Survey
* Your assessment is very important for improving the workof artificial intelligence, which forms the content of this project
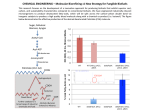
753 Biochem. J. (2003) 371, 753–760 (Printed in Great Britain) Ketone bodies disturb fatty acid handling in isolated cardiomyocytes derived from control and diabetic rats Danny M. HASSELBAINK, Jan F. C. GLATZ, Joost J. F. P. LUIKEN, Theo H. M. ROEMEN and Ger J. VAN DER VUSSE1 Department of Physiology, Cardiovascular Research Institute Maastricht, Maastricht University, Maastricht, The Netherlands According to the current paradigm, fatty acid (FA) utilization is increased in the diabetic heart. Since plasma levels of competing substrates such as ketone bodies are increased during diabetes, the effect of those substrates on cardiac FA handling was explored. Cardiomyocytes were isolated from control and streptozotocin-treated diabetic rats and incubated with normal (80 µM) and elevated (160 µM) palmitate concentrations in the absence or presence of ketone bodies, including acetoacetate (AcAc). Comparing control cardiomyocytes under normal conditions (80 µM, no AcAc) with diabetic cardiomyocytes (160 µM, 3 mM AcAc) showed that palmitate uptake was −1 increased from 35.2 + − 4.8 to 60.2 + − 14.0 nmol · 3 min · g wet weight−1 respectively. Under these conditions, palmitate oxidation rates were comparable (58.9 + − 23.6 versus 53.2 + − 18.5 nmol · 30 min−1 · g wet weight−1 ). However, in the absence of AcAc, palmitate oxidation was significantly enhanced in diabetic cardiomyocytes, indicating that ketone bodies are able to suppress cardiac FA oxidation in diabetes. The concomitantly increased FA uptake in diabetic cells, mainly due to the elevated extracellular FA levels, may be responsible for the accumulation of FA and triacylglycerol, as observed in the diabetic heart in situ. INTRODUCTION EXPERIMENTAL Diabetes mellitus is a clinical syndrome characterized by hyperglycaemia caused by a relative or absolute deficiency of insulin or by resistance to the action of insulin at the cellular level. The diabetic condition leads to a significant elevation of blood glucose and plasma lipids if left untreated [1,2]. Insulin deficiency also results in increased hepatic synthesis of ketone bodies, leading to elevated plasma levels of acetoacetate (AcAc) and 3-β-hydroxybutyrate (3HB) [3], which in turn can evoke serious pathophysiological complications, such as diabetic ketoacidosis. On the basis of studies on isolated hearts it was concluded that cardiac long-chain fatty acid (FA) oxidation is increased in the diabetic state [4–6]. These findings are, however, at variance with observations on hearts in situ of human diabetic patients [7]. This suggests that FA oxidation by the diabetic heart is governed by factors present in the in situ situation, but absent when isolated rat hearts are studied. Under normal conditions, cardiac energy demand is mainly covered by the oxidation of FA and glucose [8]. Lactate and ketone bodies are also able to serve as oxidizable substrates [9]. It is tempting to state that the discrepancy between isolated hearts and hearts of human diabetic patients with respect to cardiac FA utilization is caused by the availability of alternative substrates, such as ketone bodies. In the present study we have tested the hypothesis that ketone bodies, AcAc and 3HB, affect both uptake and intracellular handling of FA in diabetic cardiomyocytes. To this end, cardiomyocytes were isolated from diabetic rats, 4 weeks after streptozotocin (STZ) treatment. The cells were exposed to AcAc, 3HB or a combination of both. FA uptake, oxidation, and esterification were studied in the presence of normal (physiological) or increased (diabetic) extracellular FA concentrations. Treatment of animals Key words: acetoacetate, β-hydroxybutyrate, oxidation, triacylglycerols, uptake. Male Lewis rats were fed ad libitum, had free access to water and were kept under a 12 h/12 h light/dark cycle [10]. The Institutional Animal Care and Use Committee, Maastricht University, approved the experiments. Animals (200–250 g body weight) were anaesthetized with ketamine and xylazine (Eurovet, Bladel, the Netherlands), at 37.5 and 7.5 mg · kg−1 respectively via intraperitoneal injection. Subsequently, the animals were treated with 70 mg · kg−1 STZ in disodium citrate (100 mM, pH 4.5) via intravenous injection in the tail vein to induce type 1 diabetes, and received a subcutaneous injection of 2.5 ml of 20 % (w/v) glucose on both lateral sides. Controls received vehicle only. STZ-treated animals were given daily a 5 % glucose solution to drink. At 4 weeks after STZ treatment, rats were anaesthetized with a subcutaneous injection of 48 mg of sodium pentobarbital (CEVA Sante Animale B.V., Maassluis, the Netherlands) and 1000 i.u. of heparin. Thereafter, hearts were surgically excised. In a subset of experiments (n = 6) the anaesthetized animals did not receive heparin. After thoracotomy, blood was collected from the left ventricle with a citrate-rinsed syringe. Blood glucose was measured with EuroflashTM (Lifescan, Milpitas, CA, U.S.A.) and plasma FA by gas chromatography [11]. Isolation of cardiomyocytes The cardiomyocyte isolation procedure was partially adapted from that described by Luiken et al. [12]. The surgically removed hearts were placed in ice-cold buffer A (containing, in mM: 115 NaCl, 2.6 KCl, 1.2 KH2 PO4 , 1.2 MgSO4 , 10 NaHCO3 , 10 Hepes, 11 glucose, pH 7.4). The aorta was mounted on a cannula Abbreviations used: AcAc, acetoacetate; FA, fatty acid(s); FAT, fatty acid transporter; 3HB, 3-β-hydroxybutyrate; PL, phospholipids; STZ, streptozotocin; TG, triacylglycerols. 