Survey
* Your assessment is very important for improving the work of artificial intelligence, which forms the content of this project
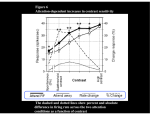
J Neurophysiol 95: 3309 –3310, 2006; doi:10.1152/jn.00163.2006. Editorial Focus Perception, Memory, and Action in Frontal and Parietal Cortex. Focus on “Selection and Maintenance of Saccade Goals in the Human Frontal Eye Fields” Vincent P. Ferrera and Jack Grinband Mahoney Center for Brain and Behavior, Center for Neurobiology and Behavior, Department of Psychiatry, Columbia University and New York State Psychiatric Institute, New York, New York Address for reprint requests and other correspondence: V. Ferrera, Mahoney Center for Brain and Behavior, Center for Neurobiology and Behavior, Department of Psychiatry, Columbia University, New York, NY (E-mail: [email protected]). www.jn.org response. A previous study (Connolly et al. 2002) showed that motor set modulated activity in FEF but not IPS. Other interpretations are possible, but the most likely of these are ruled out by the data. For example, one can imagine that during the first delay, subjects covertly prepare all four movements instead of (or in addition to) simply remembering the stimulus locations. In this case, one might infer that IPS is more closely involved in movement planning because it has the greatest activation during the first delay. This explanation can be rejected because IPS activation fell to baseline levels during the second delay despite the continued need for motor response preparation. FEF activation remained significantly above baseline. A reduction in the number of planned movements from four to one might account for the reduced activation seen in both FEF and IPS but not the observed inversion in the relative strength of activation between the two areas. The fact that the number of locations to be remembered changes once the target is cued raises the issue of whether an overall reduction in the level of BOLD activation between the first and second delays is due to response selection, per se, as opposed to the corresponding reduction in memory load. To address this, Curtis and D’Esposito looked for lateralized activation in IPS and FEF. It is well known that the majority of neurons in the parietal and prefrontal cortex prefer contralateral stimuli and/or contraversive movements (Blatt et al. 1990; Funahashi et al. 1990). Because the initial stimulus array covered both visual hemifields, the first delay interval showed bilateral brain activity that was equal across the two hemispheres. However, after presenting the target cue, activity in the second delay interval became strongly lateralized in the hemisphere contralateral to the position of the target. The lateralized IPS activation was short-lived, but FEF activation was sustained and the degree of lateralization increased over time until the response was executed. Thus the lateralization of activity in FEF increased even though there was only a single item in memory. More importantly, lateralized FEF activation just prior to the motor response predicted the direction of the eye movement; IPS activation did not. Neurophysiological studies in monkey FEF and LIP support the visual-motor distinction. These studies have revealed neurons with visual, movement and mixed visual-movement activity (Andersen et al. 1990; Barash et al. 1991; Bruce and Goldberg 1985). Visual and visual-movement neurons are found in both areas (Chafee and Goldman-Rakic 1998); however, pure movement neurons without visual responses are common in FEF but rare in LIP. Movement-related activity in parietal cortex tends to evolve slowly and is best demonstrated in delayed-response tasks. If monkeys are required to make an 0022-3077/06 $8.00 Copyright © 2006 The American Physiological Society 3309 Downloaded from http://jn.physiology.org/ by 10.220.33.1 on July 31, 2017 Visually guided behavior engages posterior parietal and lateral prefrontal cortex, yet the unique contributions of these brain regions are strongly contested. Recent experiments by Curtis and D’Esposito use functional magnetic resonance imaging to differentiate the roles of parietal and prefrontal cortex with respect to visual working memory and movement planning. The notion of functional specialization in the visual system has a long if not uncontroversial history (Goodale and Milner 1992; Scheider 1969; Trevarthen 1968; Ungerleider and Mishkin 1982). One current area of contention is the division of labor between posterior parietal (PPC) and lateral prefrontal cortex (PFC). Both are involved in visual-motor transformations, but is one inherently more “visual” and the other more “motor”? In this issue of Journal of Neurophysiology (p. 3923–3927), a functional imaging study by Curtis and D’Esposito (2006) shows that PPC and PFC perform a kind of dance during visual-motor behavior with first one area taking the lead and then the other. Their results suggest that PPC is more active during the encoding and storage of visual stimuli in working memory, but PFC becomes more active during response preparation and execution. These results lend support to the view that parietal cortex contains a general purpose visual map (Gottlieb et al. 1998), whereas prefrontal cortex is more action oriented. Curtis and D’Esposito’s experiment used a delayed-response task that evoked activity in a region of the precentral sulcus likely to be the human frontal eye field (FEF), and a region in the intraparietal sulcus (IPS) likely to be the human equivalent of monkey lateral intraparietal area (LIP). Subjects were required to remember an array of four stimulus locations during an extended delay period. At the end of this delay, a cue identified one of the four locations as the saccadic goal. A second delay interval required