Survey
* Your assessment is very important for improving the work of artificial intelligence, which forms the content of this project
/. Embryol. exp. Morph. Vol. 35, 3, pp. 577-593, 1976
Printed in Great Britain
577
An experimental investigation into
the possible neural crest origin of pancreatic APUD
(islet) cells
By ANN ANDREW 1
From the Department of Anatomy, University of the Witwatersrand,
Johannesburg
SUMMARY
It has recently been contended that pancreatic APUD cells are neural crest derivatives. In
an experimental investigation, isotopic grafts of neural tube containing neural crest cells
were transplanted from chick and quail embryos labelled with tritiated thymidine, and from
unlabelled quail embryos, to host chick embryos at the same stage of development. Transplantations were performed at various levels between somites 5 and 24 in embryos at
6- to 24-somite stages. In operated embryos at 3£ days of incubation, the pancreatic APUD
cells were not labelled; nor did their nuclei show quail features. Migration of cells from the
graft was evidenced by the presence of quail nuclei and/or radioactive label in autoradiographs,
in spinal and sympathetic ganglia in the operated region.
It is concluded that the pancreatic APUD cells of the 3f-day-old chick embryo are not
derived from the trunk neural crest up to the level of somite 24. It is unlikely that more caudal
levels contribute, because APUD cells are already concentrated in the dorsal pancreatic bud
region at the 24-somite stage, by which time no migration of crest cells has occurred caudal
to somite 24. This conclusion probably concerns A, B and D pancreatic endocrine cells.
INTRODUCTION
It has long been generally accepted that the cells of pancreatic islets are of
endodermal origin (see for instance, Liegner, 1932): they and the exocrine cells
are thought to differentiate from common 'protodifferentiated' endoderm cells
(Pictet & Rutter, 1972). Descriptions abound of the differentiation of islet
cells from primitive cell cords, ductules, acini and/or centro-acinar cells in the
endodermal pancreatic buds in fish, amphibians, birds and mammals (Liegner,
1932; Villamil, 1942; Hard, 1944; Bencosme, 1955; Frye, 1958; Robb, 1961;
Hellman, 1966; Przbylski, 1967; Like & Orci, 1972; Pictet, Clark, Williams &
Rutter, 1972; Epple & Lewis, 1973; Schweisthal & Frost, 1973; Belsare, 1974).
The only suggestion that islet cells may be mesodermal has come from Wessels
(1968) who, in the mouse, identified islet cells first at the endoderm-mesoderm
1
Author's address: Department of Anatomy, University of the Witwatersrand Medical
School, Hospital Street, Johannesburg, South Africa.
578
A. ANDREW
interface of the pancreatic anlage. He commented that it was not clear whether
they originated in the endoderm or the mesoderm.
Other workers have postulated a developmental relationship between islet
cells and endoderm cells of the gastro-intestinal tract. Feyrter (1943) suggested
that islet cells and enterochromaffin cells might share similar progenitor cells;
Adelson (1971) regarded the protein-secreting ability common to islet and gut
endocrine cells as supporting a common endodermal origin. Recognizing the
same similarity, Pearse and his co-workers have proposed that the neural crest
is the source (Pearse, 1969; Pearse & Polak, 1971; Polak, Rost & Pearse, 1971;
Pearse, Polak & Bussolati, 1972), though leaving open the possibility that not
all the pancreatic endocrine cells are so derived (Pearse & Polak, 1971; Pearse,
1973; Pearse & Takor Takor, 1975). This proposal is contributory to the concept
that APUD cells in general are neural crest derivatives (Pearse, 1966; Pearse &
Welsch, 1968; Pearse & Takor Takor, 1975). That pancreatic islet cells of one
or other type belong to Pearse's APUD (Amine Precursor Uptake and Decarboxylation) cell series (Pearse, 1966) has been well established (Falck & Hellman, 1963; Cegrell, 1967, 1968; Cegrell, Falck & Rosengren, 1967; Legg, 1968;
Trandaburu, 1972).
Like & Orci (1972) have pointed out that already in the pancreatic anlage
there may be cells committed to become islet cells, but they commented that
there was no evidence to justify claims that these were of neural crest or even of
endodermal origin. Epple & Lewis (1973), though favouring the neural crest as
the source, also recognized that no satisfactory evidence was yet available. So
far, except in the recent work of Phelps (1975), the grounds for the various proposed sources of islet cells have been morphological observations made at
successive developmental stages. Clearly, experimental evidence is more convincing.
The present study, of which a brief account has already appeared (Andrew,
1976a), is an experimental investigation into the question of whether or not
islet cells arise from the neural crest. Lengths of neural tube and neural crest
were removed from chick embryos, and replaced by neural tube containing
neural crest marked so that it could be recognized at later stages; the differentiated pancreatic APUD (islet) cells were subsequently examined for the neural
crest marker.
MATERIALS AND METHODS
Two methods were used to render neural crest cells distinguishable after their
migration away from the neural tube. Grafts of neural tube together with
neural crest were prepared from chick embryos labelled with tritiated thymidine
(a method used by Weston, 1963) or from quail embryos. Le Douarin (1971)
has shown that the nuclei of cells of the Japanese quail, Coturnix cotumix
japonica, carry large Feulgen-positive nucleoli and are thus easily distinguished
from chick cells.
