Survey
* Your assessment is very important for improving the work of artificial intelligence, which forms the content of this project
Vectors in gene therapy wikipedia , lookup
Gene therapy of the human retina wikipedia , lookup
Polycomb Group Proteins and Cancer wikipedia , lookup
Epigenetics in stem-cell differentiation wikipedia , lookup
Site-specific recombinase technology wikipedia , lookup
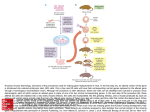
This material is protected by U.S. Copyright law. Unauthorized reproduction is prohibited. For reprints contact: [email protected] Stem Cells ® Concise Review Technical Assessment of the First 20 Years of Research Using Mouse Embryonic Stem Cell Lines Gregory J. Downing,a James F. Battey Jr.b a Office of Technology and Industrial Relations, National Cancer Institute, and bNational Institute on Deafness and Other Communication Disorders, National Institutes of Health, Bethesda, Maryland, USA Key Words. Mouse embryonic stem cell • Homologous recombination • Discovery • Technology Abstract This review assesses the effect that mouse embryonic stem (ES) cells have had on biomedical research during the 20 years that followed their isolation in 1981. Notable scientific discoveries enabled by these cell lines—including insights into cell cycle regulation, spatial and temporal relationships during development, and the roles of transcription factors and homeobox genes in developmental pathways—are discussed. The acceleration of basic discovery of gene function and the genetic basis of disease using a breakthrough technology (homologous recombination between modified gene constructs and the ES cell genome) became the principal enabling method to establish transgenic laboratory animals with single targeted genetic change. This review also examines the widespread influence of mouse ES cells as an enabling technology by highlighting their effect on drug development paradigms, directed differentiation to treat specific diseases, nuclear transfer protocols used in cloning, and establishment of methodologies for isolating non-rodent ES cells. This review concludes with a brief analysis of the most influential mouse ES cell lines of the first 20 years as viewed within the twin contexts of human disease application and contributions to the primary literature. Stem Cells 2004;22:1168–1180 Introduction These teratocarcinoma cell lines shared many morphological, biochemical, and immunological properties with pluripotent ES cells but had undergone transformation and karyotypic changes prior to establishment as cultured cell lines. Concurrently, researchers such as Brinster et al. [8–10] and Watanabe et al. [11] established the basic concepts for using chimeras with a pluripotent cell line to introduce modifications to the mouse genome, thus guiding early experiments with mouse ES cells in the following decade. In 1981, building on experience with the teratocarcinoma cell lines and with the knowledge that early postimplantation embryos transferred to ectopic sites gave rise to In the 1970s, scientists searched for a cell culture system that could serve as a platform to conduct genetic studies focusing on early embryonic development. Although it had been established that post-implantation mouse embryos contained pluripotent cells, early attempts to culture these cells in vitro proved unsuccessful [1]. Prior to 1981, in vitro model systems to study the developing embryo used cell lines, such as F9 and P10, derived from teratocarcinomas in vivo [2]. In particular, Stevens [3–7] used these mouse embryonal carcinoma (EC) cell lines to establish many of the techniques that would later be modified to isolate embryonic stem (ES) cells. Correspondence: Gregory Downing, D.O., Ph.D., Director, Office of Technology and Industrial Relations, National Cancer Institute, National Institutes of Health, Building 31, Room 10A-52, MSC 2580, 31 Center Drive, Bethesda, MD 20892-2580 USA. Telephone: 301-496-1550; Fax: 301-496-7807; e-mail: [email protected] Received May 3 2004; accepted for publication July 16, 2004. ©AlphaMed Press 1066-5099/2004/$12.00/0 doi: 10.1634/stemcells.2004-0101 STEM CELLS 2004;22:1168–1180 www.StemCells.com Downing, Battey teratomas containing pluripotent stem cells [3, 12], two groups of scientists successfully isolated and cultured ES cells from the mouse. Two simple yet elegant studies appeared that year that described the isolation and growth in culture of proliferative cells derived from mouse embryos [13, 14]. These cells were among the first to be shown to grow in an undifferentiated state for long periods of time and to be capable of differentiating into multiple cell types. With this auspicious start, mouse ES cells soon became an indispensable tool for discovery in biomedical research. There is no doubt that these cell lines have continued to represent an important research tool for many areas of biomedical science. In the two decades since Martin [13] and Evans and Kaufman [14] published their novel methods, more than 1,200 scientific publications have included mouse ES cells as a research emphasis. The era of molecular biology and recombinant DNA technology was in its infancy in 1981, making the discovery and application of this tool all the more remarkable. In the late 1970s and early 1980s, there were no genomic databases, polymerase chain reaction (PCR) techniques were only a concept, the application of homologous recombination to gene-knockout technology had yet to be proven, and applications of monoclonal antibody technology in cell biology were just emerging. As such, the discoveries enabled by mouse ES cells were remarkable from a historical perspective. Among the unique aspects of the history of this biomedical research tool is that there is a clear lineage of the source of the technology. In the 1980s, two strains of mice—129 and C57BL/6—provided the most stable sources of ES cells. The 1169 pace of discovery grew slowly in the first few years, followed by a rapid period of publication growth in the early 1990s (Fig. 1, Appendix) that some have attributed to a Cold Spring Harbor symposium and the use of the cell lines in homologous recombination, events that occurred several years earlier [15–20]. This article reviews the groundbreaking research capabilities and technologies enabled by mouse ES cells in the previous two decades. From homologous recombination to insight about cell cycle regulation, pathways for signal transduction and embryonic development, and spatial and temporal relationships during development, mouse ES cells have become a fundamental discovery tool in biomedical research. Many of the breakthroughs that will be discussed here are highlighted in Table 1. While mouse ES cells continue to inform basic research discoveries, their true impact is felt through their far-reaching application as an enabling technology. Mouse ES cells have influenced paradigms and methodologies for a variety of disciplines during the previous two decades, providing researchers with a tool that allows them to view and conduct their experiments using approaches unavailable in 1980. We highlight the part that mouse ES cells have played in breakthroughs with the potential to shape the future of medicine, including drug development paradigms, directed differentiation to treat specific diseases, nuclear transfer protocols used in cloning, and the establishment of methodologies for the isolation of non-rodent ES cells. Remarkably, the wealth of information enabled by mouse ES cells has largely resulted from work using a few key cell lines. This article concludes Figure 1. Number of publications per year that cite the use of mouse embryonic stem cell or germ cell lines. Details of search criteria are provided in the Appendix. Research Using Mouse Embryonic Stem Cell Lines 1170 Table 1. Twenty years of milestones in the application of mouse ES cells as a discovery tool Year