 103, 5161–5166. Jellinger, K.A., Attems, J., 2005. Prevalence and pathogenic role of cerebrovascular lesions in Alzheimer disease. J. Neurol. Sci. 229, 37–41. Jeynes, B., Provias, J., 2006. The possible role of capillary cerebral amyloid angiopathy in Alzheimer lesion development: a regional comparison. Acta Neuropathol. (Berl.) 112, 417–427. 111 Jolas, T., Zhang, X.S., Zhang, Q., Wong, G., Del Vecchio, R., Gold, L., Priestley, T., 2002. Long-term potentiation is increased in the CA1 area of the hippocampus of APP(swe/ind) CRND8 mice. Neurobiol. Dis. 11, 394–409. Judge, M.E., Quartermain, D., 1982. Characteristics of retrograde amnesia following reactivation of memory in mice. Physiol. Behav. 28, 585–590. Kandel, E.R., 2000. Cellular mechanisms of learning and the biological basis of individuality. In: Kandel, E.R., Schwartz, J.H., Jessel, T.M. (Eds.), Principles of Neural Science. 4th ed. McGraw-Hill, New York, pp. 1247–1279. Kandel, E.R., 2001. The molecular biology of memory storage: a dialog between genes and synapses. Biosci. Rep. 21, 565–611. Kempermann, G., 2002. Why new neurons? Possible functions for adult hippocampal neurogenesis. J. Neurosci. 22, 635–638. Kempermann, G., Kuhn, H.G., Gage, F.H., 1997. More hippocampal neurons in adult mice living in an enriched environment. Nature 386, 493–495. Kempermann, G., Wiskott, L., Gage, F.H., 2004. Functional significance of adult neurogenesis. Curr. Opin. Neurobiol. 14, 186–191. Koch, C., Laurent, G., 1999. Complexity and the nervous system. Science 284, 96–98. Kornack, D.R., Rakic, P., 1999. Continuation of neurogenesis in the hippocampus of the adult macaque monkey. Proc. Natl. Acad. Sci. U.S.A. 96, 5768–5773. Lamprecht, R., LeDoux, J.E., 2004. Structural plasticity and memory. Nat. Rev. Neurosci. 5, 45–54. Larson, J., Lynch, G., Games, D., Seubert, P., 1999. Alterations in synaptic transmission and long-term potentiation in hippocampal slices from young and aged PDAPP mice. Brain Res. 840, 23–35. Lattal, K.M., Abel, T., 2004. Behavioral impairments caused by injections of the protein synthesis inhibitor anisomycin after contextual retrieval reverse with time. Proc. Natl. Acad. Sci. U.S.A. 101, 4667–4672. LeDoux, J., 2003. The self: clues from the brain. Ann. N. Y. Acad. Sci. 1001, 295–304. Leuner, B., Gould, E., Shors, T.J., 2006. Is there a link between adult neurogenesis and learning? Hippocampus 16, 216–224. Levenson, J.M., O’Riordan, K.J., Brown, K.D., Trinh, M.A., Molfese, D.L., Sweatt, J.D., 2004. Regulation of histone acetylation during memory formation in the hippocampus. J. Biol. Chem. 279, 40545–40559. Levenson, J.M., Roth, T.L., Lubin, F.D., Miller, C.A., Huang, I.C., Desai, P., Malone, L.M., Sweatt, J.D., 2006. Evidence that DNA (cytosine-5) methyltransferase regulates synaptic plasticity in the hippocampus. J. Biol. Chem. 281, 15763–15773. Levenson, J.M., Sweatt, J.D., 2005. Epigenetic mechanisms in memory formation. Nat. Rev. Neurosci. 6, 108–118. Levenson, J.M., Sweatt, J.D., 2006. Epigenetic mechanisms: a common theme in vertebrate and invertebrate memory formation. Cell. Mol. Life Sci. 63, 1009–1016. Linch, M.A., 2004. Long-term potentiation and memory. Physiol. Rev. 84, 87– 136. Lisman, J., Schulman, H., Cline, H., 2002. The molecular basis of CaMKII function in synaptic and behavioural memory. Nat. Rev. Neurosci. 3, 175– 190. Liu, S., Wang, J., Zhu, D., Fu, Y., Lukowiak, K., Lu, Y.M., 2003. Generation of functional inhibitory neurons in the adult rat hippocampus. J. Neurosci. 23, 732–736. Lledo, P.M., Alonso, M., Grubb, M.S., 2006. Adult neurogenesis and functional plasticity in neuronal circuits. Nat. Rev. Neurosci. 