Survey
* Your assessment is very important for improving the work of artificial intelligence, which forms the content of this project
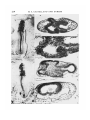
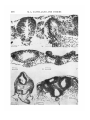
/. Embryol. exp. Morph. Vol. 29, 1, pp. 209-219, 1973 209 Printed in Great Britain The effects of isonicotinic acid hydrazide on the early chick embryo By M. A. CASTELLANO, J. L. T 6RTORA, N. I. GERMINO, F. RAMA AND C. OHANIAN 1 From the Facultad de Veterinaria, Montevideo SUMMARY 1. Unincubated White Leghorn fertile eggs were injected with isonicotinic acid hydrazide (INH) through a hole in the shell. A control group was injected with normal saline, a second group with INH plus vitamin B6, and a third group was left unopened. 2. INH, in the doses we used, proved to be lethal for more than 50% of the early chick embryos, and also produced important developmental alterations. 3. Alterations produced by INH were primarily observed at the level of the neural epithelium, particularly at its cephalic portion. The most important ones were a degenerative and necrotic process of the neural epithelium, and a distortion of the normal anatomical relations of the cephalic structures. 4. The embryonic mortality and developmental alterations induced by INH were prevented to a considerable extent by the concurrent injection of pyridoxine hydrochloride and INH. 5. The preventive action of vitamin B6 suggests that toxicity of INH in the chicken embryos is due to the antivitamin-B6 action of INH. Through this mechanism INH would block, specially, the amino acid metabolism. The developing nervous system was the embryonic area most sensitive to such metabolic alterations. INTRODUCTION It has been demonstrated that hydrazines induce a block in the metabolism of amino acids (Lewis & Izume, 1926), and that this blockade can be raised by the administration of vitamin B6 (McCormick & Snell, 1961; Simonsen & Roberts, 1967). Studies in vitro (Killam & Bain, 1957) indicated that a number of hydrazines (thiosemicarbazide, furoyl hydrazine and isonicotinic acid hydrazide) inhibit the enzymic activities of the amino acid metabolism, catalysed by pyridoxal. The blockade of pyridoxal enzymes by isonicotinic acid hydrazide (INH) has been reported by Yoneda, Kato & Okajuna (1952) and Yoneda & Asano (1953). INH combines with pyridoxal phosphate to form a hydrazone and thereby causes a competitive inhibition of aminopherases, decarboxilases, diamine oxidase and other pyridoxal enzymes (Braunstein, 1960). In the chick embryo, it has been demonstrated that vitamin B6 is essential for 1 Authors' address: Facultad de Veterinaria, Instituto de Anatomia Normal, Casilla de Correo 2351 (distrito 6) Montevideo, Uruguay. 14 EMB 29 210 M. A. CASTELLANO AND OTHERS the development of the early stages (Cravens & Snell, 1949; Cravens, 1952; Conti & Milio, 1965, 1966a, b). The pyridoxine analogs (deoxypyridoxine and methoxypyridoxine) have an inhibitory effect in development, producing 100% mortality during the first 4 days of incubation (Cravens, 1952). The purpose of this investigation is to study the action of INH in the early chick embryo during the first 4 days of development and to test the preventive action of pyridoxine hydrochloride. MATERIAL AND METHODS Fertilized White Leghorn eggs, purchased from a local hatchery, were used throughout all the experiments. A forced-draft incubator maintained at 99 °F (37-2 °C) was used. All experimental specimens were treated prior to incubation, using a slight modification of the method described by Landauer (1945): eggs were opened after being wiped with alcohol, and the shell was cut with a sterile saw-blade; a small rectangle (0-5 x 1 cm2) of the shell was removed. Experimental specimens were treated with 1 mg INH dissolved in 0-05 ml of 0-85 % NaCl, injected into the yolk sac at 5 mm of the end of blastodisc just beneath the vitelline membrane, using a sterile needle (0-5 mm external diameter). Another group was injected using the same procedure with 1 mg INH and 1 mg pyridoxine hydrochloride dissolved together in 0-05 ml of normal saline. Eggs meant to be used as saline-injected controls were opened accordingly and injected with 0-05 ml of saline. The eggs were sealed with Parafilm and melted paraffin. Unopened controls were handled but were not treated further. After injection the eggs were incubated for 24, 48 or 72 h, and at the end of these periods the embryos were dissected under a low-power binocular microscope, and we observed presence or absence of the embryonic heart beat. Only those embryos exhibiting both heart beat and circulation