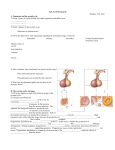
Survey
* Your assessment is very important for improving the workof artificial intelligence, which forms the content of this project
Am J Physiol Endocrinol Metab 302: E4–E18, 2012. First published October 25, 2011; doi:10.1152/ajpendo.00488.2011. Perspectives Hirsutism, virilism, polycystic ovarian disease, and the steroid-gonadotropin-feedback system: a career retrospective Virendra B. Mahesh Department of Physiology and Endocrinology, Georgia Health Sciences University, Augusta Georgia Submitted 19 September 2011; accepted in final form 20 October 2011 estrogens; androgens; progesterone IN THIS CAREER RETROSPECTIVE ARTICLE, the author describes briefly how he started working in the field of steroid hormones, which led to work in the area of hirsutism, virilism, and polycystic ovarian disease. These initial studies led to a number of interlocking questions resulting in a detailed study of the steroid-gonadotropin feedback system, leading into a lifelong career of scientific investigation from 1956 to 2010. The approach for solving the questions generated was inspired by the literature at the time the research was done as well as the incorporation of newer techniques as they developed. This article covers an extensive area of reproductive biology, and it is not possible to refer to all the literature surrounding the Address for reprint requests and other correspondence: V. B. Mahesh, Dept. of Physiology and Endocrinology, Georgia Health Sciences University, 1120 15th St., Augusta Georgia 30912 (E-mail: [email protected]). E4 research due to the breadth of the area covered. However, it is fully referenced in the articles cited and the periodic reviews on the subject. In the mid 1950s, there was no clear understanding of how steroid hormones exerted their biological action. In the area of glucocorticoids, based on the metabolism of cortisol (11,17␣,21trihydroxy-4-pregnen-3,20-dione) as well as the effectiveness of cortisol compared with cortisone (17␣,21-dihydroxy-4-pregnen-3,11,20-trione) in tissues in which the conversion of cortisone to cortisol did not take place, it was postulated that cortisol might be the active hormone (51, 82, 183). On the other hand, working with placental isocitric dehydrogenase in vitro, Talalay and Williams-Ashman in 1958 (212) explained the action of estradiol as a coenzyme in a redox system. To investigate whether a hormone was required to participate in an oxidation-reduction cascade to exert biological action, Bush and Mahesh (52) studied the factors that governed the reduction of the 11-keto group to the 11-hydroxyl group in corticosteroids. It was postulated that since the ␣-side of the molecule is flat in the 3-keto-4-ene and 3␣,5␣-reduced steroids, the enzyme involved in reducing the 11-keto group could approach the steroid from the ␣-side with ease and reduce it to the 11-hydroxyl group. On the contrary, in the 3␣,5-steroids, Ring A of the steroid is at a right angle to the rest of the molecule and thus would provide steric hindrance in reduction at the 11-position. Thus, the synthesis 3␣,11-dihydroxy-5␣androstan-17-one; 3␣-hydroxy-5␣-androstan-11, 17-dione; 3␣,11-dihydroxy-5-androstan-17-one; 3␣-hydroxy-5-androstan-11,17-dione, and 4-androsten-3,11,17-trione was carried out and their metabolism studied in the human. The results clearly showed the reduction of the 11-ketone to the 11hydroxyl compounds in 3-keto-4-ene and 3␣,5␣-steroids but not in the 3␣,5-steroids. To slow down the metabolism of cortisol in Ring A of the steroid, 2␣-methyl-cortisol and 2␣-methyl-cortisone were synthesized and tested for biological activity (70). The 2␣-methyl-cortisol was found to be very active biologically and the 2␣-methyl-cortisone had very little biological activity. Based on the structural requirements for reduction of the 11-ketone to the 11-hydroxyl group as determined by Bush and Mahesh (52), the 2␣-methyl group would provide enough steric hindrance to prevent reduction of the 11-ketone to the 11-hydroxyl group. This was verified experimentally by Bush and Mahesh (53), leading to the conclusion that the 11-hydroxyl group of cortisol did not have to undergo any metabolism for the exertion of biological activity. The higher biological activity of 9␣-fluoro steroids was also explained, in part, due to a greater persistence of the 11hydroxyl group during metabolism (55). Jensen also provided proof that estradiol exerted its biological activity without being converted to estrone (102). Further examination of the metabolism of cortisol in various rat tissues identified the kidney as 0193-1849/12 Copyright © 2012 the American Physiological Society http://www.ajpendo.org Downloaded from http://ajpendo.physiology.org/ by 10.220.33.3 on June 18, 2017 Mahesh VB. Hirsutism, virilism, polycystic ovarian disease, and the steroid-gonadotropin-feedback system: a career retrospective. Am J Physiol Endocrinol Metab 302: E4 –E18, 2012. First published October 25, 2011; doi:10.1152/ajpendo.00488.2011.—This career retrospective describes how the initial work on the mechanism of hormone action provided the tools for the study of hirsutism, virilism, and polycystic ovarian disease. After excessive ovarian and or adrenal androgen secretion in polycystic ovarian disease had been established, the question whether the disease was genetic or acquired, methods to manage hirsutism and methods for the induction of ovulation were addressed. Recognizing that steroid gonadotropin feedback was an important regulatory factor, initial studies were done on the secretion of LH and FSH in the ovulatory cycle. This was followed by the study of basic mechanisms of steroid-gonadotropin feedback system, using castration and steroid replacement and the events surrounding the natural onset of puberty. Studies in ovariectomized rats showed that progesterone was a pivotal enhancer of estrogen-induced gonadotropin release, thus accounting for the preovulatory gonadotropin surge. The effects of progesterone were manifested by depletion of the occupied estrogen receptors of the anterior pituitary, release of hypothalamic LHRH, and inhibition of enzymes that degrade LHRH. Progesterone also promoted the synthesis of FSH in the pituitary. The 3␣,5␣-reduced metabolite of progesterone brought about selective LH release and acted using the GABAA receptor system. The 5␣-reduced metabolite of progesterone brought about selective FSH release; the ability of progesterone to bring about FSH release was dependent on its 5␣-reduction. The GnRH neuron does not have steroid receptors; the steroid effect was shown to be mediated through the excitatory amino acid glutamate, which in turn stimulated nitric oxide. These observations led to the replacement of the long-accepted belief that ovarian steroids acted directly on the GnRH neuron by the novel concept that the steroid feedback effect was exerted at the glutamatergic neuron, which in turn regulated the GnRH neuron. The neuroprotective effects of estrogens on brain neurons are of considerable interest. Perspectives STEROID-GONADOTROPIN-FEEDBACK SYSTEM: CAREER RETROSPECTIVE the major organ for the oxidation of cortisol to cortisone (139, 140), a finding that proved in later years to be of great importance in the action of mineralocorticoids. These initial studies in the mechanism of steroid hormone action required the development of methods for the separation, characterization, and measurement of steroid hormone metabolites in human biological fluids and made studies in hirsutism, virilism, and polycystic ovarian disease possible. HIRSUTISM, VIRILISM AND POLYCYSTIC OVARIAN DISEASE Steroid Secretion Patterns vein, and ovarian vein steroids were measured before and after ACTH and human chorionic gonadotropin (HCG) to further establish the adrenal or ovarian source of excessive androgens (176). Even before the availability of radioimmunoassay (RIA) for the determination of serum gonadotropins, levels of luteinizing hormone (LH) secretion in urine were measured throughout the menstrual cycle using an anti-HCG antiserum. Levels of LH were found to show a peak either coinciding with or preceding a rise in basal body temperature which was then considered to be an indication of ovulation (203). With the availability of RIA for measuring serum LH, our group as well as others demonstrated a high pulsatile level of LH in patients with polycystic ovary syndrome (80).Initially, there was no explanation for this high pulsatile secretion of LH, and a hypothalamic defect was postulated. Most patients with polycystic ovary syndrome show good estrogenic vaginal smears due to ovarian secretion of estrogens and the peripheral conversion of androgens to estrogens. Using pituitary stalk-sectioned rats in which aluminum foil was placed after stalk resection to prevent regeneration of the hypothalamic blood supply, Greeley et al. (87) showed that the rats still responded to luteinizing hormone-releasing hormone [LHRH; also referred to as gonadotropin-releasing hormone (GnRH)] in the secretion of LH and FSH. Furthermore, if such rats were treated with estradiol, the pituitary showed enhanced sensitivity to the release of LH (88). Studies of Legan and Karach (110) showed that in long-term ovariectomized rats, a single injection of estradiol brought about daily LH surges, whereas a single injection of progesterone initially enhanced the estrogen-triggered surge of gonadotropins and then promptly brought about the extinction of the estrogen-induced LH surge. There were no multiple surges of gonadotropins on subsequent days of progesterone administration after the first surge took place. Thus, it appeared that persistent estrogen stimulation to the hypothalamic-pituitary axis could cause persistent LH surges in the human, which could only be dampened by luteal levels of progesterone. This was shown to occur in one patient with polycystic ovary syndrome who underwent wedge resection of the ovary; the high pulsatile levels of LH persisted only until the luteal rise of progesterone occurred after the first ovulation (138). This change in hormonal profile tends to explain the beneficial effects of wedge resection on the ovulatory process. The pivotal role of progesterone in initiating the surge of LH and FSH secretion and also the termination of the surge will be discussed in detail later in this article. Other studies on hirsutism, virilism, and steroid secretion carried out were in cases with ovarian stromal hyperplasia (131, 133), arrhenoblastoma (93, 132), adrenal rest tumor of the ovary (181), virilizing adrenal tumors (124, 127), and delayed onset of congenital adrenal hyperplasia (128). 