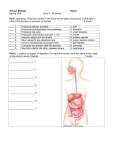
Survey
* Your assessment is very important for improving the workof artificial intelligence, which forms the content of this project
Appl Microbiol Biotechnol (2000) 53: 709±714 Ó Springer-Verlag 2000 ORIGINAL PAPER P. De Boever á R. Wouters á L. Verschaeve P. Berckmans á G. Schoeters á W. Verstraete Protective effect of the bile salt hydrolase-active Lactobacillus reuteri against bile salt cytotoxicity Received: 15 July 1999 / Received revision: 10 January 2000 / Accepted: 14 January 2000 Abstract Bacterial bile salt hydrolysis is considered a risk factor for the development of colon cancer because of the risk of forming harmful secondary bile salts after an initial deconjugation step. In this study, the in¯uence of enhanced bacterial bile salt transformation by the bile salt hydrolase-active Lactobacillus reuteri was studied in batch culture using the microbial suspension of the Simulator of the Human Intestinal Microbial Ecosystem; (SHIME), which was supplemented with oxgall at 5 g/l or 30 g/l. Changes in the fermentative capacity of the microbial ecosystem and the (geno)toxic properties of the SHIME supernatants were investigated. Increasing concentrations of oxgall inhibited the fermentation. Transient cell toxicity was observed for samples supplemented with 5 g oxgall/l, while samples with 30 g oxgall/l exhibited toxicity. The results of the haemolysis test suggest that the detrimental eects were probably due to the membrane-damaging eects of bile salts. In all cases, the adverse eects could be counteracted by the addition of 7.5 0.5 log10 CFU L. reuteri/ml. Plausible mechanisms for the protective properties of L. reuteri could involve a precipitation of the deconjugated bile salts and a physical binding of bile salts by the bacterium, thereby making the harmful bile salts less bioavailable. Introduction Colorectal cancer is the second most common cause of cancer deaths in most developed countries (Ghadirian P. De Boever á R. Wouters á W. Verstraete (&) Laboratory of Microbial Ecology and Technology, Faculty of Agricultural and Applied Biological Sciences, University Ghent, Coupure links 653, 9000 Gent, Belgium e-mail: [email protected] Tel.: +32-9-264/59/76 Fax: +32-9-264/62/48 L. Verschaeve á P. Berckmans á G. Schoeters Environmental Toxicology, Flemish Institute for Technological Research (VITO), Boeretang 200, 2400 Mol, Belgium et al. 1997). Although there have been advances in radiotherapy, chemotherapy and surgery, mortality caused by colorectal cancer remains very high and has decreased only slightly. Thirty percent of all colon cancer deaths can be linked to diet (Stone and Papas 1997). Probably one of the most cited mechanisms is that of a high-fat diet. To resorb the increased amount of dietary fat, more conjugated primary bile salts are secreted in the small intestine. This leads to an increased ¯ux of bile salts to the colon and an increased metabolism of the bile salts by the indigenous ¯ora (Giovannucci and Goldin 1997). Some of the bile salts generated by the micro-organisms have been incriminated in colonic carcinogenesis (Singh et al. 1997). One of the most important bacterial bile salt transformations is bile salt hydrolysis, which is mediated by a wide range of colon anaerobes (Hill 1995). The bile salt hydrolase enzyme releases the glycine and/or taurine moiety from the side chain of the bile salt steroid core and generates deconjugated primary bile salts, which are less water-soluble and are excreted more easily via the faeces (De Smet et al. 1994). The principle of bile salt hydrolysis was used to decrease the serum cholesterol in pigs through interaction with the host's bile salt metabolism (De Smet et al. 1998). In the latter, the bacterial bile salt hydrolase activity in pigs was enhanced by oral administration of Lactobacillus reuteri. This caused a greater drain on the bile salt pool, resulting in a loss of feedback inhibition on bile salt synthesis and an increased conversion of cholesterol into bile salts. Because of this, the serum cholesterol levels in the pigs were lowered. During this in vivo experiment, no detailed information was collected about the possible probiotic eect of this strain. An in vitro study using the Simulator of the Human Intestinal Microbial Ecosystem (SHIME) was carried out recently to evaluate