1 To whom correspondence should be addressed (e-mail [email protected]). c 2003 Biochemical Society 754 D. M. Hasselbaink and others (diameter 2.0 mm) and hearts were perfused at 37 ◦C for 5 min with buffer A at a flow rate of 7 ml · min−1 retrogradely as described by Langendorff [12a] to remove blood from the coronary circulation. Thereafter, recirculating perfusion was started using 40 ml of buffer A supplemented with 0.7 % (w/v) BSA, 0.075 % (w/v) collagenase and 15 mM butanedione monoxime. After 1 min, the flow rate was increased to 8 ml · min−1 and then increased linearly to 14 ml · min−1 at 15 min after the start of the recirculating perfusion. Flow was allowed to continue at this level for 1 min. At time points 2 and 12 min 40 µl of 0.1 mM CaCl2 was added to the perfusion buffer. At the end of the perfusion period, the heart was removed from the cannula, transferred to a Petri dish and carefully opened using forceps. The opened heart was incubated with 10 ml of perfusate and 10 ml of buffer B (buffer A supplemented with 0.2 mM Ca2+ and 2 % BSA) for 10 min in a shaking waterbath (Aquatron ; HT Infors, Bottmingen, Switzerland) (37 ◦C, 180 rev./min). The heart was subsequently transferred to a Petri dish and was further dissected with forceps. After dissection, the suspension was transferred to an Erlenmeyer flask and placed in a shaking waterbath (37 ◦C, 180 rev./min) for 5 min while the Ca2+ concentration was increased with steps of 200 µM to 1 mM. Next, cell suspension was filtered through a 0.2-mm-pore-size nylon mesh and cardiomyocytes were pelleted by centrifugation (2 min, 17 g). The cell pellet was resuspended in medium C (buffer A supplemented with 1 mM Ca2+ and 2 % BSA) and centrifuged (2 min, 23 g), and this procedure was repeated once. The cell pellet was resuspended in 20 ml of medium C and transferred to a 50 ml conical tube. Cardiomyocytes were allowed to rest for 1.5 h under mild rotation. Prior to the start of the metabolic studies, cells were washed and resuspended in medium C. At the end of the metabolic studies, viability of the cardiomyocytes was assessed by determining the percentage of rod-shaped cells. Furthermore, cellular wet weight was obtained by centrifugation in a microcentrifuge and subsequent removal of the supernatant from samples taken at the end of the performed experiments. All solutions used during the experiment were continuously gassed (O2 /CO2 , 19 : 1). performed as described by Luiken et al. [12]. Uptake experiments were performed in triplicate for each condition tested. Palmitate oxidation rate The rate of palmitate oxidation was determined in the presence or absence of ketone bodies during 30 min as described previously [13]. The reaction was started by the addition of [1-14 C]palmitate to the capped glass vial. After termination of the reaction, production of 14 CO2 was measured after base trapping in NaOH. The obtained 14 CO2 values were corrected for 14 CO2 trapped in vials supplemented with [1-14 C]palmitate but in the absence of cardiomyocytes in the incubation medium. Thereafter, this value was normalized on mg wet weight of cardiomyocytes and corrected for the percentage of non-rod-shaped cells. Oxidation experiments were performed in triplicate for each condition tested. Incorporation of palmitate into intracellular lipids A 2.0 ml suspension of cardiomyocytes was incubated in a capped glass vial in the absence or presence of ketone bodies, in a shaking waterbath (37 ◦C, 180 rev./min). The reaction was started by the addition of [1-14 C]palmitate. After 30 min the reaction was stopped as described by Luiken et al. [12]. Cells were pelleted by centrifugation (2 min, 60 g, 4 ◦C) and washed twice with 5.0 ml of ice-cold stop solution. The pellet was solubilized in buffer containing (in mM), 50 Tris/HCl, pH 7.6, 150 NaCl, 5 EDTA, and 1 % (w/v) Triton X-100. Lipids were extracted with chloroform/methanol and separated by TLC [11]. The lipid spots corresponding to triacylglycerol (TG), phospholipids (PL) and FA were scraped from the plate and the amount of radioactivity was determined with a liquid-scintillation counter. Incorporation experiments were performed in duplicate for each condition tested. Preparation of palmitate–BSA complex Choice of substrate concentrations To prepare a stock solution of palmitate–BSA complex, 100 µCi of [1-14 C]palmitate and either 1.8 or 3.6 mM palmitate in 10 ml of 100 % ethanol were mixed with water containing KOH (1.5 times the amount of palmitate on molar basis). After evaporation of ethanol at 45 ◦C under nitrogen, the saponified palmitate was added to 40 ml of buffer A supplemented with 2.5 % BSA. Prior to incubation, the stock solution was diluted 4.5-fold with buffer A supplemented with 1.0 mM CaCl2 . Final extracellular concentrations of 80 µM and 160 µM [1-14 C]palmitate were used. They correspond to the interstitial concentrations of FA in healthy and diabetic rats respectively, taking into account that the interstitial FA concentration is approx. 