Survey
* Your assessment is very important for improving the work of artificial intelligence, which forms the content of this project
143
Effect of Calcium on the Dissociation of the Mature
Rat Heart into Individual and Paired Myocytes:
Electrical Properties of Cell Pairs
BEATRICE A. WITTENBERG, ROY L. WHITE, ROSEMARY D. GINZBERG, AND DAVID C. SPRAY
Downloaded from http://circres.ahajournals.org/ by guest on May 4, 2017
The dissociation of adult rat heart into individual, functionally intact, calcium-tolerant myocytes
requires precise manipulation of extracellular calcium levels. Dissociation of intercellular connections
is achieved by lowering extracellular calcium to micromolar levels for a short period. By imposing a
very small increment in free calcium activity (from 14 to 17 fiM) during this period, we achieve a
significant yield of functionally intact pairs of myocytes still joined at the intercalated disc. We obtain
fewer intact cells, but many of these are paired end to end. Thesefindingspermit us to describe some
structural characteristics of intercellular connections between cardiac cells and to report unambiguous measurements of electrotonic coupling and dye transfer between rat cardiac cell pairs. We find
that the strength of electrical coupling between cells isolated as pairs with intact junctional contacts is
much greater than that measured between cell pairs that have formed new junctional contacts.
(Circulation Research 1986;59:143-150)
I
SOLATION techniques developed and refined
since 1980 yield populations of functionally intact single ventricular myocytes from the adult
mammalian heart (reviewed in Wittenberg and Robinson,1 Dow et al,2 and Farmer et al3).
Electrophysiological and structural properties of
cardiac gap junctions can be established without ambiguity using isolated cardiac myocytes still joined to
their neighbors by intact gap junctions. 43 While endto-end cell pairs have been isolated from guinea pig
heart,4 the standard isolation procedure applied to rat
heart' yields a negligible number of cell pairs still
attached at the intercalated disc. Modification of the
dissociation conditions permits us to optimize the yield
of either single or paired cells.
To free intact individual cells in the heart from
neighboring cells attached by junctional and nonjunctional connections, calcium concentrations must be decreased.6 However, the isolated perfused heart is insidiously damaged by low calcium perfusion, and low
calcium perturbations must be healed to prevent cell
disruption when physiological calcium levels are reestablished after the calcium-free perfusion.7 We report
here that varying calcium concentration over a narrow
range during dissociation profoundly affects maintenance of cell-to-cell connection and calcium permeability of the sarcolemma of isolated cardiac myo-
From the Department of Physiology and Biophysics and the
Department of Neuroscience, Albert Einstein College of Medicine,
Bronx, New York.
This work was supported in part by a grant-in-aid from the New
York Heart Association (B.A.W.); National Heart, Lung, and
Blood Institute Grants HL19299 (B.A.W.) and HL33655 (R.L.W.,
B.A.W.), and a grant-in-aid from the American Heart Association
(D.C.S.).
Address for reprints: Dr. Beatrice A. Wittenberg, Department of
Physiology and Biophysics, Albert Einstein College of Medicine,
Bronx, NY 10461.
Received December 5, 1985; accepted June 17, 1986.
cytes. These findings permit us to control the
dissociation of rat heart so that a significant number of
intact myocytes are still joined to a neighboring cell at
the intercalated disc. We report some of the morphological features of gap junctions in isolated pairs of rat
cardiac cells and show that these cell pairs are strongly
coupled as measured by the passage of electrical current and fluorescent dye between them.
Previously, we have presented evidence for the occurrence of low resistance pathways between "newly
formed" cell pairs.5 We report measurements of junctional conductance between such cell pairs that were
newly joined in random orientations after separation
into single individual cells, and we compare these values with junctional conductance measured between
cells that were isolated as intact pairs.
Materials and Methods
Preparation of Isolated Heart Cells
This is the current modification of the procedure of
Wittenberg and Robinson.' The solutions were supplements of modified commercial MEM Eagle Joklik
(K.C. Biological, DMC317). HEPES-MEM contained NaCl, 117 mM; KC1, 5.7 mM; NaHCO3, 4.4
mM; KH2PO4, 1.5 mM; MgCl2, 1.7 mM; HEPES,
21.1 mM; glucose, 11.7 mM; amino acids and vitamins; L-glutamine, 2 mM, and insulin, 21 mU/ml (69
x 10-' M). The pH was adjusted to pH 7.2 with
NaOH. This solution is 292 mOsm, isosmolar with rat
serum, and in the standard procedure contained no added
calcium; measured calcium activity was 5 /iM. Resuspension medium was HEPES-MEM supplemented with
0.5% bovine serum albumin, 0.3-1.0 mM calcium
chloride, and 10 mM taurine adjusted to 292 mOsm.