the eect of adding L. reuteri on the composition and activity of the gut microbiota (Nollet et al. 1999). The experiments indicated that L. reuteri was able to repress the concentration of Enterobacteriac and coliforms, which are considered potential enteropathogens (Isolauri et al. 710 1991). It was suggested that the competitiveness of L. reuteri was largely due to its bile salt hydrolase activity (Nollet et al. 1999). Although the bile salt hydrolase-active L. reuteri oers potential as a `biological' alternative to pharmaceutical interventions to treat hypercholesterolaemia and to be used as a probiotic, there is medical concern about possible side eects. According to some authors (Hylemon and Glass 1983; Kandell and Bernstein 1991), enhanced bile salt hydrolytic activity is not favourable because a subsequent dehydroxylation of these deconjugated primary bile salts by 7a-dehydroxylase-active strains could generate toxic and/or mutagenic secondary bile salts (Marteau et al. 1995; Nagengast et al. 1995). It was suggested that an increase in faecal secondary bile salts upon ingestion of bile salt hydrolase-active lactobacilli should be regarded as a potential long-term colon cancer-promoting eect (van Faassen et al. 1987). Furthermore, these toxic bile salts could disturb the normal microbiota of the gut, leading to diarrhoea, mucosal in¯ammation or activation of harmful drugs and carcinogens in the intestinal contents (Salminen et al. 1996). This study investigated the eects of enhanced bacterial bile salt modi®cations, induced by adding L. reuteri, on the fermentative capacity of the simulated microbiota of the colon. This was done by analysing the concentration of short chain fatty acids (SCFAs) and the b-galactosidase activity. The possible generation of harmful bile salts, induced by enhanced bacterial bile hydrolase activity was investigated using a haemolysis test, Vitotox and the neutral red uptake (NRU) cytotoxicity assay. Materials and methods Simulator of the Human Intestinal Microbial Ecosystem The possible adverse eects linked to bile salts and/or enhanced bacterial bile salt hydrolase activity were investigated in batch experiments using the gut microbiota cultured in the SHIME. This six-stage computer-controlled reactor was developed to simulate the bacterial communities found in the distinctive parts of the human intestine. Each of the six reactors contains the microbiota of a dierent part of the human gastro-intestinal tract, in sequence (compartment 1±6): the stomach, the duodenum, the small intestine, the ascending, the transverse and the descending colon (Molly et al. 1993). The reactor was set up and validated as described previously (Molly et al. 1994). Brie¯y, the last three vessels were inoculated on ®ve consecutive days with a faecal suspension from ®ve healthy volunteers. This inoculum was stabilised by being fed three times a day with 200 ml of a carbohydrate-based medium containing arabinogalactan (1 g/l), pectin (2 g/l), xylan (1 g/l), starch (3 g/l), glucose (0.4 g/l) and mucin (4 g/l). Isolation and identi®cation The strain was originally isolated from pig faeces and selected because of its high bile salt hydrolase activity. The bacterium was identi®ed as L. reuteri using SDS-PAGE analysis (De Smet et al. 1998). Preparation of the L. reuteri culture A 1% inoculum of the frozen L. reuteri strain was thawed and suspended in 10 ml de Man Rogosa and Sharpe (MRS) broth (Oxoid), which was reduced by adding 1 g/l sodium thioglycolate (Fluka). After 24 h incubation at 37 °C, the culture was centrifuged (10 min at 5,000 g) and washed with sterile physiological solution before use in the batch incubations. The number of colony-forming units (CFU) used in the test was determined by making ten-fold serial dilutions of the overnight culture in physiological solution (8.5 g NaCl/l) and plating the dilutions on Rogosa agar (Oxoid) supplemented with 5 mM taurodeoxycholic acid (Dashkevicz and Feighner 1989). The bile salt hydrolase activity was quanti®ed by measuring the rate with which glycocholic acid was hydrolysed using a technique described in detail elsewhere (De Smet et al. 1994). Experimental set-up Samples (10 ml) were withdrawn from compartment 5 of the SHIME reactor and centrifuged for 10 min at 5,000 g. The microbial pellet was washed with an aliquot of sterile physiological