Milestone References 1981 Establishment of mouse ES cell lines 1985 ES cells shown capable of high level of organized development 68 1987 ES cells allow targeted mutagenesis via homologous recombination 17, 20 1988 Leukemia inhibitory factor regulates differentiation in ES cell cultures 59, 60 1989 First viable chimeric mice produced from homologous recombination 23 1990 Discovery of Oct-4 in mouse embryonal carcinoma cells 90 1998 High-throughput gene-trapping methodology established 99 2000 Directed differentiation of mouse ES cells into vascular progenitors, neurons, and pancreatic islet-like cells 74–76 13, 14 Abbreviation: ES, embryonic stem. by assessing the contributions of these lines in the twin contexts of their effect on the primary literature and the insights that they have provided on the understanding of human diseases. Mouse ES Cells Usher in the Genomic Era Although mouse ES cells have supported discovery in myriad research fields, their value was established permanently in the mid-1980s as a tool to enable targeted mutagenesis. In 1985, Oliver Smithies and colleagues [19] demonstrated gene targeting using homologous recombination in a mammalian cell line. Two years later, several groups showed that homologous recombination could produce targeted genetic changes in the mouse ES cell genome that supported an animal model for Lesch-Nyhan syndrome, a rare neurological disorder [17, 20]. While Thomas and Capecchi [17] modestly referred to their demonstration of site-directed mutagenesis in a mouse ES cell line as “a protocol [that] should be useful for targeting mutations into any gene,” this gene-targeting technology would ultimately establish homologous recombination as a means to systematically modify the mouse genome. Several years earlier, it was demonstrated that, when they were injected into mouse blastocysts, genetically altered ES cells could generate transgenic offspring [21, 22]. The application of these techniques with homologous recombination technology thus provided scientists with a controlled process to generate an unlimited variety of transgenic mice with engineered, predetermined genomes. The technology developed rapidly, and the first viable chimeric mice were reported in 1989 [23]. Inspired by contemporary breakthroughs in PCR [24, 25], the development of positive-negative double selection [26], and geneknockout techniques [27, 28], researchers were able to devise increasingly subtle and sophisticated targeted genetic changes. ES cell culture was established as the first step in creating knockout and, later, transgenic (knock-in) mice. As a result of these groundbreaking efforts, a variety of protocols has been established for the precise introduction of DNA and targeted changes into ES cells, and ES cells have had a profound effect on the engineering of the mouse genome [29, 30]. The true influence of this breakthrough on scientific research is difficult to quantify, as knockout and transgenic mice have provided insight into the etiologies of hundreds of diseases, from cancers to anxiety, from diabetes to addiction, in approximately 15 years [29, 31–35] (for full listing, see the Transgenic/Targeted Mutation Database, http://tbase.jax. org). A Medline keyword search for “knockout mouse” in publications from the previous 2 years produces more than 6,500 hits; perhaps a more useful measure of the impact of homologous recombination is its almost instantaneous widespread use and application to numerous disciplines. Within 4 years of these breakthrough articles, homologous recombination in cultured ES cells, followed by reintroduction into the mouse, provided insight into the roles of many developmental factors, including proto-oncogenes, homeobox genes, specific growth factors, and transcription factors [36–47]. By 1992, homologous recombination had outgrown its initial application to Lesch-Nyhan syndrome, and mouse ES cells had been used to develop useful animal models for cystic fibrosis [48, 49] and muscular dystrophy [50]. The seminal homologous recombination experiments in the late 1980s created null mutations in particular genes, and analysis of the corresponding phenotypes generated by these mutations provided insight into gene function. Researchers soon realized that this approach could be modified slightly by introducing a given cDNA in frame with the coding sequence of the targeting gene, thereby engineering a modi- Downing, Battey 1171 fied allele that expresses the cDNA insert in place of the endogenous gene. Knock-in mice, created using this variant of the null mutation approach, have extended capabilities to study gene expression patterns [51], to monitor the fate of cells that normally express the target gene in animals homozygous for the mutation [52], to examine the function of proteins coded by related genes from a multigene family [53, 54], and to monitor neuronal activity at the synapse [55]. However, genetic diseases in humans rarely result from null mutations. Therefore, sophisticated gene-targeting strategies that can produce point mutations and gene hypomorphs are required to more precisely mimic the genetic bases of many human diseases in mice. Directed mutagenesis via mouse ES cells has been the central component of various targeting strategies to engineer highly specific mutant alleles, as reviewed elsewhere [56, 57]. Many of these systems can introduce subtle mutations with no interference in genetic expression. Moreover, there is evidence that several of these targeted mutagenesis strategies may be amenable to conditional mutagenesis, which makes them powerful tools in situations in which the expression of the altered genes provokes embryonic lethality in an embryo or fetus homozygous for the modified gene’s zygote, or renders a complex phenotype [57, 58]. of all of these factors, with a sharp increase in CDK4-associated kinase activity. These observations suggested that the induction of differentiation converts ES cells to a CDK4dependent mode of control of G1-phase progression. Differentiation also resulted in an extended G1 phase and a slowed rate of cell division, suggesting that signals that promote cell division may inhibit differentiation. Mouse ES cells have also played a part in elucidating many of the signal transduction pathways that regulate progression through the cell cycle, such as the phosphatidylinositol 3-kinase pathway and the Ras/extracellular signal-regulated protein kinase pathway [65]. In addition, increases in production of cyclin D1 have been shown to be affected by the redox state within mouse ES cells, supporting the hypothesis that a redox cycle within the mammalian cell cycle may be the mechanistic link between the metabolic processes in early G1 and the subsequent activation of regulatory proteins that signal entry into the S phase [66]. Furthermore, mouse ES cells have proven useful for experimental confirmation of mathematical models of regulation of the G1 phase [67]. The uniqueness of the mouse ES cell cycle has allowed researchers to control the onset of differentiation, thereby providing a framework for studying the integration of a differentiation program with cell cycle regulation. Mouse ES Cells Provide Insight into Cell Cycle Regulation Pathways Configuring Spatial and Temporal Relationships during Development: A Case Study of Vasculogenesis In 1988, two groups reported simultaneously that the presence of leukemia inhibitory factor (LIF) in the growth medium allowed mouse ES cells to proliferate indefinitely in vitro in an undifferentiated state in the absence of a feeder layer [59, 60]. Upon reinjection into the blastocyst, ES cells cultured in the presence of LIF contribute to the development of the whole embryo [61]. Furthermore, withdrawal from LIF induces cultured ES cells to differentiate into endodermlike cells. While this observation provided an alternative to the use of feeder cells for