The procedure consists of three main steps.
1. Low calcium perfusion: Blood washout and collagenase (selected Worthington type II) perfusion of
the adult (350-400 g) male rat heart were carried out at
32° C with HEPES-MEM gassed with 85% O 2 , 15%
144
N2. Calcium chloride 0-30 (JLM was added to HEPESMEM as specified for the experiments reported here.
2. Mechanical tissue dissociation: Collagenaseperfused tissue was subsequently shaken in resuspension medium containing creatine, collagenase, and
0.3 rnM calcium chloride. Supernatant cell suspensions were washed and resuspended in resuspension
medium.
3. Separation of intact cells: Intact cardiac cells
were enriched by centrifugation through Percoll (Pharmacia Fine Chemicals, Upsala, Sweden). About 10*
cells were suspended in 10 ml of isotonic Percoll (final
concentrations 41 % in resuspension medium) and centrifuged for 10 minutes at 34g. Intact cells were recovered from the pellet and washed and stored at 30° C in
resuspension medium.
Evaluation of Heart Cell Preparation
Downloaded from http://circres.ahajournals.org/ by guest on May 4, 2017
Analytical Methods. Free calcium ion activity was
determined with a calcium-ion-selective electrode and
a double-junction reference electrode (W. Moller,
Zurich, Switzerland). Standard curves were constructed by dilution of calcium chloride solution in distilled
deionized H2O. The electrode response was linear,
from pCa = 1 (10"' M) to pCa = 6 (10"* M) calcium
chloride. We find that if the calcium activity coefficient, y, is assumed to be equal to 1 in distilled deionized H2O, then y - 0.5 in isotonic medium. The
activity coefficient depends on the ionic composition
of the medium and on the presence of components that
specifically bind calcium. HEPES buffer alone does
not bind calcium ion. For this reason, we prefer
HEPES buffer to bicarbonate or phosphate buffer, both
of which lower calcium ion activity by binding
calcium.
Protein was determined by the method of Lowry et
al.8 ATP, creatine phosphate, and lactate were determined by enzymatic assays. 910 Reported values of
ATP, creatine phosphate, and lactate are corrected for
the fraction of cells retaining normal morphology. We
find that rounded cells in 1 mM calcium chloride contain very little ATP in our preparations. Lactate accumulation stops even under anaerobic conditions when
cells are rounded and high-energy phosphate levels are
depressed (B.A. Wittenberg, unpublished results).
Microscopy. Calcium tolerance, sarcomere length,
and the contractile response to extracellular electrical
stimulation were observed with a Nikon inverted microscope fitted with Hoffman modulation optics. Cell
counts were determined in a volume of 10^* ml with a
hemocytometer. Calcium-tolerant cells displayed clear
cross striations and were quiescent in 1 mM calcium
chloride for at least 2 hours.
For electron microscopy cell suspensions were gently pelleted 1-2 hours after dissociation. Medium was
removed and replaced with fixative (2.5% ultrapure
glutaraldehyde in 0.1 M cacodylate buffer; pH = 7.2).
Cells were fixed for 30 minutes. After a brief rinse in
buffer, the cell pellets were postfixed for 1 hour with
1% OsO4 in the same buffer. Samples were then dehydrated through a graded series of ethanols and embed-
Circulation Research
Vol 59, No 2, August 1986
ded in Epon. Thin sections of specimens were poststained with uranyl acetate and lead citrate." A Philips
300 transmission electron microscope was used to examine the sections at 60 kV.
Dye Injection. A 3% solution of Lucifer Yellow CH
(Sigma) in 150 mM LiCl was iontophoretically injected into one cell of a coupled pair of heart cells. Cells
were examined with a Nikon Diaphot microscope
equipped with a Xenon epifluorescence source and an
FTTC filter block.
Electrophysiology. To test the contractile response
of calcium-tolerant cells, platinum wire electrodes (.3
mm diameter) separated by 200 fim were dipped into
the pool of resuspension medium containing 1 mM
calcium chloride. Biphasic pulses of 3- to 8-V strength
and 3 msec duration were applied to those few cells
between the exposed ends of the electrodes at a rate of
1-2 times/second by a stimulator (Narco Biosystems,
model SI-10, Houston, Texas). Individual quiescent
cells were observed at a magnification of 300 X . Cells
that appeared to contract synchronously throughout the
cell in response to each pulse and that did not become
spontaneously active after repeated stimulation were
considered to show normal contractile response. Not
all calcium-tolerant cells contract when stimulated.
About 80% of the cells used for subsequent experiments continued to respond to stimulation for at least 2
hours in 1 mM calcium.
