Survey
* Your assessment is very important for improving the work of artificial intelligence, which forms the content of this project
Coronary artery disease wikipedia , lookup
Artificial heart valve wikipedia , lookup
Antihypertensive drug wikipedia , lookup
Electrocardiography wikipedia , lookup
Heart failure wikipedia , lookup
Hypertrophic cardiomyopathy wikipedia , lookup
Mitral insufficiency wikipedia , lookup
Myocardial infarction wikipedia , lookup
Quantium Medical Cardiac Output wikipedia , lookup
Cardiac surgery wikipedia , lookup
Lutembacher's syndrome wikipedia , lookup
Arrhythmogenic right ventricular dysplasia wikipedia , lookup
Atrial septal defect wikipedia , lookup
Dextro-Transposition of the great arteries wikipedia , lookup
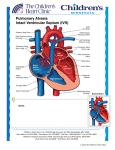
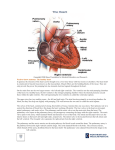
REPTILE CARDIOLOGY Schilliger, LH, DVM, Dip. ECZM (Herpetology)a a Clinique Vétérinaire du Village d’Auteuil, 35 rue Leconte de Lisle, 75016 Paris, France. ABSTRACT Reptilian cardiology differs markedly from that of mammal cardiology, both in terms of the anatomy and physiology, but also in terms of the diversity displayed across the different taxa. Thus, reptilian hearts can be loosely classified as being crocodilian hearts (crocodiles, alligators, gavials and caimans) or non-crocodilian (snakes, lizards and chelonians). In noncrocodilian reptiles, the ventricle is not physically divided but pulmonary and systemic blood flow are separated and regulated by functional plasticity. In these species, oxygenated and deoxygenated blood flowing from the aortic arches is partially mixed, the amount of mixing being determined by the degree of muscular ridge and interventricular septum development. The contraction of the muscular ridge during systole and of the interventricular septum during diastole serves to form an almost complete separation of pulmonary and systemic blood flow, particularly in monitor lizards and pythons. In cases of anoxia (e.g. prolonged diving, swallowing large preys, respiratory distress or hibernation), vasoconstriction of the pulmonary arteries/artery causes a high pulmonary outflow resistance. Together with a decreased pressure in the pulmonary veins, a right to left intracardiac shunt occurs and blood from the right atrium shifts towards both aortic arches, instead of the pulmonary trunk. This ensures bypass of the lungs and blood perfusion to vital organs during anoxia. The cardiac anatomy and physiology in crocodilian species differs from non-crocodilian reptiles but resembles that of mammals. Indeed, the ventricles are physically divided by an intraventricular muscular septum with divided pulmonary and systemic circulation. During apnea however, the crocodilian heart is able to function in a manner similar to that of reptiles with a right to left shunt. This unique versatility is made possible by the foramen of Panizza, an interventricular septal opening that connects the left and right aortic trunks, which, coupled with an active control of pressure gradients in the ventricles, allows for mixing of oxygenated and deoxygenated blood. Whilst the degree of shunting is known for many species, the functional mechanisms and physiologic role of cardiac shunting have been discussed extensively with no final or conclusive interpretations. Poïkilotherms have long been considered to suffer little from primary cardiovascular conditions, however this idea is being challenged as our knowledge of normal anatomy and physiology improves and the use of specialist diagnostic tools become more available. These include instruments readily available in small animal medicine (auscultation, electrocardiography, radiography, ultrasound, CT) applied with minor variations in techniques. INTRODUCTION In spite of our relatively good understanding of reptilian cardiac anatomy and physiology, reptile cardiology remains a poorly developed sub-specialty in herpetological medicine. Our understanding of the etiology, diagnosis and treatment of cardiac conditions in reptiles remains poor and more effort should be put into translating our present knowledge into clinical application. I. CARDIAC ANATOMY Reptilian cardiology differs markedly from that of mammal cardiology, both in terms of the anatomy and physiology but also in terms of the diversity displayed across the different taxa. Thus, reptilian hearts can be loosely classified as being crocodilian (crocodiles, alligators, gavials and caimans) or non-crocodilian (snakes, lizards and chelonians). Most published descriptions that have been performed on specific species (mainly monitor lizards, pythons, crocodilians and freshwater turtles) show very variable designs in anatomy and physiology and these are sometimes subject to controversy. Little is known about the cardiac anatomy and physiology of the other 9000+ species of reptiles.3,5,24,25,60,61-63,a In non-crocodilian reptiles, the ventricles are not physically divided, but pulmonary and systemic blood flow are separated and regulated by functional plasticity. In crocodilians however, the ventricles are fully divided by a muscular interventricular septum. They are the only reptilian species to possess an anatomically four-chambered heart.6,60-62,a,b,c A. TOPOGRAPHICAL ANATOMY The location of the heart varies within reptilian taxa and reflects their phylogenetic position and ecological niche. In most species, the heart is roughly positioned along the axial midline.30,a In snakes, the heart is found along the midline axis and its longitudinal position varies with species. Thus, in marine and freshwater snakes, it is positioned near the middle of the body (25 to 45% of total body length). In non-tree dwelling snakes it is found at about 25% of total body length, and about 15% (i.e. more cranially) in arboreal species.6,8,30,32,64,a,b,c These variations in heart positions can be understood when considering the different physical forces to which the heart is subjected to in snakes occupying different biotypes. A cranially positioned heart reduces the hydrostatic pressure above the heart and so stabilizes variations in cephalic blood pressure.6,8,a,b,c Some studies have shown that those species of snakes that spend time raising their heads (arboreal and other climbing snakes) possess relatively shorter vasculature between the heart and head than do terrestrial species. The reverse is true in aquatic species, which experience less effects of gravity in water (Lillywhite et al, 2011-In Press). Because of the lack of diaphragm, the snake’s heart is mobile within the coelomic cavity, which probably facilitates the movement of large whole prey in the esophagus5. In the coelom, the heart is found adjacent to the caudal tracheal rings, caudal to the thyroid, cranial to the bronchial bifurcation, close to the cranial pole of the lung(s) and slightly cranial to the liver.6,8,32,61,63,64 Externally, the position of the heart is indicated by percutaneous visualization of the ventral precordial tap by placing the animal in dorsal recumbency and stretching it.32,b The chelonian heart is located in the ventral midline at the level of the intersection of the humeral, pectoral and abdominal scutes of the plastron. In some species it is positioned more caudally, between the junction of the humeral-pectoral and pectoral-abdominal scute lines. This may however be an unreliable