60 % of the arterial FA level [14]. These FA conditions were combined with various concentrations of AcAc, 3HB or a combination of both ketone bodies. In general, in STZ-treated rats, plasma concentration of ketone bodies is of the order of 3–6 mM [15,16], with a ratio of 3HB to AcAc of 2 : 1 [15]. Since AcAc was added to the incubation medium as the lithium salt, the effect of 3 mM Li+ (as LiCl) was tested on FA uptake, oxidation, and deposition of label in the various lipid pools. No effect of Li+ on FA handling was found (results not shown). Palmitate uptake rate The rate of palmitate uptake was determined during 3 min as described previously [13]. In short, a 2.0 ml suspension of freshly isolated cardiomyocytes was incubated in a capped glass vial (O2 /CO2 , 19 : 1) in the absence or presence of ketone bodies, in an Aquatron shaking waterbath (37 ◦C, 180 rev./min). After thermal equilibration, 0.5 ml of labelled palmitate (80 or 160 µM final concn.) was added and incubation allowed to proceed for 3 min. Next, 2.0 ml of the incubation mixture was transferred to a tube containing 8.0 ml of ice-cold stop solution (buffer A supplemented with 0.1 % BSA, 1.0 mM CaCl2 and 200 µM phloretin). Correction for extracellular radioactivity was c 2003 Biochemical Society Materials [1-14 C]Palmitic acid was obtained from NEN Life Science Products (Boston, MA, U.S.A.). Collagenase type 2 (LS04176) was from Worthington (Lakewood, NJ, U.S.A.), and phloretin, BSA fraction V (A4503), STZ, LiCl, acetoacetic acid (lithium salt), 3-β-hydroxybutyric acid, palmitic acid and butane-2,3dione monoxime was from Sigma (St. Louis, MO, U.S.A.). Ketone bodies and cardiac fatty acid handling 755 Table 1 Initial palmitate uptake rate in control- and diabetic-rat cardiomyocytes Data are expressed as means + − S.D. (n = 5). The asterisk (∗) indicates significantly different from the value of the corresponding incubation with 80 µM palmitate (P < 0.05). Statistical analysis revealed that the higher mean values in diabetic-rat cardiomyocytes as compared with corresponding controls did not reach the level of significance. ‘6mM combination’ means a mixture of 3HB (4 mM) and AcAc (2 mM). The final incubation medium contained (in mM) NaCl (115), KCl (2.6), KH2 PO4 (1.2), MgSO4 (1.2), NaHCO3 (10), Hepes (10), glucose (11), BSA (0.3), pH 7.4, palmitate and ketone-body concentrations were as indicated. Initial uptake (nmol · 3 min−1 · g wet weight−1 ) [Palmitate] (µM) Ketone bodies Control Diabetic 80 – 3 mM AcAc 3 mM 3HB 6 mM combination 35.2 + − 4.8 33.4 + − 3.9 36.0 + − 8.0 36.0 + − 5.0 40.1 + − 7.6 39.3 + − 9.0 41.2 + − 9.4 41.4 + − 12.8 160 – 3 mM AcAc 3 mM 3HB 6 mM combination ∗ 54.8 + − 5.0 ∗ 55.3 + 8.4 − ∗ 60.7 + 7.6 − 64.0 + 10.1 − ∗ 64.0 + − 13.8 ∗ 60.2 + 14.0 − ∗ 71.5 + 14.2 − ∗ 70.1 + 27.5 − Statistics Results were obtained from at least four different cardiomyocyte isolations and are presented as means + − S.D. Analysis of experiments within one group was performed with two-tailed Student’s t tests. Comparison between groups was performed with the one-way analysis of variance. In case the F ratio obtained indicated that significant differences between groups were present, a two-tailed Student’s t test for unpaired data was carried out, applying Bonferroni’s adjustment for multiple comparison [17]. For all analyses, the level of significance was set at 0.05. Figure 1 Influence of ketone bodies on the rate of FA oxidation Cardiomyocytes derived from control and diabetic rats were incubated with [1-14 C]palmitate for 30 min. For further details see the legend to Table 1. AcAc (3 mM) and 3HB (3 mM) were either present separately or in combination [4 mM 3HB + 2 mM AcAc; ‘3HB/AcAc 6 mM (2 : 1)’]. Data are expressed as means + − S.D. (n = 6). Statistical significance : *P < 0.05, versus corresponding absence of ketone bodies (−) ; #P < 0.05, control versus corresponding diabetic : +P < 0.05, normal FA versus corresponding high FA. RESULTS General characteristics of STZ-induced diabetic rats The body weight of the diabetic rats (301 + − 49 g) was significantly lower than that of controls (394 + − 18 g). Diabetic plasma FA levels were significantly higher than control (265 + − 88 versus 59 µM). Diabetic-rat blood glucose concentrations were 134 + − significantly higher than the control-rat values, namely 20.4 + − 6.5 and 9.1 + − 1.0 mM respectively. Initial palmitate uptake rate There was no difference in the initial uptake rate of palmitate between control- and diabetic-rat cardiomyocytes at either normal (80 µM) or elevated (160 µM) palmitate concentrations (Table 1). However, increasing the palmitate concentration from 80 to 160 µM significantly enhanced its uptake. The percentage of increase did not differ between control- and diabetic-rat cardiomyocytes, being on the average 56 and 59 % respectively. Ketone bodies did not affect the initial uptake rate of FA in either control- or diabetic-rat cardiomyocytes, irrespective of the palmitate concentration. Palmitate oxidation rate The rate of palmitate oxidation at 80 µM palmitate, in the absence of ketone bodies, is about 56 % higher in diabetic-rat cardiomyocytes than in those of control rats (P < 0.05) (Figure 1). Increasing the palmitate concentration from 80 to 160 µM resulted in a 70 % increase in oxidation rate in control-rat cardiomyocytes (P < 0.05). The percentage increase in diabeticrat cardiomyocytes was comparable (68 %). Addition of 6 mM ketone bodies (3HB/AcAc, 2 : 1) to control cardiomyocytes significantly decreased the oxidation rate, both at normal and elevated palmitate concentrations, by 51 and 58 % respectively. Comparable inhibitory effects were seen in diabetic-rat cardiomyocytes. Figure 1 also shows the differential effect of 3HB and AcAc. When cardiomyocytes were exposed to 3HB (3 mM), the inhibitory effect was insignificant under all conditions analysed. In contrast, exposure to 3 mM AcAc substantially inhibited palmitate oxidation to a comparable extent in controland diabetic-rat cardiomocytes, at both palmitate concentrations. Influence of various concentrations of ketone bodies on FA oxidation Figure 2 show that palmitate oxidation can be suppressed to approx. 