Survey
* Your assessment is very important for improving the work of artificial intelligence, which forms the content of this project
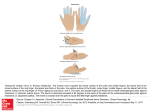
Motor Control, 2008, 12, 311-329 © 2008 Human Kinetics, Inc. Strength Training Increases Training-Specific Multifinger Coordination in Humans Jae Kun Shim, Jeffrey Hsu, Sohit Karol, and Ben F. Hurley The purpose of the current study was to investigate the effects of finger strength training (ST) on finger strength, independence, force control, and adaptations in multifinger coordination. Thirty-three healthy, young (23.0 ± 2.9 years) subjects were randomly assigned into 4 groups. Group 1 (G1) trained all fingers together, Group 2 (G2) trained individual fingers without restricting movements of the nontraining fingers, and Group 3 (G3) trained individual fingers while restricting the movement of the nontraining fingers. The control group (G0) did not undergo any training. A vertically hanging load was attached to a spring that passed through a pulley. The other end of the string extended to the horizontal plane and had thimbles attached to it. Subjects were asked to rest their forearm on the table and lift the load by inserting their fingers into the thimbles. The training protocol lasted 6 weeks. Identical experimental tests were conducted 4 times, biweekly, across the 6-week training. Force coordination and moment coordination, defined as synergies stabilizing the resultant force and the resultant moment of all finger forces, in a multifinger pressing task were quantified using the Uncontrolled Manifold (UCM) analysis. The UCM analysis allocates motor variability into two components, one in the null space of a motor task and the other perpendicular to the null space. During multifinger pressing tasks, multifinger coordination exists when the variability in the null space is greater than the variability in the subspace perpendicular to the null space. The multifinger coordination was quantified as the difference between the variance within the null space and that perpendicular to the null space, normalized by the total variance. Thus, the coordination measure in our analysis is a unitless variable. A greater coordination measure indicates better multifinger coordination. Moment-stabilizing multifinger coordination increased only in G1 (from 1.197 ± 0.004 to 1.323 ± 0.002, p < .01), and force-stabilizing coordination increased only in G3 (from 0.207 ± 0.106 to 0.727 ± 0.071, p < .01). Finger strength, measured by the maximal voluntary finger force of pressing 4 fingers, increased significantly in all training groups (from 103.7 ± 3.1 N to 144.0 ± 3.6 N for training groups, all p < .001). Finger-force errors, quantified by the deviations between the required force profiles (20% maximal voluntary force) presented to the subjects and the actual force produced, decreased significantly with ST for The authors are with the Dept. of Kinesiology, University of Maryland, College Park, MD 20742. Shim is also with the Dept. of Neuroscience and Cognitive Science and the Dept. of Bioengineering at the University of Maryland. 311 312 Shim et al. all the training groups (all p < .05). Finger independence also decreased significantly for all the training groups (p < .05). We conclude that the neuromuscular system adaptations to multifinger ST are specific to the training protocol being employed, yielding improvements in different types of multifinger coordination (i.e., coordination-specific ST), finger-force control, and finger strength and a decrease in finger independence. Finger independence, depending on the nature of the task, might or might not be favorable to certain task performances. We suggest that ST protocol should be carefully designed for the improvement of specific coordination of multieffector motor systems. Keywords: finger, training, specificity, synergy, enslaving, UCM The hand and fingers are some of the main tools the central nervous system (CNS) uses to physically interact with the external world. Multiple fingers are used in everyday manipulative tasks, some as simple as grasping a glass of water (Nowak & Hermsdorfer, 2003; Rearick, Casares, & Santello, 2003). Therefore, the CNS needs to control the kinetic redundancy caused by superfluous finger contact forces and moments of force for dexterous manipulation of an object (Shim, Latash, & Zatsiorsky, 2003, 2005). Previous studies that investigated manipulative dexterity also suggested a requirement for the CNS to coordinate the fingers for a stable performance during manipulative tasks, also known as multifinger synergy (Engel, Flanders, & Soechting, 1997; Fish & Soechting, 1992; Flanders & Soechting, 1992; Li, Dun, Harkness, & Brininger, 2004; Shim et al., 2003; Shim, Latash, et al., 2005). Specifically, multifinger synergy has been defined as the task-specific covariation of individual finger actions to stabilize a particular performance variable. Previous studies have suggested that some manipulative tasks, such as typing on a keyboard and playing piano, require not only the synergic actions of multiple fingers, but also individual finger actions, known as individual finger independence (Hager-Ross & Schieber, 2000; Schieber & Poliakov, 1998; Schieber & Santello, 2004). Previous studies of neuromuscular strength training (ST) on the hand and fingers have either employed simultaneous training to all fingers or training to individual fingers. The studies showed that ST can cause neuromuscular changes including adaptations to the neural control of finger muscles and increases in hand and finger strength (Keen, Yue, & Enoka, 1994; Laidlaw, Kornatz, Keen, Suzuki, & Enoka, 1999; Tracy, Byrnes, & Enoka, 2004). However, these studies were limited to investigating the possible effects and behavioral changes of ST on the multieffector motor system. No previous studies have investigated whether ST can change multifinger synergies when specific training protocols, intended to change such synergies (i.e., “training specificity to multieffector synergy/coordination”), are employed. Most of the previous studies have investigated the presence of multifinger synergies with respect to two important performance variables: the total force produced by different fingers and the corresponding total moments produced by those forces about the functional longitudinal axes of the hand (Kang, Shinohara, Zatsiorsky, & Latash, 2004; Shim, Kim, et al., 2005). It has been suggested that the CNS is capable of using the motor redundancy of multifinger actions and stabilizing the resultant force and/or resultant moment during pressing tasks (Kang et al., 2004; Strength Training and Multifinger Coordination 313 Shinohara, Scholz, Zatsiorsky, & Latash, 2004) as well as grasping tasks (Oliveira, Shim, Loss, Petersen, & Clark, 2006; Shim, 2005; Shim, Latash, & Zatsiorsky, 