landmark as the position of the pectoral and humeral scutes differ widely amongst species.64,a In the coelom, the heart is bordered dorsally by the lungs, laterally by the liver lobes and ventrally by the acromion and coracoid processes.8 In soft-shelled turtles of the Tryonychidae family, which possess a very streamlined carapace and hence small coelomic cavity, the heart is displaced to the right.64 This is due to long protractile neck, which when retracted into the carapace causes a displacement of the heart and liver to the right and of the stomach to the left.6 In most species of lizards, the heart is encased in the pectoral girdle, in the gular region. however in Tegus, Gila monsters and monitor lizards, the heart is located more caudally in the coelomic cavity, behind the cranial part of the sternum.6,8,32,a,b In crocodilians, the heart lies in the body midline.6 The pericardial sac occupies the caudal ventral part of the thoracic cavity. The apex lies deep between the right and left medial aspect of the hepatic lobes and apposes the liver isthmus. The base of the heart extends slightly cranial to the cranial border of the liver and lies within the mediastinum, ventral to the primary bronchi and esophagus.34 B. EXTERNAL GROSS MORPHOLOGY Despite minor variations, the external anatomy of the reptilian heart tends not to vary significantly between species. It’s interior however is complex and species specific.64 The heart represents approximately 0.2% to 0.3% of total body weight, however this varies with the effects of gravity and physical activity.6,64,a,b The non-crocodilian heart has historically been described as a three-chambered organ. This can be appreciated when examining the ventral view with the two muscular atria and single ventricle. Because the sinus venosus (see below) is now considered as an additional chamber, the non-crocodilian heart is considered as an atypical four-chambered heart. However because there is no interventricular septum proper, but rather incomplete muscular ridges that divide the ventricle, the non-crocodilian ventricle is morphogically regarded as a single chamber. This contrasts with the four-chambered crocodilian heart, where the presence of a complete interventricular septum anatomically and functionally separates the ventricle into two chambers. Nevertheless, these differences are hardly visible externally.6,63 As in all vertebrates, the reptilian heart is completely covered by a white fibrous pericardial sac and lies within a cœlomic space, the pericardial cavity. In freshwater turtles, it has been shown that the pericardial sac is filled with an alkaline fluid, rich in calcium and magnesium, which could play a protective role from acidosis during apnea.16 Within the pericardial sac, the heart is covered by the epicardium, which forms the serous lining of the pericardial cavity.6 1. Non-crocodilians In ophidians, the heart is elongated, especially in colubrid snakes, and matches the general appearance of the body.6,63,32 Unlike other reptiles (tuataras, crocodilians and most chelonians and lizards) in which the heart apex is attached to the pericardium by a ligament termed the gubernaculum cordis, the extremity of a snake’s ventricle remains free within the pericardial space, as is also the case in varanids.6 In chelonians and in varanids, the width of the heart is almost the same as its length.6,a,b Seen from the outside, the heart of non crocodilian reptiles is composed of: - Four main cavities: a single ventricle (similar to an inverted cone, it is broad and flattened dorsoventrally in many Testudines or elongated and oval in snakes, with rounded edges), two atrial chambers which can be seen on a ventral view (the right atrium being, especially in snakes and to a lesser extent in Testudines, markedly more developed and more muscular than the left one) and a sinus venosus, in which cardiac contraction is initiated. This thin sac-like structure of tubular or triangular shape, which is less developed in snakes than in lizards, is only visible from a dorsal view of the heart, and rests on the dorsal aspect of the right atrium, at the confluence of the three caval veins in most reptiles: these are the right precaval vein, the left precaval vein and the postcaval vein.6,25,52,60,61,a,b,c In the reticulated python (Python reticulatus), it seems to continue on from the postcaval vein, which is not the case, for example, in the boa constrictor (Boa constrictor) or the anaconda (Eunectes murinus).6 In chelonians (Testudines), the sinus venosus is formed at the confluence of the three caval veins and the hepatic vein.6 In the south American rattlesnake (Crotalus durissus), it is formed at the confluence of the postcaval vein (the vena cava posterior), the precaval vein (the vena cava anterior) and the jugular vein, which is much smaller than the two former ones.24 In snakes and lizards, the sinus venosus - - - is partially divided by an incomplete septum (a kind of constriction between the terminal portion of the left precaval vein and the confluence of the right precaval vein and the postcaval vein). It is undivided in tortoises and in the Tuatara (Sphenodon sp.).13,25 The ventral wall of the sinus venosus is attached to the dorsal wall of the ventricle by the dorsal ligament. All the walls of the sinus venosus are thinner than those of the other cardiac cavities.6 The atria are separated from the ventricle by quite a deep oblique-angled sulcus, called the coronary groove (Coronary Sulcus).6 This sulcus is not horizontal, because of the asymetry of the atria. In certain viperid snakes, e.g. Agkistrodon contortrix mokeson, the right atrium covers the cranial third of the ventricle.6 The atrial walls are better developed in testudines compared to other species of reptiles. In varanids, the two atria are of equal size.6,a,b,c Three arterial trunks, running from the ventricle between the two atria, rotating towards the right, and forming an angle of 180°, typical of reptiles. They are termed the left aortic arch, right aortic arch and pulmonary trunk.6,32,39,52,63,64,a,b,c The two aortic arches merge together caudally to form the common abdominal aorta (dorsal aorta).6,63,64 The right aortic arch coming from the left side of the ventricle develops into either one or two common carotid arteries. The pulmonary trunk divides up into the left and right pulmonary arteries in ophidians with two functional lungs (e.g. boid snakes). In others, such as colubrids, viperids and elapidids, the pulmonary trunk develops into one anterior pulmonary artery pointing forwards and a posterior pulmonary artery pointing backwards.6 Four afferent veins: the pulmonary vein (which, in bi-lunged snakes, proceeds from the point of confluence between the right pulmonary vein and the left pulmonary vein), the two precaval veins and the post-caval vein, draining into the sinus venosus.6,33,39,52,63,64 A well-developed coronary vascularization that arises from the systemic carotid trunk and supplies the compact myocardium, the sinus venosus and the two atria. In some reptiles (apparently especially in turtles and alligators), the supply of oxygen in the heart muscle is also provided by the blood contained in the cardiac cavities (termed transmyocardial blood perfusion, which may account for up to 30% of coronary perfusion in these species).6,24 2. Crocodilians The