30 % of its maximal rate by AcAc. This is true for cardiomyocytes derived from control and diabetic animals. The maximal percentage of inhibition was found to be independent from the two palmitate concentrations applied (Figure 2). Half-maximum inhibition is reached at 0.3–0.5 mM AcAc and c 2003 Biochemical Society 756 D. M. Hasselbaink and others Figure 2 Influence of various concentrations of ketone bodies on the rate of FA oxidation Control- and diabetic-rat cardiomyocytes were incubated with either 80 or 160 µM palmitate, both with various concentrations of ketone bodies, for 30 min. For further details see the legend to Table 1. , AcAc; , 3HB/AcAc (2 : 1, mol/mol). Data are expressed as means + − S.D. (n = 4). • maximum inhibition at 2 mM AcAc and higher. AcAc is therefore a highly potent inhibitor of cardiomyocyte FA oxidation, and the sensitivity to AcAc hardly differs between control- and diabeticrat cells. The findings in Figure 2 also indicate that it is mainly AcAc that causes the inhibitory effect of the mixture of AcAc and 3HB, supporting the results shown in Figure 1. Incorporation of labelled palmitate in various intracellular lipid pools In control cardiomyocytes, incubated under normal conditions (80 µM palmitate, no ketone bodies), incorporation of labelled palmitate in the cellular PL, TG and FA pool amounted to −1 0.27 + − 0.14, 1.46 + − 0.80 and 0.07 + − 0.05 nmol · 30 min · g wet weight−1 respectively. In diabetic-rat cardiomyocytes, incubated under diabetic conditions (160 µM palmitate, 3 mM AcAc) substantially higher amounts of labelled palmitate were incorporated in the PL, TG and FA pool, i.e. 0.69 + 5.15 + − 0.32, − −1 −1 · g wet weight . However, 3.02 and 0.19 + 0.18 nmol · 30 min − c 2003 Biochemical Society due to a relatively high degree of variation between the individual experiments, the increase in the deposition of palmitate into the various lipid pools in the diabetic cells did not reach the level of significance. To evaluate the effect of varying the ambient palmitate concentration and the addition of AcAc to the incubation medium, the values obtained in either control or diabetic-rat cells incubated in the presence of 80 µM palmitate, in the absence of AcAc, was set at 1.0. This procedure enabled us to use the cells as their own control. Raising the concentration from 80 to 160 µM tended to increase the incorporation of palmitate in the PL and TG pool of control cells (Figure 3). The increased deposition of label in the PL, TG and FA pool of diabetic cardiomyocytes was highly significant when the extracellular palmitate concentration was increased from 80 to 160 µM (P < 0.05). Exposure of cardiomyocytes, either derived from control or diabetic hearts, to 3 mM AcAc at 80 µM palmitate did not affect the incorporation of palmitate in the PL and TG pool. The FA pool in diabetic cardiomyocytes incubated in the presence of 80 µM palmitate showed a significant expansion when exposed to 3 mM AcAc. This effect was not seen in control-rat cardiac muscle cells. When the concentration of palmitate was increased from Ketone bodies and cardiac fatty acid handling 757 DISCUSSION The present observations indicate that ketone bodies, in particular AcAc, affect FA handling in the cardiac muscle cell. This finding is of interest because it allows one to fully appreciate the changes in cardiac FA utilization in the diabetic heart, since the circulating concentration of ketone bodies is substantially increased in this pathophysiological condition. AcAc causes a mismatch between FA uptake (not affected by AcAc, but increased due to enhanced supply) and FA oxidation (significantly inhibited by AcAc) when diabetic cardiomyocytes, derived from STZ-treated rats and incubated under diabetic conditions, i.e. 160 µM palmitate, 3 mM AcAc, were compared with control cardiomyocytes, incubated under normal conditions, i.e. 80 µM palmitate, no ketone bodies. This mismatch most likely explains the accumulation of lipids, FA and TG in the diabetic heart in situ. Moreover, the present findings are challenging the current paradigm that FA oxidation is enhanced in the diabetic-rat heart, since this notion is based on experiments performed on isolated hearts perfused with media devoid of ketone bodies. General background Figure 3 Deposition of radiolabelled palmitate in various lipid pools in control- and diabetic-rat cardiomyocytes After incubation for 30 min with radiolabelled palmitate, lipids were extracted and subjected to TLC. Data are presented relative to the values obtained in control- and diabetic-rat cardiomyocytes respectively, incubated in the presence of 80 µM palmitate without ketone bodies. For further details see the legend to Table 1. The asterisk () indicates significantly different from the corresponding incubation with 80 µM palmitate, without ketone bodies. Data are expressed as means + − S.D. (n = 4). 