Newly formed pairs were prepared by plating a plastic culture dish with a layer of single cells that were
then mechanically brought in contact with other single
cells and incubated overnight. The conductances of the
junctional and nonjunctional membranes of newly
formed t-shaped or x -shaped tnyocyte pairs were
measured using the dual whole-cell voltage clamp
technique.3 This technique has the advantage of enhanced sensitivity at very small current levels. However, the uncompensated series resistance of the wholecell voltage clamps (using 5-Mft patch electrodes) is
about 1 Mil (conductance = 1 /xsiemens). When the
input conductance (the parallel sum of the junctional
and nonjunctional conductances) of a cell pair exceeds
0.1 /isiemens, measurement of conductances becomes
imprecise.
For very tightly coupled cells, we voltage clamped
one cell of the pair and current clamped the other,
using independent single electrode clamp circuits for
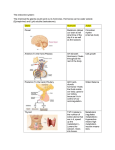
each cell (see Figure 1). The clamp circuits (Almost
Perfect Electronics, Basel, Switzerland) inject current
and measure voltage alternately, at a rate at least 10
times greater than the cell time constant (8-15 msec).
Briefly, a current pulse applied to Cell 1 permits measurement of transjunctional voltage (V^) and junctional
current (i}). Alternatively, voltage pulses applied to
Cell 2 permit the measurement of a transfer voltage in
Cell 1.
Figure 1A shows that a portion of the current pulse
applied to Cell 1 flows through the junctional resistance, rj, to Cell 2 and is there measured as ; r The
resultant voltage deflection (VH) measured in Cell 1 is
equal to V}. Thus, V, and ij are measured directly in the
Wittenberg et al
Adult Heart Cell Pairs
145
current clamp mode and g} and gt are calculated by
Equations 1 and 2:
(1)
8i = ('„
-
(2)
'J
Figure IB shows that a voltage pulse, Vn, applied to
Cell 2 by the voltage clamp results in current in flowing through ri and the nonjunctional resistances (r,, r2)
of the cell pair. A transfer voltage, V2I, is measured in
Cell 1 and the nonjunctional conductance of Cell 2 (g2)
is calculated from Equation 3:
-
A:
V2,)g,
(3)
Conventional intracellular microelectrodes were
used; resistance was 10-30 Mil for electrodes filled
with solution containing KC1, 1000 mM; EGTA 10
mM; Hepes, 2 mM. The pH was adjusted to 7.1 with
KOH.
Downloaded from http://circres.ahajournals.org/ by guest on May 4, 2017
current clamp
voltage clamp
B:
current clamp
voltage clamp
FIGURE I. Simplified schematic diagram of current flow and
voltage through a pair of coupled cells during measurement of
junctional (gj) and nonfunctional (gh g2) conductances by current pulses (A) or voltage pulses (B). The nonjunctional input
resistances of Cells I and 2 are represented by x, and r2, respectively, and Tj represents the junctional resistance between Cell 1
and Cell 2. The dotted outline is a schematic representation of a
highly magnified cell pair. The battery represents the voltage
clamp on Cell 2, and the current source represents the current
clamp on Cell I. When a pulse of current is applied to Cell I by
the current clamp (A), Vj is measured by the current clamp
voltmeter and iy is measured by voltage clamp ammeter, gj and
g/ are calculated by Equations I and 2. In order to calculate the
nonjunctional conductance in Cell 2 (g2), a voltage pulse (V22)
is applied to Cell 2 by the voltage clamp (B), and V2; and i22 are
measured. g2 is calculated by Equation 3. Currents, voltages,
and conductances are defined as follows: \n is the amplitude of
the current pulse applied to Cell 1. \j is the incremental junctional current. V, is the incremental trans junctional voltage.
V22 is the amplitude of the applied voltage pulse in Cell 2.W2I is
the amplitude of the transfer voltage pulse measured in Cell 1
when V22 is applied to Cell 2. \22 is the amplitude of the current
pulse that flows in Cell 2 in response to V22. A. Current pulse
(in) applied to Cell I by the current clamp. Junctional conductance (gj) and nonjunctional conductance in Cell 1 (g,) are
determined from Equations I and 2 (see "Materials and Methods"). B. Voltage pulse (W22) applied to Cell 2 by the voltage
clamp. Nonjunctional conductance in Cell 2 (g2) is determined
from Equation 3 (see "Materials and Methods").
Results
Effect of Calcium Concentration on the Yield
of Intact Cell Pairs and Single Cells
The results are summarized in Table 1. In the absence of added calcium during the washout step the
free calcium ion activity of the extracellular fluid drops
from about 1000 fiM. toward a value of 5 /xM, the free
calcium activity of HEPES-MEM. After perfusion
with collagenase, the measured free calcium ion activity is about 8-10 /iM.