Origin of pancreatic APUD {islet) cells
579
Following in general the procedures used by Weston (1963) and Le Douarin
& Teillet (1973), grafts from labelled Black Australorp chick embryos or from
labelled or unlabelled embryos of the African quail, Coturnix cotumix africana,
were transplanted to host Black Australorp embryos in which the corresponding
segment of the neural tube had been removed. In each case, the donor and the
host embryo were at the same stage of development. The operated embryos
were incubated until 3f days old, and then treated with the amine precursor
dihydroxyphenylalanine (DOPA) in preparation for the demonstration of the
APUD reaction in sections. After photographing the APUD cells in the dorsal
pancreatic bud, autoradiographs were prepared and/or staining by the Feulgen
method was carried out, so that cells of grafted neural crest origin could be
recognized by means of radioactive label and/or quail nuclear features. Evidence
for migration of neural crest cells from the graft was sought in the presence of
marked spinal and synpathetic ganglia in the operated region.
Various preliminary and control procedures were necessary to ascertain that
(1) the African quail also has nuclei clearly distinct from chick nuclei in the
structures to be examined; (2) at the stage of sacrifice of the operated embryos,
radioactive labelling is still adequate in the grafted neural tube, spinal and
sympathetic ganglia and pancreatic APUD cells, of chick and quail embryos;
(3) at this time, cells giving the APUD reaction have differentiated in the pancreas of both species; (4) pancreatic APUD cells do not show autofluorescence;
(5) none of the structures to be examined in autoradiographs show either positive
or negative chemography; (6) the techniques for the demonstration of the
APUD reaction, of radioactive label and of nuclear characteristics do not interfere with one another; and (7) the operation does not affect the normal development of the pancreas, including its ability to synthesize dopamine from DOPA
(APUD reaction). To serve these purposes, operated embryos which had
received unlabelled chick grafts, unlabelled quail grafts, labelled chick grafts
or labelled quail grafts, as well as unoperated embryos, chick and quail,
labelled and unlabelled, were subjected to the various procedures severally or
jointly.
Eggs were incubated at 38-5 °C. All micro-surgical procedures were carried
out under sterile conditions. Chick Ringer's solution contained antibiotics as
described previously (Andrew, 1963).
Labelling of embryos
Preliminary tests showed that a dose of 10 /tc of tritiated thymidine (thymidine-6-H3, TRA 61, specific activity 5 Ci/mmol; Amersham) had no deleterious
effect on development. The isotope was injected in 100/tl Ringer's solution
through a small window in the shell, into the air-space of eggs incubated for
24 h, blunt end upwards. The window was sealed with Sellotape. Survival of
24-h embryos was better than if the isotope was injected directly on to the blastoderm. Undoubtedly some embryos were not centrally situated under the
580
A. ANDREW
air-space: these probably took up less isotope, as found by Mawhinney,
Austin & Riley (1972).
Isotope was administered 14 to 24 h before donor embryos were used for
transplantation: practically all nuclei became labelled overnight. Labelled
unoperated chick and quail embryos showed well-labelled nuclei until 5 days
of incubation.
DOPA administration and the formaldehyde-induced
fluorescence (FIF) procedure
DOPA was administered to embryos incubated for 3f days. A dose of 150 /tg
D L - D O P A (B.D.H.) dissolved in 150/tl warm Ringer's solution was injected
through a window in the shell on to the surface of the embryo. The window was
sealed and the egg returned to the incubator. (Some control embryos received
no DOPA.)
A portion of the trunk was excised, 1-1^ h later, to include the pancreas and
the operated region of the neural tube. This was left in pre-warmed Ringer's
solution containing 100/^g/ml DOPA in the incubator for up to 15 min. The
specimens were then washed in fresh pure Ringer's solution, quenched, freezedried and fixed in hot formaldehyde vapour as described elsewhere (Andrew,
1975); the duration of fixation was 1-l^h. (Some control specimens were not
subjected to vapour fixation.) The specimens were embedded in paraplast
(M.P. 58 °C) in vacuo, serially sectioned at 8 /im and dry-mounted on lightly
albumenized slides. Wax was allowed to drain off the slides overnight in an
oven at 60 °C. Slides were stored as previously (Andrew, 1975).
Sections were examined in xylol under blue light bright field illumination on
a Reichert Fluoropan microscope equipped with an HBO 50 mercury burner.
A 6 BG 12/h exciter filter and appropriate absorption filter were used.
Autoradiography
Originally, de-waxed sections were hydrated before dipping in emulsion:
subsequently it was found that a more even layer of emulsion was produced on
slides allowed to air-dry immediately before dipping. They were de-waxed,
transferred to absolute ethanol and then exposed to the air. The drying procedure had no deleterious effects on the sections.
The emulsion used was Ilford K2; the method followed that of Rogers
(1967). In a test for negative chemography, several slides in each batch were
exposed to light after dipping; dipped slides of unlabelled tissue served as tests
for positive chemography. Both procedures are essential for valid interpretation
of autoradiographs (Rogers, 1967). All the slides (including the chemography
controls) were exposed in the dark at 4 °C. An exposure of 17 days was usually
adequate. The autoradiographs were developed in D 163 (Kodak); the method
was that recommended by Rogers (1967).
Origin of pancreatic APUD {islet) cells
581
Staining
Autoradiographs of sections of control chick embryos and of operated embryos which had received chick grafts of labelled tissue were lightly stained with
haematoxylin. Autoradiographs of sectioned control quail and some chick
embryos, and of operated embryos which had received quail grafts, were
examined before staining with the Feulgen method for DNA (Pearse, 1960).