80 to 160 µM in the incubation medium of control- or diabeticrat cells, exposed to 3 mM AcAc, the deposition of labelled palmitate tended to increase (control), or significantly increased (diabetic-rat cells), in the PL and TG pool. Since this pattern was also observed in cells incubated in the absence of AcAc, increased deposition of labelled FA in the esterified lipid pools is mainly caused by increased availability of FA rather than by a modulating effect of AcAc. The same conclusion holds for the FA pool in control cardiomyocytes. In contrast, in diabetic-rat cardiomyocytes, bulk accumulation of labelled palmitate in the FA pool seems to be mainly caused by the presence of AcAc in the incubation medium. Earlier findings in hearts isolated from STZ-treated rats and immediately freeze-clamped for tissue lipid analysis indicated a 4.5-fold accumulation of FA in the diabetic-rat heart, while blood FA levels were enhanced only two-fold [11]. Accumulation of FA in cardiac tissue, also reported by Heyliger and colleagues [18], strongly suggests a mismatch between the uptake of extracellular FA and their intracellular utilization. A common feature in patients suffering from diabetes and in experimental diabetic animals is an increased circulatory level of ketone bodies, AcAc and 3HB [19]. This increase is most striking in diabetic ketoacidosis, a major complication of type 1 diabetes. Elevated levels of ketone bodies have been also observed in poorly controlled type 2 diabetes, chronic cardiac failure and during the consumption of high-fat diet [20–23]. Pioneering studies of Randle and colleagues have shown that ketone bodies serve as alternative substrates for energy conversion in cardiac muscle [19]. Considering the enhanced availability of ketone bodies in the diabetic state, it is likely that the presence of those substances interferes with myocardial lipid metabolism. To fulfil cardiac energy requirements, two main nutrients, FA and glucose, are continuously supplied to the heart, extracted from the extracellular compartment and oxidized in the mitochondrial matrix to regenerate the myocytal ATP pool. Under normal circumstances, oxidation of plasma-borne FA contributes up to 60–70 % of energy conversion in the healthy heart [8]. In the diabetic-rat heart, fuel selection is significantly changed, but the direction and magnitude of the alterations in cardiac FA consumption appear to differ greatly between studies dealing with this subject matter [8]. These considerations prompted us to investigate the effect of ketone bodies on FA handling by cardiac muscle cells derived from STZ-treated rats. Initial palmitate uptake rate The present findings clearly indicate that the rate of palmitate uptake by isolated cardiomyocytes is not changed in the diabetic state when the cells were exposed to normal extracellular palmitate concentrations, i.e. 80 µM. Elevation of the concentration to 160 µM (diabetic conditions) caused a significant and comparable increase in the initial rate of palmitate uptake, both in diabetic- and control-rat cardiomyocytes, indicating that, under the present conditions, trans-sarcolemmal FA transport c 2003 Biochemical Society 758 D. M. Hasselbaink and others Figure 4 Initial uptake and oxidation of palmitate by control- and diabeticrat cardiomyocytes This Figure is a compilation of data shown in Table 1 and Figure 1. Control cardiomyocytes were incubated for either 3 min (initial uptake) or 30 min (oxidation) under normal conditions, i.e. 80 µM palmitate, in the absence of AcAc; diabetic-rat cardiomyocytes were incubated for similar time intervals under diabetic conditions, i.e. 160 µM palmitate in the presence of 3 mM AcAc. Data are expressed as means + − S.D. (n = 5 or 6). The asterisk () indicates a statistically significant difference between control and diabetic cells. is not altered in diabetic-rat cells. This result might appear surprising, since previous studies had shown that STZ treatment resulted in an increase in the myocardial content of proteins known to facilitate FA uptake, including fatty acid transporter (FAT)/CD36 [24,25]. A possible explanation is that the increased amount of FAT/CD36 is stored in the cytoplasmic compartment rather than incorporated into the plasma membrane. The fact that neither AcAc nor 3HB, present in the incubation medium, altered the uptake rate of palmitate in cardiomyocytes indicates that the mechanisms underlying the unidirectional influx of FA across the sarcolemma are insensitive to the presence of ketone bodies in the interstitial compartment. Since diabetic-rat cardiomyocytes in vivo are exposed to both elevated FA levels and enhanced concentrations of ketone bodies, the conclusion can be drawn that diabetic-rat cardiomyocytes take up higher amounts of FA from their surroundings than do the control, merely because the extracellular FA availability is increased (Figure 4). weight−1 ; 80 µM palmitate, no ketone bodies) (Figure 4). It is noteworthy that the inhibitory effect of ketone bodies on myocyte FA oxidation is almost exclusively caused by AcAc, since the effect of the reduced counterpart, 3HB, was found to be negligible. The lack of effect of 3HB is remarkable, considering that both 3HB and AcAc can be used as oxidizable substrate by the heart [19]. At present no conclusive explanation can be offered for this intriguing observation, although differences in redox state in the cardiomyocytes exposed to either AcAc or 3HB should be considered. In this respect, the previously obtained findings by Isales and colleagues are worth mentioning, as they also observed differential effects of 3HB and AcAc, namely on the regulation of growth factors in brain endothelial cells [26]. Detailed analysis of the inhibitory effect of AcAc on cardiomyocyte palmitate oxidation revealed that, both in controland diabetic-rat cells, AcAc is a potent inhibitor of palmitate oxidation. The maximal degree of inhibition in both cell types was on the order of 70 %, irrespective of the concentration of palmitate in the extracellular fluid. Since ketone bodies are permeant to cells via the H+ /monocarboxylate co-transporter [27–29], transport limitations of ketone bodies through this protein-mediated process could therefore contribute to the inability to depress FA oxidation