7, 179–193. Luria, A.R., 1968. The Mind of a Mnemonist. Basic Books, New York. Luria, A.R., 1976. The Neuropsychology of Memory. Winston, New York. Mactutus, C.F., Riccio, D.C., Ferek, J.M., 1979. Retrograde amnesia for old (reactivated) memory: some anomalous characteristics. Science 204, 1319– 1320. Madsen, T.M., Kristjansen, P.E., Bolwig, T.G., Wortwein, G., 2003. Arrested neuronal proliferation and impaired hippocampal function following fractionated brain irradiation in the adult rat. Neuroscience 119, 635–642. Malenka, R.C., Bear, M.F., 2004. LTP and LTD: an embarrassment of riches. Neuron 44, 5–21. Maletic-Savatic, M., Malinow, R., Svoboda, K., 1999. Rapid dendritic morphogenesis in CA1 hippocampal dendrites induced by synaptic activity. Science 283, 1923–1927. 112 Y.I. Arshavsky / Progress in Neurobiology 80 (2006) 99–113 Manns, J.R., Eichenbaum, H., 2006. Evolution of declarative memory. Hippocampus 16, 795–808. Maroun, M., Yaniv, D., Richter-Levin, G., 2001. Plasticity in local neuronal circuits. In vivo evidence from rat hippocampus and amygdala. In: Hölscher, C. (Ed.), Neuronal Mechanisms of Memory Formation. Cambridge University Press, Cambridge, MA, pp. 137–145. Martin, S.J., Grimwood, P.D., Morris, R.G., 2000. Synaptic plasticity and memory: an evaluation of the hypothesis. Annu. Rev. Neurosci. 23, 649– 711. McGaugh, J.L., 2000. Memory – a century of consolidation. Science 287, 248– 251. McGowan, P.O., Hope, T., Kelsoe, G.H., Meck, W.H., Williams, C.L., 2004. Recombination activating gene 1 (RAG1) may have a novel function in brain. In: Proceedings of the Soc. Neurosci. 34th Annual Meeting, 32221. Mel, B.W., 2002. Have we been hebbing down the wrong path? Neuron 34, 175–177. Milekic, M.H., Alberini, C.M., 2002. Temporally graded requirement for protein synthesis following memory reactivation. Neuron 36, 521–525. Millin, P.M., Moody, E.W., Riccio, D.C., 2001. Interpretations of retrograde amnesia: old problems redux. Nat. Rev. Neurosci. 2, 68–70. Milner, B., Squire, L.R., Kandel, E.R., 1998. Cognitive neuroscience and the study of memory. Neuron 20, 445–468. Ming, G.L., Song, H., 2005. Adult neurogenesis in the mammalian central nervous system. Annu. Rev. Neurosci. 28, 223–250. Misanin, J.R., Miller, R.R., Lewis, D.J., 1968. Retrograde amnesia produced by electroconvulsive shock after reactivation of a consolidated memory trace. Science 160, 554–555. Nader, K., 2003. Memory traces unbound. Trends Neurosci. 26, 65–72. Nader, K., Schafe, G.E., LeDoux, J.E., 2000. Fear memories require protein synthesis in the amygdala for reconsolidation after retrieval. Nature 406, 722–726. Nagerl, U.V., Eberhorn, N., Cambridge, S.B., Bonhoeffer, T., 2004. Bidirectional activity-dependent morphological plasticity in hippocampal neurons. Neuron 44, 759–767. Nakazawa, K., McHugh, T.J., Wilson, M.A., Tonegawa, S., 2004. NMDA receptors, place cells and hippocampal spatial memory. Nat. Rev. Neurosci. 5, 361–372. Nilsson, M., Perfilieva, E., Johansson, U., Orwar, O., Eriksson, P.S., 1999. Enriched environment increases neurogenesis in the adult rat dentate gyrus and improves spatial memory. J. Neurobiol. 39, 569–578. Nottebohm, F., 2002. Why are some neurons replaced in adult brain? J. Neurosci. 22, 624–628. Oddo, S., Caccamo, A., Shepherd, J.D., Murphy, M.P., Golde, T.E., Kayed, R., Metherate, R., Mattson, M.P., Akbari, Y., LaFerla, F.M., 2003. Tripletransgenic model of Alzheimer’s disease with plaques and tangles: intracellular Abeta and synaptic dysfunction. Neuron 39, 409–421. Parker, E.S., Cahill, L., McGaugh, J.L., 2006. A case of unusual autobiographical remembering. Neurocase 12, 35–49. Peña de Ortiz, S., Arshavsky, Y.I., 2001. DNA recombination as a possible mechanism in declarative memory: a hypothesis. J. Neurosci. Res. 63, 72– 81. Peña de Ortiz, S., Colón, M., Arshavsky, Y.I., 2004. Genomic theory of declarative memory. In: Parisi, V., De Fonzo, V., Aluffi-Pentini, F. (Eds.), Dynamical Genetics. Research Signpost, India, pp. 345–364. Peña de Ortiz, S., Colón, M., Carrasquillo, Y., Ren, K., Padilla, B., Silva, R., Arshavsky, Y.I., 2003. Experience-dependent