solution and resuspended in 10 ml of fresh SHIME feed, kept at pH 7.0 by means of a 0.1 M phosphate buer. The resuspended pellets were supplemented with 0, 5 or 30 g oxgall/l by adding an appropriate amount of a 100 g/l ®ltersterile oxgall solution (Difco). To investigate the eects of enhanced bile salt hydrolase activity, a treatment was imposed by adding L. reuteri, resulting in a viable concentration in the test of 7.5 0.5 log10 CFU/ml. A series without the oxgall addition was used as a control. Finally, the tubes were made anaerobic by ¯ushing the liquid phase for 10 min with oxygen-free nitrogen gas. After 24 h incubation at 37 °C, samples were taken to determine the concentration of SCFAs and b-galactosidase activity. The remaining liquid was centrifuged for 10 min at 5,000 g and the supernatant was sterilised using a 0.22-lm ®lter. The sterile culture supernatants were analysed for possible (geno)toxic products by means of the haemolysis test, Vitotox (Vito, Belgium) and the NRU cytotoxicity assay. Determination of SCFAs SCFAs produced by the bacterial cultures were extracted and measured with a gas chromatograph (GC; Carlo Erba Fractovap 4160) equipped with a ¯ame ionisation detector and a Delsi Nermag integrator (Nollet et al. 1999). The GC was equipped with a capillary free fatty acids packed column (25 m ´ 0.53 mm; ®lm thickness 1.2 lM). Nitrogen was used as a carrier gas at a ¯ow rate of 20 ml/min. The column temperature was 130 °C and the temperature of injection port and detector was 195 °C. The concentration of the SCFAs was expressed in millimoles per liter. Determination of b-galactosidase activity The samples were centrifuged at 10,000 g for 10 min. Cell-free supernatant (100 ll) was pipetted into a 96-well plate, with 100 ll of 5.0 mM p-nitrophenyl-b-galactopyranoside, prepared in a 0.1 M phosphate buer (pH 6.5). The plates were incubated at 37 °C and the absorbance at 405 nm was read after 30 min with a Biokinetics EL312e multi-well reader. The amount of p-nitrophenol released was measured based on a standard curve of p-nitrophenol. The results were expressed in micromoles of p-nitrophenol released per ml per min (Berg et al. 1978). Determination of haemolysis Part of the ®ltersterile supernatant (20 ll) was mixed with 140 ll phosphate-buered saline. After incubation for 1 min at 37 °C in a 711 shaking water bath, 40 ll of red blood cells (10% suspension, ICN) were added. Simultaneously, red blood cells were incubated in phosphate-buered saline (0% lysis) and in double-distilled water (100% lysis). Samples were centrifuged for 1 min at 10,000 g after incubation for 10 min at 37 °C. The supernatant was diluted four times in double-distilled water and percentage haemolysis was determined by measuring the absorption at 540 nm (Van Der Meer et al. 1991). (Geno)toxicity evaluated with Vitotox This newly developed genotoxicity test employs Salmonella typhimurium TA104 recN. This strain contains the lux operon under transcriptional control of the recN gene, which is a part of the SOS-system. Incubation of the strain with a genotoxic solution results in a derepression of the recN promotor and an increased expression of the lux operon, resulting in increased light production. Some products act on the light production or enhance the bacterial metabolism, creating false positive results. Therefore, a S. typhimurium TA104 with a promoterless lux operon (pri) was used as a control. This constitutively light-producing strain can also be used as a toxicity sensor, with a decrease in light production being interpreted as an indication of toxicity (van der Lelie et al. 1996). S. typhimurium TA104 recN and pri were grown overnight on a rotative shaker (170 rpm) at 37 °C in a normal bacterial growth medium supplemented with CaCl2. Next, the bacterial suspensions were diluted 500 times in 2.5 ml fresh growth medium. An aliquot of these dilutions (90 ll) was added to each well of a 96-well plate, already containing 10 ll SHIME supernatant diluted ten times. The plates were placed in a Microlumat LB96P luminometer (EG and G Berthold) and the light production of the strains was followed as a function of time, using the following measuring conditions: 1 s/well, cycle time of 300 s, 4 h measuring time, 30 °C. The validity of the test was veri®ed using a known genotoxic compound [0.02 lmol 