Increasing the free calcium activity of the dissociation solution from 5 /xM (zero added calcium) to about
15 fiM (20 /JM added calcium) did not affect yields of
viable heart cells. Essentially no detectable cell pairs
joined end to end were isolated by this procedure. The
addition of 25 /xM calcium chloride (aCa = 14-17
IxM) to the HEPES-MEM, however, resulted in a visible increase in the number of end-to-end cell pairs
surviving the dissociation procedure, a visible increase
in the number of irreversibly damaged single cells, and
a significant decrease in the yield of viable cells. The
high-energy phosphate level was significantly decreased probably reflecting the presence of some metabolically challenged though rectangular and calciumtolerant cells in this population.
An increase in the calcium addition from 25 /nM to
30 ju,M calcium chloride (aCa > 20 /u,M) led to irreversible contracture of all the cells and a negligible
yield of viable cells.
Morphology and Junctional Coupling of Isolated Intact
Cell Pairs
Microscopy. Cell pairs joined at the intercalated disc
were characterized by their doubled length (250 x 25
(JM) and by a distinctly visible line of demarcation at
the contact region. Cell pairs joined side to side along
the longitudinal axis were characterized by a clear difference in lateral dimension and were bidentate at their
transverse ends.
Figure 2A shows thin-section electron micrographs
of the intercalated disc region between an isolated pair
of still joined, structurally intact myocytes. As previously reported for single isolated cells, 113 mitochondrial structure, sarcomere alignment, and cell structure
146
Circulation Research
Vol 59, No 2. August 1986
TABLE 1. A Comparison of the Yield and Metabolite Content of Heart Cells After Dissociation Using Varying Calcium
Concentrations in the Perfuswn Step
Yield
Calcium in
HEPES-MEM
(MM)
added
0
n=13
20
n=l
25
, „ ..
End-toRectangu ar Ce s c . n .
2
End Pairs
(% of
(mg (% of rec-.
total)
protein) tangular)
aCa
Purified
,-, ,,
Cells
(mg
protein)
5
39±11
78±5
30.4
14
52±16
76 + 5
14-17
33±14
61 rt 18
Metabolite Content
(nmol/mg protein
of rectangular cells)
n
ATP
CP
Lactate
0.1
31.4 + 7.2
49.1 ±9.2
73.4±32.7
39.5
0.1
28 + 4
49±5
62±17
20.1*
3-5
20* + 2
38*±15
83±59
•Values are significantly lower (p < .05, Students's t test) than the control values.
Measurements are reported as mean ± SD.
Downloaded from http://circres.ahajournals.org/ by guest on May 4, 2017
FIGURE 2. A.
Thin section of a myo-
cyte pair (magnification, 9,375 X). The
desmosome separation is increased to
80 nm, but gap junctions remain intact
(see solid arrows and box). The cell coat
is intact, and mitochondria and myofilament alignment resemble these structures in intact heart. Sarcomere length
is 1.9 fun from internal calibration assuming the A-band = 1.55 jxm.'2 The
clearly defined and aligned I-bands with
defined N-lines (see clear arrow) show
that the contractile apparatus of both
cells is relaxed. Cells were isolated
using 25 yM added calcium chloride in
the perfusate. The suspension medium
contained 0.3 mM calcium chloride.
Calibration bar is 1 ixm. B. A higher
magnification (39,000 x J of a portion of
the same junctional region indicated by
the box in Figure 2A. Here, the septalaminar structure of the gap junction is
intact (see solid arrow), while the adjacent desmosome is clearly separated.
Calibration bar is 0.1 nm.
Wittenberg et al
Adult Heart Cell Pairs
147
Downloaded from http://circres.ahajournals.org/ by guest on May 4, 2017
FIGURE 3. Thin section of a portion of the
intercalated disk between another myocyte
pair (magnification, 27,625 x). The desmosomes are more widely separated than m
Figure 2A (300-500 nm) and have an almost vacuolar appearance. Note that four
gap junctions (arrows) appear to be intact,
indicating that they may be the last junctional structure to separate in the dissociation process. Despite the appearance of the
intercalated disk r.egion, the rest of the cell
appears in excellent condition, with good
myofilament alignment and intact mitochondria. Note, however, that near the separated disk, /-bands are shorter with variable length and myofilaments are in
disorder with a splayed appearance. This
suggests a strain on the ends of the cells as
the desmosome separation increases. Calibration bar is 1 fjun.
preservation is good. The A-, Z-, and M-bands as well
as the I-bands and the N-lines are clearly resolved,
demonstrating a relaxed configuration of the sarcomeres in the presence of 0.3 mM extracellular calcium.