11Hydroxyestrone was also isolated in a case of feminizing adrenal carcinoma (130). Three overall questions emerged from the aforementioned studies. The first question was whether there was a treatment to induce ovulation in patients with polycystic ovarian syndrome without undergoing surgical procedures such as the wedge resection of the ovary. The second question concerned the methods that could be employed for the management of hirsutism. The third question was whether polycystic ovary syndrome was a genetic or an acquired disorder. Initial attempts to AJP-Endocrinol Metab • doi:10.1152/ajpendo.00488.2011 • www.ajpendo.org Downloaded from http://ajpendo.physiology.org/ by 10.220.33.3 on June 18, 2017 The earliest case studied by the author was a case of twin sisters, one of whom had undergone severe psychological stress and had a sudden onset of hirsutism (54). The sister with hirsutism had a very high excretion of androgen metabolites in her urine compared with her normal sister and was hyperresponsive to adrenocorticotropic hormone (ACTH) stimulation in androgen production. That the excessive androgens were coming from the adrenal gland was demonstrated by large quantities of dehydroepiandrosterone (3-hydroxy-5-androsten-17-one) and androstenedione (4-androsten-3,17-dione) in the adrenal vein blood. In the mid 1950s, wedge resection of the polycystic ovaries was the standard way to treat polycystic ovarian disease. The ovarian wedges obtained from such patients, after obtaining informed consent, were extracted, and the steroids obtained were characterized. Several ovaries contained large quantities of dehydroepiandrosterone that was characterized by chromatographic properties and infrared spectroscopy along with 17␣-hydroxy-5-pregnenolone (3,17␣dihydroxy-5-pregnen-20-one). Some ovaries contained large quantities of androstenedione compared with normal ovaries (119, 120, 125). This was the first demonstration of androgen secretion by the polycystic ovary, and the background references are provided in two reviews (122, 125). In vitro incubation studies showed that the ovaries containing large quantities of dehydroepiandrosterone converted less substrate to androstenedione compared with normal ovaries, suggesting diminished 3-hydroxysteroid dehydrogenase activity, and those containing large quantities of androstenedione showed lower aromatase activity. Since androstenedione and dehydroepiandrosterone are weak androgens and we were not able to extract significant amounts of testosterone from the polycystic ovaries studied, it was of interest to determine whether these weak androgens could be converted peripherally into testosterone. Oral administration of dehydroepiandrosterone and androstenedione to women resulted in elevation of plasma testosterone levels (121). Since in our earlier study (54) the adrenal was the source of excessive androgens, a finding that had also been reported by others (122, 125), it was important to determine whether, in a particular patient, the adrenal or the ovary or both were the source of excessive androgens. Thus, urinary androgen metabolites were measured before and after adrenal and ovarian suppression (126). The steroids were also measured after ovarian stimulation with human pituitary follicle-stimulating hormone (FSH) (123). The results showed that different patients could have adrenal oversecretion of androgens, ovarian oversecretion of excessive androgens, or both. Human FSH enhanced ovarian hypersecretion of androgens and also caused multiple ovulations. In a subsequent study, peripheral, adrenal E5 Perspectives E6 STEROID-GONADOTROPIN-FEEDBACK SYSTEM: CAREER RETROSPECTIVE answer the third question were to determine whether there was a chromosomal abnormality in polycystic ovary syndrome patients. No chromosomal abnormalities were found in patients with polycystic ovary syndrome (56); therefore, animal models were constructed in an attempt to answer that question. Induction of Ovulation in Patients with Polycystic Ovary Syndrome Studies on the Use of Antiandrogens for the Management of Hirsutism Management of hirsutism in patients with polycystic ovary syndrome has been difficult, as the suggested approaches were ovarian suppression by use of contraceptive steroids when the source of excessive androgens was the ovary, or adrenal suppression when the source of androgens was the adrenal, or a combination of both treatments when the source of excessive androgens were the ovary and the adrenal. Establishment of an ovarian or adrenal source required extensive diagnostic tests, and long-term suppression of the adrenal or the ovary was not an easily acceptable choice. Thus, another approach was the use of an antiandrogen, 17␣-methyl-B-nor-testosterone. In male rats, 17␣-methyl-B-nor-testosterone reduced seminal ves- Animal Models for Polycystic Ovarian Disease in Humans The question whether human polycystic ovary syndrome was a genetic disorder or whether it could be caused by excessive androgen secretion is a difficult one and it is still being debated. The possibility of an androgen-related disorder was indicated by our early experiments that showed that the administration of large quantities of dehydroepiandrosterone or androstenedione in immature rats resulted in ovulatory failure and the presence of polycystic ovaries (202). A detailed examination of the effects of dehydroepiandrosterone on ovulatory failure was thus carried out (106). Administration of dehydroepiandrosterone to 27-day-old female rats resulted in ovulatory type serum FSH and LH surge on day 30 of life, and the animals exhibited either constant estrous or constant diestrous vaginal smears with either polycystic ovaries or ovaries containing corpus luteum-like structures. With time, all ovaries became polycystic. Serum FSH levels were elevated compared with control rats, serum LH levels were similar to those in control rats, and serum prolactin was elevated. The ovary was responsive to gonadotropin treatment, and the pituitary was responsive to LHRH stimulation. Discontinuation of the dehydroepiandrosterone treatment resulted in a few irregular ovulatory cycles followed by normal cyclicity. Thus, this animal model was very similar to the human one, with the exception that prolactin rather than LH was elevated. Further studies with precocious ovulation showed that the gonadotropin surge resulting in ovulation could be blocked with central nervous system blocking agents such as phenobarbital and reserpine (106). Precocious ovulation appeared to be mediated by the conversion of dehydroepiandrosterone to estrogens, because the nonaromatizable androgen 5␣-dihydrotestosterone did not cause such an ovulation (107). Furthermore, cyanoketone, an inhibitor of 3-hydroxysteroid dehydrogenase, which blocks the conversion of dehydroepiandrosterone to estrogen, prevented vaginal patency, and dehydroepiandrosterone-induced precocious ovulation (107). A study of the steroid profile after dehydroepiandrosterone administration showed an elevation in blood estradiol followed by depletion of the cytoplasmic estradiol receptors in the hypothalamus and the pituitary and the gonadotropin surge leading to ovulation (180). That these events occur in the natural process of ovulation will be discussed later in this article. The restoration of normal cyclicity in dehydroepiandrosterone-treated rats occurred only when the AJP-Endocrinol Metab • doi:10.1152/ajpendo.00488.2011 • www.ajpendo.org Downloaded from http://ajpendo.physiology.org/ by 10.220.33.3 on June 18, 2017 In attempts to search for agents other than human pituitary FSH or human postmenopausal gonadotropins that were very expensive and difficult to obtain at that time, a variety of compounds were tested for their ability to induce ovulation. Clomiphene {1-[p(-diethylaminoethoxy)phenyl]-1,2-diphenyl-2-chloroethylene} was sent to us to be tested as a contraceptive agent by a drug company. In immature female rats, this compound was found to stimulate uterine weight, and in male rats the seminal vesicle and ventral prostate weights, in very low doses, whereas it inhibited them at high doses (200). In the absence of estrogens in the ovariectomized rat, Clomiphene acted as a weak estrogen. It acted as an antiestrogen in the presence of estrogens. In unilateral ovariectomized rats, Clomiphene in low doses increased ovarian weight, thus indicating antagonism of estrogen suppression on the hypothalamic-pituitary axis and the stimulation of gonadotropin secretion (198). The antiestrogenic activity of Clomiphene was further demonstrated by its ability to inhibit the uptake of tritiated estradiol in the rat uterus and pituitary gland (201). In the human, Clomiphene was able to induce ovulation in a variety of unovulatory patients, including patients with polycystic ovary syndrome (94, 199). It was also able to stimulate spermatogenesis in men (103). Clomiphene was also able to induce ovulation in ChiariFrommel syndrome, in which estrogen levels are very low (92). Thus, Clomiphene became the accepted first treatment of choice for the induction of ovulation prior to the use of human menopausal gonadotropins followed by HCG (95). With the availability of RIA for gonadotropin secretion, Clomiphene treatment was shown to result in an ovulatory type of LH and FSH secretion in a variety of anovulatory patients (78, 79). It was also demonstrated that, in patients that were resistant to Clomiphene in the induction of ovulation, 11 of the 13 patients treated ovulated, and there were five pregnancies if the Clomiphene treatment was preceded by the synthetic glucocorticoid dexamethasone (213). It was shown previously in experimental animals that dexamethasone was able to bring about FSH release, and this perhaps aided in the ovulation (36) icle and ventral prostate weights and counteracted the growthpromoting effects of testosterone on these organs (142). However, in castrated male rats, it stimulated the growth of seminal vesicles and ventral prostate, indicating weak androgenic effects. 