in the vessels of the area vasculosa were registered as living embryos and processed. After they had been macroscopically examined and photographed under the low-power binocular microscope, they were fixed in Bouin's fluid for histological study. Material was double embedded in celloidin and paraffin, serially sectioned (10 jtim) in a transverse plane, and stained with haematoxylin and eosin. RESULTS Quantitative data According to data summarized in Table 1, there was no significant difference (P > 0-05) between the percentages of survivors in the saline-injected control group (74%) and the unopened controls (84%). The mortality in the 1NHtreated group at 2 days and 3 days of incubation (57 % and 63 % respectively) was significantly higher (P < 0-005) than in the saline-injected group (26 %). 111 Effects of isonicotinic acid hydrazide Table 1. Results of yolk-sac injection ofJNH, INH+vitamin BG, and saline in fertilized White Leghorn eggs, prior to the incubation Living embryos Age of the (h) No. of embryos recovered 48 72 72 72 72 67 70 72 93 76 embryos Treatment INH .1 mg INH 1 mg INH + vit. B6 Saline Unopened Mortality i— 0/ /o 57 63 33 26 .16 *—, No. 38 44 24 24 12 Total ,—*—, % No. 43 37 67 74 84 29 26 48 69 64 No. malformed ,—*—, % No. 100 77 25 20 5 29 20 12 14 3 No. normal ,—K —\ % 1vfo. 0 0 23 6 75 36 80 55 95 61 But the mortality after 2 days of incubation was 57 % and there was no increase in the mortality after 3 days of incubation (63%; P > 0-05). In the group injected with 1 mg INH together with 1 mg pyridoxine hydrochloride the embryonic lethality (33%) was lower (P < 0-005) than in the INH-treated group (63 %). There was no significant difference (P > 0-05) in the proportion of dead embryos in INH + vitamin B6-injected group (33 %) and the saline-injected control group (26%). However, the mortality in INH + vitamin B6-treated group (33%) was significantly higher (P < 0-025) than in the unopened group (16%). Statistically there was no difference (P > 0-05) in the proportion of malformed embryos between the saline-injected group (20%) and the unopened group (5%). Alterations by administering INH were produced in the chick embryos; the percentage of abnormal embryos was 50 % after 24 h of incubation and over 75 % after 3 days of development. The histological study of the embryos recovered after 24 h of incubation showed that 50 % were abnormal. The study of the gross morphology of 2-day embryos injected with INH showed that 81 % were abnormal, but the microscopical examination demonstrated that all the embryos had histological alterations. When the gross morphology of 3-day embryos injected with INH was studied, 54% were abnormal, but the microscopical examination showed a 77 % of malformed embryos. According to the statistical analysis by the chisquare method, the proportion of malformed embryos did not increase from the second (100%) to the third day of development (77%) (P > 0-05). Both percentages (77 and 100%) were significantly higher than the proportion of malformed embryos in the saline-injected control group (20%) (P < 0-005). A significantly lower proportion (P < 0-005) of the embryos from those injected simultaneously with INH and vitamin B6 were considered abnormal (25 %) compared to the proportion of those abnormal from the group injected with INH (77%). There was no difference (P > 0-05) between the 1NH + 14-2 212 M. A. CASTELLANO AND OTHERS Effects of isonicotinic acid hydrazide 213 vitamin B6-injected group (25%) and the normal saline-injected group (20%) in the proportion of malformed embryos. However, the INH + vitamin Boinjected group demonstrated a higher percentage (25 %) of malformed embryos compared with the unopened group (5 %) (P < 0-025). Finally, some of the INH-treated embryos, that in gross anatomy had been considered normal, in the histological study demonstrated, in all cases, varying degrees of developmental alterations, mainly a necrotic and degenerative process in the neural epithelium. Morphological data The macroscopical and microscopical study of 24 h chick embryos treated with 1 mg INH demonstrated in the abnormal cases (50 %) the absence or the rudimentary organization of the primitive streak (Fig. 1 A), or in some cases the irregular invagination of the mesoblast at the primitive groove. The macroscopical examination of 2- and 3-day chicken embryos showed that almost every INH-injected embryo had a growth and developmental retardation; they were approximately two-thirds the size of the corresponding controls. The most severe effects were found in the cranial portion of the neural tube (Fig. 