ES cell culture protocols, it also promoted the use of mouse ES cells as a discovery tool for cell cycle regulation and signal transduction pathways. Proliferation in vitro without differentiation suggests that cultured mouse ES cells exhibit an unusual cell cycle and that this cycle is regulated by LIF. Based on observations of the role of E- and D-type cyclins in the promotion of G1 phase progression and G1/S transition in cycling cells [62, 63], Savatier and colleagues [64] used mouse ES cells to probe the cell cycle factors that lead to differentiation. These researchers examined the expression of E- and D-type cyclins, cyclin-dependent kinases (CDKs) 2 and 4, and several CDK inhibitors during the process of differentiation in mouse ES cells induced by the removal of LIF from the culture medium. Withdrawal of LIF resulted in the upregulation ES cells have become a powerful tool to study spatial and temporal relationships that determine cell, tissue, and organ development. This section profiles their role in understanding vasculogenesis and angiogenesis, processes that have applications to many fields, from regenerative medicine to cancer research. The ES cell lines derived in the seminal 1981 experiments of Martin [13] and Evans and Kaufman [14] spontaneously differentiated to form embryoid bodies. However, it was not until 1985 that researchers began to examine the degree to which organized development within these embryoid bodies actually paralleled that within the developing embryo. In that year, Doetschman and colleagues [68] used cell lines derived from 129 and C57BL/6 blastocysts to show that ES cells were capable of exhibiting a high level of organized development at a high frequency. From 8 to 10 days of culture, approximately half of the embryoid bodies expanded into cystic structures analogous to the visceral yolk sac of the post-implantation embryo. Approximately one third of these cystic embryoid bodies developed myocardium, and when cultured in the presence of human cord serum, 30% developed blood islands. The authors also noted that this developmental potential and consistency of expression was observed 1172 in most ES lines, making the cells a useful tool to investigate early embryogenesis. In 1992, Wang and coworkers expanded the work of Doetschman [68] and others [69] to show that a primitive vasculature forms in these embryoid bodies, establishing them as a useful model for the study of developmental blood vessel formation [70]. Not only were the blood islands lined by endothelial cells, but also the blood island matured to form connections and channels that resemble blood vessels. Thus, the ES cell culture system retained the cellular signals for both the primary differentiation of pluripotent stem cells into endothelial cells and for some of the cellular processes involved in subsequent vascular development. Based on these discoveries, ES cells became a viable tool for developing in vitro systems to investigate vasculogenesis as it occurs in embryos. However, cell types from multiple lineages were generated when using these systems, thus hampering clear understanding of the interactions between endothelial cells and other essential cells, such as mural cells, in vascular development and maintenance [71, 72]. Interested in dissecting the cellular events that lead to formation of a primitive vasculature, Hirashima and colleagues [73] developed a system to isolate endothelial cell progenitors in vitro. From mouse ES cells, they derived and isolated endothelial cell progenitors that expressed fetal liver kinase 1 (Flk1), a marker essential for vasculogenesis. In subsequent experiments, they showed that these Flk1+ cells can differentiate into both endothelial and mural cells and can reproduce the vascular organization process [74]. As a result, the Flk1+ cells function as vascular progenitor cells capable of differentiating to form mature blood vessels. Recently, similar strategies have been employed to direct differentiation of mouse ES cells into pancreatic islet-like cells [75] and neurons [76]. As with many applications of mouse ES cells, the potential achievements enabled by stem cells extend far beyond researchers’initial visions; what began initially as a quest to understand mechanisms of development thus produced an enabling methodology for tissue engineering of the vascular system. Understanding pathways in Early Embryonic Development: Homeobox Genes and Transcription Factors Genes that contain the homeobox motif, a highly conserved, 180–base pair DNA sequence, are expressed in a temporal and tissue-specific pattern during embryogenesis [77]. Using breakthroughs in homologous recombination, researchers rapidly identified the presence, and later the identity and location, of many of the homeobox genes implicated in early development [78–82]. In 1987, Harald Eistetter [80] demonstrated that several homeobox-containing DNA sequences Research Using Mouse Embryonic Stem Cell Lines were stage-specifically regulated during in vitro differentiation of mouse ES cells, prompting a flurry of research activity that would ultimately characterize numerous murine homeobox genes, including the Hox family [36, 83–85], Csx [86], Nkx2-5 [82], and Pem [87, 88]. In particular, ES cells have enabled investigators to follow the developmental outcomes of mutations in the family of Hox genes, which are expressed in defined domains along the anterior–posterior axis of the developing embryo [89]. In 1990, LeMouellic and colleagues [83] used homologous recombination to replace the murine Hox-3.1 gene with lacZ, a reporter gene for β-galactosidase, from Escherichia coli. Because lacZ expression is easily detected in situ with high sensitivity, these researchers were able to observe localized areas of β-galactosidase activity during development. Mice homozygous for the targeted Hox-3.1 allele demonstrated phenotypic characteristics, such as the adjusted placement of the rib cage, that reflected similar segmentary transformations in Drosophila associated with loss-of-function homeotic mutations [84]. By contrast, mice deficient in Hox-1.6 exhibit profound defects in the ears, cranial nerves, and ganglia that result from a loss, rather than a homeotic transformation, of tissue [85]. In 1990, mouse EC cells enabled the identification of a novel DNA octamer-binding transcription factor, Oct-4 (also called Oct-3 and Oct 3/4), widely studied as a prototypical marker of undifferentiated, dividing cells [90]. A transcription factor capable of activating or repressing gene expression, Oct-4 is necessary for maintaining cells in an undifferentiated state. Ten years after this key discovery, Niwa and colleagues [91] shifted previous paradigms of binary transcriptional control by demonstrating that the precise level of Oct-4, rather than its mere presence, governs the fate of ES cells. In their groundbreaking experiments, these researchers showed that a less than twofold increase in Oct-4 expression caused cells to differentiate into primitive endoderm and mesoderm, while repression of Oct-4 induced loss of pluripotency and dedifferentiation into trophectoderm. As such, the transcription factor has been termed a “master regulator” of pluripotency. Oct-4 has been shown to regulate at least seven genes, activating some and repressing others. It has been postulated that the transcription factor exerts its overall impact by preventing the expression of genes that are required for differentiation [92]. Investigation of the role of Oct-4 in mouse ES cells has prompted researchers to speculate that it may have a similar role in human and primate development, as suggested by its presence in human [93] and primate [94] ES cells. However, complexities in cell culture conditions have meant that the vast majority of information about Oct-4 has been made possible by the study of mouse ES cells in vitro. Downing, Battey