17␣-Methyl-B-nor-testosterone also enhanced the effect of testosterone in suppressing testicular weight, indicating a weak antigonadotropic effect. The antigonadotropic effect was confirmed in female rats, in which it decreased ovarian weight and the number of ovulations (214). In the human, it brought about a significant decrease in sebum production rate in the forehead as well as a decrease in facial hair growth, as determined by the weight of hair shaved, starting at 30 days after it was administered (65, 221). However, the concern that a patient on an antiandrogen might become accidentally pregnant, causing fetal abnormalities, has prevented the use of antiandrogens in the management of hirsutism. Perspectives STEROID-GONADOTROPIN-FEEDBACK SYSTEM: CAREER RETROSPECTIVE THE STEROID-GONADOTROPIN FEED BACK SYSTEM The abovementioned experimental studies showed that, in animals with normal ovaries and a normal hypothalamicpituitary axis, the administration of androgens resulted in the formation of polycystic ovaries and an altered gonadotropin secretion pattern. Therefore, a detailed examination of the steroid-gonadotropin feedback system was undertaken. These studies included examining the role of FSH and LH in the ovulatory cycle, steroid-gonadotropin feedback before and after puberty, steroid and gonadotropin changes during puberty, and the ovarian signals for the preovulatory gonadotropin surge. Early Work on the Role of FSH and LH in the Ovulatory Process Before the advent of RIA for pituitary FSH and LH in serum, bioassays were used for their determination. In the cycling rat there was an 50% decrease in pituitary LH and FSH content from the morning to the late evening on the day of proestrus (84). Such a decrease was also observed in pubertal rats preceding their first ovulation. In immature rats in which follicular development was initiated with a small dose of PMSG and the endogenous gonadotropin surge blocked by phenobarbital, ovulation could be induced with purified pituitary FSH that did not contain enough LH contaminant to cause ovulation. In the hamster, antiserum prepared against a preparation of LH with ample FSH contaminant blocked ovulation (85). However, when the anti-FSH component was adsorbed out, the ovulation blocking potency of the antiserum was greatly reduced. In addition, ovulation blocked by anti-LH could be reinstituted by injection of an FSH preparation that contained very little or no LH contamination. (85). These results showed a prominent role of FSH in ovulating the mature ovarian follicle. In view of a common ␣-subunit in FSH, LH, and TSH, the ability to ovulate a mature ovarian follicle by TSH, prolactin, and the two LH subunits was also tried, but they did not induce ovulation (129). Although in the rodent the need for the secretion of FSH as a part of the preovulatory gonadotropin surge can be explained on the basis of the growth of the ovarian follicles for the next cycle, the physiological role of FSH at the time of the preovulatory gonadotropin surge, if any, in the human is still unclear. In this regard, Strott et al. (211) have reported several cases of short luteal phase in women in whom the only abnormality was a dulled or misplaced FSH peak during the ovulatory surge. The role of the ovary in regulating the preovulatory surge of gonadotropins was shown by the fact that removing the ovary on the morning of diestrus II blocked the preovulatory LH surge (86). Ovariectomy at 6:00 PM on diestrus II or 9:00 AM on proestrus resulted in a lower LH surge at proestrus. Injection of progesterone on proestrus before the preovulatory surge of gonadotropins postponed the preovulatory surge for one day (86). Examination of the Steroid-Gonadotropin Feedback System by Castration and Steroid Replacement Immature rats castrated at day 26 of age showed a rise of LH and FSH by 8 h in male rats and in 24 h for LH and 48 h for FSH in female rats (72). These levels could be suppressed by the administration of estradiol or testosterone. In the female rat, ovariectomy on day 26 of age and treatment with increasing doses of estradiol for 5 days showed a return to intact gonadotropin levels within the physiological dose range of estradiol as judged by uterine weight (145). Increasing the dose of estrogens showed an increase in gonadotropin secretion due to the positive feedback effect, followed by suppression at higher dose levels. This method also provided a means of evaluating the biological activity of several synthetic estrogens. Progesterone by itself did not suppress gonadotropins in the ovariectomized rat (144). Using seminal vesicle and ventral prostate weights, similar studies tested the steroid feedback system in male rats (73). Ovariectomy and estrogen replacement in 26-, 45-, and 95-day-old rats showed that the dose of estradiol required to decrease serum gonadotropins was four- to sixfold higher in 45- and 95-day-old rats compared with 26-day-old rats, indicating that a maturation of the feedback system after puberty even though the increase in uterine weight caused by estrogen administration was comparable in the three groups (74). This change in sensitivity to the negative feedback effect of estradiol may in part be mediated by endogenous endorphins that are suppressive of gonadotropin secretion (188). An endorphin antagonist, naloxone, brings about a large increased release of LH at all times in the rat ovulatory cycle except at the time of the gonadotropin surge. Measurement of the -endorphin content in the hypothalamus of the immature ovariectomized rat treated with estradiol and progesterone or triamcinilone acetonide (9␣-fluoro-11,21dihydroxy-1,4-pregnadiene-3,20-dione-16␣,17␣-acetonide) to induce the preovulatory type surge of gonadotropins resulted in an estradiol-induced decrease in -endorphin preceding and during the LH surge, which was maintained the following morning (37). However, progesterone and triamcinilone acetonide brought back the -endorphin levels AJP-Endocrinol Metab • doi:10.1152/ajpendo.00488.2011 • www.ajpendo.org Downloaded from http://ajpendo.physiology.org/ by 10.220.33.3 on June 18, 2017 high levels of dehydroepiandrosterone in blood were reduced (178). Because the earlier studies were done with immature rats, it was important to establish that the administration of dehydroepiandrosterone caused polycystic ovaries and ovulatory failure in adult rats as well. The effect of dehydroepiandrosterone in causing polycystic ovaries in the adult rat was shown by Ward et al. (217). Since polycystic ovary syndrome in women is often associated with obesity and insulin resistance and insulin type growth factors hyperstimulate theca cells to produce androgens, the direct effect of androgens on the ovary were examined. Hypophysectomized immature rats, when ovulated with pregnant mare’s serum gonadotropin (PMSG) and HCG, showed follicular atresia and decreased ovulation rate when treated with the nonaromatizable androgen 5␣-dihydrotestostrone (2). That the effect of the androgen was on the ovary and not gonadotropin secretion was demonstrated by the fact that in the PMSGtreated rat the preovulatory gonadotropin surge was unaffected even though the number of ovulations was reduced (62).The effect of the androgen could be reduced by pretreatment with estrogens. Thus, androgens could cause abnormal follicular development, altered ovarian steroidogenesis, and ovulatory failure. Evidence was also found for an autoregulatory effect of androgens on ovarian theca cell androgen production (207, 208). E7 Perspectives E8 STEROID-GONADOTROPIN-FEEDBACK SYSTEM: CAREER RETROSPECTIVE to control levels on the morning after the gonadotropin surge, thus appearing to reinstate the opioid inhibition. Changes Occurring in Steroid Levels During the Preovulatory Gonadotropin Surge and Initiation Of Puberty Effect of Progesterone and Corticoids in Inducing the Preovulatory Type of Surge of Gonadotropins Extensive work done in this area showed that progesterone played an important part in the preovulatory gonadotropin surge. The work consisted of defining the role of progesterone and its mechanism of action through depletion of occupied estrogen receptors in the anterior pituitary; hypothalamic control of the secretion of LHRH, regulation of peptidase activity that degrades LHRH, regulation of LH- and FSH- mRNA levels, and selective secretion of FSH and LH by progesterone metabolites. The work has been the subject of periodic reviews Fig. 1. Stimulatory effects of progesterone on the hypothalamus in the release of LHRH and stimulation of 17-hydroxysteroid dehydrogenase activity in the pituitary. It also