This was found necessary because the hydrolysis involved, caused removal or
re-distribution of some of the developed silver grains, as was shown on clean
slides (without sections) dipped, exposed to light and then developed and subjected to hydrolysis.
Photography
Photographs of fluorescent cells in the dorsal pancreatic bud were taken on the
fluorescence microscope with Kodak Tri-X Pan film (ASA 400) at an exposure
of 10 sees. As a rule, every second or third section showing the fluorescent cells
was photographed.
Transplantation
Segments of neural tube with accompanying neural crest cells were transplanted from radioactively labelled chick or quail embryos, unlabelled quail
embryos, and occasionally from unlabelled chick embryos, to the same somite
levels of chick embryos at the same stage of development. (Unlabelled grafts
from chick embryos served as controls for positive chemography.) The levels of
the transplant were determined on the basis of morphological studies of the
stage of neural crest formation at different somite levels in Black Australorp
chick embryos of relevant stages (Andrew, 1963). The levels selected for transplantation were those at which the crests had not yet formed as such, or at which
no migration from the crests proper had yet taken place. In a 12-somite embryo
for example, levels caudal to somite 8 fulfil these criteria; in a 16-somite embryo,
levels caudal to somite 13. In practice a safety margin of two or more somites'
width was usually allowed. In order to vary the levels transplanted, embryos
were used as donors at stages between 6 and 20 somites. The most likely level
of origin of pancreatic islet cells seemed that at which the duodenum develops,
i.e. somites 8 to 15 (Le Douarin, 1961). The levels of neural transplants were
therefore at first concentrated on these and then on adjoining levels (see Table 2).
The grafts varied from 4- to 8-somites' length: where they extended into the
post-somitic region, the future somite level at their caudal end was estimated.
The estimates were checked on sacrifice whenever the caudal end of the graft
could be identified, and were found to be accurate.
The desired length of neural tube was excised together with underlying notochord and endoderm. It was transferred to 0-1 % trypsin made up in calciumand magnesium-free Ringer's solution containing neutral red (2 x 10~4 mg/ml)
37
EMB
35
582
A. ANDREW
for 2 min at 37-5 °C, and then to warm calcium- and magnesium-free Ringer's
solution with neutral red for 2 min. Radioactively-labelled tissue was well rinsed
in 'cold' thymidine in Ringer's solution (4mg/ml). The neural tube was dissected clear of adherent tissue. More latterly, a transverse strip of blastoderm
bounded by the desired rostral and caudal levels of the graft was transferred
to a 0-15 % trypsin solution otherwise made up as above, at room temperature,
and placed in a refrigerator at 2 °C for 15 mins, according to the method used
by Le Douarin & Teillet (1973). Further steps were carried out as above. The
notochord is tightly adherent to the neural tube, so its removal sometimes
results in slitting of the neural tube. Therefore, occasionally small fragments of
notochord were left on the graft.
Likewise, in preparing the graft site in host embryos, tiny tags of the floor of
the neural tube were sometimes left behind. The graft was manoeuvred into
position in the prepared site by means of a very fine glass thread with a rounded
tip. After the window in the shell of the host egg had been sealed, it was returned
to the incubator. At one time, the eggs were placed straight away on turntables in the incubator (see below). However, when it was discovered that grafts
were sometimes dislodged, operated eggs were left stationary in the incubator
for an hour before being placed on the turntables. They remained there overnight.
During further incubation to reach a total incubation age of 3f days, the eggs
were stationary.
Rotation of operated eggs
Survival of operated embryos was vastly improved by incubating the eggs
horizontally on individual turntables which rotated alternately clockwise and
anticlockwise through 90° around a vertical axis at 8 cycles per minute. These
were a modification of the apparatus designed by Silver (1960). Probably the
movement was effective because it prevented adherence of the blastoderm to the
edges of the window.
RESULTS
Large Feulgen-positive nucleoli were evident in the cells of the neural tube,
spinal and sympathetic ganglia, and all cells of the dorsal pancreatic bud in
three 3|-day-old embryos of the African quail to which DOPA had been administered and in all of which the FIF procedure revealed fluorescent pancreatic
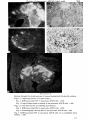
APUD cells. Fig. 2 illustrates this nuclear feature in the same pancreatic cells
shown to be fluorescent in Fig. 1. The cells in chick embryos, including those of
the pancreas and the others to be examined in this study, show only small
Feulgen-positive karyosomes, rather indistinct in the freeze-dried tissue. Comparison of the relevant chick and quail cells confirmed the reliability of the
nuclear marker for this experiment.
Two chick and two quail embryos each labelled with 10/*Ci H3-thymidine
in the same way as donor embryos, each given DOPA at 3 | days of incubation
Origin of pancreatic APUD (islet) cells
583
AK
FIGURES 1-5
Sections through the dorsal pancreas of normal (unoperated) 3J-day-old embryos.
Figs. 1, 2. Adjoining sections of a quail embryo.
Fig. 1. DOPA-provoked FIF in pancreatic APUD cells. x420.
Fig. 2. Large Feulgen-positive nucleoli in the pancreatic APUD cells, x 420.
Figs. 3, 4. The same section of a labelled quail embryo.
Fig. 3. DOPA-provoked FIF in pancreatic APUD cells. x420.
Fig. 4. Autoradiograph showing label in the pancreatic APUD cells x 420.
Fig. 5. DOPA-provoked FIF in pancreatic APUD cells of an unlabelled chick
embryo. x210.