4-nitroquinoline-1-oxide/l dimethylsulphoxide (DMSO)]. DMSO was used as a negative control. After completion of the measurements the signal to noise ratios (S/N) were plotted as a function of the incubation time. In the case of genotoxic eects, the ratio of the maximum S/N of the recN strain versus the maximum S/N of the pri strain (rec/pri) became larger than 2. In the case of toxic eects, the maximum S/N for the pri strain became smaller than 0.8. Toxicity evaluated with the NRU assay HeLa cells (ECECCC 85060701) were harvested at subcon¯uency (70±95%) and plated in 96-well plates. Each well contained 200 ll tissue culture medium (TCM) with 2,000 cells. After 24 h incubation at 5% CO2, 37 °C and 95% humidity, the TCM was removed and a TCM containing either sterile SHIME culture supernatants or a toxic compound (0.2 lM cycloheximide) was added. Sterile water was used as a negative control. The SHIME liquids were tested in six dierent concentrations at 0.04±1.25%. After 3 days incubation, the TCM was removed and replaced with a prewarmed TCM containing 1% neutral red. After a further 3 h incubation, the NRU by the HeLa cells was measured using a Cyto¯uor 2350 (PerSeptive Biosystems). The NRU of the cells incubated with diluted SHIME suspensions or cycloheximide was expressed as a percentage compared to the NRU of the negative control, with a decreased NRU indicating toxicity (Vander Plaetse and Schoeters 1995). Statistical analysis Statistical analysis of the experiments were performed using an unpaired two tailed t-test. Results Fermentation capacity of the colonic microbial ecosystem SCFAs are end-products of bacterial metabolism and a concentration decrease in comparison with the control is an indication of inhibition of bacterial activity over the total incubation period. Measuring enzyme activity after 24 h provides information about the bacterial activity at the end of the incubation period. In contrast with the measurement of SCFAs, it only re¯ects bacterial activity at the sampling time. The data about the concentration of SCFAs and b-galactosidase activity indicate that an oxgall supplementation burdened the fermentation of the gut microbiota in a concentration-dependent manner (Table 1). The concentration of SCFAs was lower in comparison with the control (0 g/l oxgall + L. reuteri), although not signi®cantly when oxgall at 5 g/l or 30 g/l was added. Adding L. reuteri led to an increased concentration of SCFAs, which was only statistically dierent from the control (P £ 0.01) in the case of 5 g oxgall/l (Table 1). A decrease in b-galactosidase activity was noticed with increasing concentrations of oxgall. This reduced activity was completely (in the case of 5 g oxgall/l) or partially (in the case of 30 g oxgall/l) counterbalanced by the addition of L. reuteri (Table 1). When the ratio for SCFAs production and b-galactosidase activity was calculated between the series with L. reuteri and the series without L. reuteri, it was observed that in all cases the values were higher than 1. This indicates that the addition of L. reuteri circumvented the negative effects of the oxgall supplementation. The presence of 7.5 0.5 log10 CFU L. reuteri/ml in the test was determined using the plating data of the overnight L. reuteri culture. The quantitative determination of the bile salt hydrolase activity indicated that cholic acid was released at a rate of 2.56 lmol per 1010 CFU per min. Haemolytic eects The haemolytic capacity of the SHIME supernatants was determined after 24 h incubation. Samples without Table 1 Short chain fatty acids (SCFAs) production (lmol/ml) and b-galactosidase activity (lmol p-nitrophenol released/ml per min) in Simulator of the Human Intestinal Microbial Ecosystem suspensions after 24 h incubation with increasing concentrations of oxgall and with or without 7.5 0.5 log10 CFU Lactobacillus reuteri/ml (n = 3). Signi®cant dierence from the series 0 g oxgall/l + L. reuteri: *P £ 0.05, **P £ 0.01 Series Parameter SCFAs 0 g oxgall/l + L. reuteri 5 g oxgall/l 5 g oxgall/l + L. reuteri 30 g oxgall/l 30 g oxgall/l + L. reuteri 58.2 55.8 68.8 53.3 63.1 b-galactosidase 0.7 0.4 0.5** 1.1 3.2 171.3 134.6 190.4 87.6 104.4 6.2 2.8* 6.7 3.5** 5.0* 712 oxgall and L. reuteri treatment did not induce lysis of the red blood cells. The supernatants originating from the liquids supplemented with 5 g