2004; Shim, Latash, et al., 2005). In previous studies that examined static grasping tasks with multiple fingers, there was evidence suggesting that multiple fingers act together to minimize linear and rotational motion of the hand-held object by minimizing the resultant force (e.g., resultant force stabilization for linear equilibrium control) and resultant moment (e.g., resultant moment stabilization for rotational equilibrium control). Although the CNS can use the kinetic redundancy to control the resultant force and the resultant moments during multifinger manipulative tasks, the ability to coordinate the motor redundancy changes with age, genetic conditions, and neurological conditions (Kang et al., 2004; Olafsdottir, Zhang, Zatsiorsky, & Latash, 2007; Reisman & Scholz, 2006; Shim, Lay, Zatsiorsky, & Latash, 2004). Thus, the development of coordination-specific ST protocols for multifinger manipulation is imperative. Neuromuscular adaptations to ST are known to be highly specific to the training protocols administered (Dons, Bollerup, Bonde-Petersen, & Hancke, 1979; Enoka, 1997; Sale, 1988). However, whether different ST protocols can induce coordination patterns that are specific to the resultant force or resultant moment stabilization during multifinger pressing tasks is unknown. The purpose of this study is to investigate whether specifically designed training protocols can be employed to induce changes in the coordinative actions of a multieffector system. Simultaneous training of all fingers in the same direction encourages positive covariation between individual finger forces. The positive covariation stabilizes the rotational equilibrium (i.e., moment-stabilizing synergy) during multifinger pressing and prehension. Individual-finger training without movement of nontraining fingers might increase negative covariation among finger forces (e.g., force-stabilizing synergy; Latash, Shim, Gao, & Zatsiorsky, 2004; Latash, Shim, Smilga, & Zatsiorsky, 2005; Shim, Latash, et al., 2005; Shim, Olafsdottir, Zatsiorsky, & Latash, 2005). The current study employed three different finger-training protocols for different training groups: four-finger simultaneous training in an attempt to increase the moment-stabilizing synergy, individual-finger training without restricting movement of nontraining fingers, and individual-finger training while self-restricting nontraining finger movements in an attempt to increase the force-stabilizing synergy. In this study, we tested whether the different training protocols specifically designed to change force-stabilizing synergy or moment-stabilizing synergy could induce the intended adaptations in multifinger control strategies. If the training adaptations for multifinger control strategies are specific to the training protocols, control strategies during multifinger actions are expected to change. We hypothesized that the neuromuscular adaptations will occur in favor of moment-stabilizing synergy after four-finger simultaneous training and force-stabilizing synergy after individualfinger training (Hypothesis 1). We also hypothesized that finger independence (FI) will improve with the individual-finger training because individual-finger training encourages independent finger control (Hypothesis 2). Finally, based on the findings of previous studies (Bilodeau, Keen, Sweeney, Shields, & Enoka, 2000; Kornatz, Christou, & Enoka, 2005; Laidlaw et al., 1999), we hypothesized that finger ST will increase finger strength and performance (Hypothesis 3). 314 Shim et al. Methods Subjects Forty young, healthy right-handed subjects were recruited for the study. Ten subjects were assigned into four groups, each group with different training protocols. Seven subjects discontinued the study before completing the study, and 33 subjects (23.0 ± 2.9 years; gender matched) completed the entire training protocol. Of the subjects that completed the study, Group 1 trained four fingers at the same time (G1: n = 7), Group 2 trained individual fingers without restriction of nontraining fingers (G2: n = 8), and Group 3 trained individual fingers with restriction of nontraining finger movements (G3: n = 10). The control group did not endure a training program (G0: n = 8). The group assignments were performed so the average four-finger maximal isometric force was similar across groups. All subjects gave informed consent based on the procedures approved by the University of Maryland’s Internal Review Board. Training and Experimental Setups For finger training, we used a pulley training system (Figure 1A). A cable, which ran through the central groove of the pulley, was connected to a vertically hanging load on one side and aluminum thimble(s) on the other side. The thimble(s) were designed to traverse the horizontal plane as the training fingers pulled the hanging load against gravity. For group G1, the subjects inserted all four fingers in four individual thimbles and pulled the thimbles against the hanging load, whereas for groups G2 and G3, the subjects inserted one training finger inside one thimble for the specific finger training. The hand and forearm were fixed on a table using a hand-wrist brace and Velcro strap. For measuring finger synergy, independence, and strength, we used another device that included four force sensors (black rectangle in Figure 1B), with amplifiers (Models 208 M182 and 484B, Piezotronics, Inc.) for each of the four fingers (second through fifth digits). The sensors were mounted on a customized aluminum frame (14.0 × 9.0 × 1.0 cm) along four slits. The slits allowed adjustments of the sensors antero-posteriorly along the long axis of the fingers. Adjustments were made according to the individual hand and finger sizes of the subjects. Adjacent slits were separated medio-laterally by 20 mm. The frame for each hand was attached to a large aluminum panel (21.0 × 16.0 × 2.0 cm) with a vertical slit (14.0 cm), which allowed the frame two degrees of freedom: one for vertical translation and the other for rotation about an axis perpendicular to the aluminum panel. A C-shaped aluminum thimble was attached to the bottom of each sensor. The frame was tilted at 25° with respect to the antero-posterior axis such that all finger joints (distal interphalangeal, proximal interphalangeal, and metacarpo-phalangeal joints) were slightly flexed when the distal phalanges were positioned inside the thimbles. After the position adjustments, the frame was mechanically fixed to the panel using a nut–bolt structure. Signals from the sensors were conditioned, amplified, and digitized at 100 Hz using a 16-bit A/D board (PCI 6034E, National Instruments Corp.) and a custom software program made in LabVIEW (LabVIEW 7.1, National Instruments Corp.). Figure 1 — Training and experimental settings. (A) The setup for the index finger training. The hand, wrist, and forearm were immobilized with a plastic hand brace and Velcro. The hand brace was mechanically fixed to the table, and the brace firmly held both the palm and the dorsal sides of the hand. For training, the subject used either individual fingers or all four fingers to pull against the weight, which was attached to the pulley system. (B) The experimental settings for the right hand: two-directional (tension and compression) sensors (black rectangles) were attached to an aluminum frame, and the C-shaped thimbles were attached to the bottom of the sensors. The subject inserted the distal phalange of each finger in the thimbles. The sensor positions were adjustable along the aluminum frame. (C) The subject watched the computer screen to perform a task while sitting in a chair. 