Sarcomere length measured in the myocyte pair shown
in Figure 2A is comparable to that which we measure
in living cells without fixation. In contrast, the appearance of the intercalated disc differs significantly from
that typically observed in the intact heart. There is a
visible, distinct separation between the cells that is not
observed in whole tissue. However, cell-to-cell contact is structurally maintained at typical gap-junction
structures. Gap junctions between cells at the intercalated disc remain intact (see Figure 2B) even when the
desmosomes have a larger than normal separation (see
Figure 3). Considerable strain is evident in the last Aand I-band regions next to the widely separated desmosome (see Figure 3).
Dye Transfer Between Cell Pairs. Figure 4 demonstrates sarcoplasmic continuity between cells still
joined at the intercalated disc. Lucifer Yellow CH injected iontophoretically into one cell of an end-to-end
pair diffused into the other cell within a few minutes,
specifically through the intercalated disc content. Dye
does not diffuse out of Cell 1 at any other region. Dye
coupling in newly formed pairs was slower and was
never seen within 5 minutes of injection, indicating
that gj was very low.
Electrical Coupling Between Isolated Intact Pairs.
.Functional conductance was measured between cell
pairs. For these measurements three different types of
pairs were examined:
1. end-to-end pairs isolated with some intact structural contact at the intercalated disc;
2. side-to-side pairs isolated with structural contact
intact along the longitudinal axis; and
3. cells first isolated as single cells and subsequently mechanically brought into contact to form randomly
joined new pairs.
Cells isolated as pairs aligned end to end or side to
side were almost always electrically coupled, and
junctional conductance (gj) was often very high. Newly formed pairs mechanically joined (as judged by coordinated motion) were rarely electrically coupled,
and when coupling was observed, gt was much lower
(see Table 2).
Figure 5 shows typical records of electrical coupling
from several pairs of cells. Figure 5A is a dual wholecell voltage clamp recording using patch electrodes
from cells which were dissociated in nominally calcium-free medium as single cells and were subsequently joined to form new low-resistance pathways after
Circulation Research
148
Downloaded from http://circres.ahajournals.org/ by guest on May 4, 2017
FIGURE 4. Upper panel: Light micrograph showing the injection ofLucifer Yellow into the left-hand member of a pair of cells
with trans- and epi-illumination. Lower panel: the same field
several minutes after injection viewed with fluorescence optics.
The injected cell fluoresces brilliantly. Transfer of fluorescent
dye through the intercalated disc into the right-hand cell is
clearly observed. Note that dye diffusion throughout the injected cell on the left appears to be faster than dye movement
through the intercalated disc. Cells were dissociated with 25
fiM CaCl2 (aCa = 17 fiM) and finally resuspended in I mM
CaCl2.
incubation overnight. Junctional currents are seen as
upward deflections in the current traces (I) of the
paired cell and represent current flowing to ground
through the junctional membrane when either Cell 1 or
Cell 2 was hyperpolarized by its respective voltage
clamp circuit. Junctional conductance was .007 /Ltsiemens, while nonjunctional conductances were .018
/usiemens (Cell 1) and .067 /asiemens (Cell 2).
Figure 5B shows traces from a tightly coupled isolated cell pair, dissociated using the modified technique with 25 fiM calcium chloride added to the
HEPES-MEM during the low calcium perfusion. The
single electrode switching clamp (see "Materials and
Methods" and Figure 1) was used. One cell was injected with current pulses (top trace), and the voltage
deflection in that cell (Vt) was recorded (second trace
from top). At the same time, current (i) was measured
in the second, voltage clamped, cell (upward deflections in the third trace from top). A voltage pulse (V^)
(bottom trace) was alternately applied to the second
cell (by its voltage clamp circuit) with resultant downward deflections in the current measured in Cell 2 (in)
TABLE 2.
Junctional Conductance Measured Between Cell Pairs
Calcium
Added to
HEPESMEM
Type of Intercellular Contact (pM)
Junctional Conductance
(Siemens x 10" 6 )
mean ± SEM
Newly formed pairs
(random orientation)
0
Laterally coupled pairs
25
1.20 ±0.40
(n = 7)*-t
End-to-end coupled pairs
25
2.53 ±0.55
(n=12)*
0.007 ±0.004 (n=10)
•Significantly different from g, measured in newly formed pairs
(p < 0.01).
tNot significantly different from g measured in end-to-end coupled cells (p > .05, Student's / test).
Vol 59, No 2, August 1986
and the voltage recorded in Cell 1 (V2I) while /,,, the
current in Cell 1, remained constant (at zero). Cell 2
was voltage clamped to —40 mV, while Cell 1 remained current clamped; but because the cells were
tightly coupled, the voltage measured in Cell 1 was
also - 4 0 mV. Junctional conductance measured in
this pair was 2.24 /xsiemens. Nonjunctional conductances were 0.107 /isiemens (Cell 1) and 0.028 /xsiemens (Cell 2).