any further. Half-maximal inhibition was obtained at about 0.3–0.5 mM AcAc, indicating that, on the one hand, FA oxidation by cardiomyocytes is highly sensitive to AcAc in the surrounding medium, since this relatively low concentration of AcAc is readily reached under a variety of conditions [30] and, on the other, that the sensitivity to AcAc did not differ between diabeticrat and healthy-rat cardiomyocytes. The latter indicates that cardiomyocytes in diabetic-rat hearts do not adjust to chronically elevated levels of ketone bodies by altering their sensitivity towards the inhibitory action of these compounds. This kind of adjustment might have been expected, since previous observations by Grinblat and colleagues [31] and Kante and co-workers [32] showed that the expression and activity of two enzymes involved in ketone-body handling, namely 3-β-hydroxybutyrate dehydrogenase and 3-oxoacid-CoA transferase, are significantly decreased in the diabetic-rat heart as compared with controls. Obviously, not merely the intracellular conversion, but also other, as yet unidentified properties may be involved in the inhibitory action of ketone bodies on cardiac FA oxidation. Palmitate oxidation rate The rate of palmitate oxidation was found to be significantly higher in cardiomyocytes isolated from diabetic-rat hearts than in control cardiomyocytes when incubated under comparable normal conditions (80 µM palmitate, no ketone bodies). These findings support some earlier observations in isolated rat hearts, namely that the rate of FA oxidation is increased in hearts obtained from diabetic animals [4,6]. Increasing the extracellular palmitate concentration from 80 to 160 µM enhanced the FA oxidation rate about 70 % both in control and diabetic cardiomyocytes. Comparing control cells at normal ambient FA levels with diabetic-rat cardiomyocytes subjected to diabetic FA concentrations revealed that the rate of palmitate oxidation is approx. 3 times higher in the latter (Figure 1). However, this picture substantially changes when ketone bodies were added to the incubation medium to mimic the diabetic conditions as much as possible. Co-incubation of diabetic-rat cardiac muscle cells with 160 µM palmitate and 3 mM AcAc inhibited palmitate oxidation by approx. 65 %. The resulting oxidation rate −1 −1 (53.2 + − 18.5 nmol · 30 min · g wet weight ) was not significantly different from that measured in control cells incubated −1 under control conditions (58.9 + − 23.6 nmol · 30 min · g wet c 2003 Biochemical Society Incorporation of palmitate in cellular lipid pools In general, the amount of labelled palmitate sequestered in the esterified lipid pool (TG and PL) depended on the extracellular FA concentration. Increasing the supply of palmitate resulted in a higher degree of esterification, both in control- and diabetic-rat cardiomyocytes. AcAc exerted no significant effect on the rate of palmitate esterification under these conditions. The observation that the TG content is enhanced in the diabetic-rat heart in situ [33] is supported by the present findings. It is noteworthy that the increased supply of extracellular FA, rather than the presence of ketone bodies, is instrumental in the enhanced deposition of palmitate into the intracellular TG pool (Figure 3). Previous studies also indicated that ketone bodies are able to inhibit lipolysis [34], which may contribute to the net expansion of the TG pool in the diabetic-rat heart in situ. The present findings, however, do not support this notion, since (additive) effects of AcAc on the incorporation of palmitate into the intracellular TG pool were absent. The present study clearly shows that, in diabetic-rat cardiomyocytes AcAc increases the intracellular level of nonesterified palmitate, both at the extracellular concentration of Ketone bodies and cardiac fatty acid handling 80 and 160 µM palmitate (Figure 3). It is noteworthy that the 4-fold increase in deposition of labelled palmitate in the cellular FA pool matches very well with the 4.5-fold increase in the FA content of diabetic hearts in situ [11]. Collectively, the present findings strongly suggest that accumulation of FA in cardiac tissue is caused by a combination of two separate changes in the diabetic cardiomyocyte: the initial uptake rate of FA by diabeticrat cardiomyocytes is enhanced, due to the increased supply of extracellular palmitate, on the one hand, and the inhibitory action of ketone bodies, in particular AcAc, on the intracellular rate of FA oxidation, on the other. This notion is illustrated in Figure 4, which summarizes the results shown in Table 1 and Figure 1. It should be emphasized, however, that the present data are obtained in resting cardiac muscle cells. It would be interesting to explore how, at enhanced workload (e.g. the beating heart), circulating ketone bodies interact with cardiac FA handling. Possible implications of the present findings One may speculate on the pathophysiological consequences of chronically elevated FA levels inside the cardiomyocytes of the diabetic-rat heart. Pioneering studies of Feuvray and colleagues have shown that accumulation of lipotoxic FA and their intermediates are associated with mechanical dysfunction and cell damage in diabetic hearts subjected to ischaemia [35]. Furthermore, other studies indicated that FA at elevated concentrations exert cytotoxic effects, including aberrations in signaltransduction cascades [36–38]. More recent observations by Van der Lee and colleagues [10] and others [39–41] have convincingly shown that chronic exposure of cardiomyocytes to FA modulates the expression of a