heart of crocodilians has two atria and a ventricle that is physically divided by a distinct muscular septum into right (pulmonary + systemic) and left (strictly systemic) sides. It is visibly externally as the interventricular sulcus, which marks the junction between right and left ventricles. On gross examination, the heart is comparable that of mammals and birds, except for the presence of the left and right aortic arches. The gubernaculums cordis extends from the right ventricle (not from the apex as in Testudinae and Sauria) and anchors the heart to the pericardium. The right and left atria are of similar size. The sinus venosus resembles an elongated pyramid, with the base lying cranially and apex caudally. In the saltwater crocodile (Crocodylus porosus), the sinus venosus is formed is formed by the union of the hepatic vein, postcaval vein and right precaval vein. This latter vein is separated from the sinus venosum by a dorsal sinus septum.6,62 Three major vessels exit the ventricle: the right aorta leaves the left ventricle and gives off cranially the carotid and subclavian arteries. It continues caudally as the dorsal aorta. The pulmonary artery leaves the right ventricle and supplies the left and right lungs. The left aorta also leaves the right ventricle. As they exit the ventricle, all three vessels are of similar diameter. As they run cranially, the aortae and the pulmonary artery are bound together by a connective tissue sheath.6,62,64,a C. INNER STRUCTURES The cavities of the heart are lined by the endocardium, a membrane continuous with blood vessel endothelium. The outer heart is covered by the epicardium, and between these two layers is the myocardium, which forms the bulk of the heart mass. It comprises of specialized muscle and connective tissue cells. The muscle cells are arranged in three layers, which are, from the exterior to the interior: a thin layer of longitudinally arranged myocytes, followed by spirally arranged myocytes (poorly arranged in young animals and Tuataras) and finally by the spongy myocardium that form the traberculae (better developed in species with round and short ventricles).64 1. Non-crocodilians Seen from the inside, the heart is composed of: - Three ventricular sub-chambers, recessed within the ventricular cavity and communicating with each other: the cavum venosum (right dorsal chamber of the ventricle), the cavum arteriosum (left dorsal chamber of the ventricle) and the cavum pulmonale (left chamber or “ ventral ventricle ”).6,33,39,52,60-64 Reptiles belonging to the genus Varanus possess an enlarged cavum arteriosum and reduced cavum venosum. This gives the heart a symmetrical appearance, in contrast to other squamates.6 - Several openings and valves: the pulmonary vein drains into the dorsal wall of the left atrium through the valve-free pulmonary vein orifice, located close to the interatrial septum. The sinus venosus proceeds into the right atrium at the sinoatrial opening. This orifice has a pair of flap valves derived from the endocardium that are membranous or muscular, depending upon the species, and are known as sinoatrial valves.6,53 The atrial chambers communicate with the single ventricle through the left and right atrioventricular orifices. These orifices are both equipped with a single atrioventricular valve, known as the single septal monocuspid atrioventricular valve.6,33,39,52,60-64 The left and right aortic arches arise from the cavum venosum at two orifices with bicuspid valves, rather than tricuspid valves as in mammals. The pulmonary trunk is a continuation of the cavum pulmonale and its base also contains small semilunar valves.6 In some species of turtles and squamates, a foramen connects the left and right aortas. The functional significance is unknown as both aortas originate from the cavum venosum.53 - two main septa: one vertical septum, so called “interventricular septum”, located in the ventricular cavity, between the cavum arteriosum and the cavum venosum. Because of its position facing the interatrial septum, it contributes to the separation of pulmonary veinous blood from systemic flow during ventricular diastole. A second relatively strong muscular ridge, the muscular ridge (also called horizontal septum), originates between the pulmonary trunk and the left aortic arch, and twists towards the dorso-apical region of the ventricle. This ridge marks a separation between the cavum venosum and the cavum pulmonale. It is more developed in varanids and pythons compared to other squamates and most chelonians (with the exception of sea turtles and giant tortoises).6,33,39,52,60-64 - another septum, the bulbuslamelle, which lies opposite to the muscular ridge, curving over and around it. It is situated immediately to the right of the vertical septum.24 2. Crocodilians Seen from the inside, the heart is composed of: - - a left and right ventricle, separated by an interventricular septum. The right ventricle is further separated into a ventral and lateral chamber by a ventral branch of the muscular septum (the aorticopulmonary septum). It separates the pulmonary artery from the left systemic one (the left aorta). Because the vertical (interventricular) septum in non-crocodilians lies approximately opposite to the caudal edge of the interatrial septum, it is analogous to the muscular part of the ventricular septum in crocodilians.6,8,32,62,64,a several openings and valves: the base of each efferent arteries has a bicuspid valve, the medial cusp being longer than the lateral one. Significantly, the medial cusp of the right aorta covers the foramen of Panizza during ventricular systole. The bicuspid atrioventricular valves, which guard the connection between the atria and ventricles, are composed of a left septal and a right marginal cusp (they are bicuspid valves). The left atrioventricular valves are membranous as the marginal cusp of the right atrioventricular valve is a thick, lip-like structure. As in mammals, the pulmonary veins are devoid of valves. The right and left aorta are connected via the foramen of Panizza and the dorsal connecting artery. The former structure lies just above the bicuspid valves at the base of both aortas. The dorsal connecting artery is located more distally, but is rostral to the convergence of both aorta into the single aorta.6,8,32,62,64,a II. PHYSIOLOGY A. Conduction and heart rate Unlike mammals, reptiles do not have a specialized cardiac conduction system such as pacemaker nodes and Purkinje fibers. Instead, the contractions are initiated by cardiac muscle fibers in the sinus venosus of the right atrium, which spread in sequential coordination, first to the left and caudally.6,64 The ventricle is depolarized starting at its base and then proceeding to the left. Repolarization starts from the base and spreads equally to the right and left towards the apex.33,b The heart receives innervations from both parasympathetic and sympathetic fibers. The parasympathetic fibers run in the vagus nerve and provide cholinergic (inhibitory) control. The less well-developed sympathetic fibers cause positive chronotropism via adrenergic innervation.6,58,61,64 Heart rates (HR) are in general slower in reptiles compared to mammals or birds. Heart rate is dependant on numerous factors: body temperature (increasing during basking and lowering during cooling, myocardial efficiency