Measurements recorded from a series of cell pairs in
several preparations are shown in Table 2. While g
varied from 0.47 to 3.10 /^Siemens in laterally coupled
pairs and from 0.60 to 8.00 ^isiemens in end-to-end
A:
B.
Newly formed
Tightly coupled
cell pairs
isolated cell pairs
I
V
11
, iomy
[lOO pA
,1 nA
EJ5 mV
1 sec
1 sec
FIGURE 5. Electrical coupling recorded between pairs of
myocytes. The dotted horizontal line separates traces recorded
from Cell 1 from traces recorded from Cell 2 in both Figure 5A
and Figure 5B. V designates voltage records: I designates
current records. A. Electrical coupling recorded between newly
formed pairs was measured using two independent whole cell
voltage clamps. Gigaohm seals to the sarcolemma of each cell
were formed using low-resistance electrodes. The membrane
enclosed by the pipette tip was ruptured using suction.5 A hyperpolarizing voltage pulse (Vj) was applied to Cell 1 (top trace),
while Cell 2 was maintained at constant voltage. Junctional
current, ij, is observed as a resultant upward deflection in the
current trace of Cell 2 (bottom trace). Cell 1 was pulsed alternately with Cell 2. Cells were in medium containing 0.3 mM
CaCl2. Junctional conductance was 0.007 fxsiemens. Nonjunctional conductances were 0.018 (Cell 1) and 0.067 ftsiemens
for Cell 2. Calibration: vertical bar, voltage = 10 mV, current
= 100 pA; horizontal bar, time = I second. B. Electrical
coupling recorded between tightly coupled heart cells isolated
as end-to-end pairs, using the voltage/current clamp method
with two single electrode clamp circuits described in Figure 1
(see "Materials and Methods"). Here, a current pulse (i,,)
applied to Cell 1 results in a voltage deflection (WJ in Cell I and
a current deflection (ij) in Cell 2 while voltage is maintained
constant in Cell 2. When a voltage pulse (V22) was applied to
Cell 2, current fijjj flowed through junctional and nonjunctional conductance pathways to produce a voltage deflection
V2; in Cell 1. Junctional conductance was2.24 ftsiemens, while
nonjunctional conductances were 0.107 fisiemens for Cell 1
and 0.028 ftsiemens for Cell 2. Calibration: vertical bar, voltage = 2.5 mV, current = 1,000 pA: horizontal bar, time = /
second.
Wittenberg et al
Adult Heart Cell Pairs
pairs, the junctional conductance of laterally coupled
cell pairs was not significantly different from that of
end-to-end pairs (p > 0.05 Student's t test). Mean gj
between all cells isolated as intact pairs was 2.05 ±
0.46 ^Siemens (n = 19; mean ± SEM), a value nearly
300 times greater than that observed between newly
formed pairs (0.007 ± .004 /^Siemens, n = 10) (see
Table 2).
Downloaded from http://circres.ahajournals.org/ by guest on May 4, 2017
Discussion
The manipulation of extracellular calcium ion activity has been crucial to the isolation of functionally
intact cardiac myocytes. U4~16 We have defined the limits over which calcium concentration can be varied
during the low calcium perfusion in the initial dissociation step.
Thin sections, displaying the sarcolemma of single
cells isolated after dissociation in nominally calciumfree medium, show that the surface coat, separated
halves of desmosomes, and remnant gap junctions are
intact. ll7 Therefore, dissociation in low calcium medium (aCa = 5-14 fiM) does not split the gap junctions
down the middle. Rather, the adjacent nonjunctional
membranes were ruptured and thereafter incorporated
with the whole gap junction into one or the other cell
through a pinocytotic mechanism.17
When aCa of the dissociation medium is increased
from 14 to 17 /iM, fewer cells survive the dissociation
process, and the occurrence of cell pairs that remain
attached at the intercalated disc increases strikingly.
We cannot tell whether these two effects are related or
independent. The sharp calcium dependence of cell
pair production may represent the lower limit of calcium required for maintenance of intact membranes
surrounding the gap junctional plaques. Cells still connected to their neighbors by weakened intercellular
connections may be more vulnerable to the shearing
forces of the dissociation procedure after calcium concentration is increased to 300 (AM. Figure 3 shows that
cells still connected by several intact gap junctions
demonstrate signs of structural strain in the erratic arrangement of myofilaments and non-uniform I-bands
on both sides of the intercalated disc when the desmosome separation increases.
The dissociation of guinea pig heart yields a higher
percentage of end-to-end paired cells,4 and the period
of increased calcium overload susceptibility is apparently much shorter16 than we observe for rat heart.