37-2
584
A. ANDREW
and submitted to the FIF procedure followed by autoradiography, showed
adequate isotope labelling in the neural tube, spinal and sympathetic ganglia and
pancreatic APUD cells (Figs. 3, 4), provided observations on embryos with
quail grafts were made prior to Feulgen staining. Labelling of pancreatic cells
was particularly intense. Development of all these structures was normal in
the labelled embryos.
Under the same conditions, none of the relevant structures showed negative
chemography in the normal chick and quail embryos tested; nor did positive
chemography occur over sections of comparable unlabelled embryos. Examination of autoradiographs of operated embryos, four of which had received chick
grafts and two, quail grafts, confirmed that positive chemography was no problem. Nor were any signs of negative chemography present in the test slides of
operated embryos with labelled grafts, except for one specimen, which was
discarded.
The necessity for administration of DOPA for demonstration of the APUD
reaction of pancreatic cells at 3 | days was confirmed for chick embryos (see
Andrew, 1975) and shown for quail embryos. Five embryos of each species
treated with DOPA showed fluorescence of the cells, whereas in three chick and
two quail unoperated embryos, and one operated chick embryo, none of which
was given DOPA, fluorescence was lacking. In three chick and two quail unoperated embryos and two operated (chick) embryos all treated with DOPA,
but not subjected to formaldehyde vapour fixation, there was no autofluorescence
in any pancreatic cells. The presence of isotope did not affect the above results.
Pancreatic APUD cell development (Figs. 1, 3, 5) in five unoperated chick
and five unoperated quail embryos showed 3 | days of incubation to be a suitable
time for sacrifice in relation to the duration of radioactive label. APUD cells
are present in the chick dorsal pancreatic bud considerably earlier, at the time
of its evagination at or shortly before the 27-somite stage (Andrew, 1975); in
quails they are already present at the 31-somite stage. Even if surgical intervention had retarded development, pancreatic APUD cells should therefore
still have been demonstrable, though in smaller numbers.
Operated embryos
Transplantations were performed on 128 embryos, of which 73 % survived.
Three were discarded due to poor or abnormal development, one due to negative chemography, and a few were spoilt during technical procedures. Of the
rest, labelled chick grafts had been transplanted to 40, labelled quail grafts to
26, unlabelled quail grafts to 12 and unlabelled chick grafts to 6.
In most cases, the grafts healed well in the host embryos. Small fragments of
donor notochord were occasionally identified in sectioned operated embryos,
and in two cases only, a little donor mesoderm adjacent to the graft. More
often, remnants of host neural tube were present in theoperated region, generally
ventral to the grafted tube. The latter was usually well-formed, though junctions
585
Origin of pancreatic APUD (islet) cells
Table 1. The absence of graft-derived cells among pancreatic APUD cells
Pancreas
Grafts
Presence of
ganglia in
operated
region
Labelled
Chick
Quail
Fluorescent
UnTotal
APUD
labelled no. of
operated Well de- cells
Quail embryos veloped present
Spinal and
13
10
10
33
30
29
sympathetic
1
1
5
7
6
7
Spinal or
sympathetic
Total no. of
14
11
15
40
36
36
embryos
The figures represent numbers of successfully operated embryos.
Labelled
and/or
quail
nuclei in
APUD
cells
0
0
0
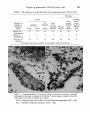
Figs. 6, 7. Autoradiograph of a 3f-day-old embryo which had received a labelled
chick graft at the levels of somites 16-23 at the 17-somite stage. In order to show the
silver grains, the tissue is slightly out of focus.
Fig. 6. Labelled grafted neural tube(AT)and labelled spinal ganglion (SG). x 620.
Fig. 7. Labelled sympathetic ganglion (SyG). x 620.
586
A. ANDREW
Figs. 8-11. Sections through the dorsal pancreas of 3J-dny-old operated embryos.
Figs. 8, 9. The same section through an embryo which had received a labelled chick
graft at the levels of somites 16-23 at the 17-somite stage.
Fig. 8. DOPA-provoked FIF in pancreatic APUD cells, x 420.
Fig. 9. Autoradiograph showing no label in the pancreatic APUD cells. Only a few
background silver grains are present, x 420.
Figs. 10, 11. The same section of an embryo which had received an unlabelled
quail graft at the levels of somites 17-22 at the 17-somite stage, x 420.
Fig. 10. DOPA-provoked FIF in pancieatic APUD cells.
Fig. 11. Lack of large Feulgen-positive nucleoli in the pancreatic APUD cells.
x420.
with the host tube were not always neat. In all but a few cases, development of
the pancreas was normal (Tables 1,3; Figs. 8-11). The general appearance and
stage of development of successfully operated embryos (for criteria see below)
at 3J days of incubation were almost always normal.
In eight embryos with well-formed chick grafts and six with well-developed
quail grafts, the intensity of radioactive labelling in autoradiographs of the
donor neural tube was considered inadequate. Those which bore quail grafts
are considered in Table 1, together with the embryos having unlabelled quail
grafts. The embryos with chick grafts could not be used.