being optimum when the reptile is within its preferred optimal temperature zone), body size (heart rate is inversely proportional to the animal’s size), activity (heart rate is proportional to metabolic level), respiratory rate (bradycardia is observed during apnea as pulmonary resistance increases and blood flow to the lungs decreases), hypovolemia (reptiles that experience blood loss as a result of surgery or trauma can become tachycardic to ensure that the tissues remain oxygenated), digestion, gravidity and sensory stimulations such as handling, postural and gravitational stress.6,32,b Heart rate also plays a significant role in thermoregulation: it increases when the animal lies in the sun, and vice versa. Thus, tachycardia and peripheal vasodilatation increase heat loss from cutaneous vascularization when ambient temperature increases, whereas bradycardia and peripheal vasoconstriction decrease heat loss when ambient temperature drops. This mechanism, which often anticipates internal temperature variations, is regulated by cutaneous thermoreceptors.6 Heart rate (HR) can be predicted using the following formula32,d : HR (beats per minute) = 33,4 x W(kg)-0,25 B. Intracardiac blood flow during normal breathing 1. Non-crocodilians (three chambered hearts) During atrial systole, both atrioventricular valves open simultaneously, blocking the interventricular canal between the cavum arteriosum and the cavum venosum. The deoxygenated blood contained in the right atrium first fills the cavum venosum and the cavum pulmonale, incompletely separated by the muscular ridge, and the oxygenated blood contained in the left atrium then flows into the cavum arteriosum.6,32,39,52,64 Blood from the right atrium enters the cavum venosum slightly before blood from the left atrium fills the cavum arteriosum.53 Thus, during ventricular diastole, systemic and pulmonary blood flow are partially separated by the interventricular septum. During ventricular systole, both atrioventricular valves close the atrioventricular funnels and open the interventricular canal, allowing blood from the cavum arteriosum to fill the cavum venosum (once the cavum pulmonale is filled). The muscular ridge moves cranially and separates the cavum venosum and the cavum pulmonale. Oxygenated blood is then pumped out of the cavum venosum on to the left and right aortic arches and deoxygenated blood is pumped out from the cavum pulmonale towards the pulmonary arterial trunk.6,32,53,64 The diastolic blood pressure is lower in the pulmonary arches than in the systemic ones, hence blood flows in the pulmonary arties before the systemic arteries.6 Ventricular systole tends to be long and diastole is short.64 No outflow vessel emerge from the cavum arteriosum (the two aortic arches originate exclusively from the cavum venosum) and no inflow vessel drain into the cavum pulmonale (this sub-chamber receives blood only from the the cavum venosum).53 Hence, the cavum venosum has direct systemic arterial outputs as well as a direct right atrial input of systemic venous blood.6,53 In non-crocodilian reptiles, blood flowing along the two aortic arches is mixed because the cavum venosum contains both oxygenated and deoxygenated blood. However, this mixture is dependant on the degree of interventricular septum and muscular ridge development. The contraction of the interventricular septum during diastole and of the muscular ridge during systole can ensure an almost complete separation of the pulmonary and systemic circulation, especially in pythons and varanids. In these species, the cavum pulmonale becomes functionally separated from the cavum venosum and the cavum arteriosum during systole.6,13,24,32,53,59,60-65 In most reptiles (most of snakes, lizards and chelonians), blood pressures in the pulmonary and systemic arteries are relatively low and are identical during systole. However, in pythons and varanid lizards, in which the asymetry in wall thickness and the muscular ridge create a pressure differential between the right and the left sides, systemic blood pressure is much larger than pulmonary blood pressure (as is the case in mammals and birds).13,25,53,59 One study showed that ball pythons (Python regius) achieve a complete separation of the pulmonary and systemic circulation.58,59 This dual pressure pump system has also been demonstrated in varanids.3,31 2. Crocodilians (four chambered hearts) When the crocodilian has free access to air, the heart functions in a similar way to mammals with separate systemic and pulmonary circulations. Because the blood pressure in pulmonary vasculature is much lower than in the systemic vasculature, the pressure generated in the right ventricle is much lower than in the left ventricle. The blood pressures in the left and right aorta are equalized via the foramen of Panizza (which opens during ventricular diastole) and the dorsal connecting artery. Consequently, the bicuspid valve at the orifice of the left aorta (remember that this exits the right ventricle) remains closed, as the right ventricular pressure does not exceed left aortic blood pressure. Hence, all the blood in the right ventricle is pumped into the pulmonary circulation. In addition, the blood in the left ventricle goes primarly into the right aorta as one of the valves guarding the right aorta folds back to cover the entire foramen of Panizza during systole.6,8,32,62,64,a,b,c C. Intracardiac blood flow during anoxia (ex: apnea) 1. Non-crocodilians Non-crocodilian reptiles exhibit an interesting distinctive cardio-circulatory feature in that intracardiac blood flow may vary according to systemic blood oxygenation levels (PO2 levels) and to the relative vascular resistance of the pulmonary and systemic circuits. In cases of anoxia (prolonged underwater diving, slowly swallowing large preys, hibernation or respiratory distress), an increase in pressure of the pulmonary artery (due to vasoconstriction) and a decreased blood pressure in the pulmonary veins are observed, creating a high pulmonary outflow resistance. Subsequently, a right to left intracardiac shunt may occur and blood from the right atrium shifts towards both aortic arches, instead of the pulmonary trunk. This shunt ensures blood perfusion of vital organs during anoxia.13,25,64 This pulmonary bypass has also been shown in fasting and resting ball pythons (Python regius) where up to twice as much blood flows into the systemic circulation than into the pulmonary one.53 Inversely, a left to right shunt occurs in case of low systemic blood oxygen levels, when pulmonary resistance is low or when a physiological or environmental event (such as a variation in temperature, exercise or digestion) increases metabolism rate.13,25,64 This L-R shunting (directing more blood into the pulmonary circulation) has been clearly shown in digesting ball pythons (Python regius), whose oxygen demand increases considerably during digestion.53 It is thought that this shunting is only possible only during the early phase of ventricular systole, before blood flows from the cavum arteriosum into the cavum venosum and before the muscular ridge closes the connection between the cavum venosum and cavum pulmonale.53 The ability of cardiac shunting is species dependant. For example, the poorly subdivided ventricle of freshwater turtles enables large cardiac shunts whereas pythons and monitor lizards generate blood pressures similar to mammals thanks to their atypically « divided » heart (the muscular ridge separates pulmonary and systemic circuits during ventricular systole and the cavum venosum is much reduced in pythons, approximately 10% of the cavum arteriosum).6,25 In these reptiles, shunts are less frequent than in other species (20% compared to more than 50%).25 Whilst the degree of shunting is known for many species, the functional mechanisms and physiologic role of cardiac shunting have been discussed extensively with no final or conclusive interpretations. 