These data suggest that there may be differences in the
calcium sensitivity of the sarcolemma and of intercellular connections in different animals.
The maintenance of high-energy phosphate levels
(ATP plus creatine phosphate) provides a sensitive
index of the ability of the cells to maintain high-energy
phosphate synthesis at rates commensurate with utilization. Heart cells maintain normal ionic gradients18
and continue to maintain structural integrity, calcium
tolerance, and other criteria of viability when highenergy phosphate levels are diminished to 67% of control for several hours.I9-2U The decreased high-energy
phosphate levels measured in cells prepared in
149
HEPES-MEM with aCa = 14-17 /iM suggests that
the maintenance of ionic gradients is challenging the
metabolic regeneration of ATP in this population of
cells.
The electron micrographs shown here display separation of desmosomes but preservation of gap junctions in axially oriented cells that have survived the
dissociation procedure as pairs. These large gap junctions are apparently similar to those in intact tissue and
satisfy criteria of functional integrity; that is, they
show junctional conductance (g^ that is higher than
that measured between newly formed pairs and they
pass the fluorescent dye Lucifer Yellow CH.
Page and Shibata21 conclude that fewer gap junctions occur along the lateral borders than at the longitudinal ends of cardiac cells in the heart. Thus, one might
have expected that gt between end-to-end coupled cells
would be higher than that between laterally coupled
cells. We found no significant difference in g between
pairs of cells that were coupled end to end and cells that
were joined along their lateral edges. Kameyama4 reported measurements of gt between myocyte pairs isolated from guinea pig hearts without manipulation of
calcium levels. Our measured values of g} are 3—4
times greater than those reported by Kameyama; however, his data also show no significant difference in g}
between end-to-end and side-to-side pairs. We measured junctional current and transjunctional voltage by
voltage/current clamp techniques directly, so that calculation of gj does not become "imprecise at higher
coupling ratios."4 The magnitude of junctional conductance depends on the conductance per channel and
the proportion of existing channels that are open.22
Experimental discrepancies may reflect species differences or changes in junctional conductance induced
during the dissociation procedure.
Two separated cells that have been isolated as individual single cells can be brought into contact to form a
mechanically coupled cell pair oriented at random.
Remarkably, some of these cell pairs demonstrate conducting pathways after overnight incubation, although
gi measured between such pairs is about 300 times
lower than that recorded between cells isolated as intact pairs. The fact that conductance can be measured
between these pairs establishes the existence of lowresistance pathways between cells; the magnitude of
the measured conductances suggests relatively few
open channels between cells. In accord with this conclusion we have found that thin sections and freeze
fracture replicas of newly coupled cells reveal only
punctate oppositions and small particle aggregates
(Mazet F, Ginzberg RD, Wittenberg BA, and Spray
DC, unpublished observations).
Pairs of cardiac myocytes, joined end to end and
connected by gap junctions, have been isolated from
the adult rat heart. Gap junctions between these paired
cells are functionally similar to those in intact tissue, as
judged by the gj values recorded (Figure 5) as well as
the morphology of the intercalated disk region (Figure
2). Adult heart cells isolated as individuals may form
new low-resistance pathways when they are brought
150
Circulation Research
together in random orientation after isolation. The
magnitude of conductance measured between such
newly paired cells is much lower than that measured
between cells isolated as intact pairs.
11.
Acknowledgments
12.
We thank Dr. Thomas F. Robinson for helpful suggestions and
constructive criticism particularly concerning interpretations of the
electron micrographs, and we thank Eleanor A. Morales for fixation
and processing of the heart cells for electron microscopy. The
technical assistance of J. Zavilowitz is gratefully acknowledged.
We are particularly grateful for the constructive technical assistance of Chui Fan Wong in the preparation and analytical aspects of
this study.
References
Downloaded from http://circres.ahajournals.org/ by guest on May 4, 2017
1. Wittenberg BA. Robinson TF: Oxygen requirements, morphology, cell coat and membrane-permeability of calciumtolerant myocytes from hearts of adult rats. Cell Tissue Res