expression of the gene encoding terminal deoxynucleotidyl transferase in the mouse brain. Neuroreport 14, 1141–1144. Penfield, W., 1958. Some mechanisms of consciousness discovered during electrical stimulation of the brain. Proc. Natl. Acad. Sci. U.S.A. 44, 51–66. Power, A.E., Berlau, D.J., McGaugh, J.L., Steward, O., 2006. Anisomycin infused into the hippocampus fails to block ‘‘reconsolidation’’ but impairs extinction: the role of re-exposure duration. Learn. Mem. 13, 27–34. Price, D.L., 2000. Aging of the brain and dementia of the Alzheimer type. In: Kandel, E.R., Schwartz, J.H., Jessel, T.M. (Eds.), Principles of Neural Science. 4th ed. McGraw-Hill, New York, pp. 1149–1161. Quartz, S.R., Sejnowski, T.J., 1997. The neural basis of cognitive development: a constructivist manifesto. Behav. Brain Sci. 20, 537–556. Rakic, P., 1985. Limits of neurogenesis in primates. Science 227, 1054–1056. Rakic, P., 2002. Neurogenesis in adult primate neocortex: an evaluation of the evidence. Nat. Rev. Neurosci. 3, 65–71. Ren, K., Peña de Ortiz, S., 2002. Nonhomologous DNA end joining in the mature rat brain. J. Neurochem. 80, 949–959. Roder, S., Danober, L., Pozza, M.F., Lingenhoehl, K., Wiederhold, K.H., Olpe, H.R., 2003. Electrophysiological studies on the hippocampus and prefrontal cortex assessing the effects of amyloidosis in amyloid precursor protein 23 transgenic mice. Neuroscience 120, 705–720. Rosenbaum, R.S., Kohler, S., Schacter, D.L., Moscovitch, M., Westmacott, R., Black, S.E., Gao, F., Tulving, E., 2005. The case of K.C.: contributions of a memory-impaired person to memory theory. Neuropsychologia 43, 989–1021. Ross, R.S., Eichenbaum, H., 2006. Dynamics of hippocampal and cortical activation during consolidation of a nonspatial memory. J. Neurosci. 26, 4852–4859. Routtenberg, A., Rekart, J.L., 2005. Post-translational protein modification as the substrate for long-lasting memory. Trends Neurosci. 28, 12–19. Sadowski, M., Pankiewicz, J., Scholtzova, H., Li, Y.S., Quartermain, D., Duff, K., Wisniewski, T., 2004. Links between the pathology of Alzheimer’s disease and vascular dementia. Neurochem. Res. 29, 1257–1266. Sara, S.J., 2000. Retrieval and reconsolidation: toward a neurobiology of remembering. Learn. Mem. 7, 73–84. Schinder, A.F., Gage, F.H., 2004. A hypothesis about the role of adult neurogenesis in hippocampal function. Physiology (Bethesda) 19, 253–261. Scoville, W.B., Milner, B., 1957. Loss of recent memory after bilateral hippocampal lesions. J. Neurol. Neurosurg. Psychiatry 20, 11–21. Selkoe, D.J., 2001. Alzheimer’s disease: genes, proteins, and therapy. Physiol. Rev. 81, 741–766. Selkoe, D.J., 2002. Alzheimer’s disease is a synaptic failure. Science 298, 789– 791. Shors, T.J., Miesegaes, G., Beylin, A., Zhao, M., Rydel, T., Gould, E., 2001. Neurogenesis in the adult is involved in the formation of trace memories. Nature 410, 372–376. Si, K., Giustetto, M., Etkin, A., Hsu, R., Janisiewicz, A.M., Miniaci, M.C., Kim, J.H., Zhu, H., Kandel, E.R., 2003a. A neuronal isoform of CPEB regulates local protein synthesis and stabilizes synapse-specific long-term facilitation in Aplysia. Cell 115, 893–904. Si, K., Lindquist, S., Kandel, E.R., 2003b. A neuronal isoform of the aplysia CPEB has prion-like properties. Cell 115, 879–891. Silva, A.J., 2003. Molecular and cellular cognitive studies of the role of synaptic plasticity in memory. J. Neurobiol. 54, 224–237. Small, D.H., Mok, S.S., Bornstein, J.C., 2001. Alzheimer’s disease and Abeta toxicity: from top to bottom. Nat. Rev. Neurosci. 