Origin of pancreatic APUD {islet) cells
587
Table 2. Successful transplantation or deletion of neural tube and crest
at various levels
Somite
level
No. of
times
transplanted
No. of
times
deleted
1
1
2
4
5
6
6
6
—
—
1
1
1
2
2
2
4
5
7
7
1
2
3
4
5
6
7
8
9
10
II
12
13
14
15
8
10
10
11
13
14
13
Somite
level
No. of
times
transplanted
No. of
times
deleted
13
13
15
14
14
13
11
9
5
4
5
3
3
3
3
4
1
1
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
6
6
4
2
1
1
1
1
1
1
1
1
1
1
1
Table 3. Pancreatic APUD cells in embryos lacking neural crest
at various levels
Pancreas
Ganglia present in operated region
Total no. of
embryos
Spinal
8
5
Sympathetic
Well
developed
Fluorescent
APUD cells
present
7
7
7
The figuresrepresent numbers of embryos.
Operations were regarded as successful if there was good evidence of migration of graft cells from the transplant. Of the 40 such embryos, at least some
spinal ganglia and/or sympathetic ganglia were present in the operated region
(Table 1); they showed the markets of the grafted tissue, i.e. radioactive label
(Figs. 6, 7) and/or quail nuclei. In many of these, migration was normal or almost
normal. In most cases, neurilemmal (Schwann) cells of graft origin were identified in the spinal nerve roots. In a further eight operated embryos there was little
or no evidence of cell migration from the graft. These are not included among
the successful operations and are not shown in Table 2. In 36 of the 40 successful
cases, FIF was demonstrated in normal numbers of cells in the dorsal pancreatic
bud (Figs. 8, 10). In none of these did any pancreatic APUD cells show radioactive label (Fig. 9), nor did any have nuclei with quail features (Fig. 11; Table 1).
The number of times neural tube and neural crest of each somite level were
588
A.ANDREW
included in a transplant is recorded in Table 2. Levels from somite 5 to somite 24
are well-represented.
The grafted pieces of neural tube escaped from the site of transplantation in
eight embryos. These may be regarded as embryos from which the neural crest
was deleted at the operated levels. The somite levels affected by such deletions
are listed in Table 2. Spinal and sympathetic ganglia were very seldom present
in the operated regions. FIF revealed normal development of A PUD cells in
all except one of these embryos (Table 3).
DISCUSSION
In this experiment, it was vital that any migration of host neural crest cells
from operated levels to the pancreas should be forestalled. Previous experiments
showed that the grounds for the selection of levels for transplantation in embryos
at the various stages (see Materials and Methods) was sound: chorio-allantoic
grafts of blastoderm designed - on the same basis - to exclude neural crest,
were shown to be free of crest derivatives (Andrew, 1963).
In operated embryos in which a radioactively labelled graft was recognizable
at the time of sacrifice, label was nevertheless absent from the pancreatic APUD
cells. This was not attributable to dilution of label in these cells as they are
well-labelled in normal embryos; it was not due to negative chemography, nor
to interference with latent image formation by the APUD-FIF procedure. Since
quail cell nuclei are markedly different from chick nuclei in pancreatic APUD
cells of normal embryos, the absence of quail features was a reliable indication
that no graft cells had entered the pancreas in those operated embryos bearing
quail grafts. The development of the operated embryos, and of the pancreas
and the pancreatic APUD cells in particular, were unaffected by the experimental procedure in all but a few cases, and the cells appeared to be present in
normal numbers.
In operated embryos, normal migration of neural crest cells did not always
occur. Only embryos were scored which showed evidence of migration of marked
cells to form known derivatives of the neural crest and perhaps neural tube, i.e.
spinal and sympathetic ganglia, and neurilemmal cells. As judged by these
criteria, the migration of neural crest cells from quail grafts was as extensive
as from chick grafts. It is apparent from many similar experiments of Le
Douarin and her co-workers, that isotopic quail to chick grafts are prolific in
the formation of migratory neural crest cells (Le Douarin & Teillet, 1970, 1973,
1974; Le Lievre & Le Douarin, 1975).
In the operated embryos, the transplants represented the levels between
somites 5 and 24 well enough to justify the conclusion that the pancreatic APUD
cells present on sacrifice were not derived from these levels of the neural crest.
The presence of apparently normal numbers of pancreatic APUD cells in
the embryos lacking segments of trunk neural crest is in line with the above
Origin of pancreatic APUD {islet) cells
589
conclusion. Alone, such evidence would be inconclusive, because neural crest
from adjoining areas is known to migrate into levels from which the crests have
been deleted (see Andrew, 1971). The rare occurrence of spinal and sympathetic
ganglia at levels deprived of neural crest, however, suggests only limited
penetration from adjoining crests.
It seems unlikely that levels of the crest caudal to somite 24 give rise to pancreatic APUD cells. At the 24-somite stage (about 2 days of incubation), when
the evagination of the dorsal pancreatic bud is imminent, APUD cells are
already present in the region which will evaginate (Andrew, 1975). No migration
of crest cells occurs at post-somitic levels (see Andrew, 1963). Therefore, at least
those APUD cells present at the 24-somite stage could not have arisen from
the neural crest of levels caudal to somite 24. From previous morphological
observations, it seems that APUD cells appearing in the primitive gut groove
from the 16-somite stage are the progenitors of pancreatic APUD cells, since
they appear to aggregate in the presumptive dorsal pancreatic region (Andrew,
1975). Tt is therefore more likely that the APUD cells in the dorsal bud when it
evaginates will proliferate to form the full complement of pancreatic APUD
progenitor cells, than that others arrive there later from caudal levels of the
neural crest.
Rhombencephalic levels of the neural crest are still to be tested. Pearse
(1973) on one occasion mentioned these levels of the crest as the source of
pancreatic APUD cells, but gave no reason for doing so. Certainly neural crest
cells from hindbrain levels do reach the gastro-intestinal tract caudal to the
pancreas (where they become enteric ganglion cells (Andrew, 1969; Le Douarin
& Teillet, 1973)).