Mouse ES cells have also recently enabled the identification of the homeobox gene, nanog, a unique pluripotencysustaining factor in ES cells [95, 96]. Nanog mRNA is present in pluripotent mouse and human ES cells yet absent in differentiated cells. The nanog gene is capable of maintaining mouse ES cell self-renewal independent of the LIF/Stat3 pathway associated with transcription factors such as Oct-4, Sox2, and FoxD3. Elevated nanog expression from transgene constructs is sufficient for clonal expansion of ES cells, and nanog-deficient ES cells differentiate into extraembryonic endoderm lineage. Understanding of the molecular mechanisms that govern pluripotency is a key step toward generating pluripotent cells from somatic stem and other cells. The identification and characterization of nanog demonstrates the central role of mouse ES cells as an enabling technology to understand the pathways that govern early embryonic development. The Widespread Effect of Technologies Enabled by Mouse ES Cells The true influence of mouse ES cells can be seen in their widespread application as an enabling technology. In both subtle and overt ways, mouse ES cells have provided researchers with a tool that allows them to view their experiments through a set of lenses unavailable in 1980. Mouse ES cells have influenced paradigms and methodologies for a diverse set of applications during their first 20 years, leading to refinements in existing protocols and offering the basis for breakthroughs with wide future application. This section highlights the effect of mouse ES cells on areas that will shape the future of medicine, including drug development, directed differentiation to treat specific diseases, nuclear transfer protocols used in cloning, and the establishment of methodologies for the isolation of non-rodent ES cells. Discovering Molecular Targets and Drug Development Paradigms In the post-genomic era, strategies that use homologous recombination to target single genes have become augmented by a battery of gene-trapping approaches with the potential for probing the entire genome simultaneously [97]. When inserted into a gene, these gene-trap vectors generate fusion RNA transcripts that facilitate identification of the trapped endogenous gene from each clone of interest by sequencing. Gene trapping is therefore advantageous in mammalian cells with complex genomic organization, and the technique provides a convenient method for tagged random mutagenesis in mice [98]. Gene-trap vectors can be introduced into mouse ES cells by retroviral integration [99] or by physical methods such as electroporation and transfection [100]. The technique provides great promise for the 1173 large-scale screening efforts associated with drug discovery and development. For example, one recent report describes a high-throughput gene-trapping method that allows the automated identification of sequence tags from the mutated genes [99]. Using this approach, researchers created a library of mouse ES cells that contains sequence-tagged mutations in 2,000 genes. Genetically engineered mice have become an enabling technology in the private sector, spurring the processes of drug discovery and development. High-throughput construction and evaluation of genetically engineered mice have become standard practices for investigating molecular mechanisms of disease, evaluating prospective therapeutic targets, and screening candidate agents for efficacy and safety [101]. For example, a drug thought to work by modifying the activity of a specific protein should behave differently in a mouse model where the gene encoding the specific protein is targeted. In this manner, scientists can determine if a drug is working as predicted or by modulating a different cellular pathway. Looking forward, phenotypes of genetically engineered mice may reveal potential target organs and uncover toxic effects of potential therapies. Furthermore, constructs in which the engineered gene does not result in significant therapeutic potential may provide insight into molecular mechanisms and signaling pathways with numerous other applications. The widespread use of knockout mice has shifted paradigms in drug development, enabling a supporting biotechnology industry in the process. A recent retrospective evaluation of the 100 best-selling drugs demonstrates that knockout phenotypes for the drugs’ targets correlate well with known drug efficacy, thus supporting continued use of the technology as a tool for target discovery and validation [102]. For example, specific inhibitors of cyclooxygenase-2 (COX2), such as celecoxib and rofecoxib, have achieved high sales as treatments for arthritis. The role of COX2 in the inflammatory process has been elucidated in part from extensive study of Cox2 knockout mice, which exhibit reduced inflammation and a significant reduction in collagen-induced arthritis. The observation that Cox2 knockout mice also have decreased polyp formation has suggested additional uses for currently marketed COX2 inhibitors in areas such as cancer. Furthermore, new molecular targets for drug discovery can be identified by systematically mutating genes and determining the phenotypic consequences, such as obesity, arthritis, and cardiovascular disease. The sequencing of the human genome will provide the pharmaceutical industry with a plethora of potential new drug targets, thereby encouraging the development of large-scale mouse knockout or genetrapping programs. Questions about the utility of the knockout mouse have now been supplanted by concerns related to 1174 scale such as cost, labor intensity, and timeliness. As the 21st century unfolds, more demands will be made on highthroughput platforms and screening processes, and mouse ES cells therefore will influence another level of biotechnology development. Directed Differentiation for Specific Disease Applications The ability to induce stable genetic modifications in ES cells offers the potential to direct the course of differentiation in vivo through a series of in vitro manipulations. Human ES cells may one day serve as a source of specialized cells useful in numerous regenerative medicine applications. One of the most widely studied examples of directed differentiation, the formation in vivo of putative dopaminergic and serotonergic neurons from mouse ES cells, has great potential for the understanding and treatment of neurodegenerative diseases such as Parkinson’s disease [103]. Breakthroughs with neuronal differentiation in vivo have largely been realized within the last decade, and advances could not have been possible without the wealth of basic research on the culture and modification of mouse ES cells in vitro. Using established techniques, researchers have shown that, when they are transplanted into the mouse brain, mouse ES cells can be induced to differentiate into dopaminergic and serotonergic neurons [104]. Advances in cell culture techniques have also provided efficient methods for generating dopaminergic and serotonergic neurons in high yields from mouse ES cells in vitro [76, 105]. Dopaminergic neurons produced from mouse ES cells share the electrophysiological and behavioral properties expected of neurons from the midbrain [106], and transplantation of ES cell–derived dopaminergic neurons has been shown to partially correct the phenotype of a murine model of Parkinson’s disease [107]. Techniques to induce efficient neuronal differentiation of mouse ES cells have recently been extended to derive dopaminergic neurons from primate ES cells [108]. While the application of these breakthroughs to the treatment of motor disorders in humans has yet to be realized, mouse ES cells are pivotal tools that enable researchers to investigate factors that govern differentiation and to apply this knowledge to putative models of human disease. Somatic Cell Nuclear Transfer Protocols and Cloning Recent