2. Crocodilians During apnea, the crocodilian heart functions similarly to a non-crocodilian heart with a right to left shunt, and up to 25% of blood flow may bypass the lungs. Dual mode of action is made possible by the foramen of Panizza, together with an active control of pulmonary vasculature outflow. When in apnea, an increase pressure of lung tissue together with coglike valves in the pulmonary artery causes an increase in pulmonary resistance. These results in the pressure of the right ventricle overcoming that of the pressure in the left systemic aorta. Then, there can be simultaneous output from the right ventricle into the left aorta and the pulmonary artery. Most of the blood probably follows the course of the left aorta. However, some blood could pass into the right aorta via the foramen of Panizza and at the confluence of the left and right aortic arches (resulting in some mixing of oxygenated and deoxygenated blood in the systemic circulation).6,8,32,62,64,a,b,c III. CARDIOVASCULAR DISEASES IN REPTILES Clinical signs of cardiac disease in reptiles include swelling in the area of the heart, cyanosis, peripheral oedema, ascites, and exercise intolerance. Non-specific clinical signs are however more frequent, such as lethargy, depression, anorexia, weight loss, weakness, dyspnoea and sudden death. Poïkilotherms have long been considered to suffer little from primary cardiovascular conditions, nevertheless, the reptilian heart can be subject to various lesions such as cardiomyopathy2,e, septic endocarditis20,48,f, valvular insufficiency39,46,f, myocarditis12,18,21,36,43,22,f,g, pericardial effusion38,45, infarct41, atherosclerosis45, aneurysms41,h, Gout17,27, arteries calcification49,e, thrombus20, parasitic infestation5,7,9,10,26,29, congenital heart defects23,41,i, and even tumors14,28,47,50,j In many cases, the diagnosis is obtained post mortem. When case of suspected heart disease first presents, the clinician should first of all determine if the cause of the disease is primary or secondary to systemic illness. IV. DIAGNOSTICS TOOLS Diagnostic tools commonly used in reptile cardiology include most instruments typically used in small animal medicine but applied with minor variations in techniques. Hence, auscultation, electrocardiography, radiography, ultrasound and CT are the most used tools for investigating cardiac disease in reptiles.33,b,k-r It is important to note that when performing a clinical examination, care should be taken to consider the environmental temperature as this can influence cardiopulmonary parameters and distort the final results.64,a,b,k A. Auscultation Heart sounds in reptiles are of very low amplitude and hence cannot be consistently auscultated with a standard stethoscope. This problem can be overcome in some species by using a pressure sensitive acoustic stethoscope (Ultrascope) that permits diagnosis of heart murmurs. This equipment is however only suitable in large species of snakes (e.g. large boids) and species of lizards where the heart lies in the mid-coelomic region and is not hidden under the bony pectoral girdle, as for example in monitor lizards. Doppler flow ultrasound, used to determine blood pressures in small mammals, can be used for cardiac auscultation in all species of reptiles. The ultrasound emitter is placed with an acoustic gel coating on the epidermal surface at the level of the heart or large efferent arteries. The sounds obtained represent the systolic blood flow and enables the clinician to evaluate the heart rate and rhythm.33,a,b,k B. Electrocardiography Electrocardiography (ECG) can greatly enhance the diagnosis of cardiac disease in reptiles and is also beneficial for monitoring patients under anesthesia. The main challenge associated with the use of ECG in reptiles are the low electric amplitudes (usually <1.0mV) which do not always provide readings of diagnostic quality. Moreover, standard parameters are not established for many species, thus interval and segments values may not always be of much use. Performing routine ECGs on healthy patients can help to build up baseline values for future comparison.33,55-57a,b,k Interpretations of ECGs in reptiles are very similar to those in mammals, with P, QRS and T complexes. An SV wave represented by the depolarization of the sinus venosus (and the postcaval vein) may be measured just before the P wave.The SV wave is followed by sinal contraction, the P wave is followed by atrial contraction and the R wave is followed by ventricular contraction. The T wave indicates ventricular repolarization.33,55-57a,b,k It should be noted that correct placement of the probe is primordial as bad positioning of the electrodes can influence ECG readings. The electrodes can be self-adhering skin electrodes (designed for human use), stainless steel hypodermic needles, stainless steel suture material or alligator clips. Placement of the electrodes varies according to species, but is usually inspired from the traditional four limb lead placement. In snakes they should be placed two heart lengths cranial and caudal to the heart. In lizards where the heart is hidden under the pectoral girdle (e.g. iguanas, bearded dragons, chameleons, water dragons and skinks), the two cranial electrodes should be placed in the cervical region instead of on the forelimbs. Lizards such monitors and Tegus, as well as crocodilians, which have a heart positioned caudally to the pectoral girdle, are suited for the electrodes being placed on limbs or torso. The four limb placement is however not appropriate in chelonians due to the low surface voltage generated. In these species, the two cranial electrodes should be placed in the cervical region, lateral to the neck and medial to the forelimbs.33,55-57a,b,k C. Radiography The use of diagnostic imaging is an essential part of the examination of the reptile patient. Although conventional radiography has historically been the main imaging tool for reptiles, it remains challenging to deliver images of high diagnostic value, especially for assessing the cardiac silhouette. Radiographs can rarely be used to evaluate reptilian heart, mainly due to the poor differences in tissue contrast (compared to domestic mammals) and anatomical positioning of organs. Indeed, in chelonians and crocodilians the heart is superimposed with other visceral organs, and in lizards and chelonians, the bony pectoral girdle and shell respectively, form a barrier to x-rays. Some of these difficulties may be overcome by using corrected settings, however the use of computed radiography (CR) and direct digital radiography (DR) allow for post-photographic digital enhancement and can greatly enhance the diagnostic value of the image. Ways in which the image can be manipulated include altering size, sharpness, contrast