315 316 Shim et al. A desktop computer (Dimension 4700, Dell, Inc.) with a 19-in. monitor was used for data acquisition. The single-finger force (for the task finger) or the total of allfour finger forces applied on the sensors was displayed on the monitor as online visual feedback. After recording the forces, the data were processed and analyzed in MatLAB (MatLAB 7, MathWorks, Inc.). The force data were digitally low-pass filtered with a second-order, zero-lag Butterworth filter at 25 Hz cutoff frequency (Shim, Olafsdottir, et al., 2005; Winter, 1990). Training and Experimental Procedure All subjects sat in a chair and positioned the right forearm on a table. The subjects pronated the forearm about 90°, such that the palm of the right hand faced left. The forearm was secured by Velcro straps. The distal phalange(s) of the task finger(s) were positioned inside the thimble(s). All subjects performed 6 sets of 10 repetitions at 70% of their one repetition maximum (1-RM) for each session. Each repetition of finger training involved finger flexion and extension about the metacarpophalangeal joints. G3 subjects were asked to self-restrict motions of the nontraining fingers. There were three sessions of training per week (Kornatz et al., 2005). 1-RMs of task finger(s) were tested and readjusted every two weeks to accommodate training adaptations (e.g., a decrease in training response to a constant stimulation or resistance) and provide a constant training overload (Zatsiorsky & Kraemer, 2006). For testing, all subjects sat in a chair facing a computer screen with their shoulder abducted 35° in the frontal plane and elbow flexed 45° in the sagittal plane, such that the forearm was parallel to the frame (Figure 1B). The forearms rested on the customized wrist-forearm braces fixed to a wooden panel (29.8 × 8.8 × 3.6 cm). Velcro straps were used to avoid forearm and wrist movements. The subjects were asked to rest the distal phalange of each finger in a thimble such that all joints were slightly flexed (Figure 1A). There were two main tests: maximal voluntary force (MVF) production and constant force production. Subjects performed five conditions for the isometric MVF test: four single-finger conditions and one fourfinger condition using the right hand. Subjects were shown a horizontal bar on the screen as force feedback for MVF tasks, and the bar moved vertically downward when subjects pressed the sensors. One trial was performed for each condition of the MVF test. Different conditions were presented to the subjects in a pseudorandomized order, with a 1-min interval between consecutive conditions. For the constant force-production test, a fixed horizontal line, which represented 20% of the four-finger MVF value for a particular subject, was shown on the computer screen as the force profile template. The actual force produced on the sensor, by the subject, was also shown on the computer screen as a different color for force feedback. The line representing the force feedback moved vertically upward and downward as the four-finger force increased and decreased, respectively. Subjects were asked to produce four-finger pressing forces to match the force profile template over a 10-s interval. Each subject performed 12 trials with at least 1-min intervals between trials and 2-min intervals between tasks. The tests were conducted every 2 weeks. Strength Training and Multifinger Coordination 317 Data Processing Maximal Voluntary Force (MVF). The peak magnitude of four-finger force was used as the MVF, which was used to estimate finger strength. Finger Independence (FI). For each MVF trial of the single-finger flexion force- production task, the nontask fingers produced forces. This phenomenon has been called finger force enslaving (Zatsiorsky, Li, & Latash, 2000). The forces produced by the nontask fingers have been called enslaving forces and are usually expressed as a percentage of the task-finger force. Because finger force enslaving is opposite to FI, FI was calculated by subtracting the averaged value of enslaving forces from 100% (Equation 1). FI = 100% − n n j −1 i =1 ∑ [100% × ∑ (F ij i / Fmax ) / (n − 1)] (1) n ij where i ≠ j, n = 4, F is the maximal force produced by the finger i, and F is the involuntary force produced by the nontask finger i, during the j finger MVF task. i max Constant Error (CE). The average absolute difference between the time trajectory of the four-finger total force and target force during the constant force-production task was calculated between the fifth and eighth second of each trial. The values were averaged across all 12 trials and normalized by the four-finger MVF (Equation 2). (2) where F(t) is the time trajectory of four-finger force, Ftarget is the target force (20% of four-finger MVF), ∆t is the time in seconds with which the analysis was performed (∆t = 3), n is the total number of trials (n = 12), and MVF is the maximal voluntary force of four fingers. Uncontrolled Manifold (UCM) Analysis. For quantifying force and moment synergy, subsequent analyses were done under the framework of the UCM hypothesis (Latash, Scholz, Danion, & Schoner, 2001; Schöner, 1995). As described earlier, a voluntary command to produce force by the task finger leads to an involuntary force production by nontask fingers. This phenomenon is called finger force enslaving and can be written in the form of a 4 × 4 matrix [E]. The elements of the enslaving matrix were computed from the regression analysis between finger forces during individual-finger MVF tasks. The diagonal and nondiagonal elements of the matrix represent the task-finger force changes and nontask-finger force changes, normalized by the changes in total force of four fingers, respectively. The four-dimensional (i.e., four-finger) vector F of finger force data, after subtracting the mean values, was converted into finger force modes m, the hypothetical commands by the central controller to the fingers to generate force (Danion et al., 2003; Equation 3). Thus, a four-dimensional force mode space was generated. 