had direct action on the pituitary in increasing the sensitivity of the pituitary to LHRH in release of LH and synthesis of new LH and FSH. Progesterone also inhibited peptidase activity in the hypothalamus and pituitary, slowing degradation of LHRH and increasing greater availability of LHRH for LH secretion. Enhanced 17-hydroxysteroid dehydrogenase activity brings about increased conversion of estradiol to estrone in the pituitary, resulting in decreased nuclear occupancy of the estrogen receptor. Progesterone thus decreased the inhibitory action of estrogens on the pituitary on LH release and decreased progesterone receptor synthesis. Decreased progesterone receptors prevent the stimulatory effect of progesterone on LH secretion. Stimulatory pathways are shown in green, inhibitory pathways in red. AJP-Endocrinol Metab • doi:10.1152/ajpendo.00488.2011 • www.ajpendo.org Downloaded from http://ajpendo.physiology.org/ by 10.220.33.3 on June 18, 2017 Ovarian and uterine histology and serum gonadotropins were measured in Holtzman rats from days 22 to 40 of age, in which ovulation occurred most consistently on day 38 of age (64).The percentage of large follicles showed a linear growth from day 22 onward. The endometrial stroma, myometrium, and luminal epithelium started growing on day 32 of age along with an increase in uterine weight. These changes in uterine histology were early indicators of ovarian estrogen secretion. An early small increase in serum LH occurred on day 34, with the preovulatory LH and FSH surge on day 38. These observations suggested that ovarian estrogens may be the trigger for the initiation of puberty. Measurement of steroids during the natural onset of puberty showed an increase in blood estradiol, progesterone, and testosterone preceding vaginal opening and an increased sensitivity of the pituitary to LHRH (179). Correlated with the gonadotropin surge was a fall in the cytoplasmic estrogen receptors of the pituitary and the hypothalamus due to nuclear translocation by the secreted estradiol. The rise in estradiol and the depletion of cytoplasmic estrogen receptors in the pituitary and hypothalamus were similar to what occurred in normally ovulating rat (89) and immature rats induced to ovulate by the administration of PMSG (177). Similar events also took place in the precocious ovulation induced by the administration of dehydroepiandrosterone (180). In the estrogen-primed ovariectomized rat, an injection of 2 g of estradiol brought about a decrease in the estrogen receptor content of the pituitary without altering the estrogen receptor mRNA levels, which only increased 12 h after the estrogen injection in preparation for the estrogen receptor replenishment that started at 18 h. Thus, the initial decline in estrogen receptor content was not due to loss of estrogen receptor mRNA but to accelerated estrogen receptor processing (61). In the uterus, similar changes took place, and progesterone was shown to delay estrogen receptor replenishment (223, 224). Pituitary ultrastructure of the gonadotropes during the preovulatory surge also showed degranulation of LH-containing gonadotropes a few hours prior to the degranulation of FSH-containing gonadotropes, which was consistent with the start of the LH surge a little earlier than the FSH surge. (63). Extensive work was also done on the role of androgens and LHRH in the initiation of puberty in the male rat (135, 160, 161–164). A detailed description, however, is outside the scope of this article. on the subject (14, 16, 17 113–115, 134). The results of these studies are also summarized in Figs. 1 and 2. Defining the role of progesterone in the preovulatory surge of gonadotropins. Even though the administration of estrogens to ovariectomized rats was able to induce an LH surge, the LH surge was only of a magnitude of ⬃10% of the preovulatory gonadotropin surge. Thus, experiments were done using doses of estrogens that were able to reduce the postcastration rise of gonadotropins but not prevent it, in combination with various doses of progesterone (146). The dose of estradiol used was enough to induce progesterone receptors but only cause a minimal if any estrogen-induced LH surge. The doses of progesterone used were able either to stimulate the release of LH and FSH or to suppress them with the next higher dose. The pattern of the lower dose being stimulatory and the next higher dose being inhibitory was persistent in several protocols and several experiments. Such a pattern has been shown to be important in the human menstrual cycle where a low level of progesterone enhanced the preovulatory LH surge while the higher level helped terminate the surge (101). The stimulatory dose of progesterone was able to mount a preovulatory-type gonadotropin surge of LH and FSH similar to what is seen at the time of proestrus. The sensitivity of the pituitary to LHRH was enhanced by the stimulatory dose of progesterone and suppressed by the inhibitory dose of progesterone (148). This pattern was also exhibited by the ability of progesterone to decrease occupied estrogen receptors of the anterior pituitary and the effect of progesterone and 5␣-dihydroprogesterone to attenuate estrogen-induced prolactin release, which will be discussed later in this article. In the immature castrated male rat, the dose of estradiol needed to reduce the levels of gonadotropins comparable to what was achieved with the 0.1 g/kg body wt dose of estradiol in female rats was 2.0 g/kg body wt (149). This was Perspectives STEROID-GONADOTROPIN-FEEDBACK SYSTEM: CAREER RETROSPECTIVE In the adult rat, measurement of both occupied and unoccupied estrogen receptor content was determined in the pituitary and the uterus after 0.8, 2.0, and 4.0 mg/kg body wt progesterone. The 0.8 and 4.0 mg/kg body wt dose of progesterone was stimulatory to gonadotropin release, whereas the 2.0 mg/kg body wt dose was inhibitory. The finding in the immature rat (146, 148) and the adult rat (57, 75) of a smaller dose of progesterone to be stimulatory to gonadotropin release and the next higher dose to be inhibitory was of great interest. It was shown that in the human menstrual cycle the initial increase in progesterone around the time of ovulation was stimulatory to gonadotropin release, whereas a secondary rise coincided with the termination of the LH surge (101). High pulsatile levels of LH were also inhibited by rising levels of progesterone after ovulation in a patient with wedge resection of the ovary (138). In the pituitary and the uterus, the decrease in estrogen receptors occurred only in the occupied form and not the unoccupied estrogen receptors (75). However, whereas the pituitary showed a dose dependency of progesterone of the effect, the 0.8 and 4.0 mg/kg body wt doses being more effective than the 2.0 mg/kg body wt dose in reducing occupied estrogen receptors, the uterus showed a systematic dose response. This was an interesting tissue difference. In the estrogen-primed progesterone-treated group, 0.8 and 4.0 mg/kg body wt progesterone increased estrogen receptor mRNA levels in the pituitary within 1 h of administration, whereas the 2.0 dose was ineffective. Despite this early increase in estrogen receptor mRNA levels by the 0.8 and 4.0 mg/kg body wt progesterone, these doses still depleted the occupied estrogen receptors of the pituitary. In the uterus, progesterone treatment in estrogen-primed rats did not alter the estrogen receptor mRNA levels but depleted the occupied estrogen receptors (197). The mechanism for the decrease of occupied estrogen receptors of the pituitary and the uterus was by the stimulation of 17-hydroxysteroid dehydrogenase by progesterone (71, 76). This converted estradiol to estrone, which had a lower affinity for the estrogen receptor than estradiol, thereby reducing the suppressive effect of estrogens on gonadotropin secretion at the level of the pituitary. Accelerated estrogen receptor processing was also indicated. The antagonism of estrogen action by progesterone and 5␣-dihydrotestosterone and their dose dependency was also shown in the inhibition of estrogen-induced prolactin release (30, 31, 38). Such dose dependence was also found in the inhibition of estrogen-induced prolactin release by 5␣-dihydroprogesterone (34). Surprisingly, the antiandrogen flutamide could not only block the action of 5␣-dihydrotestosterone but that of progesterone as well (32). This was also true for RU-486, which blocked progesterone as well as 5␣-dihydrotestosterone effects. HYPOTHALAMIC REGULATION OF LHRH SECRETION REGULATED BY PROGESTERONE. It has already been stated that progesterone given to estrogen-primed ovariectomized rats increases the sensitivity of the pituitary to LHRH in the release of LH (148). Progesterone also stimulates the secretion of LHRH from the hypothalamus in the estrogen-primed rat (182). The regulators of LHRH secretion, catecholamine as well as neuropeptide Y, are also released by progesterone in the medial basal hypothalamus by progesterone and the glucocorticoid triamcinilone acetonide in the estrogen-primed ovariectomized rat (15, 26). The release of the above are more acutely related to LH AJP-Endocrinol Metab • doi:10.1152/ajpendo.00488.2011 • www.ajpendo.org Downloaded from http://ajpendo.physiology.org/ by 10.220.33.3 on June 18, 2017 perhaps due to the masculinization of the hypothalamus neonatally by estrogens, resulting in the change in sensitivity to estrogens. With this dose of estradiol, progesterone was able to induce a preovulatory-type gonadotropin surge even in the male rat (149). That progesterone played a pivotal role in ovulation was further demonstrated by the use of the progesterone and glucocorticoid antagonist RU-486 (17-hydroxy-11-[4-dimethylaminopgenyl]17␣-[prop-1-ynyl]-estra-4,9-diene-3-one) (193). RU-486 