1B-E, Fig. 2G-K): some of them showed the absence of an encephalic nervous system (Fig. 1B-E) and sometimes partial absence of closure of the neural folds (Fig. 1F). The macroscopical analysis also demonstrated the scanty amount of circulating blood in INH-treated embryos; whereas the vascular system did not show any remarkable anomalies. The histological study of 2- and 3-day chick embryos showed that the most substantial morphological changes in the abnormal cases of the INH-injected FIGURE 1 (A) Transverse section of a 24 h chick embryo at the primitive-streak stage treated with INH. Note a rudimentary primitive streak formed in a blastoderm which belongs to experimental series, x 250. (B) 48 h chick embryo treated with J mg INH. The forebrain is suppressed and represented only by a mass of neural tissue with a tiny neural canal. Note the failure to close of the neural tube, h, Embryonic heart, x 18. (C) The same embryo as (B), section made at the level of the arrows in B. The forebrain is suppressed and represented by a tiny neural tube (n) located under the primitive pharynx (ph) and surrounded by the embryonic heart (h). x 130. (D) 48 h chick embryo treated with INH, showing disorganized brain and the irregular outline of the neural groove at the caudal level, h, Heart, x 18. (E) Transverse section of the embryo of D, the arrows point out the level of this section. Note the disorganized brain, the defect of the floor (arrow) of neural tube and numerous degenerate cells in the neural epithelium and surrounding tissues, x 85. (F) Transverse section through the caudal portion of the same embryo (D), showing a disorganized neural epithelium with picnotic cells and without the characteristic histological pattern of this epithelium, x 200. 214 M. A. CASTELLANO AND OTHERS Effects of isonicotinic acid hydrazide 215 group were anomalies of the developing nervous system, while the other embryonic tissues only occasionally showed alterations. The most frequent anomalies observed in the nervous system of the INH-treated embryos were: (a) Foci of degenerate cells and necrosis affecting a variable number of cells (Fig. 1 E, F; Fig. 2H, I, K, L; Fig. 3 M-P). This degenerative process consists chiefly of picnosis and cariorrexis (Fig. 2, I; Fig. 3, M-P); the degenerate cells are frequently seen desquamated and occasionally fill the lumen of the neural tube (Fig. 2H, K, L; Fig. 3N, P). In some embryos the floor of the neural tube showed an intense degeneration which occasionally produced the rupture of this sector (Fig. IE, arrow). Only from time to time does the notochord participate in this degenerative process (Fig. I F ; Fig. 3O, P). In all these cases the neural epithelium lost its characteristic histological pattern (Fig. I F ; Fig. 3 M-P) and the cells appeared with no special orientation. Over 50 % of the INH-treated embryos showed degeneration of the neural epithelium, while only 20% of INH + vitamin B6-injected embryos showed the same process. (b) Failure of the neural tube closure, with persistence of the neural groove (Fig.lD,F;Fig.2J;Fig.3P). (c) Multiple accessory lumen or duplication of the neural tube (Fig. 3R). (d) Asymmetric folding of the neural groove (Fig. 2H). (e) Complete anatomical disorganization in the cephalic structures as shown in Fig. 1C and H. In this embryo (Fig. 1B, C) the rudimentary cranial portion of nervous system developed between the primitive pharynx and the cardiac region. DISCUSSION In order to study the action of INH in early chick embryos, eggs were injected prior to incubation. Some authors, such as Carter (1964) and Gebhardt (1968), found that the insertion of a needle in the yolk sac, prior to incubation, causes a high mortality even in the controls. Our results indicate that the salineinjected controls developed until the third day of incubation with no significant FIGURE 2 (G) 48 h chick embryo treated with INH, showing defects in forebrain (/). x 15. (H, I) Transverse section through the forebrain of the same embryo as (G); note the irregular forebrain formation, and the degenerative and necrotic process in the higher magnification, x 130 and x 340. (J) 48 h chick embryo treated with INH; note the neural tube open at the posterior level (arrow) and the disorganized forebrain. x 18. (K) Transverse section through the forebrain of the embryo of (J), showing the intense degenerative and necrotic process of the neural epithelium and the lumen filled with desquamated cells, x 140. (L) Section through the embryo of (J) at the level of primitive pharynx; note the floor of the neural tube disorganized and the cells desquamated in the lumen, x 140. 