318 Shim et al. m = [E]–1F (3) Change in performance variable (dPV), expressed in terms of modes, can be written dPV = [R] × [E] × m, (4) where the elements of the 1 × 4 matrix [R] depend on the performance variable (PV): the elements are unity when the total force of four fingers is stabilized (i.e., when the error between the target motor outcome and produced motor outcome is minimized) by individual finger forces (“force stabilization hypothesis”). The elements are the moment arms of each finger, with respect to the longitudinal axis, which pass between the middle and the ring finger when the total moment is stabilized by individual-finger moments of force (“moment stabilization hypothesis”; see Kang et al. [2004] and Olafsdottir et al. [2007] for detailed computations). The hypotheses were tested with respect to two performance variables, total force and the total moment about the longitudinal axes of the hand. For a stable value of performance variable, dPV = 0, and each manifold (e.g., a vector space generated by all the combinations of finger force modes that do not change the performance variable) was approximated linearly by the null space spanned by the basis vector e (Equation 5). 0 = [R] × [E] × e (5) Thus, two separate manifolds were computed in the finger mode space: one related to total force value and the other related to the total moment of force value. The total variance (VTOT) at each time interval across all the trials was resolved into two components. The mode vectors m from each interval were resolved into their projection parallel and perpendicular to both the force-stabilization manifold and moment-stabilization manifold. The squared length of the deviation vector lying within the UCM is called variability within the UCM (VUCM) in the subsequent analysis. The direction along the manifold is called the “uncontrolled” direction because the component of variability parallel to this manifold does not affect the resultant PV. The squared length of the deviation vector perpendicular to the UCM is called variability orthogonal to the UCM (VORT). This component of variability changes the resultant PV. These two components were first divided by their degrees of freedom. The synergy index (∆V) was computed by normalizing the difference of these two components by the total variance per degree of freedom (Equation 6). ∆V = [(VUCM/3 – VORT/1)]/ [(VUCM + VORT)/4] (6) Thus, positive values of ∆V correspond to greater VUCM than VORT, reflecting the synergy between different modes to stabilize a particular performance variable (i.e., the individual modes would covary to produce a stable value of the performance variable being investigated). On the other hand, negative values of ∆V would indicate destabilization of the performance variable by the interactions between modes. ∆V was calculated for force stabilization (∆Vforce) and moment stabilization (∆Vmoment). ∆V was averaged between the fifth and eighth seconds and averaged across all 12 trials. Strength Training and Multifinger Coordination 319 Statistics Two-way ANOVAs were performed with the between-subject factor of GROUP [4 levels: G0 (control group), G1 (four-finger simultaneous training), G2 (individualfinger training without restriction), and G3 (individual-finger training with restriction of nontraining finger movements)] and the within-subject factor of SESSION (4 levels: S0, S1, S2, and S3 with 2-week intervals between consecutive testing sessions). The critical value for significant difference was set at p = .05. When appropriate, Bonferonni corrections were used for multiple comparisons. ShapiroWilk test and Kolmogorov-Smirnov test showed that the ANOVA assumption of normality was not violated (p > .05). Levene’s homogeneity test showed that the homogeneity assumption was not violated (p > .05). Results Uncontrolled Manifold (UCM) Analysis In UCM analysis, an increase in ∆V is associated with the strengthening of multifinger synergies for a specific performance variable, such as the resultant force or resultant moment of force. UCM analysis confirmed our first hypothesis that neuromuscular adaptations would improve the moment synergy and force synergy. ∆Vforce increased significantly with training in G3, while remaining unchanged in G0, G1, or G2 (Figure 2A). Significant increases of ∆Vforce in G3 were observed after 2 weeks of individual-finger training, and the increase was more pronounced after 6 weeks, indicating improved force-stabilizing synergies. These findings were supported by a two-way ANOVA, which showed a significant GROUP × SESSION interaction [F(9, 87) = 2.1, p < .05] and GROUP effect [F(3, 29) = 5.2, p < .01], but not a significant SESSION effect [F(3, 87) = 2.2, p = .09]. ∆Vmoment increased in G1, but not in G0, G2, or G3 (Figure 2B). A significant increase of ∆Vmoment in G1 was already observed after 2 weeks of training, and the increase became larger after 4 and 6 weeks, thus indicating better moment-stabilizing synergies with training. The increase became more pronounced after 6 weeks. These findings were supported by a two-way ANOVA, which showed a significant GROUP × SESSION interaction [F(9, 87) = 3.1, p < .01], GROUP effect [F(3, 29) = 132.0, p < .001], and SESSION effect [F(3, 87) = 5.2, p = .01]. The significant improvements in ∆Vforce only for G3 and in ∆Vmoment only for G1 provide support for Hypothesis 1 (i.e., force synergy and moment synergy improve with four-finger and individualfinger training, respectively). Finger Independence (FI) All training groups (G1, G2, and G3) showed a significant decrease in FI after 6 weeks of training (Figure 3). The significant decrease in FI was observed after 2 weeks of training in G1 and G3; whereas, significant reductions were not observed until 4 weeks in G2. Moreover, the decrease of FI in G1 was significantly greater than that in G2 and G3. After the 6 weeks of training, FI of G1, G2, and G3 decreased 320 Shim et al. Figure 2 — (A) ∆Vforce and (B) ∆Vmoment during all finger pressing tasks. G0, G1, G2, and G3 represent the control group, the group of four-finger simultaneous training, the group of individual-finger training without restriction, and the group of individual-finger training with restriction to nontraining-finger movements, respectively. S0, S1, S2, and S3 represent consecutive test sessions with 2-week intervals. * signifies significant (p < .05) differences of ∆V between sessions. to 85%, 93%, and 91%, respectively, of baseline (S0) FI values for each group. These findings were supported by a two-way ANOVA, which showed a significant SESSION effect [F(3, 87) = 136.4, p < .001], GROUP effect [F(3, 29) = 3.5, p < .05], and GROUP × SESSION interaction [F(9, 87) = 10.4, p < .001]. Our second hypothesis was that FI would increase with ST. However, the decrease in FI with Strength