and brightness, as well as the application of filters (digital algorithms), which can be used to enhance edges of structures. Nevertheless, radiographs can be used to evaluate heart size in snakes and monitor lizards because in these species the heart is not located under a radio-opaque organ. Thus, in snakes and monitor lizards, radiography can be used to confirm the presence of cardiomegaly. Another disadvantage of cardiac radiography is the lack of normal reference parameters. This can however be overcome by acquiring routine x-rays of healthy patients in order to build up a personal (or publishable) database of standard parameters.33,55-57a,b,k When assessing for cardiac enlargement, it is worth noting that cardiomegaly may be normal under some circumstances. For example, some studies have demonstrated that in Burmese pythons (Python molurus bivittatus), the ventricular muscle mass can hypertrophy by up to 40% in less than 48 hours post-prandialy. This is thought to occur because of the increase in oxygen consumption to meet the heavy metabolic demands.1 As with any routine radiographic examination, two orthogonal projections should be taken: a dorso-ventral view and lateral view. In chelonians, a cranio-caudal view can be useful, but is most useful for visualizing lung fields.11,33,40,51,k,l,q,r Mineralization of the great vessels can sometimes be observed , which may result from hypervitaminosis D or other metabolic disturbances.b The shoulder girdles provide good landmarks for increasing image quality in chelonians. The heart is usually just caudal to the level of the acromion processes and cranial to the distal procoracoid process-procoracoid cartilage junction.64 D. Echocardiography (Ultrasonography) Ultrasound and Doppler examination are diagnostic tools of choice for ante-mortem evaluation of heart diseases.4,15,42,44-46,48,51,54,l-r A great range of ultrasound equipment is currently available to the veterinary practitioner. Most of these units are suitable to reptiles. Selection of the appropriate transducer is a trade-off between spatial resolution of the image and imaging depth: lower frequencies produce less resolution but image deeper into the body. That’s why the size of the patient dictates the transducer probes that are needed: smaller animals (snakes, most lizards and small chelonians) require the use of a 10 or 15 MHz transducer, a 5 to 8 MHz transducer is used for medium to large size patients. A 3.5 MHz probe may be warranted for very large reptiles such as big crocodiles, giant tortoises, large monitors or sea turtles. A linear array is usually used in snakes and lizards. In chelonians, the use of the biconvex sector scanner is recommended because of the narrow space of the acoustic windows in these reptiles: the linear transducer is too large to fit in the cervical, axillary or prefemoral areas.l-r In lizards whose heart is located at the level of the base of the neck, the ultrasonographic window is cranio-caudal: the probe must be positioned in front of and under the bony pectoral girdle (this neck window is relatively narrow and thus restricts a good visualization of the whole organ). In varanidae and teiidae, the heart is located more caudally in the coelomic cavity and can be easily accessed via ventral approach.l-r In chelonians, placing the transducer in the axillo-cervical (cervico-brachial) fossa will provide access to the heart, which is caudal to the thyroïd. It lies immediatly above the plastron ventrally. The left and right atria are of similar size with thin walls. The ventricle is thick walled and the two atrioventricular valves, whose motion is clearly discernible, are very echogenic. They can be seen as highly reflective ribbons.l-r Echocardiographic examination can also be performed on crocodilians, but rather on small or juvenile animals in which ultrasound waves can penetrate the bony ventral scutes.l-r A standardization of the two-dimensional echocardiography in snakes based on the approach used for humans or other companion carnivorous mammals has been suggested and published. 4,44,p In snakes, the position of the heart is indicated by visualization of the ventral precordial tap. As the snake’s heart is mobile inside the coelomic cavity, the transducer may be moved by a few centimeters, either cranially or caudally in relation to the initial position during the ultrasound examination. A thick layer of acoustic coupling gel has to be applied ventrally to ensure perfect cohesion between the probe and the snake’s scales.4,15,39,42,44,51,52,54,m-p,q Three approaches to define the ultrasound windows can be used in succession (Figure 9).15,33,k Most of the ultrasound examination is carried out by placing the probe ventral to the heart. This ventral approach makes it possible to view the organ from the caudal ventricular apex to the cranial atria, and examine the sinus venosus, the atrioventricular junctions and the three arterial trunks. Two other approaches, known as right and left intercostal approaches, are obtained by lateral positioning of the probe. These approaches are used to complete the ventral cranial examination and obtain a lateral visualization of the three arterial trunks and both atria that proceed into the single ventricle.4,15,39,42,44,51,52,54,m-p,q By placing the probe ventral to the heart, the operator can scan or “sweep” the organ from the apex to the arterial trunks along its short axis (short-axis views). Thus two transventricular sections can be obtained by ventral approach.4,44,p On moving the probe caudal to cranial, from the apex towards the base: the apical or transapical short-axis section shows a transverse section of the apical myocardium and the pericardium from behind, in the form of an echogenic line. the transventricular subarterial short-axis section shows a transverse section of the three Cava surrounded by the peripheral myocardium (the ventral cavum pulmonale the right dorsal cavum venosum, the left dorsal cavum arteriosum). The interventricular (vertical) septum, located in the ventricular cavity between the cavum arteriosum and the cavum venosum can be partially visualized, as can the muscular ridge (horizontal septum) marking the separation between the cavum venosum and the cavum pulmonale.4,44,p By continuing to move the probe cranially, a transarterial short-axis section is obtained giving a transverse section of the three large arterial trunks: the two aortic arches of equal diameter and the pulmonary trunk of larger diameter (having the appearence of a “Mickey-Mousehead”). By leaving the probe in a ventral position, but by moving it slightly towards the right, the right transatrial short-axis section shows the opening of the sinus venous into the right atrium and enables the aspect of both sinoatrial valves to be assessed.4,44,p Long-axis views are obtained by turning the probe 90° in relation to the previous projections (Figure 10).4,44,p Thus, starting from the transventricular or subarterial section, the rotation of the probe enables two long-axis sections of the heart to be obtained. These are called atrioventricular sections and they show both atrial cavities opening into the single ventricle. The left atrioventricular junction can be observed by orienting the ultrasound plane ventro-dorsally from the right to the left. Similarly, the right atrioventricular junction