improvements in somatic cell nuclear transfer (SCNT) methodologies have suggested numerous potential applications for cloning in basic research, medicine, and agriculture [109]. Although currently a relatively inefficient process, nuclear transfer using embryo-derived nuclei [110] Research Using Mouse Embryonic Stem Cell Lines and fetal and adult cell nuclei [111] has produced viable sheep, including the well-known “Dolly.” SCNT has been used successfully in the genetic modification of cattle [112], mice [113], goats [114], and pigs [115], suggesting that, once optimized, the technique may have a central role in research on disease models, xenotransplantation, and the genetic engineering of livestock. The technique has recently been used with homologous recombination to create the first nonrodent knockout animal, a sheep with deletion of the prion protein (PrP) gene [116]. While this technology does not require mouse ES cells per se, the results of gene-modification strategies using mouse ES cells are a clear motivation for these techniques in mammals. Understanding of developmental processes in mouse ES cells provides a guidepost to researchers as they develop state-of-the-art SCNT methodologies. Mouse ES cells are commonly used as a comparative tool in SCNT protocols to assess the characteristics of the newly modified cells. For example, in their seminal papers on the creation of cloned sheep, Campbell and colleagues [110] often describe their embryonic cells in terms of the morphology and expression of markers of differentiation present in mouse ES cells at complementary stages of development. When Campbell suggests that somatic cell nuclear transfer will offer opportunities for genetic manipulation of livestock analogous to gene modification in knockout mice, it is evident that mouse ES cells have not only enabled a developing technology but also inspired a vision for the technology’s future direction and potential. Furthermore, novel mouse ES cell lines have been created via nuclear transfer from adult mouse somatic cells from inbred, hybrid, and mutant strains [105]. These ES cells can be induced to differentiate into numerous cell types, including dopaminergic and serotonergic neurons in vitro and germ cells in vivo. Non-Mouse Stem Cells Although mouse ES cells offer a versatile tool to study mechanisms that control differentiation, significant differences between early human and early mouse development suggest that other models may be more appropriate for the study of human development and the application to human disease. By modifying cell culture techniques developed for mouse ES cells, Thomson and colleagues [117] successfully isolated the first primate ES cell line from rhesus monkeys in 1995. Because of their similarity to humans, rhesus monkeys may provide an appropriate model to investigate strategies to prevent immune rejection of transplanted cells and to demonstrate the safety and efficacy of ES cell–based therapies [118]. Just as studies of mouse teratocarcinoma-derived stem cells preceded the isolation and characterization of mouse Downing, Battey 1175 ES cells, studies of human teratocarcinoma-derived stem cells preceded the isolation of human ES cells. Definition of human teratocarcinoma stem cell morphology and expression of the stage-specific embryonic antigens SSEA-3 and SSEA-4 (but not SSEA-1) [119–121] prepared the way for Thomson and coworkers to isolate human ES cells from human blastocysts in 1998 [118]. These cells demonstrated the ability to form trophoblasts and derivatives of all three embryonic germ layers in teratomas after injection into immunodeficient mice. Ethical and practical concerns are raised by experiments that employ human ES cell lines, thus suggesting that some future applications of primate and human ES cells will be preceded by mouse models and complementary experiments that use mouse ES cells. From One to Many: The Trajectory of Mouse ES Cell Lines in the First Two Decades In the early years of mouse ES cell research, a substantial body of knowledge was obtained using relatively few cell lines. Between 1981 and 2001, approximately two thirds of published mouse ES cell original research articles feature combinations of six key cell lines (Table 2), and subclones and modifications of these lines continue to spawn new cell lines and research directions. From their early application to Lesch-Nyhan syndrome, these seminal cell lines have contributed to the creation of transgenic animal models of many human diseases, including osteoporosis, Huntington’s disease, and myeloid leukemia. In addition, transgenic mice derived using ES cells have provided insight into complex human disorders from diabetes to cystic fibrosis. The dynamic, expanding application of mouse ES cells as a discovery tool is reflected in the steady annual increase in the number of publications that feature mouse ES cells (Fig. 1, Appendix). Although novel mouse ES cell lines continue to be created, cell lines developed in the 1980s, such as CCE, D3, and E14, have remained useful. In a sense, these cell lines are the engines that have driven the first two decades of mouse ES cell research. It must be noted, however, that these lines were created by modifying techniques and conditions established for murine EC cell lines such as F9 and P19. The varied and continual subcloning of these fundamental lines has resulted in a traceable lineage for each mouse ES cell line. A diagram showing selected subclones that have been developed from the E14 line is provided in Figure 2. While mouse ES cells have contributed to the development of transgenic animal models of human disease, they cannot be linked directly to the development of a well-known drug or diagnostic tool. However, ES cell–based breakthroughs, such as homologous recombination, underscore many patented technologies, including disease-specific mouse ES cell lines, gene-trapping techniques, and highthroughput platforms. In the two decades since they were first created, murine ES cell lines have proved to be a versatile enabling tool, and trends in the literature suggest that new applications of this technology continue to proliferate. As the 21st century unfolds, it is clear that emerging stem cell– based technologies will continue to create new breakthroughs and establish new milestones in biomedical research and application. Conclusion The isolation of murine ES cell lines in 1981 enabled a plethora of research opportunities. From gene targeting by homologous recombination to new insights on cell cycle reg- Table 2. Publications and disease insights provided by mouse ES cell lines Cell line Year established [reference] F9a 1973 [122] 77 1982 [123] 51 P19 a No. of featured publications (1981–2001) Key transgenic animal models / human disease applications [reference] CCE 1981 [14] 147 Lesch-Nyhan syndrome [18], Hemophilia B [124], Myeloid leukemia [125] D3 1985 [68] 279 Muscular dystrophy [50], Human B0 thalassemia [126], Inflammatory bowel disorders [127] E14 lines 1987 [128] 195 Lesch-Nyhan syndrome [128], Cystic fibrosis [48] AB lines 1990 [40] 76 Osteoporosis [44], Hypercholesterolemia [129], Citrullinemia [130] J1 1992 [131] 75 Tay-Sachs disease [132], Sandhoff disease [132] R1 1993 [133] 129 Diabetes [134], Rickets [135], Paroxysmal nocturnal hemoglobinuria [136] a Embryonal carcinoma cell lines. 