Training and Multifinger Coordination 321 Figure 3 — Finger independence (FI) during individual-finger maximal voluntary force (MVF) production tasks. G0, G1, G2, and G3 represent the control group, the group of four-finger simultaneous training, the group of individual-finger training without restriction, and the group of individual-finger training with restriction to nontraining-finger movements, respectively. S0, S1, S2, and S3 represent consecutive test sessions with 2-week intervals. * signifies significant (p < .05) differences of FI between sessions. training, especially in G2 and G3, does not support Hypothesis 2 (i.e., FI will improve with the individual-finger training). Constant Errors (CE) All training groups (G1, G2, and G3) showed a significant decrease in finger CE after 6 weeks of training (all p < .05; Figure 4). The significant decrease in CE was first observed after 2 weeks of training in G3; whereas, this was not observed until 4 weeks of training in G1 and G2. After 4 weeks, CE in G1, G2, and G3 decreased to 85%, 93%, and 91%, respectively, of the baseline session (S0) CE values for each group. These findings were supported by a two-way ANOVA, which showed the significant SESSION effect [F(3, 87) = 130.4, p < .001] and GROUP × SESSION interaction [F(9, 87) = 9.5, p < .001]. The GROUP effect was not significant. Our third hypothesis that ST will improve the finger performance was supported by the decrease in CE. Finger Strength Figure 5 shows the changes in isometric MVF of all fingers over 6 weeks of training. All training groups (G1, G2, and G3) increased MVF significantly with training (all p < .05); whereas MVF remained relatively constant in the control group (G0). The earliest observed significant increases from baseline MVF were after 4 weeks of training in G1 and G3 and after 6 weeks in G2. When all groups were combined, MVF increased by 39% with training. These findings were supported by the significant SESSION effect [F(3, 87) = 105.6, p < .001] and GROUP × SESSION interaction [F(9, 87) = 11.9, p < .001]. The GROUP effect was not Figure 4 — Constant error (CE) during constant finger force-production tasks. G0, G1, G2, and G3 represent the control group, the group of four-finger simultaneous training, the group of individual-finger training without restriction, and the group of individual-finger training with restriction to nontraining-finger movements, respectively. S0, S1, S2, and S3 represent consecutive test sessions with 2-week intervals. * signifies significant (p < .05) differences of CE between sessions. Figure 5 — Maximal voluntary force (MVF) during all finger pressing tasks. G0, G1, G2, and G3 represent the control group, the group of four-finger simultaneous training, the group of individual-finger training without restriction, and the group of individual-finger training with restriction to nontraining-finger movements, respectively. S0, S1, S2, and S3 represent consecutive test sessions with 2-week intervals. * signifies significant (p < .05) differences of MVF between sessions. 322 Strength Training and Multifinger Coordination 323 significant. The increases in MVF values after the training support Hypothesis 3 (i.e., finger strength increases with strength training). Discussion Increases in Multifinger Coordination With Training Whereas previous studies showed that finger coordination could be changed with task-specific practice, the current study demonstrates that finger coordination, quantified by multifinger synergies using the UCM analysis, can be improved by coordination-specific ST protocols as well. The increases in ∆Vforce and ∆Vmoment found in the current study after four-finger ST and individual-finger ST suggest that adaptations of the multifinger neuromuscular system from regular resistance exercise are specific to the training protocols. The specific training protocols would yield changes to different multifinger synergies (e.g., specificity of training for multifinger synergies). Our previous study showed that older adults have deficits in stabilizing the resultant moment of finger forces (Shim, Lay, et al., 2004). The results of the current study, showing moment-stabilizing synergy with individual-finger training, suggest that individual-finger training would likely serve as a beneficial intervention for reversing these deficits in the elderly. Previous studies reported that repetitive practice at relatively low force levels can also change multifinger coordination (Kang et al., 2004; Scholz, Kang, Patterson, & Latash, 2003). For example, Kang et al. (2004) showed that after 2 days of practice in controlling relatively low magnitudes of finger force (<30% MVF), there could be an accompanied increase in coordinative actions of finger forces (∆Vforce) without affecting coordination of finger moments (∆Vmoment). It was shown that persons with Down’s syndrome, who exhibit a decreased level of multifinger force coordination when compared with normal subjects, could increase the finger force coordination (∆Vforce) with only a few days of practice (Latash, Kang, & Patterson, 2002). Considering that 2 days of nonoverloading practice is unlikely to induce structural changes of muscles, such as hypertrophy (Kamen, 2004), the changes in multifinger synergies and finger strength appear to be associated more with neurological adaptations. Although the experimental task of keeping a constant sum of finger forces did not have an explicit requirement for moment stabilization, the finger training induced an increase in moment-stabilizing synergy. In fact, moment-stabilizing synergy and force-stabilizing synergy are somewhat conflicting despite both being capable of simultaneous stabilization through motor redundancy (Latash et al., 2005). The increases in moment-stabilizing synergy induced by finger training suggest that a specific synergic action of the multifinger motor system can occur regardless of the motor task. This might have important implications on day-to-day pressing and prehension tasks with four fingers. Common pressing and prehension tasks demand more moment-stabilizing synergy as compared with force-stabilizing synergy. For example, in holding a glass of water, controlling the angular position of the glass is critical to avoid spilling, whereas any thumb and finger forces are acceptable as far as the forces prevent the slipping and crushing of the glass. However, the moments must be very precisely controlled because a small moment error will 324 Shim et al. cause immediate tilting of the glass (Latash et al., 2004). Moreover, our finding of training-induced improvements in moment-stabilization synergy supports the hypothesis that both moment-stabilizing synergy and force-stabilizing synergy can increase with finger training in a task that predominantly requires moment stabilization, such as