can be observed by orienting the ultrasound plane ventro-dorsally from the left to the right. The atrioventricular valves, also known as septal monocuspid valves, can hence be observed.4,44,p Similarly, starting from the short-axis transarterial section, the probe is rotated 90° to obtain a long-axis transarterial section which reveals the right aortic arch and the pulmonary trunk with parallel paths. The path of the pulmonary artery can be located by moving the probe caudally. The path of this artery comes closer to the probe ventrally and then opens out on the right with the cavum pulmonale.4,44,p Lastly, starting from the right transatrial short-axis section, the probe is rotated 90°, and moved caudally, to display the caudal vena cava running parallel to the pulmonary vein: a long-axis transcaval section, starting from the sinus venosus can thus be obtained.4,44,p When imaging small snakes, the examination may be completed by two intercostal approaches. Thus the transarterial long-axis section, obtained by right intercostal approach, provides clear visualization of the left atrium. The probe is placed laterally on the right so that the cross-section is parallel to the animal’s body, and the left atrium is removed from the proximal field occupied by the large arterial trunks. Conversely, the left symmetrical intercostal section provides a good approach for observing the right atrium.4,44,p REFERENCES 1. Andersen JB, Rourcke BC, Caiozzo VJ, Bennett AF, Hicks JW. Physiology: postprandial cardiac hypertrophy in pythons. Nature 2005; 434:37-8. 2. Barten SL. Cardiomyopathy in a king snake (Lampropeltis calligaster rhombomaculata). Vet Med Small Anim Clin 1980;75:125-29. 3. Burgreen WW, Johansen K. Ventricular haemodynamics in the monitor lizard Varanus exanthematicus: Pulmonary and systemic pressure separation. J Exp Biol 1982;96:343-54. 4. Chetboul V, Schilliger L, Tessier D, et al. Specific features of echocardiographic examination in ophidians. Schweiz Archiv Tierheilk 2004;146:327-34. 5. Cordero-Tapia A, Gardner SC, Arellano-Blanco J, et al. Laeredius learedi infection in black turtles (Chelonia mydas agassizii), Baja California Sur, Mexico. J Parasitol 2004;90:645-47. 6. Farrell AP, Gamperl AK, Francis ETB. Comparative aspects of heart morphology. In: Gans C, Gaunt AS, eds. Biology of the Reptilia, vol. 19, Morphology G, Visceral organs. Ithaca: Society for the Study of Amphibians and Reptiles,1998;375-424. 7. Frye FL. Reptile Care. An atlas of diseases and treatments, vol 1&2. Neptune City: TFH Publications, Inc., 1991. 8. Girling SJ, Hynes B. Cardiovascular and haemopoietic systems In: Girling SJ, Raiti P, eds. BSAVA Manual of Reptiles, Second Edition. Gloucester (UK), British Small Animal Veterinary Association, 2004:243-260. 9. Glazebrook JS, Campbell RS, Blair D. Studies on cardiovascular fluke (Digenea: Spirorchiidae) infections in sea turtles from the Great Barrier Reef, Queensland, Australia. J Comp Pathol 1989;101:231-50. 10. Gordon N, Kelly NR, Cribb TH. Lesions caused by cardiovascular flukes (Digenea: Spirorchiidae) infections in stranged green turtles (Chelonia mydas). Vet Pathol 1998;35:2130. 11. Hernandez-Divers SJ. Diagnostic techniques In: Mader DR, eds. Reptile Medicine and Surgery, 2nd ed. St Louis: Saunders Elsevier, 2006;490-532. 12. Homer BL, Jacobson ER, Schumacher J, et al. Chlamydiosis in marineculture-reared green see turtles (Chelonia mydas). Vet Pathol 1994;31:1-7. 13. Hicks JW. Cardiac shunting in reptiles : mechanisms, regulation and physiological functions. In: Gans C, Gaunt AS, eds. Biology of the Reptilia, vol. 19, Morphology G. Visceral Organs. Ithaca, NY : Society for the Study of Amphibians and Reptiles,1998;425-483. 14. Hruban Z, Vardiman E, Meehan T, et al. Hematopoietic neoplasms in zoo animals. J Comp Pathol 1992;106:15-24. 15. Isaza R, Ackerman N, Jacobson ER. Ultrasound imaging of the coelomic structures in the Boa constrictor (Boa constrictor). Vet Radiol 1993;34:445-50. 16. Jackson DC, Heisler N. The contribution of the alkaline pericardial fluid of freshwater turtles to acid buffering during prolonged anoxia. J Exp Biol 1984;109:55-62. 17. Jacobson ER. Gentamicin related visceral gout in two boid snakes. Vet Med Small Anim Clin 1976;71: 361-63. 18. Jacobson ER., Seely JC, Novilla MN, et al. Heart failure associated with unusual hepatic inclusions in a Deckert’s rat snake. J Wild Dis 1979;15: 75-81. 19. Jacobson ER, Gaskin JM, Mansell J. Chlamydial infection in puff adders (Bitis arietans). J Zoo Wild Med 1989;20: 364-69. 20. Jacobson ER, Homer B, Adams W. Endocarditis and congestive heart failure in a burmese python (Python molurus bivittatus). J Zoo Wild Med 1991;22: 245-48. 21. Jacobson ER, Origgi F, Heard D, et al. Immunohistochemical staining of chlamydial antigen in emerald tree boas (Corallus caninus). J Vet Diagn Invest 2002;14:487-94. 22. Jacobson ER, Ginn PE, Troutman JM, et al. West Nile virus infection in farmed American alligators (Alligator mississipiensis) in Florida. J Wild Dis 2005;41:96-106. 23. Jensen B, Wang T. Hemodynamic consequences of cardiac malformations in two juvenile ball pythons (Python regius). J Zoo Wild Med 2009;40:752-56. 24. Jensen B, Abe AS, Andrade DV, et al. The heart of the south American rattlesnake, Crotalus durissus. J Morphol 2010;271:1066-1077. 25. Jensen B, Nyengaard JR, Pedersen M, et al. Anatomy of the python heart. Anat Sci Int 2010;85(4):194-203. 26. Johnson CA, Griffith JW, Tenorio P, et al. Fatal trematodiasis in research turtles. Lab Anim Sci 1998;48:340-343. 27. Mader D. Gout. In: Mader DR, ed. Reptile Medicine and Surgery, 2nd ed. St Louis: Saunders Elsevier, 2006;793-800. 28. Mauldin GN, Done LB. Oncology In: Mader DR, eds. Reptile Medicine and Surgery, 2nd ed. St Louis: Saunders Elsevier, 2006;299-322. 29. McAllister CT, Cordes JE, Conn DB, Singleton J, Walker JM. Helminth parasites of unisexual and bisexual whiptail lizards (Teiidae) in North America. V. Mesocestoides sp. tetrathyridia (Cestoidea: Cyclophyllidea) from four species of Cnemidophorus. J Wild Dis 1991;27:494-97. 30. McCracken HE. Organ location in snakes for diagnostic and surgical evaluation. In: Miller RE, ed. Zoo and wildlife medicine, current therapy 4. Philadelphia: WB Saunders, 1999; 243248. 31. Millard RW, Johansen K. Ventricular outflow dynamics in the lizard, Varanus niloticus: responses to hypoxia, hypercarbia and diving. J Exp Biol 1974;60:871-80 32. Murray MJ. Cardiopulmonary Anatomy and Physiology In: Mader DR, ed. Reptile Medicine and Surgery, 2nd ed. St Louis: Saunders Elsevier, 2006;124-134. 33. Murray MJ. Cardiology In: Mader DR, ed. Reptile Medicine and Surgery, 2nd ed. St Louis: Saunders Elsevier, 2006;181-195. 34. Mushonga B, Horowitz A. Serous Cavities of the Nile Crocodile (Crocodylus niloticus). Journal of Zoo and Wildlife Medicine 1996;27(2):170-179. 35. Newell SM, Roberts G. Diagnostic imaging. In: Jacobson ER, ed. Infectious Diseases and Pathology of Reptiles. Boca Raton: CRC Press, 2007;168-177. 36. Obendorf DL, Carson J, McManus TJ. Vibrio damsela infection in a stranded leatherback turtle. J Wildl Dis 1987;23: 666-68. 37. Poupa O, Lindström L. Comparative and scaling aspects of heart and body weights with reference to the conducting system. Comp Bioch Physiol 1983;76A:413-421. 38. Redrobe SP, Scudamore CL. Ultrasonographic diagnosis of pericardial effusion and atrial dilatation in a spur-thighed tortoise (Testudo graeca). Vet Rec 2000;146:183-85. 