abolished the preovulatory gonadotropin surge in PMSG-treated immature rat and in the normally cycling adult rat. In this regard, it is of interest to note that Goldman et al. (83) found an elevation of progesterone in ovarian vein blood at 1400 h on proestrus prior to the LH surge. Progesterone and LH levels also showed an increase at 1400 h in the study of Nequin et al. (165), although several other investigators found no rise in preovulatory progesterone, possibly because they did not sample blood at 1400 h. A preovulatory increase in serum progesterone has been found in women 12 h before the initiation of the rise of LH (101) and 6 h before the rise of LH in rhesus monkeys (204). In the estradiol-primed immature rat, deoxycorticosterone (21-hydroxy-4-pregnene-3,20-dione) and the synthetic glucocorticoid triamcinilone acetonide were also able to induce a preovulatory type of gonadotropin surge, whereas cortisol was unable to do so (35, 36). The ability of deoxycorticosterone and triamcinilone acetonide to induce the surge was considered to be due to their interaction with the progesterone receptor. These steroids, like progesterone, were also very effective in stimulating fluid loss from the distended uterus caused by estrogen treatment. Low doses of the synthetic steroid dexamethasone were able to selectively bring about FSH release in the estrogen-primed rat (36). Deoxycorticosterone and triamcinilone acetonide were also able to induce ovulation in PMSG-primed rats, similar to progesterone, which served as a biological test for their effect (35). In the estrogen-primed ovariectomized rat, acute administration of ACTH also resulted in the preovulatory type of gonadotropin release. The surge required the presence of the adrenal glands and was antagonized by the progesterone and glucocorticoid antagonist RU-486. ACTH increased both progesterone and deoxycorticosterone, and the gonadotropin surge was considered to be mediated by these steroids (190). Mechanism of action of progesterone on the release of gonadotropins. ANTAGONISM OF ESTROGEN ACTION ON THE PITUITARY. An examination of the effects of progesterone on the estrogen receptor of the anterior pituitary in ovariectomized estrogen-primed rats showed that progesterone brought about a decrease in the estrogen receptors of the anterior pituitary (210). This created a controversy in the literature, as several other investigators had failed to find an effect of progesterone on estrogen receptors of the anterior pituitary. A detailed examination of the progesterone effect showed that it occurred only during the period of the nuclear occupancy of the progesterone receptor and not before or after the event (57). Furthermore, only during the period of nuclear occupancy of the progesterone receptor in the pituitary did progesterone attenuate the effect of a second injection of estradiol on the increase in cytosolic progesterone receptors. The effect of progesterone on estrogen receptors was also confirmed in in vitro experiments and in the adult rats (75, 209). E9 Perspectives E10 STEROID-GONADOTROPIN-FEEDBACK SYSTEM: CAREER RETROSPECTIVE secretion than to FSH secretion. The direct effect of neuropeptide Y on anterior pituitary cells in culture in sensitizing the pituitary to LHRH has also been demonstrated (168). It was also found that progesterone modulates neuropeptide Y levels in the anterior pituitary during the progesterone-induced surge in the estrogen-primed rat (169). Changes in galanin mRNA also correlate to the progesterone-induced gonadotropin surge (8). The above-mentioned experiments clearly indicate that progesterone in estrogen-primed rats can bring about LHRH secretion, which is responsible for the preovulatory surge of gonadotropins. REGULATION OF PEPTIDASE ACTIVITY INVOLVED IN THE DEGRADATION OF LHRH IN THE HYPOTHALAMUS AND THE PITUITARY. The REGULATION OF LH- MRNA AND FSH- MRNA LEVELS BY PROGESTERONE DURING THE PREOVULATORY GONADOTROPIN SURGE. Initial experiments did not show any consistent effect of progesterone on the pituitary LH- mRNA and FSH- mRNA levels in the ovariectomized estrogen-primed rat treated with progesterone, whereas dexamethasone increased the level of FSH- mRNA before the rise of serum FSH (27). This may be due to the fact that the gonadotropin subunit levels in ovariectomized estrogen-primed rats were severalfold higher than those in intact estrogen-primed rats. In the estrogen-primed intact rat, progesterone administration brought about an elevation of LH- mRNA and FSH- mRNA in parallel with the preovulatory gonadotropin surge (28). This also occurred in the PMSG-primed immature rat. The changes in the mRNA levels were blocked by the antiprogestin RU-486 (28). Further work showed that a 361 base pair region of the FSH promoter gene contained several progesterone response elements and these mediated the progesterone effect on the FSH gene (170, 171). SELECTIVE RELEASE OF LH AND FSH BY PROGESTERONE METABOLITES. In the estrogen-primed immature ovariecto- mized rat, the administration of the progesterone metabolite 5␣-dihydroprogesterone brought about the selective release of FSH (155) and the progesterone metabolite 3␣,5␣-tetrahydroprogesterone brought about a selective release of LH (156). The selective release of LH and FSH by the above-mentioned progesterone metabolites also took place in the PMSG-treated immature rats exposed to constant light (157). Initially, the effect of 3␣,5␣-tetrahydroprogesterone was unexpected, as the Fig. 2. Progesterone is converted into two major metabolites, 5␣-dihydroprogesterone and 3␣,5␣-tetrahydroprogesterone. The former uses the progesterone receptor and brings about selective release of FSH; the latter uses the GABAA receptor and brings about selective release of LH. Administration of a 5␣-reductase inhibitor in vivo or in vitro reduced FSH release without affecting LH release. Stimulatory pathways are shown in green; inhibitory pathways in red. AJP-Endocrinol Metab • doi:10.1152/ajpendo.00488.2011 • www.ajpendo.org Downloaded from http://ajpendo.physiology.org/ by 10.220.33.3 on June 18, 2017 biological activity of LHRH in the release of LH and FSH is also dependent upon the amount of LHRH that survives degradation by the peptidase activity in the hypothalamus and the pituitary. The peptidase activity of the hypothalamus and the pituitary was lower in the hypothalamus and the pituitary on proestrus and diestrus 1 compared with other parts of the ovulatory cycle, thus leaving the maximum amount of LHRH available during that time for biological action (166). To examine the role of progesterone on changes in the peptidase activity in the hypothalamus, pituitary and serum, the activity was measured in estrogen-primed ovariectomized rats before and after progesterone administration. Estrogen administration increased the peptidase activity of the hypothalamus, which was antagonized by progesterone (167). Progesterone similarly decreased the peptidase activity of serum but had no effect on the peptidase activity of the pituitary. These results indicate that progesterone administration brings about a suppression of hypothalamic peptidase activity, permitting the availability more LHRH for mounting the preovulatory gonadotropin surge. compound does not interact with the progesterone receptor. However, several steroidal anesthetics that have a 3␣,5␣reduced Ring A structure had been shown to use the ␥-amino butyric acid A (GABAA) receptor system for their biological action. Further work showed that the action of 3␣,5␣-tetrahydroprogesterone on LH release could be blocked by GABAA receptor antagonists but not by RU-486 (33). Progesterone metabolites were also able to dampen the estrogen-induced uterine contractions. Once again, similar to the FSH and LH release, the dampening effect of 5␣-dihydroprogesterone was blocked by the progesterone receptor antagonist RU-486, whereas the 3␣,5␣-tetrahydroprogesterone effect was blocked by the GABAA antagonist (189). That the effect of progesterone in the release of FSH was mediated by its 5␣-reduction to 5␣-dihydroprogesterone was demonstrated by the use of a 5␣-reductase inhibitor N,N-diethyl-4-methyl-3oxo-4-aza-5␣androstane-17-carboxamide. The use of this inhibitor blocked the reduction of progesterone in the 5␣ position and the progesterone induced a FSH surge without affecting the progesterone-induced LH surge (Fig. 2) (191). 5␣-Dihydroprogesterone also brought about the depletion of occupied estrogen receptors of the anterior pituitary and the uterus similar to progesterone (77). A significant amount of the stimulatory and inhibitory effects of LHRH on gonadotropin secretion by progesterone occurred at the level of the pituitary, as shown in pituitary cell cultures (112). In estrogen-primed pituitary cells, progesterone exposure for 1– 6 h brought about the potentiation of LHRH in the release of LH and FSH, whereas progesterone exposure of 12 h or more was inhibitory. The 5␣-reductase inhibitor blocked the effect of progesterone on FSH release. It was also of interest to note that in the estrogen-primed rat the administration of the opioid antagonist naloxone brought about an increase in LH secretion. An inhibitor of GABA degradation aminooxyacetic acid and both GABAA and GABAB agonists blocked this action (44). In this regard, a decreased opioid response to naloxone was observed in patients with premenstrual tension (196). 