2, 595–598. Snyder, J.S., Hong, N.S., McDonald, R.J., Wojtowicz, J.M., 2005. A role for adult neurogenesis in spatial long-term memory. Neuroscience 130, 843–852. Squire, L.R., 1992. Memory and the hippocampus: a synthesis from findings with rats, monkeys, and humans. Psychol. Rev. 99, 195–231. Squire, L.R., Clark, R.E., Knowlton, B.J., 2001. Retrograde amnesia. Hippocampus 11, 50–55. Squire, L.R., Kandel, E.R., 1999. Memory: From Mind to Molecules. Sci. Am. Library, New York. Squire, L.R., Slater, P.C., Chace, P.M., 1976. Reactivation of recent or remote memory before electroconvulsive therapy does not produce retrograde amnesia. Behav. Biol. 18, 335–343. Sweatt, J.D., 2003. Mechanisms of Memory. Elsevier, Amsterdam. Tashiro, A., Sandler, V.M., Toni, N., Zhao, C., Gage, F.H., 2006. NMDAreceptor-mediated, cell-specific integration of new neurons in adult dentate gyrus. Nature 442, 929–933. Taubenfeld, S.M., Milekic, M.H., Monti, B., Alberini, C.M., 2001. The consolidation of new but not reactivated memory requires hippocampal C/ EBPbeta. Nat. Neurosci. 4, 813–818. Teixeira, C.M., Pomedli, S.R., Maei, H.R., Kee, N., Frankland, P.W., 2006. Involvement of the anterior cingulate cortex in the expression of remote spatial memory. J. Neurosci. 26, 7555–7564. Terry, R.D., Masliah, E., Salmon, D.P., Butters, N., DeTeresa, R., Hill, R., Hansen, L.A., Katzman, R., 1991. Physical basis of cognitive alterations in Alzheimer’s disease: synapse loss is the major correlate of cognitive impairment. Ann. Neurol. 30, 572–580. Y.I. Arshavsky / Progress in Neurobiology 80 (2006) 99–113 Teskey, G.C., Valentine, P.A., 1998. Post-activation potentiation in the neocortex of awake freely moving rats. Neurosci. Biobehav. Rev. 22, 195–207. Thomas, G.M., Huganir, R.L., 2004. MAPK cascade signaling and synaptic plasticity. Nat. Rev. Neurosci. 5, 173–183. Thompson, P.J., 1991. Memory function in patients with epilepsy. Adv. Neurol. 55, 369–384. Toni, N., Buchs, P.A., Nikonenko, I., Bron, C.R., Muller, D., 1999. LTP promotes formation of multiple spine synapses between a single axon terminal and a dendrite. Nature 402, 421–425. Treffert, D.A., Christensen, D.D., 2005. Inside the mind of a savant. Sci. Am. 293, 108–113. Trinchese, F., Liu, S., Battaglia, F., Walter, S., Mathews, P.M., Arancio, O., 2004. Progressive age-related development of Alzheimer-like pathology in APP/PS1 mice. Ann. Neurol. 55, 801–814. Ujiie, M., Dickstein, D.L., Carlow, D.A., Jefferies, W.A., 2003. Blood–brain barrier permeability precedes senile plaque formation in an Alzheimer disease model. Microcirculation 10, 463–470. van Praag, H., Christie, B.R., Sejnowski, T.J., Gage, F.H., 1999. Running enhances neurogenesis, learning, and long-term potentiation in mice. Proc. Natl. Acad. Sci. U.S.A. 96, 13427–13431. Walsh, D.M., Selkoe, D.J., 2004. Deciphering the molecular basis of memory failure in Alzheimer’s disease. Neuron 44, 181–193. 113 Wang, H., Hu, Y., Tsen, J.Z., 2006. Molecular and systems mechanisms of memory consolidation and storage. Prog. Neurobiol. 79, 123–135. Wang, J., Ren, K., Pérez, J., Silva, A.J., Peña de Ortiz, S., 2003. The antimetabolite ara-CTP blocks long-term memory of conditioned taste aversion. Learn. Mem. 10, 503–510. Wiltgen, B.J., Brown, R.A., Talton, L.E., Silva, A.J., 2004. New circuits for old memories: the role of the neocortex in consolidation. Neuron 44, 101–108. Wittenberg, G.M., Tsien, J.Z., 2002. An emerging molecular and cellular framework for memory processing by the hippocampus. Trends Neurosci. 25, 501–505. Yi, J.J., Ehlers, M.D., 2005. Ubiquitin and protein turnover in synapse function. Neuron 47, 629–632. Zipp, F., Aktas, O., 2006. The brain as a target of inflammation: common pathways link inflammatory and neurodegenerative diseases. Trends Neurosci. 29, 518–527. Zlokovic, B.V., 2005. Neurovascular mechanisms of Alzheimer’s neurodegeneration. Trends Neurosci. 28, 202–208. Zola-Morgan, S.M., Squire, L.R., 1990. The primate hippocampal formation: evidence for a time-limited role in memory storage. Science 250, 288–290. Zuo, Y., Lin, A., Chang, P., Gan, W.B., 2005. Development of long-term dendritic spine stability in diverse regions of cerebral cortex. Neuron 46, 181–189.