oxgall/l caused total lysis (100 12%) during the 10 min incubation. The addition of L. reuteri reduced the lytic eect to 2.1 2.8%. All samples caused total lysis when liquids with 30 g oxgall/l were tested, even when L. reuteri was added. (Geno)toxicity evaluated with Vitotox In the Vitotox assay, the reference substance nitroquinoline-1-oxide brought about a rec/pri signal of 5.05 0.61, indicating genotoxicity. Without the L. reuteri supplement, the rec/pri ratios were 1.13 0.09 and 1.01 0.09 for 5 g oxgall/l and 30 g oxgall/l respectively. When L. reuteri was added, the ratios were 1.04 0.35 and 0.92 0.04 respectively. The data indicate that none of the sterile SHIME culture supernatants exhibited genotoxic eects. The S/N ratio for the pri strain became lower than 0.8 when 5 g oxgall/l was added, indicating physiological cell toxicity. The pri strain was able to recover from this toxic shock as the maximum S/N became higher than 1 during the test. The addition of L. reuteri prevented this toxic eect and temporarily stimulated the light production of the test strain. In the case of 30 g oxgall/l, the Salmonella strain was only able to recover when the supernatant originated from a series supplemented with 7.5 0.5 log10 CFU L. reuteri/ml (Fig. 1). Toxicity evaluated with the NRU assay The NRU assay was validated by verifying that 0.2 lM cycloheximide induced an inhibition of NRU by HeLa Fig. 1 Kinetics of the Signal/Noise ratio of the Vitotox Salmonella typhimurium TA104 pri strain incubated with sterile Simulator of the Human Intestinal Microbial Ecosystem (SHIME) supernatants supplemented with 5 g oxgall/l, with (s) or without (d) 7.5 0.5 log10 CFU Lactobacillus reuteri/ml and 30 g oxgall/l with (,) or without 7.5 0.5 log10 CFU L. reuteri/ml (.) cells of 9±18%. No dierence in NRU was seen between the control and the series incubated with dilutions of ®ltersterile SHIME supernatants supplemented with 5 g oxgall/l with and without the bile salt hydrolaseactive L. reuteri (results not shown). When the highest three dilutions (0.31, 0.62 and 1.25%) of the series supplemented with 30 g oxgall/l were tested, a reduced NRU by the HeLa cells could be observed. This eect was not observed when the suspension was preincubated with 7.5 0.5 log10 CFU L. reuteri/ml (Fig. 2). Discussion In a faecal suspension simulated by the SHIME reactor (Molly et al. 1994), we investigated whether enhancement of bacterial bile salt modi®cation, by the addition of the bile salt hydrolase-active L. reuteri, would lead to increased dehydroxylation and the generation of harmful bile salts. The SHIME liquids (with and without L. reuteri) were incubated for 24 h with 5 g oxgall/l and 30 g oxgall/l. According to the manufacturer of the oxgall, the concentrations supplied to the suspensions corresponded to about 1 mM and 6 mM bile salts respectively. Addition of 7.5 0.5 log10 CFU L. reuteri/ml to the test tubes with oxgall at 5 g/l and 30 g/l counteracted the detrimental eects. The SCFAs concentration increased to levels higher than the control, while the b-galactosidase activity recovered only partially. The results of the haemolysis test suggest that the detrimental eects of adding oxgall were probably due to membranedamaging eects. Any lysis of the erythrocytes used in this test can be attributed to membrane-damaging eects as these cells do not possess cell organelles and lack speci®c mechanisms for the uptake and metabolism of bile salts (Pazzi et al. 1997). Due to the L. reuteri addition, haemolysis disappeared almost completely in the case of 5 g oxgall/l. Fig. 2 Neutral red uptake (%NRU) by HeLa cells treated with sterile SHIME culture supernatants supplemented with 30 g oxgall/l (d) and 30 g oxgall/l with 7.5 0.5 log10 CFU L. reuteri/ml (s). Data are the mean values for two separate experiments. Error bars indicate standard deviations. Signi®cant dierence from the series without L. reuteri: **P £ 0.01 713 The potential generation of genotoxic bile salts after an enhanced bile salt hydrolysis was investigated with Vitotox. None of the samples revealed genotoxic eects. At the concentration of 5 g oxgall/l, all samples exhibited transient toxicity on the Vitotox strain. When the solutions with 30 g oxgall/l were tested, the Vitotox strain only recovered from the toxic pulse when the solution was preincubated with L. reuteri. Possible