The APUD cells present in the chick pancreas at 3 | days of incubation
probably include A, B and D cells. In different species, different islet cell types
are APUD (see Andrew, 1975): in chick embryos of between 9 and 18 days of
incubation all three types are APUD (Andrew, unpublished observations).
A and B cells have been identified in the dorsal pancreatic bud shortly before
3 | days by electron microscopy (Dieterlen-Lievre, 1965; Przbylski, 1967) and
D cells at the 31 -somite stage (2% days of incubation) by light microscopy
(Andrew, unpublished observations). It is therefore likely that A, B and D cells
may be included among the APUD cells of the present study, but direct
evidence is still required.
Completely in line with the results of this investigation are the findings of
Phelps (1975). He has shown experimentally that B cells in the rat pancreas
are not derived from the neural crest. He cultured endoderm and mesoderm
from which ectoderm had been removed before formation of the neural crests.
Insulin was detected in the differentiated pancreas; B cells were identified
ultrastructurally.
Pearse & Polak (1971) proposed that APUD cells in the mouse pancreas are
derived from the neural crest, because they saw cells showing DOPA-provoked
590
A. ANDREW
FIF first in a position attributed to the central stream of the neural crest cells,
in the mesenchyme between the neural tube and the pharynx, then in the
pharynx, and later in the dorsal pancreatic bud. The earliest APUD cells illustrated seem to the author to be in too large a mass, and to lie too far laterally,
to be neural crest cells. Also, although observations made on cells with the
same features at successive stages of development are highly suggestive evidence
that the cells follow the route of migration mapped out in this way, such
evidence alone is not conclusive.
Later, Pearse, Polak & Heath (1973) apparently abandoned the idea that crest
cells invade the dorsal pancreatic bud directly, and traced pancreatic APUD
cells in the mouse from the 'primitive endocrine cell' in the gut wall, claimed
by Pearse & Polak (1971) to be of neural crest origin. Granules (described as
pleomorphic), characteristic of these non-argentafnn APUD cells, were found
later on in development in pancreatic cells, together with round granules regarded as characteristic of islet cells. The latter were also shown to be APUD
cells. These findings tie in with the observations of the present author on the
occurrence of APUD cells successively in the gut wall of chick embryos, the
presumptive dorsal pancreas and the bud itself (see above) and are in accord
with the suggestion that the differentiation of the islet cells in chick embryos
may begin before evagination of the pancreatic bud (Przbylski, 1967). Pearse
et ah (1973) conclude that the islet cells (all three types according to a later
statement of Pearse, 1973), are derived from the neural crest. The present study
does not support this contention for the trunk neural crest at least.
The pancreatic APUD cells referred to in the present investigation are localized in the dorsal pancreatic bud. None are present in the ventral buds before
these fuse with the dorsal bud (Andrew, 1975). (Fusion occurs at a stage later
than the time of sacrifice in this experiment.) Many workers maintain that
islets arise only from the dorsal bud in vertebrates (see Gianelli, 1908; WolfHeidegger, 1936-cited by Frye, 1962; Frye, 1962), though some attribute
islet formation to ventral as well as dorsal buds (Hard, 1944, in the rat). From
the distribution in the adult, Bencosme & Liepa (1955) contend that derivatives
of both buds in the dog and cat form islets. However, islet cells arising from one
bud could easily pass into and proliferate in the other after fusion. Extirpation
and transplantation of buds, or of presumptive regions from which the buds
arise, have produced evidence that the islets arise from the dorsal bud only, in
amphibians (Frye, 1962) but from both dorsal and ventral buds in chicks
(Sandstrom, 1934; Dieterlen-Lievre, 1970).
The lack of APUD cells in chick ventral pancreatic buds seems to be at
variance with participation of ventral buds in islet formation. If, however, the
ventral buds do indeed contribute to the definitive islets, then it is clear that the
present study has not dealt with their origin. It would be strange, though, if
their source were different from those of the dorsal bud.
The conclusion reached from this experiment is that the pancreatic APUD
Origin of pancreatic APUD (islet) cells
591
(islet) cells present in the chick embryo at 3 days 18 h of incubation are not
derived from the neural crest of the levels of somites 5 to 24, and probably
not from more caudal levels. It is likely that these pancreatic APUD cells include A, B and D cells, The pathological significance of the embryological origin
of APUD cells is discussed elsewhere (Andrew, 19766).
The author would like to thank the Atomic Energy Board of South Africa for a grant
for isotope and nuclear emulsion and the South African Medical Research Council and the
Council for Scientific and Industrial Research for grants for equipment. For advice on operative procedures, the author is very grateful to Professor N. Le Douarin of the Laboratoire
d'Embryologie, Universite de Nantes, and Dr K. Hara of the Hubrecht Laboratory, Utrecht.
Thanks are due to Mr D. W. Roberts of the University of the Witwatersrand's Animal Unit
at Frankenwald for obtaining and maintaining a colony of quails to supply fertile eggs, to
Mr J. Bunning for identifying the quails, Mr R. G. Klomfass for constructing the turntable
system, Mr R. J. Herman, Mrs N. G. de Maar, Mr E. Favini. Mrs B. Levitan and Mr
W. Tadiello for competent technical assistance, and, finally, to Professor P. V. Tobias, Head
of the Department of Anatomy, for his continued interest and encouragement.