1176 Research Using Mouse Embryonic Stem Cell Lines Figure 2. Selected subclones and cell lines derived from E14. Key cell lines such as E14 have produced numerous embryonic stem cell lines cited in the literature. ulation and development, these ES cells provided researchers with the toolkit needed to better understand the genetic basis of many diseases. This review demonstrates the diverse influence of mouse ES cells on molecularly based therapeutic development, cloning, directed differentiation, and the establishment of mammalian ES cell lines. Moreover, the understandings of the pluripotent and proliferative nature of these cells provide the foundations for pioneering work with their human counterparts, including the overall frontier of stem cell biology that aims to regenerate and restore form and function to patients. As applications of ES cell biology progress from a discovery tool to applied technologies for medicine, research with mouse ES cells continues to enable many extraordinary achievements. Acknowledgments The authors thank Charles A. Goldthwaite Jr. for his assistance in preparing this manuscript. The database referenced here is available as a searchable Web-based resource on the NIH stem cell Website at http://stemcells.nih.gov/research/ mouselit/. Appendix Criteria for the Selection of Scientific Publications That Feature Mouse Embryonic Stem Cell Lines A PubMed search of the scientific literature from 1981 to 2001 was conducted using the keyword “mouse embryonic stem cells” and yielded over 2,700 citations. Although the literature includes publications that feature cell lines developed from other species (e.g., rat, hamster, cow, pig), our search focused only on cell lines developed from mice because of the ubiquitous nature in which they are used. Publications were collected and reviewed to determine if they met the entry criteria for the analysis. There were 1,338 original papers deemed acceptable for review. Thus, more than 1,400 review papers, methods papers, publications describing work on isolated cell preparations (not cultures), and incorrectly coded PubMed citations were excluded. Thirty other eligible publications were identified that were not captured via the key words in the PubMed search, and these were included in the analysis. References 1 Sherman MI. The culture of cells derived from mouse blastocysts. Cell 1975;5:343–349. 2 Martin GR. Teratocarcinomas and mammalian embryogenesis. Science 1980;209:768–776. 3 Stevens LC. The development of transplantable teratocarcinomas from intratesticular grafts of pre- and postimplantation mouse embryos. Dev Biol 1970;21:364–382. 4 Stevens LC. A new inbred subline of mice (129-terSv) with a high incidence of spontaneous congenital testicular teratomas. J Natl Cancer Inst 1973;50:235–242. 5 Stevens LC. Animal model of human disease: benign cystic and malignant ovarian teratoma. Am J Pathol 1976;85: 809–813. 6 Stevens LC, Varnum DS, Eicher EM. Viable chimeras produced from normal and parthenogenetic mouse embryos. Nature 1977;269:515–517. 7 Stevens LC. Totipotent cells of parthenogenetic origin in a chimaeric mouse. Nature 1978;276:266–267. 8 Moustafa LA, Brinster RL. Induced chimaerism by transplanting embryonic cells into mouse blastocysts. J Exp Zool 1972;181:193–201. Downing, Battey 9 Brinster RL. The effect of cells transferred into the mouse blastocyst on subsequent development. J Exp Med 1974; 140:1049–1056. 10 Brinster RL. Participation of teratocarcinoma cells in mouse embryo development. Cancer Res 1976;36:3412– 3414. 11 Watanabe T, Dewey MJ, Mintz B. Teratocarcinoma cells as vehicles for introducing specific mutant mitochondrial genes into mice. Proc Natl Acad Sci U S A 1978;75:5113– 5117. 12 Solter D, Skreb N, Damjanov I. Extrauterine growth of mouse egg-cylinders results in malignant teratoma. Nature 1970;227:503–504. 13 Martin GR. Isolation of a pluripotent cell line from early mouse embryos cultured in medium conditioned by teratocarcinoma stem cells. Proc Natl Acad Sci U S A 1981;78: 7634–7638. 14 Evans MJ, Kaufman MH. Establishment in culture of pluripotential cells from mouse embryos. Nature 1981;292: 154–156. 15 Hogan B, Constantini F, Lacy E. Manipulating the Mouse Embryo: A Laboratory Manual. Cold Spring Harbor, NY: Cold Spring Harbor Laboratory Press, 1986. 16 Lovell-Badge RH, Bygrave AE, Bradley A et al. Transformation of embryonic stem cells with the human type-II collagen gene and its expression in chimeric mice. Cold Spring Harb Symp Quant Biol 1985;50:707–711. 17 Thomas KR, Capecchi MR. Site-directed mutagenesis by gene targeting in mouse embryo-derived stem cells. Cell 1987;51:503–512. 18 Kuehn MR, Bradley A, Robertson EJ et al. A potential animal model for Lesch-Nyhan syndrome through introduction of HPRT mutations into mice. Nature 1987;326:295– 298. 19 Smithies O, Gregg RG, Boggs SS et al. Insertion of DNA sequences into the human chromosomal beta-globin locus by homologous recombination. Nature 1985;317:230–234. 20 Doetschman T, Gregg RG, Maeda N et al. Targetted correction of a mutant HPRT gene in mouse embryonic stem cells. Nature 1987;330:576–578. 21 Gossler A, Doetschman T, Korn R et al. Transgenesis by means of blastocyst-derived embryonic stem cell lines. Proc Natl Acad Sci U S A 1986;83:9065–9069. 22 Robertson E, Bradley A, Kuehn M et al. Germ-line transmission of genes introduced into cultured pluripotential cells by retroviral vector. Nature (London) 1986;323:445– 448. 23 Thompson S, Clarke AR, Pow AM et al. Germ line transmission and expression of a corrected HPRT gene produced by gene targeting in embryonic stem cells. Cell 1989;56:313– 321. 24 Saiki RK, Scharf S, Faloona F et al. Enzymatic amplification of β-globin genomic sequences and restriction site analysis for diagnosis of sickle-cell anemia. Science 1985; 230:1350–1354. 1177 25 Erlich HA, Gelfand DH, Saiki RK. Specific DNA amplification. Nature (London) 1988;331:461–462. 26 Mansour SL, Thomas KR, Capecchi MR. Disruption of the proto-oncogene int-2 in mouse embryo-derived stem cells: a general strategy for targeting mutations to non-selectable genes. Nature (London) 1988;336:348–352. 27 Jasin M, Berg P. Homologous integration in mammalian cells without target gene selection. Genes Develop 1988;2: 1353–1363. 28 Sedivy JM, Sharp PA. Positive genetic selection for gene disruption in mammalian cells by homologous recombination. Proc Natl Acad Sci U S A 1989;86:227–231. 29 Babinet C, Cohen-Tannoudji M. Genome engineering via homologous recombination in mouse embryonic stem (ES) cells: an amazingly versatile tool for the study of mammalian biology. An Acad Bras Cienc 2001;73:365–383. 30 Torres M. The use of embryonic stem cells for the genetic manipulation of the mouse. Curr Top Dev Biol 1998;36: 99–114. 31 Muller MB, Keck ME. Genetically engineered mice for studies of stress-related clinical conditions. J Psychiatr Res 2002;36:53–76. 32 Fan J, Watanabe T. Inflammatory reactions in the pathogenesis of atherosclerosis. J Atheroscler Thromb 2003;10:63– 71. 33 Holschneider DP, Chen K, Seif I et al. Biochemical, behavioral, physiologic, and neurodevelopmental changes in mice deficient in monoamine oxidase A or B. Brain Res Bull 2001;56:453–462. 34 Harada N, Tamai Y, Ishikawa T et al. Intestinal polyposis in mice with a dominant stable mutation of the beta-catenin gene. EMBO J 1999;18:5931–5942. 35 Kulkarni RN, Bruning JC, Winnay JN et al. Tissue-specific knockout of the insulin receptor in pancreatic beta cells creates an insulin secretory defect similar to that in type 2 diabetes. Cell 1999;96:329–339. 36 Chisaka O, Capecchi MR. Regionally restricted developmental defects resulting from targeted disruption of the mouse homeobox gene hox-1.5. Nature 1991;350:473– 479. 37 DeChiara TM, Efstratiadis A, Robertson EJ. A growth-deficiency phenotype in heterozygous mice carrying an insulinlike growth factor II gene disrupted by targeting. Nature 1990;345:78–80. 38 Joyner AL, Herrup K, Auerbach BA et al. Subtle cerebellar phenotype in mice homozygous for a targeted deletion of the En-2 homeobox. Science 1991;251:1239–1243. 39 Koller BH, Marrack P, Kappler JW et al. Normal development of mice deficient in B2M, MHC class I proteins, and CD8+ T cells. Science 1990;248:1227–1229. 40 McMahon AP, Bradley A. The wnt-1(int-1) proto-oncogene is required for development of a large region of the mouse brain. Cell 1990;62:1073–1085. 