cytotoxic eects of the SHIME supernatants were traced using the NRU cytotoxicity assay. The HeLa cells employed in this test are human epithelial cells derived from a cervix carcinoma. They are non-specialised cells, suitable for measuring general toxicity caused by substances interfering with non-specialised housekeeping cell functions. Again, increasing concentrations of oxgall caused detrimental eects and these were circumvented by the addition of L. reuteri. It is currently thought that the promotion of colon cancer by dietary fat involves the excess production of bile salts and the bacterial conversion of conjugated primary bile salts to potentially dangerous unconjugated secondary bile salts (Morotomi et al. 1997). Intestinal bacteria ®rst deconjugate bile salts before they are further metabolised (Batta et al. 1990). The predominant metabolism of the deconjugated primary bile salts (cholic acid and chenodeoxycholic acid) is the 7a-dehydroxylation into the secondary bile salts (deoxycholic acid and litocholic acid). Because of the risk of forming harmful secondary bile salts after an initial deconjugation step, bacterial bile salt hydrolysis has been considered a risk factor for the development of colon cancer. The precise mechanism through which these molecules exert their eect is not fully understood. On one hand, it is thought that they might act through co-mutagenic and co-carcinogenic eects, thereby enhancing colorectal neoplasm (Martin et al. 1981). On the other hand, the hydrophobic nature of the secondary bile salts could be responsible for disrupting the membrane integrity of mucosal cells, ultimately leading to cytotoxicity. This would increase cell proliferation, which is considered to be a biomarker for the development of cancer (Preston-Martin et al. 1990). There are numerous examples that bile salt hydrophobicity correlates well with toxic eects on all kinds of cells: bacterial cells, isolated hepatocytes, gastric mucosa, colonic mucosa, etc. (llani and Granoth 1990; Heuman et al. 1996; Pazzi et al. 1997). In the case of toxicity on bacterial cells, De Smet et al. (1995) suggested that the protonated form of the bile salt exhibits its eect through the same mechanism as organic acids, namely by causing intracellular acidi®cation. The conjugated bile salts, which are protonated, enter the cell via passive diusion and are dissociated in the bacterial cytoplasm due to the higher pH. This causes acidi®cation of the cytoplasm and collapse of the proton motive force, resulting in an inhibition of the nutrient transport and thus toxicity. The results presented in this paper clearly indicate that bile salts, supplemented under the form of oxgall at millimolar concentrations, cause toxic eects on bacterial cells, blood cells and epithelial cells. In par- ticular, the data from the red blood cells lead to the reasonable assumption that the mode of action is toxicity through membrane damage. This phenomenon has been reported in literature (Albalak et al. 1996; Shekels et al. 1996). Based on literature data, it was assumed that enhanced bile salt hydrolysis would make more deconjugated bile salts available for 7a-dehydroxylation. This would lead to a higher concentration of secondary bile salts which are (geno)toxic and mutagenic compounds (Nagengast et al. 1995). Most of this evidence was gained from in vitro tests and laboratory animal models. In the latter, secondary bile salts have been shown to induce hyperproliferation of the colonic epithelium in rats, which makes the animals more susceptible to experimentally induced colon cancer (Christl et al. 1995). However, this hypothesis is nevertheless considered controversial because of contradictory case control studies (Owen 1997). In our experiments, addition of the bile salt hydrolase-active L. reuteri did not result in an increase in detrimental eects, but on the contrary brought protection against the bile salts. Based on the bile salt hydrolase data, it was estimated that during the 24 h incubation period about 1 mmol/l glycocholic acid was hydrolysed. If glycocholic acid was the sole bile salt present in oxgall, then approximately 100% and 20% would have been hydrolysed in the tubes to which 5 g oxgall/l and 30 g oxgall/l was added. A plausible mechanism by which detrimental eects of bile salts could have been avoided is