REFERENCES
J. W. (1971). Enterosecretory proteins. Nature, Lond. 289, 321-325.
A. (1963). A study of the developmental relationship between enterochromaffin
cells and the neural crest. /. Embryol. exp. Morph. 11, 307-324.
ANDREW, A. (1969). The origin of intramural ganglia. III. The vagal source of enteric ganglion
cells. /. Anat. 107, 327-336.
ANDREW, A. (1971). The origin of intramural ganglia. IV. The origin of enteric ganglia:
a critical review and discussion of the present state of the problem. /. Anat. 108, 169-184.
ANDREW, A. (1975). APUD cells in the endocrine pancreas and the intestine of chick embryos. Gen. comp. Endocr. 26, 485-495.
ANDREW, A. (1976a). Evidence that pancreatic APUD cells in chick embryos are not derivatives of the neural crest. IRCS Medical Science 4, 27.
ANDREW, A. (19766). APUD cells, Apudomas and the neural crest: a critical review.
S. Afr. med. J. 50 (in the Press).
BELSARE, D. K. (1974). Studies on the development of endocrine glands in fishes. III.
Morphogenesis of the endocrine pancreas in Clarias batrachus L. Z. mikrosk.-anat.
Forsch. 88, 981-986.
BENCOSME, S. A. (1955). The histogenesis and cytology of the pancreatic islets in the rabbit.
Am. J. Anat. 96, 103-152.
BENCOSME, S. & LIEPA, E. (1955). Regional differences of the pancreatic islet. Endocrinology
57, 588-593.
CEGRELL, L. (1967). Dopamine in the pancreas of albino and pigmented newborn guinea-pigs.
Life Sci. 6, 2491-2495.
CEGRELL, L. (1968). Adrenergic nerves and monoamine-containing cells in the mammalian
endocrine pancreas. Acta physiol. scand. Suppl. 314, 17-23, 14-16, 35-39, 40-4.1.
CEGRELL, L., FALCK:, B. & ROSENGREN, A.-M. (1967). Dopamine and 5-hydroxytryptamine in
the guinea-pig pancreas. Life Sci. 6, 2483-2489.
DIETERLEN-LIEVRE, F. (1965). Etude morphologique et experimentale de la differenciation du
pancreas chez l'embryon de poulet. Bull. biol. Fr. Belg. 99, 31-16.
DIETERLEN-LIEVRE, F. (1970). Tissus exocrine et endocrine du pancreas chez l'embryon de
poulet: origine et interactions tissulaires dans la differenciation. Devi Biol. 22, 138-156.
EPPLE, A. & LEWIS, T. L. (1973). Comparative histophysiology of pancreatic islets. Am. Zool.
13, 567-590.
FALCK, B. & HELLMAN, B. (1963). Evidence for the presence of biogenic amines in pancreatic
islets. Experientia 19, 139-140.
ADELSON,
ANDREW,
592
A. ANDREW
F. (1943). Uber das Inselorgan des Menschen. Ergebn. allg. Path. Anat. 36, 3.
(Cited by Weichert, 1970.)
FRYE, B. E. (1958). Development of the pancreas in Amblystoma opacum. Am. J. Anat. 102,
117-140.
FRYE, B. E. (1962). Extirpation and transplantation of the pancreatic rudiments of the
salamanders, Amblystoma punctatum and Eurycea bislineata. Anat. Rec. 144, 97-103.
GIANELLI, K. (1908). Contributo allo studio dello sviluppa del pancreas negli Uccelli. Monit.
zool. ital. 19, 1986-1999.
HARD, W. L. (1944). The origin and differentiation of the alpha and beta cells in the pancreatic islets of the rat. Am. J. Anat. 75, 369-402.
HELLMAN, B. (1966). The development of the mammalian endocrine pancreas. Biol. Neonat.
9, 263-278.
LE DOUARIN, N. (1961). Radiodestructions partielles chez l'embryon de poulet aux stades
jeunes et localisation des ebauches digestives. /. Embryol. exp. Morph. 9, 1-8.
LE DOUARIN, N. (1971). Caracteristiques ultrastructurales du noyau interphasique chez le
caille et chez le poulet et utilisation de cellules de caille comme 'marqueurs biologiques'
en embryologie experimentale. Annls Embryol. Morph. 4, 125-135.
LE DOUARIN, N. & TEILLET, M.-A. (1970). Sur quelques aspects de la migration des cellules
neurales chez l'embryon de poulet etudiee par la methode des greffes heterospecifiques de
tube nerveux. C. r. Seam. Soc. Biol. 164, 390-397.
LE DOUARIN, N. & TEILLET, M.-A. (1973). The migration of neural crest cells to the wall of
the digestive tract in avian embryo. /. Embryol. exp. Morph. 30, 31—48.
LE DOUARIN, N. & TEILLET, M.-A. (1974). Experimental analysis of the migration and differentiation of neuroblasts of the autonomic nervous system and of neurectodermal mesenchymal derivatives, using a biological cell marking technique. Devi Biol. 41, 162-184.
LEGG, P. G. (1968). Fluorescence studies on structures and endocrine cells in the pancreas of
the cat. Z. Zellforsch. mikrosk. Anat. 116, 205-227.
LE LIEVRE, C. S. & LE DOUARIN, N. M. (1975). Mesenchymal derivatives of the neural crest:
analysis of chimaeric quail and chick embryos. /. Embryol. exp. Morph. 34, 125-154.