216 M. A. CASTELLANO AND OTHERS R Effects of isonicotinic acid hydrazide 217 difference from the unopened controls. The different results we obtained could be due either to the brief incubation period we used or to the differences in the genetic background of the White Leghorn eggs used. On the other hand, McLaughlin et ah (1963) injected thousands of eggs prior to incubation without apparent loss in the controls. The present study indicates that 1NH causes a high mortality in the early chick embryos (over 50% at 2 or 3 days of incubation). The preventive effect of pyridoxine hydrochloride suggests that this lethal action is due, at least partially, to the antivitamin B6 effect of INH. These results agree with those developed by Cravens (1952) and Conti & Milio (1965, 1966#, b) using vitamin B6 analogs. Moreover, the present work indicates that INH can modify the processes of development. The modifications produced by INH have some characteristic features; this effect may be shown by early death or by arrested or delayed development, and developmental abnormalities chiefly at the neural epithelium, including the absence of cephalic structures of the nervous system. We wish to emphasize the presence of a necrotic and degenerative process at the neural epithelium, as well as a disorder of the normal pattern of cell distribution in this embryonic epithelium. In the chick embryo, Duraiswami (1961) found that INH administered on the 3rd, 4th and 5th days of development causes limb deformities, beak defects and ectopia viscerum. The disagreement between these results and those presented in this paper could be attributed to the stage during development of the embryo when the drug is administered, giving differential susceptibility to INH (Murphy, 1967). The simultaneous injection of pyridoxine hydrochloride together with the INH prevented these developmental abnormalities, which suggests that they are produced by a vitamin B6 deficiency. The toxicity of hydrazines has been attributed to an acute vitamin B6 deficiency, and pyridoxal reverses this toxic effect (Cornish, 1969). A deficiency of pyridoxine has been considered to be in the adult, the cause of the 1NH- FIGURE 3 (M) INH-treated 48 h chick embryo. The transverse section shows the presence of picnotic and degenerate cells, and disorganization of the neural epithelium, x 400. (N) Transverse section through the forebrain of a 48 h INH-treated chick embryo; note desquamated cells filling the lumen, x 340. (O) INH-treated 48 h chick embryo; note the degenerative process affecting the neural epithelium and the notochord. x 220. (P) Failure of the caudal neural tube to close in a 48 h chick embryo INH-treated. x200. (Q) INH-treated 72 h chick embryo, showing the irregular rombencephalon formation and the desquamation to the lumen, x 85. (R) A 72 h chick embryo treated with INH + vitamin B6; note the multiple neural tube. x225. 218 M. A. CASTELLANO AND OTHERS induced neuropathy. However, some of the neurological disorders produced by INH are not prevented by pyridoxine (Lampert & Schochet, 1968; Yonezawa, Mori & Nakatani, 1969; Carlton & Kreutzberg, 1966). It has been demonstrated both in vitro and in vivo, that INH forms a hydrazone with pyridoxal phosphate and thereby causes a competitive inhibition of several pyridoxal enzymes (Braunstein, 1960). An additional explanation for the inhibitory effect of INH on enzyme systems catalysed by vitamin B6 emerges from the inhibition of pyridoxal kinase enzyme by INH, so that, while pyridoxal accumulates, there is insufficient pyridoxal phosphate to saturate the apoenzymes (Levy, 1969). The interference by INH with the vitamin B6 enzymes produces primarily a block in the amino acid metabolism. Our results indicate that the normal development of the nervous system at its early stages probably depends on a normal amino acid metabolism. Regional differences in sensitivity to several metabolic inhibitors during development have been described (Spratt, 1952, 1956), and particular events of development such as brain formation, heart formation, etc., seem to be underlain by both quantitatively and qualitatively different metabolic processes. The inhibition of amino acid metabolism by INH in our experiments indicates the importance of this metabolism for the normal development of the nervous system in early stages. On the contrary, the remaining embryonic structures with no major alterations during the first days of incubation. However, the 1NHinduced disorders in the chick embryo are not totally prevented by supplemental pyridoxine; therefore, the