multifinger grasping. Changes in Finger Independence and Interfinger Connection With Training Previous studies have shown that humans are not capable of moving a single finger without moving other fingers (Hager-Ross & Schieber, 2000; Li et al., 2004; Schieber & Santello, 2004) or producing a single-finger force without generating forces with other fingers (Li, Latash, & Zatsiorsky, 1998; Reilly & Hammond, 2000; Shim et al., 2007). The current study shows that both ST regimens of four fingers as a group and single fingers individually are associated with decreases in FI. The greater decrease of FI found with four-finger training as compared with single-finger training appears to be a logical outcome considering that simultaneous finger actions during four-finger training encourage positive covariations of finger forces. This is an important factor because FI is a critical aspect of manual dexterity involved in everyday tasks. For example, typing on a keyboard requires individual movements of fingers to avoid typing unwanted keys. Considering the negative effects of ST to FI, ST might not be beneficial to pianists, guitarists, or professional typists, who require independent finger movements or force production. Owing to the central and peripheral constraints for independent finger actions (see Schieber & Santello, 2004, for review), the CNS is required to produce actions of nontask fingers opposing the actions of task fingers to avoid involuntary actions of nontask fingers conflicting with the task-finger actions. For example, if a task finger is flexed, the CNS might prevent the flexion of nontask fingers by extending them. The decrease in FI after the individual-finger training does not support our hypothesis that stimulating the CNS to practice nontask-finger actions opposite to the task-finger actions would increase independence of fingers. The mechanism for the training-induced decrease in FI is unclear and beyond the scope of this investigation. However, strength (MVF) appears to be related to independent actions of the finger. Shinohara, Latash, and Zatsiorsky (2003) showed that the increase in finger MVF was associated with an increase in finger enslaving (e.g., a decrease in FI). Moreover, our recent study in children also showed that FI decreases with greater finger strength (Shim et al., 2007). In addition, a previous study showed that after 2 days of applying relatively low magnitudes of finger forces, there was both an increase in finger strength and a decrease in FI (Kang et al., 2004). The changes in FI observed in the current study and other studies may be induced by neurological changes in the hand neuromechanical system, especially during the early stages of nonoverloaded practice and overloaded (i.e., strength training) conditions. An interfinger connection matrix or enslaving matrix (i.e., a 4 × 4 matrix whose diagonal elements are task-finger forces and other elements are nontaskfinger forces) has been used in previous studies to investigate kinetic and kinematic interactions between fingers and the CNS’s control on them (Danion et al., 2003; Scholz, Danion, Latash, & Schoner, 2002; Scholz et al., 2003). The previous studies considered the enslaving matrix to be a constant matrix. However, the current study Strength Training and Multifinger Coordination 325 showed that both the diagonal elements (i.e., finger strength) and nondiagonal elements (i.e., interfinger connections) can be plastic with training while demonstrating the increases of task-finger forces (i.e., MVF increases) and the disproportionate increases of nontask-finger forces (i.e., FI decreases). Previous studies showed that the interfinger connection matrix is dependent on manipulation tasks (e.g., flexion versus extension) and interfinger connection changes with human development (e.g., children versus adults; Shim et al., 2007). Thus, the UCM analysis of multifinger force-production tasks requires adjustments to the interfinger connection matrix as practice, training, and development progress and as different manipulation tasks are employed. Changes in synergies can depend on both changes in the force modes and changes in force enslaving. Because the changes of FI values over training were similar across all training groups, the changes in force synergies appear to be contributed more by changes in the force modes. Finger Strength and Performance When nontask fingers are required to avoid involuntary movements as in the G3 group, as compared with the G2 group, the task-finger muscles should produce a greater force to overcome the enslaving effects on the task finger caused by nontask-finger actions. Thus, one may expect to find greater increases from training effects on MVF in the nontask-finger movement-restriction condition than the nonrestriction condition. However, our results showed that all training conditions provided very similar responses in finger strength increases. Identifying the exact role of finger extensors is beyond the scope of this study, but the finger extensor muscle activities might have contributed to counteract the enslaving effect during training. A previous study on knee extensor ST showed that the training of the agonist muscle group (quadriceps) induced a 20% decrease in electromyographic activities of the antagonist muscle group (biceps femoris; Carolan & Cafarelli, 1992). Thus, a future study on electromyographic activities of finger flexors and extensors might provide further insight into the role of finger extensors in the training effects of the finger flexors. The complex biomechanical structures and tendon connections of the hand and forearm muscles pose a technical difficulty in monitoring individual finger muscles. Although data from the current study cannot directly determine the mechanism(s) for increases in finger strength with training, the significant improvements in strength after only 2 weeks of training indicate initial neurological rather than morphological adaptations. Others have attributed strength gains to both neural adaptations of the CNS (e.g., changes in motor-unit recruitment and firing frequency; Carroll, Riek, & Carson, 2001; Macefield, Fuglevand, & Bigland-Ritchie, 1996; Thomas, Bigland-Richie, & Johansson, 1991) and muscle morphological adaptations (e.g., changes in contractile elements and muscle architectures; Folland & Williams, 2007; Narici et al., 1996; Shoepe, Stelzer, Garner, & Widrick, 2003; Widrick et al., 2003). However, neurological adaptations are more pronounced during the early stages of training (Kamen, 2004). The increase in finger motor-task performance with ST has been reported in several previous studies (Bilodeau et al., 2000; Kornatz et al., 2005; Laidlaw et al., 1999). The decrease in force errors can also be considered an improvement to the performance of a multifinger motor system