precipitation of the deconjugated bile salts, thereby decreasing the bioavailable concentration. It is well known that bile salt transformations have an impact on physicochemical properties such as ionisation, solubility and micelle formation, which are correlated with the structure of the bile salt molecule. Removal of the amino acid moiety from the side chain by deconjugation results in compounds that are less resistant to precipitation at low pH or by divalent cations such as Ca2+ (Fini and Roda 1987; De Boever and Verstraete 1999). A second mechanism for the protective properties of L. reuteri could involve a physical binding of bile salts by the bacterium, thereby making them less bioavailable. Evidence from in vitro as well as in vivo studies supports the concept that Lactobacilli and other lactic acid bacteria are able to bind a wide range of mutagens and (geno)toxins (Hosoda et al. 1992; Orrhage et al. 1994; Pool-Zobel et al. 1996). All tests were done with a simulator of the gastrointestinal microbial ecosystem (SHIME). Therefore, these in vitro studies should be considered as a preliminary evaluation of the putative protective action exerted by L. reuteri when it is consumed as a probiotic. More profound research has to be conducted about the type of bile salt removed from the liquid phase and the precise mechanism involved in this process. Nevertheless, the potential decrease of the bioavailable concentration of toxic bile salts is a point in favour of the use of L. reuteri as a human probiotic (Salminen et al. 1996; De Smet et al. 1998; Nollet et al. 1999). 714 Acknowledgements This research has been funded by a scholarship from the Flemish Institute for the Improvement of Scienti®c± Technological Research in Industry (IWT). The experiments carried out at the VITO were sponsored by AVECOM (Belgium). References Albalak A, Zeidel ML, Zucker SD, Jackson AA, Donovan JM (1996) Eects of submicellar bile salt concentrations on biological membrane permeability to low molecular weight non-ionic solutes. Biochemistry 35: 7936±7945 Batta AK, Salen G, Arora R, Shefer S, Batta M, Person A (1990) Side chain conjugation prevents bacterial 7a-dehydroxylation of bile salts. J Biol Chem 265: 10925±10928 Berg JO, Nord CE, Wadstrom T (1978) Formation of glycosidases in batch and continuous cultures of Bacteroides fragilis. Appl Microbiol Biotechnol 35: 269±273 Christl SU, Bartram HP, Ruckert A, Scheppach W, Kasper H (1995) In¯uence of starch fermentation on bile acid metabolism by colonic bacteria. Nutr Cancer 24: 67±75 Dashkevicz MP, Feighner MP (1989) Development of a dierential medium for bile salt hydrolase-active Lactobacillus spp. Appl Microbiol Environ 53: 331±336 De Boever P, Verstraete W (1999) Bile salt deconjugation by Lactobacillus plantarum 80 and its implication for bacterial toxicity J Appl Micorbiol 87: 345±352 De Smet I, Van Hoorde L, De Sayer N, Vande Woestyne M, Verstraete W (1994) In vitro study of bile salt hydrolase (BSH) activity of BSH isogenic Lactobacillus plantarum 80 strains and estimation of cholesterol lowering through enhanced BSH activity. Microb Ecol Health D 7: 315±329 De Smet I, Van Hoorde L, Vande Woestyne M, Christiaens H, Vestraete W (1995) Signi®cance of bile salt hydrolytic activities of lactobacilli. J Appl Bacteriol 79: 292±301 De Smet I, De Boever P, Verstraete W (1998) Cholesterol lowering in pigs through enhanced bacterial bile salt hydrolase activity. Br J Nutr 79: 185±194 Fini A, Roda A (1987) Chemical properties of bile acids IV. Acidity constants of glycine-conjugated bile acids. J Lipid Res 28: 755± 759 Ghadirian P, Laxroix A, Maisonneuve P (1997) Nutritional factors and colon carcinoma. Cancer 80: 858±864 Giovannucci E, Goldin B (1997) The role of fat, fatty acids and total energy intake in the etiology of human colon cancer. Am J Clin Nutr 66: 1564±1571 Heuman DM, Bajaj RS, Lin Q (1996) Adsorption of mixtures of bile salt taurine conjugates to lecithin-cholesterol membranes: implications for bile salt toxicity and cytoprotection. J Lipid Res 37: 562±573 Hill MJ (1995) Bacteria and fat digestion. In: Hill MJ (ed) Role of gut bacteria in human toxicology and pharmacology. Taylor and Francis, London, pp 131±142 Hosoda M, Hashimoto H, Morita H, Chiba M, Hosono A (1992) Antimutagenicity of milk cultured with lactic acid bacteria against N-methyl-n¢-nitro-N-nitrosoguanidine. J