39. Rishniw M, Carmel BP. Atrioventricular valvular insufficiency and congestive heart failure in a carpet python. Aust Vet J 1999;77:580-583. 40. Rübel A, Kuoni W. Radiology and imaging. In: Frye FL, ed. Reptile Care. An atlas of diseases and treatments. Vol 1. Neptune City: TFH Publications, Inc, 1991; 185-208. 41. Rush EM, Donnelly TM, Walberg J. What’s your diagnosis? Cardiopulmonary arrest in a Burmese python. Aortic aneurysm Lab Anim 2001;30: 24-27. 42. Schildger BJ, Tenhu H, Kramer M, et al. Ultraschalluntersuchung bei Reptilien. Berl Münch Tierärztl Wschr 1996;109:136-141. 43. Schilliger L, Vanderstylen D, Pietrain J, et al. Granulomatous myocarditis and coelomic effusion due to Salmonella enterica arizonae in a Madagascarian Dumeril’s boa (Acrantophis dumerili, Jan. 1860). J Vet Cardiol 2003;5:43-45. 44. Schilliger L, Tessier D, Pouchelon JL, et al. Proposed Standardization of the TwoDimensional Echocardiographic Examination in Snakes. J Herp Med Surg 2006;16: 90-102. 45. Schilliger L, Lemberger K, Chai N, et al. Atherosclerosis associated with pericardial effusion in a central bearded dragon (Pogona vitticeps, Ahl. 1926), J Vet Diagn Invest 2010;22(5):789-92. 46. Schilliger L, Tréhiou E, Petit A, et al. Double valvular insufficiency in a Burmese python (Python molurus bivittatus, Linnaeus, 1758) suffering from concomittant bacterial pneumonia. J Zoo Wild Med 2010;41(4):742-44. 47. Schilliger L, Selleri P, Frye FL. Lymphoreticular neoplasm and leukemia in a red-tail boa (Boa constrictor constrictor) associated with concurrent inclusion body disease, J Vet Diagn Invest 2011;23:159-162. 48. Schroff S, Schmidt V, Kiefer I, et al. Ultrasonographic diagnosis of an endocarditis valvularis in a Burmese python (Python molurus bivittatus) with pneumonia. J Zoo Wild Med 2010;41(4):721-24. 49. Schuman SM, Taylor DO. Arteriosclerosis in an iguana (Iguana iguana), J Am Vet Med Assoc 1970;157:614-16. 50. Schumacher JR, Bennett A, Fox LE, et al. Mast cell tumor in an eastern kingsnake (Lampropeltis getulus getulus). J Vet Diagn Invest 1998;10:101-04. 51. Silverman S. Diagnostic imaging In: Mader DR, ed. Reptile Medicine and Surgery, 2nd ed. St Louis: Saunders Elsevier, 2006;471-489. 52. Snyder PS, Shaw NG, Heard DJ. Two-dimensional echocardiographic anatomy of the snake heart (Python molurus bivittatus). Vet Radiol Ultrasound 1999;40:66-72. 53. Starck JM. Functional morphology and patterns of blood flow in the heart of Python regius. J Morphol 2009;270:673-87. 54. Stetter MD. Ultrasonography. In: Mader DR, eds. Reptile Medicine and Surgery, 2nd ed. St Louis: Saunders Elsevier, 2006;665-674. 55. Vallentinuzzi ME, Hoff MD, Geddes ME. Observations on the electrical activity of the snake heart. J Electrocardiology 1969;2(1):39-50. 56. Vallentinuzzi ME, Hoff MD, Geddes ME. Electrocardiogram of the snake: intervals and duration. J Electrocardiology 1969;2(4):343-352. 57. Vallentinuzzi ME, Hoff MD, Geddes ME. Electrocardiogram of the snake: effect of vagal stimulation on the Q-T duration. J Electrocardiology 1970;3(1):21-27. 58. Wang T, Warburton S, Abe A, et al. Vagal control of heart rate and cardiac shunts in reptiles: relation to metabolic state. Exp Physiol 2001;86:777-84. 59. Wang T, Altimiras J, Klein W, et al. Ventricular haemodynamics in Python molurus : separation of pulmonary and systemic pressures. J Exp Biol 2003;206:4241-4245. 60. Webb GJW, Heatwole H, de Bavay J. Comparative cardiac anatomy of the Reptilia. I. The chambers and septa of the varanid ventricle. J Morphol 1971;134:335-350. 61. Webb GJW, Heatwole H, de Bavay J. Comparative cardiac anatomy of the Reptilia. II. A critique of the literature on the Squamata and Rhynchocephalia. J Morphol 1974;142:1-20. 62. Webb GJW. Comparative cardiac anatomy of the Reptilia. III. The heart of crocodilians and an hypothesis on the completion of the interventricular septum of crocodilians and birds. J Morphol 1979;161:221-240. 63. White FN. Functional anatomy of the heart of reptiles. Am Zool 1968;8:211-219. 64. Wyneken J. Normal Reptile Heart Morphology and Function. Veterinary Clin North Am Exot Anim Pract 2009;12:51-63. 65. Zaar M, Overgaard J, Gesser H, et al. Contractile properties of the functionally divided python heart: two sides of the same matter. Comp Biochem Physiol A Mol Integr Physiol 2007;146:163-73. ENDNOTES a . Wyneken J. Anatomy and Physiology Cardiopulmonary Systems in Reptiles, in Proceedings. 17th Annu Conf Assoc Rept Amph Vet 2010;99-106. b . Kik, MJL, Mitchell MA. Reptile Cardiology: A Review of Anatomy and Physiology, Diagnostic Approaches, and Clinical Diseases, in Proceedings. Semin Av Exotic Pet Med 2005;14(1):52-60. c . Williams DL. Cardiovascular system. In: Beynon PH, Lawton MPC, Cooper JE, eds. Manual of Reptiles. Cheltenham: British Small Animal Veterinary Association 1996;80-87. d . Sedgwick CJ. Allometrically scaling the data base for vital sign assessment used in general anesthesia of zoological species, in Proceedings. Assoc Zoo Vet 1991;360-369. e . Frye FL. Characteristics of Cardiomyopathy in Two Pythons: Aortic Valvular Stenosis and Secondary Cardiomyopathy in a Children's Python, Liasis Childreni, and Ventricular Wall Hypoplasia, First-Degree Heart Block, and Plasmacytic Pericarditis in a Juvenile Burmese Python, Python molurus bivittatus, in Proceedings. IV Int. Coll. Path. of Reptiles and Amph, Bad Nauheim, Germany 1991. f . Schmidt R, Reavill D. Cardiopulmonary disease in Reptiles, in Proceedings. 17th Annu Conf Assoc Rept Amph Vet 2010;90-98. g . Innis CJ. Myocardial abcess and hemopericardium in a green iguana (Iguana iguana), in Proceedings. 7th Annu Conf Assoc Rept Amph Vet 2000;185-188. h . Rush EM, Donnelly TM, Walberg J. Aneurysm and subsequent cardiopulmonary arrest in a Burmese python (Python molurus bivittatus), in Proceedings. Am Assoc Zoo Vet 1999;134138. i . Clippinger TL. Aortic stenosis and atrioventricular dilatation in a green iguana (Iguana iguana), in Proceedings. Am Assoc Zoo Vet 1993;342-344. j . Schmidt RE, Reavill DR. Metastatic chondrosarcoma in a corn snake (Elaphe guttata), in Proceedings. 1st Int Conf Rept and Amph Medicine Münich 2010;147. k . Mader DR. Cardiopulmonary diagnostics in Reptiles, in Proceedings. 17th Annu Conf Assoc Rept Amph Vet 2010;107-111. l . Schilliger L. Diagnostic imaging in turtles and tortoises: what a headache, I mean challenge !, in Proceedings. North Am Vet Conf 2009:1798-1801. m . Schilliger L. Ultrasound examination in reptiles: restraint, positioning, artifacts and patient examination, in Proceedings. 1st Int Conf Rept and Amph Medicine Münich 2010;179-187. n . Hochleitner C, Hochleitner M. Ultrasound in reptiles, in Proceedings. 11th Annu Conf Assoc Rept Amph Vet 2004;41-44. o . Schildger BJ, Caesares M, Kramer M, et al. Technique of ultrasonography in lizards, snakes and chelonians, in Proceedings. Seminars in Avian and Exotic Pet Med 1994:147155. p . Schilliger L, Chetboul V, Tessier D. Standardizing two-dimensional echocardiographic examination in snakes, in Proceedings. Int Cong on Exotics 2005;63-74. q . Hernandez-Divers S, Hernandez-Divers S. Diagnostic imaging of reptiles. In Practice 2001;July/August:370-390. r . Bennett RA. Clinical, diagnostic and therapeutic techniques, in Proceedings. 5th Annu Conf Assoc Rept Amph Vet 1998;35-40.