3␣,5␣-reduced steroids have an anesthet- Perspectives STEROID-GONADOTROPIN-FEEDBACK SYSTEM: CAREER RETROSPECTIVE ic- and anxiety-reducing effect. In women with premenstrual tension, the metabolism of progesterone to 3␣,5␣-tetrahydroprogesterone was compromised (195). EXCITATORY AMINO ACIDS AS MEDIATORS OF THE STEROID-GONADOTROPIN FEED BACK SYSTEM Estabilishing the Possible Role of Excitatory Amino Acids in Gonadotropin Secretion rat showed a decline in pituitary NMDA mRNA. However, progesterone treatment to the estrogen-primed rat resulted in an increase in pituitary NMDA mRNA, which is in keeping with the direct effects of progesterone on the pituitary during the gonadotropin surge mentioned earlier in this article (9). During the rat ovulatory cycle, NMDA receptor 1 (NMDAR1) and glutamate receptor 1 (GluR1) did not change much, but the levels of the kainite receptor GluR6 decreased just preceding the surge. This decrease was demonstrated to be due to progesterone action (42) Administration of NMDA advanced puberty in the rat by 2.5 days, whereas the non-NMDA agonist kainite or the nonNMDA antagonist had no effect on puberty, indicating that NMDA was primarily involved in the process of puberty (43). The NMDA receptors did not change significantly in the pituitary during puberty, whereas the AMPA receptors showed an increase (219). This indicated selective excitatory amino acid receptor involvement during puberty. Determining Whether Glutamate Was Actually Secreted During the Gonadotroipn Surge The next important question was to determine whether the role of glutamate, which was indicated to be important in the gonadotropin surge by the study of various agonist and antagonist studies of various receptors of glutamate action, could be established by the actual secretion of glutamate during the ovulatory surge of gonadotropins. To achieve this, microdialysis was done in vivo from the preoptic area through a cannula implanted in the estrogen-primed rat given progesterone to induce ovulation. There was a significant increase in the preoptic area release of glutamate and aspartate immediately preceding the preovulatory gonadotropin surge (187). Naloxone has been shown to bring about LH release by inhibiting the opioid inhibitory mechanism in several studies. In male rats, administration of naloxone brought about an increase in serum LH and an increase in nitric oxide synthase (NOS) in the preoptic area and medial basal hypothalamus within 20 min (5). This increase was blocked by the NMDA antagonist MK801. Microdialysis experiments in vivo of the preoptic area also showed a significant increase in glutamate secretion within 15 min of naloxone administration. Thus, the opioid block of gonadotropin secretion appeared by their suppression of glutamate release. Localization of NMDAR1 Receptors in the Hypothalamus and the Role of NOSs Physiochemical localization of NMDAR1-containing neurons showed extensive distribution in the hypothalamus, including the organum vasculosum of the lamina terminalis (OVLT), median preoptic nucleus, and median preoptic area (4). This NMDAR1 staining area did not colocalize with the GnRH neuron but surrounded several of them; they colocalized with NOS-1 activity. Central administration of a NOS inhibitor abolished the steroid-induced preovulatory surge, indicating that NMDA acted through NO in the release of LHRH. NMDAR1 was also found in almost all cell types of the anterior pituitary, suggesting some pituitary action of NMDA (6). Further work showed that the hypothalamus contained all three isoforms of NOS, namely brain NOS (NOS-1), microphage NOS (NOS-2), and endothelial NOS (NOS-3). Of AJP-Endocrinol Metab • doi:10.1152/ajpendo.00488.2011 • www.ajpendo.org Downloaded from http://ajpendo.physiology.org/ by 10.220.33.3 on June 18, 2017 Early work had suggested that glutamate may have a role in the release of LH (175). To determine whether excitatory amino acids mediated the steroid-induced gonadotropin surge, estrogenprimed immature rats treated with progesterone or triamcinilone acetonide were injected with the N-methyl-D-aspartate (NMDA) antagonist MK801 [(⫹)-5-methyl-10,11-dihydro-5H-dibenzo(a,d) cyclohepten-5,10-imine maleate] (18). MK801 blocked the progesterone- or triamcinilone acetonide-induced LH and FSH surge. MK801 also reduced the number of ova ovulated in the PMSG-treated rat along with a decrease in the LH and FSH surge (19). MK801 also decreased serum LH levels in the adult cycling rat. The excitatory amino acid agonist NMDA given to estrogen-primed rats brought about prompt elevations of LH and FSH. The effect seemed to be at the hypothalamus, because medial basal hypothalamus/preoptic area fragments showed an elevated release of LHRH in vitro 5 and 7 min after the administration of NMDA (19 –21). These findings are of considerable importance, as it was shown that the GnRH neuron did not appear to have steroid receptors whereas there were steroid receptors in glutamate-containing neurons (100, 111). Thus, the GnRH neuron was regulated by other neurons in the hypothalamus that had steroid receptors. The subject has been reviewed extensively (22–24). To determine whether non-NMDA neurotransmission also regulated the gonadotropin surge, the non-NMDA receptor antagonist DNQX (6,7dinitroquinoxaline-2,3-dione) was administered via a third ventricular cannula in estrogen-primed adult rats treated with progesterone and PMSG-primed immature rats. DNQX attenuated the LH and the prolactin surges without much effect on the FSH surge (29). Both NMDA and non-NMDA receptors played a role in pulsatile LH release, as shown by injection of the specific NMDA receptor antagonist AP5 (2-amino-5-phosphono-pentanoic acid) and the non-NMDA antagonist DNQX in adult rats ovariectomized for 2 wk via a third ventricular cannula (185). Similar results were obtained in the male rat as well, although the NMDA receptor antagonist was more effective than the non-NMDA receptor antagonist (186). AMPA (␣-amino-3-hydroxy-5-methylisooxazole-4-propionoc acid) also brought about LH release in the estrogen-primed rat, indicating a role for AMPA receptors. The steroid-induced gonadotropin surge was blocked by the selective AMPA receptor antagonist NBQX (184). Since NMDA was more effective in gonadotropin release in the estrogen-primed female rat compared with the ovariectomized rat, the NMDA receptor in the hypothalamus was characterized (141), and the NMDA receptor binding and NMDA receptor mRNA weremeasured in the hypothalamus upon castration and after estrogen priming (41). No change was observed in castrated female rats after estrogen treatment or in castrated male rats with testosterone treatment. The effect of steroids on NMDA receptor mRNA levels in the pituitary was also examined. Estrogen treatment of the ovariectomized E11 Perspectives E12 STEROID-GONADOTROPIN-FEEDBACK SYSTEM: CAREER RETROSPECTIVE these, NOS-1 was the major isoform and acted through the activation of cGMP (3). The role of NOS-1 in the release of steroid-induced gonadotropin surge was further confirmed by the administration of antisense nucleotides to NOS-1, which attenuated the steroid-induced gonadotropin surge (1). NOS levels were also shown to increase during the proestrus gonadotropin surge in the rat (108). A regulatory role of carbon monoxide on LH release has also been suggested (109). The role of gaseous neurotransmitters in the release of gonadotropins has been reviewed (7). Mechanism of Action of Progesterone in Releasing Glutamate and Suppressing GABA ROLE OF HYPOTHALAMIC ASTROCYTES IN STEROID-GONADOTROPIN FEEDBACK To determine whether astrocytes played any role in stimulating hypothalamic LHRH secretion, conditioned medium from hypothalamic astrocytes was incubated with either hypothalamic fragments or the immortalized GnRH neuronal cells the GT1-7 cells (49), The hypothalamic conditioned medium brought about the release of LHRH. Ultra filtration of the astrocytes conditioned medium to remove peptides greater than 10 kDa resulted in loss of activity. The peptide responsible for the release of LHRH was found to be transforming growth factor- (TGF). Antibodies to TGF abolished the ability of OTHER RELATED STUDIES This author has been involved in a number of related studies, a detailed description of which is outside the scope of this article. They are studies of steroid gonadotropin feedback in aging (25, 42, 104, 147, 225), testicular feminization (154), gonadal dysgenesis (90, 91, 143), metabolism of oral contraceptives (47, 48, 151, 152, 153), heterogeneity in granulosa cells (192, 194), regulation of follicular development by diethyl stilbesterol (59, 60), estrogen and progesterone receptors in the human endometrium (158, 159), and pregnancy in the rat (172–174). Other studies refer to the role of kinins in the ovulatory cycle and pregnancy and the possible role of bradykinin in the preovulatory gonadotropin surge (12, 81, 205, 206). The role of leptin in reproduction is also of considerable interest (10, 40, 50, 58, 218, 220). Finally, computer modeling of partially unwound DNA structures caused by receptor binding shows a stereochemical fit of estradiol in the cavity in the center of the hormone response element. The fit in the DNA cavity is a much more accurate prediction of biological activity than estrogen binding to its receptor. The technique has led to the development of search engines that can be used in drug design (46, 96 –99, 117, 118, 136, 137) SUMMARY AND CONCLUSIONS Fig. 3. Progesterone inhibition of glutamic acid dehydrogenase-67 (GAD67) permits more glutamate release and less GABA release, thus permitting an LH surge. Stimulatory events are shown in green, inhibitory events in red. Early work on polycystic ovary syndrome was a major breakthrough, as it established the ovary with or without the involvement of the adrenal as a source of androgens. The reasons for the occurrence of hirsutism and virilism in a variety of endocrine patients was also established. Experimental models for creating polycystic ovaries as well as other experiments established the detrimental effects