effect of INH cannot be interpreted purely as an inhibitory mechanism acting on pyridoxine. Unrelated side effects of the INH must be considered. This point needs further investigation. Finally, we wish to emphasize the scanty amount of circulating blood in INHtreated embryos, which was interpreted as the result of a severe anaemia, and which could probably lead to hypoxia. We cannot dismiss the fact that the effects of INH, including the embryonic death, could be attributed to anaemia. We attribute this anaemia to the antivitamin B6 action of INH, because it is prevented by supplemental pyridoxine. REFERENCES A. E. (1960). Pyridoxal phosphate. In The Enzymes, 2nd ed. vol. n (ed. P. D. Boyer, H. H. Lardy & K. Myrback), pp. 113-184. New York and London: Academic Press. CARLTON, W. W. & KREUTZBERG, G. (1966). Isonicotinic acid hydrazide-induced spongy degeneration of the white matter in the brains of Pekin ducks. Am. J. Path. 48, 91—105. CARTER, S. B. (1964). Problems of interpreting teratogenic effects in eggs. Proc. Eur. Soc. Study Drug Tox. 5, 142-148. CONTI, G. & MILIO, G. (1965). Action de la vitamine et de l'antivitamine B6 sur le developpement de l'embryon de Poulet. /. Embryol. exp. Morph. 14, 37-51. CONTI, G. & MILIO, G. (1966a). Action de la vitamine et de l'antivitamine B6 sur l'intestin d'embryon de Poulet cultive in vitro. Archs BioL, Paris 77, 79-91. CONTI, G. & MILIO, G. (19666). Action de la vitamine et de l'antivitamine B6 sur le poumon d'em bryon de Poulet cultive in vitro. Archs BioL, Paris 77,93-106. BRAUNSTEIN, Effects of isonicotinic acid hydrazide 219 H. H. (1969). The role of vitamin Bc in the toxicity of hydrazines. Ann. N.Y. Acad. Sci. 166, 136-145. CRAVENS, W. W. (1952). Vitamin deficiencies and antagonists. Ann. N.Y. Acad. Sci. 55, 188-195. CRAVENS, W. W. & SNELL, E. E. (1949). Effects of desoxypyridoxine and vitamine B6 on development of the chick embryo. Proc. Soc. exp. Biol. Med. 71, 73-76. DURAISWAMI, P. K. (1961). Experimental induction of congenital defects with isonicotinic acid hydrazide. Indian J. med. Res. 49, 631-634. GEBHARDT, D. O. E. (1968). The teratogenic action of propylene glycol (propanediol-1,2) and propanediol-1,3 in the chick embryo. Teratology 1, 153-162. KILLAM, K. F. & BAIN, j . A. (1957). Convulsant hydrazides. I. In vitro and in vivo inhibition of vitamin B6 enzymes by convulsant hydrazides. /. Pharmac. exp. Ther. 119, 255-262. LAMPERT, P. W. & SCHOCHET, S. S. (1968). Electron microscopic observations on experimental spongy degeneration of the cerebellar white matter. /. Neuropath, exp. Neurol. 27, 210-220. LANDAUER, W. (1945). Rumplessness of chicken embryos produced by the injection of insulin and other chemicals. /. exp. Zool. 98, 65-77. LEVY, L. (1969). Mechanism of drug-induced vitamin B6 deficiency. Ann. N.Y. Acad. Sci. 166, 184-190. LEWIS, H. B. & IZUME, S. S. (1926). The influence of hydrazine and its derivatives on metabolism. H. Changes in the non-proteins nitrogenous constituents of the blood and in the metabolism of injected glycine in hydrazine intoxication. /. biol. Chem. 71, 33-49. MCCORMICK, D. B. & SNELL, E. E. (1961). Pyridoxal phosphokinases. II. Effects of inhibitors. /. biol. Chem. 236, 2085-2088. CORNISH, MCLAUGHLIN, J., MARLIAC, J. P., VERRET, M. J., MUTCHLER, M. K. & FITZHUGH, O. G. (1963). The injection of chemicals into the yolk sac of fertile eggs prior to incubation as a toxicity test. Toxic, appl. Pharmac. 5, 760-771. MURPHY, L. (1967). Factors influencing teratogenic response to drugs. In Teratology: Principles and Techniques (ed. J. G. Wilson & J. Warkany). Chicago and London: University of Chicago Press. SIMONSEN, D. G. & ROBERTS, E. (1967). Influence of hydrazine on the distributions of free amino acids in mouse liver. Proc. Soc. exp. Biol. Med. 124, 806—811. SPRATT, N. T. JR (1952). Metabolism of the early embryo. Ann. N.Y. Acad. Sci. 55, 40-50. SPRATT, N. T. JR (1956). Nutrient necessities in chick development. In Symposium on Embryonic Nutrition, Brown University (ed. D. Rudnick). Chicago and London: University of Chicago Press. YONEDA, M. & ASANO, N. (1953). Competitive action of isonicotinic acid hydrazide and pyridoxal in the amino acid decarboxylation of Escherichia coli. Science, N. Y. Ill, 277-279. YONEDA, M., KATO, N. & OKAJUNA, K. (1952). Competitive action of isonicotinic acid hydrazide and vitamin B6 in the formation of indole by E. coli. Nature, Lond., 170, 803. YONEZAWA, T., MORI, T. & NAKATANF, Y. (1969). Effects of pyridoxine deficiency in nervous tissue maintained in vitro. Ann. N.Y. Acad. Sci. 166, 146-157. (Manuscript received 25 April 1972)