1981;216:231-251
2. Dow JW, Harding NG, Powell T: Isolated cardiac myocytes: I.
Preparation of adult myocytes and their homology with intact
tissue. Cardiovasc Res 1981 ;15:483—514
3. Farmer BB, Mancina M, Williams ES, Watanabe AM: Isolation of calcium tolerant myocytes from adult rat hearts: Review
of the literature and description of a method. Life Sciences
1983;33:1-18
4. Kameyama M: Electrical coupling between ventricular paired
cells isolated from guinea-pig heart. J Physiol 1983;336:
345-357
5. White RL, Spray DC. Campos de Carvalho AC, Wittenberg
BA, Bennett MVL: Some electrical and pharmacological properties of gap junctions between adult ventricular myocytes. Am
J Physiol 1985;249:C447-C455
6. Muir A: The effects of divalent cations on the ultra-structure of
the perfused rat heart. J Anal 1967; 101:239-261
7. Zimmerman ANE, Hulsmann WC: Paradoxical influence of
calcium ions on the permeability of the cell membranes of the
isolated rat heart. Nature 1966;211:646-647
8. Lowry OH, Rosebrough NJ, Farr AL, Randall RJ: Protein
measurement with the Folin phenol reagent. J Biol Chem
1951; 193:265-275
9. Lowry OH, Passeneau JV: A Flexible System of Enzymatic
Analysis. New York, Academic Press, 1972, pp 151-153
10. Gutmann I, Wahlefeld AW: L - ( + ) - Lactate. Determination
13.
14.
15.
16.
17.
18.
19.
20.
21.
22.
Vol 59, No 2, August 1986
with lactate dehydrogenase and NAD, in Bergmeyer HU (ed):
Methods of Enzymatic Analysis, ed # 2 , vol 3. Deerfield
Beach, Florida, Verlag Chemie International, 1974, pp
1464-1468
Venable JH, Coggeshall R: A simplified lead citrate stain for
use in electron microscopy. J Cell Biol 1965;25:407-408
Page SG: Measurement of structural parameters in cardiac
muscle, in The Physiological Basis of Starling's Law of the
Heart. Summit, N.J.. Ciba Foundation Symposium # 2 4 . pp
13-26
Severs NJ, Slade AM, Powell T, Twist VW, Warren RL:
Correlation of ultrastructure and function in calcium-tolerant
myocytes isolated from the adult rat heart. J Ultraslruct Res
1982;8I:222-239
Powell T, Terrar DA, Twist VW: Electrical properties of individual cells isolated from adult rat ventricular myocardium. J
Physiol 1980;302:13l-153
Hohl CM, Altschuld RA, Brierley GP: Effects of calcium on
the permeability of isolated adult rat heart cells to sodium. Arch
Biochem Biophys 1983;221:197-205
Bendukidge Z, Isenberg G, Klockner U: Ca-tolerant guineapig ventricular myocytes as isolated by pronase in the presence
of 250 fjiM free calcium. Basic Res Cardiol l985;80(suppl
1): 13— 17
MazetF, Wittenberg BA, Spray DC: Fate of intercellular junctions in isolated adult rat cardiac cells. Circ Res 1985;56:
195-204
Wittenberg BA, Gupta RK: NMR studies of intracellular sodium ions in mammalian cardiac myocytes. J Biol Chem 1985;
260:2031-2034
KatzIR, Wittenberg JB, Wittenberg BA: Monoamineoxidase,
an intracellular probe of oxygen pressure in isolated cardiac
myocytes. J Biol Chem 1984;259:7504-7509
Wittenberg BA, Wittenberg JB: Oxygen pressure gradients in
isolated cardiac myocytes. J Biol Chem 1985;260:6548-6554
Page E, Shibata Y: Permeable junctions between cardiac cells.
Ann Rev Physiol 1981;43:431-441
Spray DC, White RL, Mazet F, Bennett MVL: Regulation of
gap junctional conductance. Am J Physiol 1985;248: H753H764
KEY WORDS • ventricular myocytes • gap junctions • dye transfer • calcium sensitivity •
electrophysiology • gap junctional conductance
Effect of calcium on the dissociation of the mature rat heart into individual and paired
myocytes: electrical properties of cell pairs.
B A Wittenberg, R L White, R D Ginzberg and D C Spray
Downloaded from http://circres.ahajournals.org/ by guest on May 4, 2017
Circ Res. 1986;59:143-150
doi: 10.1161/01.RES.59.2.143
Circulation Research is published by the American Heart Association, 7272 Greenville Avenue, Dallas, TX 75231
Copyright © 1986 American Heart Association, Inc. All rights reserved.
Print ISSN: 0009-7330. Online ISSN: 1524-4571
The online version of this article, along with updated information and services, is located on the
World Wide Web at:
http://circres.ahajournals.org/content/59/2/143
Permissions: Requests for permissions to reproduce figures, tables, or portions of articles originally published in
Circulation Research can be obtained via RightsLink, a service of the Copyright Clearance Center, not the
Editorial Office. Once the online version of the published article for which permission is being requested is
located, click Request Permissions in the middle column of the Web page under Services. Further information
about this process is available in the Permissions and Rights Question and Answer document.
Reprints: Information about reprints can be found online at:
http://www.lww.com/reprints
Subscriptions: Information about subscribing to Circulation Research is online at:
http://circres.ahajournals.org//subscriptions/