panel of cardiac enzymes involved in carbohydrate and fatty acid handling. The down-regulation of hexokinase II and the up-regulation of long-chain acyl-CoA dehydrogenase, especially, may explain the decline in glucose utilization and enhanced FA oxidation commonly observed in isolated hearts obtained from STZ-treated diabetic rats [10]. The present findings, however, challenge the current paradigm that FA oxidation is enhanced in the intact diabetic heart [42,43]. The latter notion is based on experimental findings showing a significant increase in the oxidation rate of radiolabelled palmitate by hearts isolated from STZ-treated rats [44]. Results of other studies, however, do not support these findings. The rate of FA oxidation was found to be decreased in hearts from spontaneously diabetic rats [43], and unchanged in hearts obtained from insulinresistant obese rats [45]. Moreover, the utilization of FA by the hearts in vivo of diabetic swine was found to be not significantly different from control [44], while the uptake and oxidation of iodine-labelled FA analogues by hearts of patients with impaired glucose tolerance was decreased [7]. Finally, cardiac FA uptake and oxidative utilization of patients suffering from type 1 and type 2 diabetes did not significantly differ from healthy controls [7]. Since the contrasting findings are most pronounced between isolated hearts (enhanced FA utilization) and intact hearts in situ (no changes or even a decline in FA uptake and/or oxidation), the regulatory role of other factors present in blood plasma, but not in buffers used in perfusion experiments, on isolated hearts should be considered. These factors may be, among others, elevated blood concentration of ketone bodies in the intact experimental animal or subjects suffering from diabetes. The present findings might also have ramifications for other conditions associated with enhanced plasma ketone-body levels, such as starvation, high-fat diets and cardiac failure. Under these conditions cardiac FA handling may be compromised by elevated exposure of the heart to circulating ketone bodies. 759 REFERENCES 1 Stearns, S. B., Tepperman, H. M. and Tepperman, J. (1979) Studies on the utilization and mobilization of lipid in skeletal muscles from streptozotocin-diabetic and control rats. J. Lipid Res. 20, 654–662 2 Wahren, J., Hagenfeldt, L. and Felig, P. (1975) Splanchnic and leg exchange of glucose, amino acids, and free fatty acids during exercise in diabetes mellitus. J. Clin. Invest. 55, 1303–1314 3 Owen, O. E., Markus, H., Sarshik, S. and Mozzoli, M. (1973) Relationship between plasma an muscle concentrations of ketone bodies and free faaty acids in fed, starved and alloxan-diabetic states. Biochem. J. 134, 499–506 4 Christe, M. E. and Rodgers, R. L. (1995) Cardiac glucose and fatty acid oxidation in the streptozotocin-induced diabetic spontaneously hypertensive rat. Hypertension 25, 235–241 5 Rodrigues, B. and McNeill, J. H. (1992) The diabetic heart: metabolic causes for the development of a cardiomyopathy. Cardiovasc. Res. 26, 913–922 6 Saddik, M. and Lopaschuk, G. D. (1994) Triacylglycerol turnover in isolated working hearts of acutely diabetic rats. Can. J. Physiol. Pharmacol. 72, 1110–1119 7 Turpeinen, A. K., Kuikka, J. T., Vanninen, E. and Uusitupa, M. I. (1997) Abnormal myocardial kinetics of 123 I-heptadecanoic acid in subjects with impaired glucose tolerance. Diabetologia 40, 541–549 8 van der Vusse, G. J., Glatz, J. F., Stam, H. C. and Reneman, R. S. (1992) Fatty acid homeostasis in the normoxic and ischemic heart. Physiol. Rev. 72, 881–940 9 Neely, J. R. and Feuvray, D. (1981) Metabolic products and myocardial ischemia. Am. J. Pathol. 102, 282–291 10 van der Lee, K. A., Vork, M. M., De Vries, J. E., Willemsen, P. H., Glatz, J. F., Reneman, R. S., Van der Vusse, G. J. and Van Bilsen, M. (2000) Long-chain fatty acid-induced changes in gene expression in neonatal cardiac myocytes. J. Lipid Res. 41, 41–47 11 Hasselbaink, D. M., Roemen, T. H. and van der Vusse, G. J. (2002) Determination of long-chain fatty acids, triacylglycerols and phospholipids by capillary gas chromatography in cardiac tissue and blood plasma of normal and diabetic rats. Anal. Chim. Acta 465, 351–357 12 Luiken, J. J., van Nieuwenhoven, F. A., America, G., van der Vusse, G. J. and Glatz, J. F. (1997) Uptake and metabolism of palmitate by isolated cardiac myocytes from adult rats: involvement of sarcolemmal proteins. J. Lipid Res. 38, 745–758 12a Langendorff, O. (1895) Untersuchungen am uberlebenden Saugethierherzen. Pflügers Arch. Gesamte Physiol. Menschen Tiere 61, 291–332 13 Luiken, J. J., Willems, J., van der Vusse, G. J. and Glatz, J. F. (2001) Electrostimulation enhances FAT/CD36-mediated long-chain fatty acid uptake by isolated rat cardiac myocytes. Am. J. Physiol. Endocrinol. Metab. 281, E704–E712 14 Julien, P., Gailis, L., Lepage, M. and Roy, P. E. (1979) A comparison of fatty acid patterns of arterial plasma, pericardial fluid and cardiac lymph in dog. Artery 5, 37–44 15 Charlton, J. A., Thompson, C. J. and Baylis, P. H. (1988) Possible mechanisms responsible for the rise in plasma vasopressin associated with diabetic ketoacidosis in the rat. J. Endocrinol. 116, 343–348 16 Kashiwaya, Y., King, M. T. and Veech, R. L. (1997) Substrate signaling by insulin : a ketone bodies ratio mimics insulin action in heart. Am. J. Cardiol. 80, 50A–64A 17 Wallenstein, S., Zucker, C. L. and Fleiss, J. L. (1980) Some statistical methods useful in circulation research. Circ. Res. 47, 1–9 18 Heyliger, C. E., Scarim, A. L., Eymer, V. P., Skau, K. A. and Powell, D. M. (1997) Characteristics of the myocardial PM-FABP: effect of diabetes mellitus. Mol. Cell. Biochem. 