41 Mucenski ML, McLain K, Kier AB et al. A functional c-myb gene is required for normal murine fetal hepatic hematopoiesis. Cell 1991;65:677–689. 1178 42 Pevny L, Simon MC, Robertson E et al. Erythroid differentiation in chimaeric mice blocked by a targeted mutation in the gene for transcription factor GATA-1. Nature 1991;349: 257–260. 43 Schwartzberg PL, Stall AM, Hardin JD et al. Mice homozygous for the ablm1 mutation show poor viability and depletion of selected B and T cell populations. Cell 1991;65: 1165–1175. 44 Soriano P, Montgomery C, Geske R et al. Targeted disruption of the c-src proto-oncogene leads to osteoporosis in mice. Cell 1991;64:693–702. 45 Tybulewicz VLJ, Crawford CE, Jackson PK et al. Neonatal lethality and lymphopenia in mice with a homozygous disruption of the c-abl proto-oncogene. Cell 1991;65:1153– 1163. 46 Thomas KR, Capecchi MR. Targeted disruption of the murine int-1 proto-oncogene resulting in severe abnormalities in midbrain and cerebellar development. Nature 1990; 346:847–850. 47 Zijlstra M, Bix M, Simister NE et al. β2-microglobulin deficient mice lack CD4–8+ cytolytic T cells. Nature 1990;344: 742–746. 48 Koller BH, Kim HS, Latour AM et al. Toward an animal model of cystic fibrosis: targeted interruption of exon 10 of the cystic fibrosis transmembrane regulator gene in embryonic stem cells. Proc Natl Acad Sci U S A 1991;88:10730– 10734. 49 Ratcliff R, Evans MJ, Cuthbert AW et al. Production of a severe cystic fibrosis mutation in mice by gene targeting. Nat Genet 1993;4:35–41. 50 Rapaport D, Fuchs O, Nudel U et al. Expression of the Duchenne muscular dystrophy gene products in embryonic stem cells and their differentiated derivatives. J Biol Chem 1992;267:21289–21292. 51 Colucci-Guyon E, Portier M-M, Dunia I et al. Mice lacking vimentin develop and reproduce without an obvious phenotype. Cell 1994;79:679–694. 52 Tajbakhsh S, Rocancourt D, Buckingham M. Muscle progenitor cells failing to respond to positional cues adopt nonmyogenic fates in myf-5 null mice. Nature 1996;384:266– 270. 53 Wang Y, Jaenisch R. Myogenin can substitute for Myf5 in promoting myogenesis but less efficiently. Development 1997;124:2507–2513. 54 Suda Y, Nakabayashi J, Matsuo I et al. Functional equivalency between Otx2 and Otx1 in development of the rostral head. Development 1999;126:743–757. 55 Bozza T, McGann JP, Mombaerts P et al. In vivo imaging of neuronal activity by targeted expression of a genetically encoded probe in the mouse. Neuron 2004;42:9–21. Research Using Mouse Embryonic Stem Cell Lines 58 Gu H, Marth JD, Orban PC et al. Deletion of a DNA polymerase beta gene segment in T cells using cell type-specific gene targeting. Science 1994;265:103–106. 59 Smith AG, Heath JK, Donaldson DD et al. Inhibition of pluripotential embryonic stem cell differentiation by purified polypeptides. Nature 1988;336:688–690. 60 Williams RL, Hilton DJ, Pease S et al. Myeloid leukaemia inhibitory factor maintains the developmental potential of embryonic stem cells. Nature 1988;336:684–687. 61 Nichols J, Evans EP, Smith AG. Establishment of germline-competent embryonic stem (ES) cells using differentiation inhibiting activity. Development 1990;110:1341– 1348. 62 Sherr CJ. G1 phase progression: cycling on cue. Cell 1994; 79:551–555. 63 Weinberg RA. The retinoblastoma protein and cell cycle control. Cell 1995;81:323–330. 64 Savatier P, Lapillonne H, van Grunsven LA et al. Withdrawal of differentiation inhibitory activity/leukemia inhibitory factor upregulates D-type cyclins and cyclin-dependent kinase inhibitors in mouse embryonic stem cells. Oncogene 1996;12:309–322. 65 Jirmanova L,Afanassieff M, Gobert-Gosse S et al. Differential contributions of ERK and PI3-kinase to the regulation of cyclin D1 expression and to the control of the G1/S transition in mouse embryonic stem cells. Oncogene 2002;21: 5515–5528. 66 Menon SG, Sarsour EH, Spitz DR et al. Redox regulation of the G1 to S phase transition in the mouse embryo fibroblast cell cycle. Cancer Res 2003;63:2109–2117. 67 Bai S, Goodrich D, Thron CD et al. Theoretical and experimental evidence for hysteresis in cell proliferation. Cell Cycle 2003;2:46–52. 68 Doetschman TC, Eistetter H, Katz M et al. The in vitro development of blastocyst-derived embryonic stem cell lines: formation of visceral yolk sac, blood islands and myocardium. J Embryol Exp Morph 1985;87:27–45. 69 Risau W, Sariola H, Zerwes HG et al. Vasculogenesis and angiogenesis in embryonic-stem-cell-derived embryoid bodies. Development 1988;102:471–478. 70 Wang R, Clark R, Bautch VL. Embryonic stem cell-derived cystic embryoid bodies form vascular channels: an in vitro model of blood vessel development. Development 1992; 114:303–316. 71 Folkman J, D’Amore PA. Blood vessel formation: what is its molecular basis? Cell 1996;87:1153–1155. 72 Carmeliet P. Mechanisms of angiogenesis and arteriogenesis. Nature Med 2000;6:389–395. 56 Sauer B. Inducible gene targeting in mice using the Cre/lox system. Methods 1998;14:381–392. 73 Hirashima M, Ketaoka H, Nishikawa S et al. Maturation of embryonic stem cells into endothelial cells in an in vitro model of vasculogenesis. Blood 1999;93:1253–1263. 57 Cohen-Tannoudji M, Babinet C. Beyond “knock-out” mice: new perspectives for the programmed modification of the mammalian genome. Mol Hum Reprod 1998;4:929–938. 74 Yamashita J, Itoh H, Hirashima M et al. Flk1-positive cells derived from embryonic stem cells serve as vascular progenitors. Nature 2000;408:92–96. Downing, Battey 1179 75 Lumelsky N, Blondel O, Laeng P et al. Differentiation of embryonic stem cells to insulin-secreting structures similar to pancreatic islets. Science 2001;292:1389–1394. 92 Pesce M, Gross MK, Scholer HR. In line with our ancestors: Oct-4 and the mammalian germ. Bioessays 1998;20:722– 732. 76 Lee SH, Lumelsky N, Studer L et al. Efficient generation of midbrain and hindbrain neurons from mouse embryonic stem cells. Nat Biotechnol 2000;18:675–679. 93 Reubinoff BE, Pera MF, Fony CY et al. Embryonic stem cell lines from human blastocysts: somatic differentiation in vitro. Nat Biotechnol 2000;18:399–404. 77 McGinnis W, Krumlauf R. Homeobox genes and axial patterning. Cell 1992;68:283–302. 94 Mitalipov SM, Kuo HC, Hennebold JD et al. Oct-4 expression in pluripotent cells of the Rhesus monkey. Biol Reprod 2003;69:1785–1792. 78 Tsonis PA, Adamson ED. Specific expression of homeobox-containing genes during induced differentiation of embryonal carcinoma cells. Biochem Biophys Res Commun 1986;137:520–527. 79 Colberg-Poley AM, Puschel AW, Dony C et al. Post-transcriptional regulation of a murine homeobox gene transcript in F9 embryonal carcinoma cells. Differentiation 1987;35: 206–211. 80 Eistetter HR. A mouse pluripotent embryonal stem cell line stage-specifically regulates expression of homeo-box containing DNA sequences during differentiation in vitro. Eur J Cell Biol 1987;45:315–321. 81 Thomas PQ, Rathjen PD. HES-1, a novel homeobox gene expressed by murine embryonic stem cells, identifies a new class of homeobox genes. Nucleic Acids Res 1992;20:5840. 82 Lyons I, Parsons LM, Hartley L et al. Myogenic and morphogenic defects in the heart tubes of murine embryos lacking the homeo box gene Nkx2-5. Genes Dev 1995;9:1654– 1666. 83 LeMouellic H, Lallemand Y, Brulet P. Targeted replacement of the homeobox gene Hox-3.1 by the Escherichia coli lacZ in mouse chimeric embryos. Proc Natl Acad Sci U S A 1990;87:4712–4716. 84 Le Mouellic H, Lallemand Y, Brulet P. Homeosis in the mouse induced by a null mutation in the Hox-3.1 gene. Cell 1992;69:251–264. 85 Chisaka O, Musci TS, Capecchi MR. Developmental defects of the ear, cranial nerves and hindbrain resulting from targeted disruption of the mouse homeobox gene Hox1.6. Nature 1992;355:516–520. 95 Chambers I, Colby D, Robertson M et al. Functional expression cloning of nanog, a pluripotency sustaining factor in embryonic stem cells. Cell 2003;113:643–655. 