of excessive androgens on follicular development that resulted in cystic ovaries. The studies also resulted in establishing the use of Clomiphene for the induction of ovulation in anovulatory women. Animal studies showing the ability of dexamethasone in releasing FSH resulted in the use of dexamethasone in combination with Clomiphene to induce ovulation in women not shown to have excessive androgen secretion but who had failed to ovulate on treatment with Clomiphene alone. A link between premen- AJP-Endocrinol Metab • doi:10.1152/ajpendo.00488.2011 • www.ajpendo.org Downloaded from http://ajpendo.physiology.org/ by 10.220.33.3 on June 18, 2017 The next question was how progesterone brought about the release of glutamate during the preovulatory gonadotropin surge. This was shown to be due to the suppression of glutamic acid decarboxylase-67 (GAD67) by progesterone in the estrogen-primed rat, resulting in the decrease of glutamate converted to GABA, thus permitting more glutamate and less GABA to be secreted (Fig. 3) (215). The suppression of GAD67 during the preovulatory surge of gonadotropins was also shown to occur in the adult rat (39). In addition, the synaptic terminals’ appositions to the GnRH neuron of the vesicular GABA transporter (VGAT) was decreased while the vesicular glutamate transporter 2 (VGLUT2) was increased during the proestrus gonadotropin surge, thus further confirming the role of glutamate in the surge (104). the conditioned medium to release LHRH. Furthermore, the hypothalamus and the GT1-7 cells possessed TGF receptors. Estrogens stimulated astrocytes to release TGF, and this action was attenuated with estrogen receptor antagonists. Astrocytes also possessed estrogen receptors. Astrocytes’ conditioned medium also protected GT1-7 cell cultured in serumfree medium from death, and this was attributed to TGF. The neuroprotective pathway appeared to be via the activation of the c-Jun/AP-1 pathway (66). Similar to estradiol, tamoxifen was also able to reduce the extent of stroke in experimental animals in which the medial carotid artery was blocked (150). The neuroprotective effects of estrogens were exerted in part by the stimulation of TGF secretion using a nongenomic mechanism (68). This was another example of the nongenomic effect of steroids (13). The role of astrocytes in reproduction and neuroprotection has also been reviewed (116). Extensive work on the mechanisms involved in estrogen effects on neuroprotection has continued but is outside the scope of this article but is described in detail elsewhere (11, 45, 67, 69, 105, 216, 222). Perspectives STEROID-GONADOTROPIN-FEEDBACK SYSTEM: CAREER RETROSPECTIVE ACKNOWLEDGMENTS I wish to express my gratitude to numerous graduate students, postdoctoral fellows, and faculty colleagues for their untiring collaboration and help in introducing new techniques in the laboratory from time to time. GRANTS This research was supported by Grants AM-04429, HD-04626, HD-10795, HD-16688, HD-14196 and HD-16396 from the National Institutes of Health. DISCLOSURES No conflicts of interest, financial or otherwise, are declared by the author(s). AUTHOR CONTRIBUTIONS Author contributions: V.B.M. conception and design of research; V.B.M. performed experiments; V.B.M. analyzed data; V.B.M. interpreted results of experiments; V.B.M. drafted manuscript; V.B.M. edited and revised manuscript; V.B.M. approved final version of manuscript. REFERENCES 1. Aguan K, Mahesh VB, Ping L, Bhat G, Brann DW. Evidence for a physiological role for nitric oxide in the regulation of the LH surge: effect of central administration of antisense oligonucleotides to nitric oxide synthase. Neuroendocrinology 64: 449 –455, 1996. 2. Bagnell CA, Mills TM, Costoff A, Mahesh VB. A model for the study of androgen effects on follicular atresia and ovulation. Biol Reprod 27: 903–914, 1982. 3. Bhat G, Mahesh VB, Aguan K, Brann DW. Evidence that brain-nitric oxide synthase is the major nitric oxide synthase isoform in the hypothalamus of the adult female rat and that nitric oxide potently regulates hypothalamic cGMP levels. Neuroendocrinology 64: 93–102, 1996. 4. Bhat GK, Mahesh VB, Lamar CA, Ping L, Aguan K, Brann DW. Histochemical localization of nitric oxide neurons in the hypothalamus: association with gonadotropin-releasing hormone neurons and co-localization with N-methyl-D-aspartate receptors. Neuroendocrinology 62: 187–197, 1995. 5. Bhat GK, Mahesh VB, Ping L, Chorich LP, Wiedmeier V, Brann DW. Opioid-glutamate-nitric oxide connection in the regulation of luteinizing hormone secretion in the rat. Endocrinology 139: 955–960, 1998. 6. Bhat GK, Mahesh VB, Zhen WC, Chorich LP, Zamorano PL, Brann DW. Localization of the N-methyl-D-aspartate R1 receptor subunit in specific anterior pituitary hormone cell types of the female rat. Neuroendocrinology 62: 178 –186, 1995. 7. Brann DW, Bhat GK, Lamar CA, Mahesh VB. Gaseous transmitters and neuroendocrine regulation. Neuroendocrinology 65: 385–395, 1997. 8. Brann DW, Chorich LP, Mahesh VB. Effect of progesterone on galanin mRNA levels in the hypothalamus and the pituitary: correlation with the gonadotropin surge. Neuroendocrinology 58: 531–538, 1993. 9. Brann DW, Chorich LP, Zamorano PL, Mahesh VB. Presence of NMDA receptor mRNA in the anterior pituitary of the female rat: steroid modulation and changes during gonadotropin surge induction. Mol Cell Neurosci 4: 571–575, 1993. 10. Brann DW, De Sevilla L, Zamorano PL, Mahesh VB. Regulation of leptin gene expression and secretion by steroid hormones. Steroids 64: 659 –663, 1999. 11. Brann DW, Dhandapani K, Wakade C, Mahesh VB, Khan MM. Neurotrophic and neuroprotective actions of estrogen: basic mechanisms and clinical implications. Steroids 72: 381–405, 2007. 12. Brann DW, Greenbaum L, Mahesh Gao X VB. Changes in kininogens and kallikrein in the plasma, brain and uterus during pregnancy in the rat. Endocrinology 136: 46 –51, 1995. 13. Brann DW, Hendry LB, Mahesh VB. Emerging diversities in the mechanism of action of steroid hormones. J Steroid Biochem Mol Biol 52: 113–133, 1995. 14. Brann DW, Mahesh VB. The role of corticosteroids in reproduction. FASEB J 5: 2691–2698, 1991. 15. Brann DW, Mahesh VB. Detailed examination of the mechanism and site of action of progesterone and corticosteroids in the regulation of gonadotropin secretion: hypothalamic gonadotropin releasing hormone and catecolamine involvement. Biol Reprod 44: 1005–1015, 1991. 16. Brann DW, Mahesh VB. Validation of the mechanisms proposed for the stimulatory and inhibitory effects of progesterone on gonadotropin secretion in the estrogen-primed rat. A possible role for adrenal steroids. Steroids 56: 103–111, 1991. 17. Brann DW, Mahesh VB. Regulation of gonadotropin secretion by steroid hormones. Front Neuroendocrinol 12: 165–207, 1991. AJP-Endocrinol Metab • doi:10.1152/ajpendo.00488.2011 • www.ajpendo.org Downloaded from http://ajpendo.physiology.org/ by 10.220.33.3 on June 18, 2017 strual tension and abnormal metabolism of progesterone was also established. The above are excellent examples of translational applications of basic science research. There is no clear understanding of the importance of the FSH surge at the time of ovulation in the human. The finding that FSH can bring about the rupture of the mature ovarian follicle and that several women with short luteal phase have a decreased or displaced FSH surge is intriguing, and further work needs to be done in this area. The studies described in this article also provide the sequence of events starting with follicular growth and an increase in estrogens resulting in the occurrence of puberty in the female and similar effects with androgens in the male rat. Although the studies provide a solid foundation of the events that take place during puberty, the mechanisms that trigger the initial increase in follicular growth and estrogen secretion in female rats and androgen secretion in male rats are still obscure and require further investigation. The role of estradiol in triggering the preovulatory surge of gonadotropins has been well recognized in the literature. The studies described in this article clearly show the pivotal role of progesterone in the process of modulating the preovulatory surge of gonadotropins. The dependence of progesterone action on estrogens is due to the synthesis of progesterone receptors by estrogen action. Progesterone acts by stimulating hypothalamic release of LHRH, suppressing the peptidase activity and thus making more LHRH available, increasing the pituitary sensitivity to LHRH along with synthesis of new LH and FSH, and decreasing the occupied nuclear estrogen receptors of the pituitary, thus overriding the suppressive effects of estrogens on the pituitary in gonadotropin release. The differential regulation of FSH and LH has also been a subject of considerable interest. The demonstration of selective effects of progesterone metabolites on FSH and LH secretion and the use of not only the progesterone receptor but the GABAA receptor as well in this process is therefore of considerable importance. The work described in this article has resulted in the replacement of the long-held belief, that gonadal steroids act directly on the GnRH neuron to regulate gonadotropin secretion, with the novel concept that estrogens and progesterone act on the glutamatergic neurons of the hypothalamus that contain steroid receptors and that these neurons in turn regulate the GnRH neuron via NOS-1. Thus, this new pathway adds to the multiple pathways known or currently under discovery of the regulation of reproduction, which is essential for the preservation and continuation of the species. Finally, the concept of the neuroprotective effect of estrogens on brain function is of considerable importance. This neuroprotective effect is lost during years of estrogen deprivation during menopause, leading to increased incidence of stroke and dementia. Studies on the mechanism of loss of the neuroprotective effect of estrogens during estrogen deprivation and methods to circumvent it would be of considerable value for the health and well-being of our aging population. E13 Perspectives E14 STEROID-GONADOTROPIN-FEEDBACK SYSTEM: CAREER RETROSPECTIVE 42. 