for controlling finger force. Shinohara 326 Shim et al. et al. (2003) proposed the “strength-dexterity trade-off hypothesis” by showing that FI decreases as strength increases. However, this assumption was based solely on the FI change with ST. The current study also observed a decrease in FI with an increase in finger strength. However, our finding that ST induces an increase in strength and force-control performance (e.g., a decrease in CE and increase in multifinger synergy) undermines the strength-dexterity trade-off hypothesis. Moreover, the synergic interactions of multiple fingers, reflected by ∆Vforce and ∆Vmoment, increased or remained constant after ST, despite the decrease in FI. Negative and positive covariations between finger forces are associated with stabilization and destabilization of a constant resultant force, respectively. The decrease in FI found in this study can be interpreted as an increase in positive covariations between finger forces, which might interfere with the overall performance of all finger forces. Despite the increase in positive covariations between finger forces, we found increases in ∆Vforce (increase in negative covariance) and multifinger force-control performance after training in G3. It appears that an increase in synergic actions between finger forces had a greater impact on multifinger performance than the impact from the decrease in FI. Two muscle groups contribute to finger-flexion actions: extrinsic muscles that have common origins with insertions into multiple fingers (e.g., flexor digitorum profundus and flexor digitorum superficialis) and intrinsic muscles that have single origins inserting into single fingers (i.e., lumbricals and interossei). The extrinsic muscles have insertions into the distal phalanges, whereas intrinsic muscles have insertions into the proximal phalanges (Landsmeer & Long, 1965; Landsmeer, 1955; Li, Zatsiorsky, & Latash, 2000). This causes differential uses of intrinsic and extrinsic muscles depending on the external force application points (Li et al., 2000; Shinohara et al., 2003). The external forces were applied at the distal phalanges during finger flexion about the metacarpophalangeal joints, which caused activations of extrinsic muscles. The decrease in FI appears to be caused by the extrinsic muscles with multifinger insertions because activations of the flexor digitorum profundus and flexor digitorum superficialis induce simultaneous kinetic and kinematic actions of all fingers. Considering the decreased FI after ST at the finger tips (i.e., extrinsic muscle training) found in the current study, intrinsic muscle training might be more beneficial for people who require highly independent finger actions, such as those who play keyboard and string instruments and professional typists. In the future, we intend to investigate the effects of intrinsic muscle ST on finger independence. Acknowledgment The authors would like to thank Patrick Bengero and Emma Scott for their help on recruiting and training subjects. Support for this research was provided by a grant to the University of Maryland from the Howard Hughes Medical Institute Undergraduate Science Education Program. References Bilodeau, M., Keen, D.A., Sweeney, P.J., Shields, R.W., & Enoka, R.M. (2000). Strength training can improve steadiness in persons with essential tremor. Muscle & Nerve, 23(5), 771–778. Strength Training and Multifinger Coordination 327 Carolan, B., & Cafarelli, E. (1992). Adaptations in coactivation after isometric resistance training. Journal of Applied Physiology, 73(3), 911–917. Carroll, T.J., Riek, S., & Carson, R.G. (2001). Neural adaptations to resistance training: Implications for movement control. Sports Medicine (Auckland, N.Z.), 31(12), 829–840. Danion, F., Schoner, G., Latash, M.L., Li, S., Scholz, J.P., & Zatsiorsky, V.M. (2003). A mode hypothesis for finger interaction during multi-finger force-production tasks. Biological Cybernetics, 88(2), 91–98. Dons, B., Bollerup, K., Bonde-Petersen, F., & Hancke, S. (1979). The effect of weightlifting exercise related to muscle fiber composition and muscle cross-sectional area in humans. European Journal of Applied Physiology and Occupational Physiology, 40(2), 95–106. Engel, K.C., Flanders, M., & Soechting, J.F. (1997). Anticipatory and sequential motor control in piano playing. Experimental Brain Research, 113(2), 189–199. Enoka, R.M. (1997). Neural strategies in the control of muscle force. Muscle & Nerve. Supplement, 5, S66–S69. Fish, J., & Soechting, J.F. (1992). Synergistic finger movements in a skilled motor task. Experimental Brain Research, 91(2), 327–334. Flanders, M., & Soechting, J.F. (1992). Kinematics of typing: Parallel control of the two hands. Journal of Neurophysiology, 67(5), 1264–1274. Folland, J.P., & Williams, A.G. (2007). The adaptations to strength training: Morphological and neurological contributions to increased strength. Sports Medicine (Auckland, N.Z.), 37(2), 145–168. Hager-Ross, C., & Schieber, M.H. (2000). Quantifying the independence of human finger movements: Comparisons of digits, hands, and movement frequencies. The Journal of Neuroscience, 20(22), 8542–8550. Kamen, G. (2004). Neural issues in the control of muscular strength. Research Quarterly for Exercise and Sport, 75(1), 3–8. Kang, N., Shinohara, M., Zatsiorsky, V.M., & Latash, M.L. (2004). Learning multi-finger synergies: An uncontrolled manifold analysis. Experimental Brain Research, 157(3), 336–350. Keen, D.A., Yue, G.H., & Enoka, R.M. (1994). Training-related enhancement in the control of motor output in elderly humans. Journal of Applied Physiology, 77(6), 2648–2658. Kornatz, K.W., Christou, E.A., & Enoka, R.M. (2005). Practice reduces motor unit discharge variability in a hand muscle and improves manual dexterity in old adults. Journal of Applied Physiology, 98(6), 2072–2080. Laidlaw, D.H., Kornatz, K.W., Keen, D.A., Suzuki, S., & Enoka, R.M. (1999). Strength training improves the steadiness of slow lengthening contractions performed by old adults. Journal of Applied Physiology, 87(5), 1786–1795. Landsmeer, J.M., & Long, C. (1965). The mechanism of finger control, based on electromyograms and location analysis. Acta Anatomica, 60(3), 330–347. Landsmeer, J.M.F. (1955). Anatomical and functional investigations on the articulation of the human fingers. Acta Anatomica, 25, 1–69. Latash, M.L., Kang, N., & Patterson, D. (2002). Finger coordination in persons with down syndrome: Atypical patterns of coordination and the effects of practice. Experimental Brain Research, 146(3), 345–355. Latash, M.L., Scholz, J.F., Danion, F., & Schoner, G. (2001). Structure of motor variability in marginally redundant multifinger force production tasks. Experimental Brain Research, 141(2), 153–165. Latash, M.L., Shim, J.K., Gao, F., & Zatsiorsky, V.M. (2004). Rotational equilibrium during multi-digit pressing and prehension. Motor Control, 8(4), 392–404. Latash, M.L., Shim, J.K., Smilga, A.V., & Zatsiorsky, V.M. (2005). A central back-coupling hypothesis on the organization of motor synergies: A physical metaphor and a neural model. Biological Cybernetics, 92(3), 186–191. 