the maintenance of the target location until a movement execution cue was given. IPS and FEF showed opposite patterns of blood-oxygen-level-dependent (BOLD) activitation during the two delay intervals. During the “visual memory” delay, activation was stronger in IPS than in FEF. However, during the “movement preparation” delay, the pattern of activation reversed. These results support the idea that neural activity in IPS is primarily influenced by the processing and storage of visual stimuli, whereas FEF is more heavily recruited during the preparation of the motor Editorial Focus 3310 REFERENCES Andersen RA, Bracewell RM, Barash S, Gnadt JW, and Fogassi L. Eye position effects on visual, memory, and saccade-related activity in areas LIP and 7a of macaque. J Neurosci 10: 1176 –1196, 1990. Andersen RA, Brotchie PR, and Mazzoni P. Evidence for the lateral intraparietal area as the parietal eye field. Curr Opin Neurobiol 2: 840 – 846, 1992. Barash S, Bracewell RM, Fogassi L, Gnadt JW, and Andersen RA. Saccade-related activity in the lateral intraparietal area. I. Temporal properties; comparison with area 7a. J Neurophysiol 66: 1095–1108, 1991. Blatt GJ, Andersen RA, and Stoner GR. Visual receptive field organization and cortico-cortical connections of the lateral intraparietal area (area LIP) in the macaque. J Comp Neurol 299: 421– 445, 1990. Bruce CJ and Goldberg ME. Primate frontal eye fields. I. Single neurons discharging before saccades. J Neurophysiol 53: 603– 635, 1985. J Neurophysiol • VOL Bruce CJ, Goldberg ME, Bushnell MC, and Stanton GB. Primate frontal eye fields. II. Physiological and anatomical correlates of electrically evoked eye movements. J Neurophysiol 54: 714 –734, 1985. Chafee MV and Goldman-Rakic PS. Matching patterns of activity in primate prefrontal area 8a and parietal area 7ip neurons during a spatial working memory task. J Neurophysiol 79: 2919 –2940, 1998. Connolly JD, Goodale MA, Menon RS, and Munoz DP. Human fMRI evidence for the neural correlates of preparatory set. Nat Neurosci 5: 1345–1352, 2002. Curtis CE and D’Esposito M. Selection and maintenance of saccade goals in the human frontal eye fields. J Neurophysiol 95: 3923–3927, 2006. Everling S and Munoz DP. Neuronal correlates for preparatory set associated with pro-saccades and anti-saccades in the primate frontal eye field. J Neurosci 20: 387– 400, 2000. Funahashi S, Bruce CJ, and Goldman-Rakic PS. Visuospatial coding in primate prefrontal neurons revealed by oculomotor paradigms. J Neurophysiol 63: 814 – 831, 1990. Goldberg ME, Bisley J, Powell KD, Gottlieb J, and Kusunoki M. The role of the lateral intraparietal area of the monkey in the generation of saccades and visuospatial attention. Ann NY Acad Sci 956: 205–215, 2002. Goodale MA and Milner AD. Separate visual pathways for perception and action. Trends Neurosci 15: 20 –25, 1992. Gottlieb J and Goldberg ME. Activity of neurons in the lateral intraparietal area of the monkey during an antisaccade task. Nat Neurosci 2: 906 –912, 1999. Gottlieb, J, Kusunoki, M, and Goldberg, M.E. The representation of visual salience in monkey parietal cortex. Nature 391: 481– 484, 1998. Moore T, Armstrong KM, and Fallah M. Visuomotor origins of covert spatial attention. Neuron 40: 671– 683, 2003. Posner MI and Petersen SE. The attention system of the human brain. Annu Rev Neurosci 13: 25– 42, 1990. Schneider GE. Two visual systems: brain mechanisms for localization and discrimination are dissociated by tectal and cortical lesions. Science 163: 895–902, 1969. Shibutani H, Sakata H, and Hyvarinen J. Saccade and blinking evoked by microstimulation of the posterior parietal association cortex of the monkey. Exp Brain Res 55: 1– 8, 1984. Snyder LH, Batista AP, and Andersen RA. Intention-related activity in the posterior parietal cortex: a review. Vision Res 40: 1433–1441, 2000. Thompson KG, Biscoe KL, and Sato TR. Neuronal basis of covert spatial attention in the frontal eye field. J Neurosci 25: 9479 –9487, 2005. Trevarthen CB. Two mechanisms of vision in primates. Psychol Forsch 31: 229 –337, 1968. Ungerleider LG and Mishkin, M. (1982). Two cortical visual systems: separation of appearance and location of objects. In: Analysis of Visual Behavior, edited by Ingle DL, Goodale MA, and Mansfield RJW. Cambridge, MA: MIT Press, 1982, p. 549 –586. 95 • JUNE 2006 • www.jn.org Downloaded from http://jn.physiology.org/ by 10.220.33.1 on July 31, 2017 immediate “anti-saccade” (a saccade directed away from the visual stimulus) when a target appears, FEF neurons are able to reliably signal the direction of the movement (Everling and Munoz 2000), but LIP neurons are not (Gottlieb et al. 1999). Finally, whereas eye movements can be elicited by stimulating either FEF or LIP, stimulation thresholds in FEF are much lower than LIP (Bruce et al. 1985; Shibutani et al. 1984). FEF and LIP are key components of a fronto-parietal network that is involved in spatial attention, working memory, and eye movements (Posner and Petersen 1990). Different labs have come to almost diametrically opposed views on the role of parietal cortex in this circuit. One group has proposed that parietal cortex represents the attentional salience of visual objects (Goldberg et al. 2002), whereas others have emphasized its role in motor planning (Snyder et al. 2000). The view that FEF is primarily “motor” has likewise been called into question by studies showing the involvement of FEF in attentional tasks (Moore et al. 2003; Thompson et al. 2005). Electrophysiologically, FEF and IPS show as many similarities as differences. The value of the Curtis and D’Esposito study is that it clearly shows, at the level of fMRI, a movement planning signal in FEF that is not present in IPS, thus providing evidence for functional specialization in the fronto-parietal circuit.