Dairy Sci 75: 976±981 Hylemon PB, Glass TL (1983) Biotransformations of bile acids and cholesterol by the intestinal micro¯ora. In: Hentges DJ (ed) Human intestinal ¯ora in health and disease. Academic Press, New York, pp 79±99 IIani A, Granoth R (1990) The pH dependence of the hemolytic potency of bile salts. Biochim Biophys Acta 1027: 199±204 Isolauri E, Juntunen M, Rautanen T, Sillanaukee P, Koivula TA (1991) Human Lactobacillus strain (Lactobacillus casei sp. strain GG) promotes recovery from acute diarrhea in children. Pediatrics 88: 90±97 Kandell RL, Bernstein C (1991) Bile salt/acid induction of DNA damage in bacterial and mammalian cells: implications for colon cancer. Nutr Cancer 16: 227±238 Marteau P, Gerhardt MF, Myara A, Bouvier E, Trivin F, Rambaud JC (1995) Metabolism of bile salts by alimentary bacteria during transit in the human small intestine. Microb Ecol Health D 8: 151±157 Martin MS, Justrabo E, Jeannin JF, Leclerc A, Martin F (1981) Eect of dietary chenodeoxycholic acid on intestinal carcinogenesis induced by 1,2 dimethylhydrazine in mice. Br J Cancer 43: 884±886 Molly K, Vande Woestyne M, Verstraete W (1993) Development of a 5-step multi-chamber reactor as a simulation of the human intestinal microbial ecosystem. Appl Microbiol Biotechnol 39: 254±258 Molly K, Vande Woestyne M, De Smet I, Verstraete W (1994) Validation of the simulator of the human intestinal microbial ecosystem (SHIME) reactor using microorganism-associated activities. Microb Ecol Health D 7: 191±200 Morotomi M, Sakaitani Y, Satou M, Takahashi T, Takagi A, Onoue M (1997) Eects of a high-fat diet on azoxymethaneinduced aberrant crypt foci and fecal biochemistry and microbial activity in rats. Nutr Cancer 27: 84±91 Nagengast FM, Grobben MJ, van Munster IP (1995) Role of bile acids in colorectal carcinogenesis. Eur J Cancer 31: 1067± 1070 Nollet LJA, Pereira DI, Verstraete W (1999) Eect of a probiotic bile salt hydrolytic Lactobacillus reuteri on the human gastrointestinal microbiota as simulated in the SHIME reactor system. Microb Ecol Heath D 11: 13±21 Orrhage K, Sillerstrom E, Gustafsson J-A, Nord CE, Rafter J (1994) Binding of mutagenic heterocyclic amines by intestinal and lactic bacteria. Mutat Res 311: 239±248 Owen RW (1997) Faecal steroids and colorectal carcinogenesis. Scand J Gastroenterol 32: 76±82 Pazzi P, Puviani AC, Dalla Libera M, Guerra G, Ricci D, Gullini S, Ottolenghi C (1997) Bile salt-induced cytotoxicity and ursodeoxycholate cytoprotection: in-vitro study in perifused rat hepatocytes. Eur J Gastroenterol Hepatol 9: 703±709 Pool-Zobel BL, Neudecker C, Domizla I, Ji S, Schillinger U, Rumney C, Moretti M, Vilarini I, Scassellati-Sforzollini R, Rowland I (1996) Lactobacillus- and Bi®dobacterium-mediated antigenotoxicity in the colon of rats. Nutr Cancer 26: 365± 380 Preston-Martin S, Pike MC, Ross RK, Jones PA, Henderson BE (1990) Increased cell division as a cause of human cancer. Cancer Res 50: 7415±7421 Salminen S, Isolauri E, Salminen E (1996) Clinical uses of probiotics for stabilizing the gut mucosal barrier: Successful strains for future challenges. Antonie van Leeuwenhoek 70: 347±358 Shekels LL, Lyftogt CT, Ho SB (1996) Bile acid-induced alterations of mucin production in dierentiated human colon cancer cell lines. Int J Biochem Cell Biol 28: 193±201 Singh J, Hamid R, Reddy BS (1997) Dietary fat and colon cancer: modulating eect of types and amount of dietary fat on ras-p21 function during promotion and progression stages of colon cancer. Cancer Res 57: 253±258 Stone WL, Papas AM (1997) Tocopherols and the etiology of colon cancer. J Natl Cancer Inst 89: 1006±1014 van der Lelie D, Regniers L, Borremans B, Provoost A, Verschaeve L (1996) The VITOTOX test, an SOS bioluminescence Salmonella typhimurium test to measure genotoxicity kinetics. Mutat Res 389: 279±290 Van Der Meer R, Termont DSML, De Vries HT (1991) Dierential eects of calcium ions and calcium phosphate on cytotoxicity of bile acids. Am J Physiol 260: G142±G147 Vander Plaetse F, Schoeters G (1995) A rapid method for identi®cation of the cellular targets of xenobiotics. Clin Chem 41: 1906±1908 van Faassen A, Bol J, van Dokkum W, Pikaar NA, Ockhuizen T, Hermus RJJ (1987) Bile acids, neutral steroids and bacteria in faeces as aected by a mixed, a lacto-ovovegetarian and a vegan diet. Am J Clin Nutr 46: 962±967