96 Mitsui K, Tokuzawa Y, Itoh H et al. The homeoprotein nanog is required for maintenance of pluripotency in mouse epiblast and ES cells. Cell 2003;113:631–642. 97 Durick K, Mendlein J, Xanthopoulos KG. Hunting with traps: genome-wide strategies for gene discovery and functional analysis. Genome Res 1999;9:1019–1025. 98 Zambrowicz BP, Friedrich GA. Comprehensive mammalian genetics: history and future prospects of gene trapping in the mouse. Int J Dev Biol 1998;42:1025–1036. 99 Zambrowicz BP, Friedrich GA, Buxton EC et al. Disruption and sequence identification of 2,000 genes in mouse embryonic stem cells. Nature 1998;392:608–611. 100 Skarnes WC, Auerbach BA, Joyner AL. A gene trap approach in mouse embryonic stem cells: the lacZ reported is activated by splicing, reflects endogenous gene expression, and is mutagenic in mice. Genes Dev 1992;6:903–918. 101 Bolon B, Galbreath E. Use of genetically engineered mice in drug discovery and development: wielding Occam’s razor to prune the product portfolio. Int J Toxicol 2002;21: 55–64. 102 Zambrowicz BP, Sands AT. Knockouts model the 100 bestselling drugs: will they model the next 100? Nat Rev Drug Discov 2003;2:38–51. 103 Sasai Y. Generation of dopaminergic neurons from embryonic stem cells. J Neurol 2002;249(suppl 2):II41–II44. 86 Komuro I, Izumo S. Csx: a murine heart homeobox-containing gene specifically expressed in the developing heart. Proc Natl Acad Sci U S A 1993;90:8145–8149. 104 Deacon T, Dinsmore J, Costantini LC et al. Blastula-stage stem cells can differentiate into dopaminergic and serotonergic neurons after transplantation. Exp Neurol 1998;149: 28–41. 87 Sasaki AW, Doskow J, MacLeod CL et al. The oncofetal gene Pem encodes a homeodomain and is regulated in primordial and pre-muscle stem cells. Mech Dev 1991;34: 155–164. 105 Wakayama T, Tabar V, Rodriguez I et al. Differentiation of embryonic stem cell lines generated from adult cells by nuclear transfer. Science 2001;292:740–743. 88 Fan Y, Melhem MF, Chaillet JR. Forced expression of the homeobox-containing gene Pem blocks differentiation in embryonic cells. Dev Biol 1999;210:481–496. 106 Kim JH, Auerbach JM, Rodriguez-Gomez JA et al. Dopamine neurons derived from embryonic stem cells function in an animal model of Parkinson’s disease. Nature 2002; 418:50–56. 89 Akam M. Hox and HOM: homologous gene clusters in insects and vertebrates. Cell 1989;57:347–349. 90 Okamoto K, Okazawa H, Okuda A et al. A novel octamer binding transcription factor is differentially expressed in mouse embryonic cells. Cell 1990;60:461–472. 91 Niwa H, Miyazaki J, Smith AG. Quantitative expression of Oct-3/4 defines differentiation, dedifferentiation or selfrenewal of ES cells. Nat Genet 2000;24:372–376. 107 Barberi T, Klivenyi P, Calingasan NY et al. Neural subtype specification of fertilization and nuclear transfer embryonic stem cells and application in parkinsonian mice. Nat Biotechnol 2003;21:1200–1207. 108 Kawasaki H, Suemori H, Mizuseki K et al. Generation of dopaminergic neurons and pigmented epithelia from primate ES cells by stromal cell-derived inducing activity. Proc Natl Acad Sci U S A 2002;99:1580–1585. 1180 109 Paterson L, DeSousa P, Ritchie W et al. Application of reproductive biotechnology in animals: implications and potentials. Applications for reproductive cloning. Anim Reprod Sci 2003;79:137–143. 110 Campbell KHS, McWhir J, Ritchie WA et al. Sheep cloned by nuclear transfer from a cultured cell line. Nature 1996;380:64–66. 111 Wilmut I, Schnieke AE, McWhir J et al. Viable offspring derived from fetal and adult mammalian cells. Nature 1997;385:810–813. 112 Cibelli JB, Stice SL, Golueke PJ et al. Cloned transgenic calves produced from nonquiescent fetal fibroblasts. Science 1998;280:1256–1258. 113 Wakayama T, Perry ACF, Zuccotti M et al. Full term development of mice from enucleated oocytes injected with cumulus cell nuclei. Nature 1998;394:369–374. 114 Baguisi A, Behboodi E, Melican DT et al. Production of goats by somatic cell nuclear transfer. Nat Biotechnol 1999; 17:456–461. 115 Polejaeva IA, Chen SH, Vaught TD et al. Cloned pigs produced by nuclear transfer from adult somatic stem cells. Nature 2000;407:86–90. 116 Denning C, Burl S, Ainslie A et al. Deletion of the alpha(1,3)galactosyl transferase (GGTA1) gene and the prion protein (PrP) gene in sheep. Nat Biotechnol 2001; 19:559–562. 117 Thomson JA, Kalishman J, Golos TG et al. Isolation of a primate embryonic stem cell line. Proc Natl Acad Sci U S A 1995;92:7844–7848. 118 Thomson JA, Itskovitz-Eldor J, Shapiro SS et al. Embryonic stem cell lines derived from human blastocysts. Science 1998;282:1145–1147. 119 Shevinshky LH, Knowles BB, Damjanov I et al. Monoclonal anitbody to murine embryos defines a stage-specific embryonic antigen expressed on mouse embryos and human teratocarcinoma cells. Cell 1982;30:697–705. 120 Kannagi R, Cochran NA, Ishigami F et al. Stage-specific embryonic antigens (SSEA-3 and -4) are epitopes of a unique globo-series ganglioside isolated from human teratocarcinoma cells. EMBO J 1983;2:2355–2361. 121 Andrews PW, Goodfellow PN, Shevinshky LH et al. Cell surface antigens of a clonal human embryonal carcinoma cell line: morphologic and antigenic differentiation in culture. Int J Cancer 1982;29:523–531. 122 Bernstine EG, Hooper ML, Grandchamp S et al. Alkaline phosphatase activity in mouse teratoma. Proc Natl Acad Sci U S A 1973;70:3899–3903. 123 McBurney MW, Rogers BJ. Isolation of male embryonal carcinoma cells and their chromosome replication patterns. Dev Biol 1982;89:503–508. Research Using Mouse Embryonic Stem Cell Lines 124 Kundu RK, Sangiorgi F, Wu LY et al. Targeted inactivation of the coagulation factor IX gene causes hemophilia B in mice. Blood 1998;92:168–174. 125 Sitailo S, Sood R, Barton K et al. Forced expression of the leukemia-associated gene EVI1 in ES cells: a model for myeloid leukemia with 3q26 rearrangements. Leukemia 1999;13:1639–1645. 126 Ciavatta DJ, Ryan TM, Farmer SC et al. Mouse model for human beta zero thalassemia: targeted deletion of the mouse beta maj- and beta min-globin genes in embryonic stem cells. Proc Natl Acad Sci U S A 1995;92:9259–9263. 127 Hermiston ML, Gordon JI. Inflammatory bowel disease and adenomas in mice expressing a dominant negative N-cadherin. Science 1995;270:1203–1207. 128 Hooper ML, Hardy K, Handyside A et al. HPRT-deficient (Lesch-Nyhan) mouse embryos derived from germline colonization by cultured cells. Nature (London) 1987;326: 292–295. 129 Ishibashi S, Brown MS, Goldstein JL et al. Hypercholesterolemia in low density lipoprotein receptor knockout mice and its reversal by adenovirus-mediated gene delivery. J Clin Invest 1993;92:2883–2893. 130 Patejunas G, Bradley A, Beaudet AL et al. Generation of a mouse model for citrullinemia by targeted disruption of the argininosuccinate synthetase gene. Somat Cell Mol Genet 1994;20:55–60. 131 Li E, Bestor TH, Jaenisch R. Targeted mutation of the DNA methyltransferase gene results in embryonic lethality. Cell 1992;69:915–926. 132 Sango K,Yamanaka S, Hoffmann A et al. Mouse models of Tay-Sachs and Sandhoff diseases differ in neurologic phenotype and ganglioside metabolism. Nat Genet 1995;11: 170–176. 133 Nagy A, Rossant J, Nagy R et al. Derivation of completely cell-culture derived mice from early-passage embryonic stem cells. Proc Natl Acad Sci U S A 1993;90:8424–8428. 134 Soria B, Roche E, Berna G et al. Insulin-secreting cells derived from embryonic stem cells normalize glycemia in streptozotocin-induced diabetic mice. Diabetes 2000;9: 157–162. 135 Dardenne O, Prud’homme J, Arabian A et al. Targeted inactivation of the 25-hydroxyvitamin D(3)-1(alpha)-hydroxylase gene (CYP27B1) creates an animal model of pseudovitamin D-deficiency rickets. Endocrinology 2001;142: 3135–3141. 136 Kawagoe K, Kitamura D, Okabe M et al. Glycosylphosphatidylinositol-anchor-deficient mice: implications for clonal dominance of mutant cells in paroxysmal nocturnal hemoglobinuria. Blood 1996;87:3600–3606.