LIEGNER, B. (1932). Studien zur Entwicklung des Pankreas, besonders der Langerhanschen
Inseln. Z. mikrosk.-anat. Forsch. 90, 494-529.
LIKE, A. A. & ORCI, L. (1972). Embryogenesis of the human pancreatic islets: a light and
electron microscope study. Diabetes 21, Suppl. 2, pp. 511-534.
MAWHINNEY, B. S., AUSTIN, B. T. & RILEY, E. F. (1972). The uptake of topically applied
tritiated thymidine by chick embryos. Proc. Soc. exp. Biol. Med. 140, 208-211.
PEARSE, A. G. E. (1960). Histochemistry. Theoretical and Applied, 2nd ed., p. 822. London:
Churchill.
PEARSE, A. G. E. (1966). Common cytochemical properties of cells producing polypeptide
hormones with particular reference to calcitonin and the thyroid C cells. Vet. Rec. 79,
587-590.
PEARSE, A. G. E. (1969). The cytochemistry and ultrastructure of polypeptide-hormone producing cells of the APUD series and the embryologie, physiologic and pathologic implications of the concept. /. Histochem. Cytochem. 17, 303-313.
PEARSE, A. G. E. (1973). Cell migration and the alimentary system: endocrine contributions
of the neural crest to the gut and its derivatives. Digestion 8, 372-385.
PEARSE, A. G. E. & POLAK, J. M. (1971). Neural crest origin of the endocrine polypeptide
(APUD) cells of the gastrointestinal tract and pancreas. Gut 12, 783-788.
PEARSE, A. G. E., POLAK, J. M. & BUSSOLATI, G. (1972). The neural crest origin of gastrointestinal and pancreatic endocrine polypeptide cells and their distinction by sequential
immunofluorescence. Folia Histochem. Cytochem. 10, 115-120.
PEARSE, A. G. E., POLAK, J. M. & HEATH, C. M. (1973). Development, differentiation and
derivation of the endocrine polypeptide cells of the mouse pancreas. Immunofluorescence,
cytochemical and ultrastructural studies. Diabetologia 9, 120-129.
PEARSE, A. G. E. & TAKOR TAKOR, T. (1975). Neurocrine embryology and the APUD concept. Endocrinology 1975: Proc. 6th Int. Symp., London. (Abstract.)
FEYRTER,
Origin of pancreatic APUD {islet) cells
593
A. G. E. & WELSCH, U. (1968). Ultrastructural characteristics of thyroid C cells in
the summer, autumn and winter states of the hedgehog (Erinaceus europaeus L.), with
reference to other mammalian species. Z. Zellforsch. mikrosk. Anat. 92, 596-609.
PHELPS, P. (1975). Evidence that the endocrine pancreatic cells are derived from the neural
crest. Anat. Rec. 181, 449.
PICTET, R. L., CLARK, W. R., WILLIAMS, R. H. & RUTTER, W. J. (1972). An ultrastructural
analysis of the developing embryonic pancreas. Devi Biol. 29, 436-467.
PrcTET, R. & RUTTER, W. J. (1972). Development of the embryonic pancreas. In Handbook
of Physiology, section 7, vol. 1 (ed. D. F. Steiner & N. Freinkel), pp. 25-66. Washington
D.C., Am. Physiol. Soc. (Cited by Epple and Lewis, 1973.)
POLAK, J. M., ROST, F. W. D. & PEARSE, A. G. E. (1971). Fluorogenic amine tracing of
neural crest derivatives forming the adrenal medulla. Gen. comp. Endocrinol. 16, 132-136.
PRZBYLSKI, R. J. (1967). Cytodifferentiation of the chick pancreas. I. Ultrastructure of the
islet cells and the initiation of granule formation. Gen. Comp. Endocr. 8, 115-128.
ROBB, P. (196.1). The development of the islets of Langerhans in the human fetus. Q. Jl exp.
Physiol. 46, 335-343.
ROGERS, A. W. (1967). Techniques of Autoradiography, pp. 201-202, 269-271. Amsterdam:
Elsevier.
SANDSTROM, R. H. (1934). The differentiation of hepatic and pancreatic tissues of the chick
in chorio-allantoic grafts. Physiol. Zool. 7, 226-246.
SCHWEISTHAL, M. R. & FROST, C. C. (1973). Differentiation of alpha cells in the fetal rat
pancreas grown in organ culture. Am. J. Anat. 136, 527-532.
SILVER, P. H. S. (1960). Special problems of experimenting in ovo on the early chick embryo,
and a solution. /. Embryol. exp. Morph. 8, 369-375.
TRANDABURU, T. (1972). Comparative observations on adrenergic innervation and monoamine content in endocrine pancreas of some amphibians, reptiles and birds. Endokrinologie 59, 260-264.
VILLAMIL, M. F. (1942). Citogenesis del pancreas exo y endocrino en embriones de polio.
Rev. Soc. arg. Biol. 18, 416-424.
WESSELS, N. K. (1968). Problems in the analysis of determination, mitosis and differentiation.
In Epithelio-Mesenchymal Interactions, 18th Hahnemann Symp. (ed. R. Fleischmajer &
R. E. Billingham), pp. 132-151. Baltimore: Williams & Wilkins.
WESTON, J. A. (1963). A radioautographic analysis of the migration and localisation of trunk
neural crest cells in the chick. Devi Biol. 6, 279-310.
PEARSE,
(Received 17 December 1975, revised 13 January 1976)