328 Shim et al. Li, Z.M., Dun, S., Harkness, D.A., & Brininger, T.L. (2004). Motion enslaving among multiple fingers of the human hand. Motor Control, 8(1), 1–15. Li, Z.M., Latash, M.L., & Zatsiorsky, V.M. (1998). Force sharing among fingers as a model of the redundancy problem. Experimental Brain Research, 119(3), 276–286. Li, Z.M., Zatsiorsky, V.M., & Latash, M.L. (2000). Contribution of the extrinsic and intrinsic hand muscles to the moments in finger joints. Clinical Biomechanics (Bristol, Avon), 15(3), 203–211. Macefield, V.G., Fuglevand, A.J., & Bigland-Ritchie, B. (1996). Contractile properties of single motor units in human toe extensors assessed by intraneural motor axon stimulation. Journal of Neurophysiology, 75(6), 2509–2519. Narici, M.V., Hoppeler, H., Kayser, B., Landoni, L., Claassen, H., Gavardi, C., et al. (1996). Human quadriceps cross-sectional area, torque and neural activation during 6 months strength training. Acta Physiologica Scandinavica, 157(2), 175–186. Nowak, D.A., & Hermsdorfer, J. (2003). Sensorimotor memory and grip force control: Does grip force anticipate a self-produced weight change when drinking with a straw from a cup? The European Journal of Neuroscience, 18(10), 2883–2892. Olafsdottir, H., Zhang, W., Zatsiorsky, V.M., & Latash, M.L. (2007). Age-related changes in multifinger synergies in accurate moment of force production tasks. Journal of Applied Physiology, 102(4), 1490–1501. Oliveira, M.A., Shim, J.K., Loss, J.F., Petersen, R.D., & Clark, J.E. (2006). Effect of kinetic redundancy on hand digit control in children with DCD. Neuroscience Letters, 410(1), 42–46. Rearick, M.P., Casares, A., & Santello, M. (2003). Task-dependent modulation of multi-digit force coordination patterns. Journal of Neurophysiology, 89(3), 1317–1326. Reilly, K.T., & Hammond, G.R. (2000). Independence of force production by digits of the human hand. Neuroscience Letters, 290(1), 53–56. Reisman, D.S., & Scholz, J.P. (2006). Workspace location influences joint coordination during reaching in post-stroke hemiparesis. Experimental Brain Research, 170(2), 265–276. Sale, D.G. (1988). Neural adaptation to resistance training. Medicine and Science in Sports and Exercise, 20(Suppl. 5) S135–S145. Schieber, M.H., & Poliakov, A.V. (1998). Partial inactivation of the primary motor cortex hand area: Effects on individuated finger movements. The Journal of Neuroscience, 18(21), 9038–9054. Schieber, M.H., & Santello, M. (2004). Hand function: Peripheral and central constraints on performance. Journal of Applied Physiology, 96(6), 2293–2300. Scholz, J.P., Danion, F., Latash, M.L., & Schoner, G. (2002). Understanding finger coordination through analysis of the structure of force variability. Biological Cybernetics, 86(1), 29–39. Scholz, J.P., Kang, N., Patterson, D., & Latash, M.L. (2003). Uncontrolled manifold analysis of single trials during multi-finger force production by persons with and without down syndrome. Experimental Brain Research, 153(1), 45–58. Schöner, G. (1995). Recent developments and problems in human movement science and their conceptual implications. Ecological Psychology, 7(4), 291–314. Shim, J.K. (2005). Rotational equilibrium control in multi-digit human prehension. University Park: The Pennsylvania State University. Shim, J.K., Kim, S.W., Oh, S.J., Kang, N., Zatsiorsky, V.M., & Latash, M.L. (2005). Plastic changes in interhemispheric inhibition with practice of a two-handforce production task: A transcranial magnetic stimulation study. Neuroscience Letters, 374(2), 104–108. Shim, J.K., Latash, M.L., & Zatsiorsky, V.M. (2003). Prehension synergies: Trial-to-trial variability and hierarchical organization of stable performance. Experimental Brain Research, 152, 173–184. Shim, J.K., Latash, M.L., & Zatsiorsky, V.M. (2004). Prehension synergies in three dimensions. Journal of Neurophysiology, 93, 766–776. Strength Training and Multifinger Coordination 329 Shim, J.K., Latash, M.L., & Zatsiorsky, V.M. (2005). Prehension synergies: Trial-to-trial variability and principle of superposition during static prehension in three dimensions. Journal of Neurophysiology, 93(6), 3649–3658. Shim, J.K., Lay, B.S., Zatsiorsky, V.M., & Latash, M.L. (2004). Age-related changes in finger coordination in static prehension tasks. Journal of Applied Physiology, 97(1), 213–224. Shim, J.K., Olafsdottir, H., Zatsiorsky, V.M., & Latash, M.L. (2005). The emergence and disappearance of multi-digit synergies during force-production tasks. Experimental Brain Research, 164(2), 260–270. Shim, J.K., Oliveira, M.A., Hsu, J., Huang, J., Park, J., & Clark, J.E. (2007). Hand digit control in children: Age-related changes in hand digit force interactions during maximum flexion and extension force production tasks. Experimental Brain Research, 176(2), 374–386 . Shinohara, M., Latash, M.L., & Zatsiorsky, V.M. (2003). Age effects on force produced by intrinsic and extrinsic hand muscles and finger interaction during mvc tasks. Journal of Applied Physiology, 95(4), 1361–1369. Shinohara, M., Scholz, J.P., Zatsiorsky, V.M., & Latash, M.L. (2004). Finger interaction during accurate multi-finger force production tasks in young and elderly persons. Experimental Brain Research, 156(3), 282–292. Shoepe, T.C., Stelzer, J.E., Garner, D.P., & Widrick, J.J. (2003). Functional adaptability of muscle fibers to long-term resistance exercise. Medicine and Science in Sports and Exercise, 35(6), 944–951. Thomas, C.K., Bigland-Richie, B., & Johansson, R.S. (1991). Force-frequency relationships of human thenar motor units. Journal of Neurophysiology, 65(6), 1509–1516. Tracy, B.L., Byrnes, W.C., & Enoka, R.M. (2004). Strength training reduces force fluctuations during anisometric contractions of the quadriceps femoris muscles in old adults. Journal of Applied Physiology, 96(4), 1530–1540. Widrick, J.J., Maddalozzo, G.F., Lewis, D., Valentine, B.A., Garner, D.P., Stelzer, J.E., et al. (2003). Morphological and functional characteristics of skeletal muscle fibers from hormone-replaced and nonreplaced postmenopausal women. The Journals of Gerontology. Series A, Biological Sciences and Medical Sciences, 58(1), 3–10. Winter, D.A. (1990). Biomechanics and motor control of human movement (2nd ed.). New York: John Wiley & Sons Inc. Zatsiorsky, V.M., & Kraemer, W.J. (2006). Science and practice of strength training (2nd ed.). Champaign, IL: Human Kinetics. Zatsiorsky, V.M., Li, Z.M., & Latash, M.L. (2000). Enslaving effects in multi-finger force production. Experimental Brain Research, 131(2), 187–195.