176, 281–286 19 Randle, P. J., Newsholme, E. A. and Garland, P. B. (1964) Regulation of glucose uptake by muscle. 8. Effects of fatty acids, ketone bodies and pyruvate, and of alloxan-diabetes and starvation, on the uptake and metabolic fate of glucose in rat heart and diaphragm muscles. Biochem. J. 93, 652–665 20 Edens, N. K. and Friedman, M. I. (1984) Response of normal and diabetic rats to increasing dietary medium-chain triglyceride content. J. Nutr. 114, 565–573 21 Lommi, J., Koskinen, P., Naveri, H., Harkonen, M. and Kupari, M. (1997) Heart failure ketosis. J. Intern. Med. 242, 231–238 22 Lommi, J., Kupari, M. and Yki-Jarvinen, H. (1998) Free fatty acid kinetics and oxidation in congestive heart failure. Am. J. Cardiol. 81, 45–50 23 Umpierrez, G. E., Watts, N. B. and Phillips, L. S. (1995) Clinical utility of β-hydroxybutyrate determined by reflectance meter in the management of diabetic ketoacidosis. Diabetes Care 18, 137–138 24 Glatz, J. F., van Breda, E., Keizer, H. A., de Jong, Y. F., Lakey, J. R., Rajotte, R. V., Thompson, A., van der Vusse, G. J. and Lopaschuk, G. D. (1994) Rat heart fatty acid-binding protein content is increased in experimental diabetes. Biochem. Biophys. Res. Commun. 199, 639–646 c 2003 Biochemical Society 760 D. M. Hasselbaink and others 25 Pelsers, M. M., Lutgerink, J. T., Nieuwenhoven, F. A., Tandon, N. N., van der Vusse, G. J., Arends, J. W., Hoogenboom, H. R. and Glatz, J. F. (1999) A sensitive immunoassay for rat fatty acid translocase (CD36) using phage antibodies selected on cell transfectants: abundant presence of fatty acid translocase/CD36 in cardiac and red skeletal muscle and up-regulation in diabetes. Biochem. J. 337, 407–414 26 Isales, C. M., Min, L. and Hoffman, W. H. (1999) Acetoacetate and β-hydroxybutyrate differentially regulate endothelin-1 and vascular endothelial growth factor in mouse brain microvascular endothelial cells. J. Diabetes Complications 13, 91–97 27 Poole, R. C. and Halestrap, A. P. (1993) Transport of lactate and other monocarboxylates across mammalian plasma membranes. Am. J. Physiol. 264, C761–C782 28 Wang, X., Levi, A. J. and Halestrap, A. P. (1996) Substrate and inhibitor specificities of the monocarboxylate transporters of single rat heart cells. Am. J. Physiol. 270, H476–H484 29 Zorzano, A., Fandos, C. and Palacin, M. (2000) Role of plasma membrane transporters in muscle metabolism. Biochem. J. 349, 667–688 30 Laffel, L. (1999) Ketone bodies: a review of physiology, pathophysiology and application of monitoring to diabetes. Diabetes Metab. Res. Rev. 15, 412–426 31 Grinblat, L., Pacheco Bolanos, L. F. and Stoppani, A. O. (1986) Decreased rate of ketone-body oxidation and decreased activity of D-3-hydroxybutyrate dehydrogenase and succinyl-CoA: 3-oxo-acid CoA transferase in heart mitochondria of diabetic rats. Biochem. J. 240, 49–56 32 Kante, A., Malki, M. C., Coquard, C. and Latruffe, N. (1990) Metabolic control of the expression of mitochondrial D-β-hydroxybutyrate dehydrogenase, a ketone body converting enzyme. Biochim. Biophys. Acta 1033, 291–297 33 Paulson, D. J. and Crass, 3rd, M. F. (1982) Endogenous triacylglycerol metabolism in diabetic heart. Am. J. Physiol 242, H1084–H1094 34 Larsen, T. S. and Severson, D. L. (1990) Influence of exogenous fatty acids and ketone bodies on rates of lipolysis in isolated ventricular myocytes from normal and diabetic rats. Can. J. Physiol. Pharmacol. 68, 1177–1182 Received 16 October 2002/20 November 2002; accepted 18 December 2002 Published as BJ Immediate Publication 18 December 2002, DOI 10.1042/BJ20021617 c 2003 Biochemical Society 35 Feuvray, D., Idell-Wenger, J. A. and Neely, J. R. (1979) Effects of ischemia on rat myocardial function and metabolism in diabetes. Circ. Res. 44, 322–329 36 de Vries, J. E., Vork, M. M., Roemen, T. H., de Jong, Y. F., Cleutjens, J. P., van der Vusse, G. J. and van Bilsen, M. (1997) Saturated but not mono-unsaturated fatty acids induce apoptotic cell death in neonatal rat ventricular myocytes. J. Lipid Res. 38, 1384–1394 37 Listenberger, L. L., Ory, D. S. and Schaffer, J. E. (2001) Palmitate-induced apoptosis can occur through a ceramide-independent pathway. J. Biol. Chem. 276, 14890–14895 38 Mallov, S. (1983) Role of calcium and free fatty acids in epinephrine-induced myocardial necrosis. Toxicol. Appl. Pharmacol. 71, 280–287 39 Barger, P. M. and Kelly, D. P. (2000) PPAR signaling in the control of cardiac energy metabolism. Trends Cardiovasc. Med. 10, 238–245 40 Latruffe, N. and Vamecq, J. (1997) Peroxisome proliferators and peroxisome proliferator activated receptors (PPARs) as regulators of lipid metabolism. Biochimie 79, 81–94 41 Taegtmeyer, H. (2000) Genetics of energetics: transcriptional responses in cardiac metabolism. Ann. Biomed. Eng. 28, 871–876 42 Stanley, W. C., Lopaschuk, G. D. and McCormack, J. G. (1997) Regulation of energy substrate metabolism in the diabetic heart. Cardiovasc. Res. 34, 25–33 43 Lopaschuk, G. D. and Tsang, H. (1987) Metabolism of palmitate in isolated working hearts from spontaneously diabetic “BB" Wistar rats. Circ. Res. 61, 853–858 44 Hall, J. L., Stanley, W. C., Lopaschuk, G. D., Wisneski, J. A., Pizzurro, R. D., Hamilton, C. D. and McCormack, J. G. (1996) Impaired pyruvate oxidation but normal glucose uptake in diabetic pig heart during dobutamine-induced work. Am. J. Physiol. 271, H2320–H2329 45 Lopaschuk, G. D. and Russell, J. C. (1991) Myocardial function and energy substrate metabolism in the insulin-resistant JCR: LA corpulent rat. J. Appl. Physiol. 71, 1302–1308