43. 44. 45. 46. 47. 48. 49. 50. 51. 52. 53. 54. 55. 56. 57. 58. 59. 60. 61. 62. 63. 64. 65. the hypothalamus and cerebral cortex of the adult rat. Neuroendocrinology 58: 666 –672, 1993. Brann DW, Zamorano PL, DeSevilla L, Mahesh VB. Expression of glutamate receptor subunits in the hypothalamus of the female rat during the afternoon of the proestrous LH surge and effects of antiprogestin treatment and aging. Neuroendocrinology 81: 120 –128, 2005. Brann DW, Zamorano PL, Ping L, Mahesh VB. Role of excitatory amino acid neurotransmission during puberty in the female rat. Mol Cell Neurosci 4: 107–112, 1993. Brann DW, Zamorano PL, Putnam-Roberts CD, Mahesh VB. Gamma aminobutyric acid-opioid interactions in the regulation of gonadotropin secretion in the immature female rat. Neuroendocrinology 56: 445–452, 1992. Brann DW, Zhang QG, Wang RM, Mahesh VB, Vadlamudi RK. PELP1—a novel estrogen receptor-interacting protein in the brain. Mol Cell Endocrinol 290: 2–7, 2008. Bransome ED Jr, Hendry LB, Muldoon TG, Mahesh VB, Hutson MS, Campbell LK. Apparent stereochemical complementarity of estrogens and helical cavities between DNA base pairs: implications for the mechanism of action of steroids. J Theoret Biol 112: 97–108, 1985. Braselton WE, Lin TJ, Mills TM, Ellegood JO, Mahesh VB. Identification and measurement by gas chromatography-mass spectrometry of Norethindrone and metabolites in human urine and blood. J Steroid Biochem Mol Biol 8: 9 –18, 1977. Braselton WE Jr, Lin TJ, Ellegood JO, Mills TM, Mahesh VB. Accumulation of Norethindrone and idividual metabolites in human plasma during short- and long-term administration of a contraceptive dosage. Am J Obstet Gynecol 133: 154 –160, 1979. Buchanan CD, Mahesh VB, Brann DW. Estrogen-astrocyte-luteinizing hormone releasing hormone signaling : a role for transforming growth factor-beta-1. Biol Reprod 62: 1710 –1721, 2000. Buchanan CL, Mahesh VB, Zamorano PL, Brann DW. Central nervous system effects of leptin. Trends Endocrinol Metab 9: 146 –150, 1998. Bush IE. The 11-oxygen function in steroid metabolism. Experientia 12: 325–331, 1956. Bush IE, Mahesh VB. Metabolism of 11-oxygenated steroids. 1. Influence of the A/B ring junction on the reduction of 11-oxo groups. Biochem J 71: 705–717, 1959. Bush IE, Mahesh VB. Metabolism of 11-oxygenated steroids. 2. 2-Methyl steroids. Biochem J 71: 718 –742, 1959. Bush IE, Mahesh VB. Adrenocortical hyperfunction with sudden onset of hirsutism. J Endocrinol 18: 1–25, 1959. Bush IE, Mahesh VB. Metabolism of 11-oxygenated steroids. 3. Some 1-dehydro and 9alpha-fluoro steroids. Biochem J 93: 236 –255, 1964. Byrd JR, Mahesh VB, Greenblatt RB. Chromosomal studies in the Stein-Leventhal Syndrome. J Clin Endocrinol Metab 24: 939 –940, 1964. Calderon JJ, Muldoon TG, Mahesh VB. Receptor mediated interrelationship between progesterone and estradiol action on the anterior pituitary-hypothalamic axis of the ovariectomized Immature rat. Endocrinology 120: 2428 –2435, 1987. Cannady WE, Brann DW, Mahesh VB. The potential role of periovarian fat and leptin the initiation of puberty. Int J Obes Relat Metab Dis Suppl 12: S146 –S147, 2000. Chakravorty A, Mahesh VB, Mills TM. Regulation of follicular development by diethylstilboestrol in the immature rat ovary. J Reprod Fertil 92: 307–321, 1991. Chakravorty A, Mahesh VB, Mills TM. Inhibition of proliferative potential of specific-sized follicles in the rat ovary by diethylstilboestrol. J Reprod Fertil 92: 323–332, 1991. Chorich L, Rosser M, Howard E, Zamorano P, Mahesh VB. Rat anterior pituitary estrogen receptor mRNA levels after estrogen and progesterone treatment. Mol Cell Neurosci 3: 487–496, 1992. Conway BA, Mahesh VB, Mills TM. Effect of dihydrotestosterone on the growth and function of ovarian follicles in intact immature female rats primed with PMSG. J Reprod Fertil 90: 267–277, 1990. Costoff A, Eldridge JC, Mahesh VB. Pituitary ultrastructure and serum gonadotropin levels in the PMS-primed immature rat. Cell Tissue Res 151: 79 –92, 1974. Costoff A, Knudsen JF, Mahesh VB. Correlation of serum gonadotropins, ovarian and uterine histology in immature and prepubertal rats. Anat Rec 180: 497–507, 1974. Dalla Pria S, Greenblatt RB, Mahesh VB. An antiandrogen in acne and idiopathic hirsutism. Invest Dermatol 52: 348 –350, 1969. AJP-Endocrinol Metab • doi:10.1152/ajpendo.00488.2011 • www.ajpendo.org Downloaded from http://ajpendo.physiology.org/ by 10.220.33.3 on June 18, 2017 18. Brann DW, Mahesh VB. Endogenous excitatory amino acid regulation of the progesterone-induced LH and FSH surge in estrogen-primed ovariectomized rats. Neuroendocrinology 53: 107–110, 1991. 19. Brann DW, Mahesh VB. Endogenous excitatory amino acid involvement in the preovulatory and steroid-induced surge of gonadotropins in the female rat. Endocrinology 128: 1541–1547, 1991. 20. Brann DW, Mahesh VB. Excitatory amino acid regulation of gonadotropin secretion: modulation by steroid hormones. J Steroid Biochem Mol Biol 41: 847–850, 1992. 21. Brann DW, Mahesh VB. Excitatory amino acid neurotransmission: evidence for a role in neuroendocrine regulation. Trends Endocrinol Metab 3: 120 –124, 1992. 22. Brann DW, Mahesh VB. Excitatory amino acids: function and significance in reproduction and neuroendocrine regulation. Front Neuroendocrinol 15: 3–49, 1994. 23. Brann DW, Mahesh VB. Glutamate: a major neuroendocrine excitatory signal mediating steroid effects on gonadotropin secretion. J Steroid Biochem Mol Biol 53: 325–329, 1995. 24. Brann DW, Mahesh VB. Excitatory amino acids: evidence for a role in the control of reproduction and anterior pituitary hormone secretion. Endoc Rev 18: 678 –700, 1997. 25. Brann DW, Mahesh VB. The aging reproductive neuroendocrine axis. Steroids 70: 273–283, 2005. 26. Brann DW, McDonald JK, Putnam CD, Mahesh VB. Regulation of hypothalamic gonadotropin-releasing hormone and neuropeptide Y concentrations by progesterone and corticosteroids in immature rats: correlation with Luteinizing hormone and follicle-stimulating hormone release. Neuroendocrinology 54: 425–432, 1991. 27. Brann DW, O’Conner JL, Wade M, Mahesh VB. LH and FSH subunit mRNA concentrations during the progesterone-induced gonadotropin surge in ovariectomized estrogen-primed immature rats. Mol Cell Neurosci 3: 171–178, 1992. 28. Brann DW, O’Conner JL, Wade MF, Zamorano PL, Mahesh VB. Regulation of anterior pituitary gonadotropin subunit mRNA levels during the preovulatory gonadotropin surge: a physiological role of progesterone in regulating LH- and FSH- mRNA levels. J Steroid Biochem Mol Biol 46: 427–437, 1993. 29. Brann DW, Ping L, Mahesh VB. Possible role of non-NMDA receptormediated neurotransmission in steroid-induced and preovulatory gonadotropin surges in the rat. Mol Cell Neurosci 4: 292–297, 1993. 30. Brann DW, Putnam CD, Mahesh VB. Antagonism of estrogen-induced prolactin release by dihydrotesterone. Biol Reprod 40: 1201–1208, 1989. 31. Brann DW, Putnam CD, Mahesh VB. Dose-related effects of progesterone and 5alpha-dihydroprogesterone upon estrogen-induced prolactin release. J Neuroendocrinol 2: 341–345, 1990. 32. Brann DW, Putnam CD, Mahesh VB. Similarities and differences in progesterone and androgens in modulation of LH, FSH and PRL release: unexpected properties of flutamide. J Steroid Biochem Mol Biol 36: 287–294, 1990. 33. Brann DW, Putnam CD, Mahesh VB. GABAA receptor mediate 3␣-hydroxy-5␣-pregnan-20-one induced gonadotropin secretion. Endocrinology 126: 1854 –1859, 1990. 34. Brann DW, Putnam CD, Mahesh VB. Dose-related effects of progesterone and 5alpha-dihydroprogesterone upon estrogen-induced prolactin release. J Neuroendocrinol 2: 341–345, 1990. 35. Brann DW, Putnam CD, Mahesh VB. Corticosteroid regulation of gonadotropin secretion and induction of ovulation in the rat. Proc Soc Expt Biol Med 193: 176 –180, 1990. 36. Brann DW, Putnam CD, Mahesh VB. Corticosteroid regulation of gonadotropin and prolactin secretion in the rat. Endocrinology 126: 159 –166, 1990. 37. Brann DW, Putnam-Roberts CD, Mahesh VB. Progesterone and corticosteroid regulation of hypothalamic and pituitary opioid content during LH surge induction. Mol Cell Neurosci 3: 191–198, 1992. 38. Brann DW, Rao IM, Mahesh VB. Antagonism of estrogen-induced prolactin release by progesterone. Biol Reprod 39: 1067–1073, 1988. 39. Brann DW, Unda R, Mahesh VB. Role of progesterone receptor in restrained glutamic acid decarboxylase gene expression in the hypothalamus during the preovultory luteinizing hormone surge. Neuroendocrinology 76: 283–289, 2002. 40. Brann DW, Wade MF, Dhandapani KM, Mahesh VB, Buchanan CD. Leptin and reproduction. Steroids 67: 95–104, 2002. 41. Brann DW, Zamorano PL, Chorich LP, Mahesh VB. Steroid hormone effects on NMDA receptor binding and NMDA receptor mRNA levels in Perspectives STEROID-GONADOTROPIN-FEEDBACK SYSTEM: CAREER RETROSPECTIVE 90. 91. 92. 93. 94. 95. 96. 97. 98. 99. 100. 101. 102. 103. 104. 105. 106. 107. 108. 109. 110. 111. 112. trogen receptor concentration in the anterior pituitary and hypothalmus of the cycling rat. Biol Reprod 13: 505–512, 1975. Greenblatt RB, Byrd JR, McDonough PG, Mahesh VB. The spectrum of gonadal dysgenesis: a clinical, cytogenetic and pathologic study. Am J Obstet Gynecol 98: 151–172, 1967. Greenblatt RB, Dominquez H, Mahesh VB, Demos RJ. Gondal dysgenesis intersex with X0/XY mosaicism. J Am Med Assoc 188: 221–224, 1964. Greenblatt RB, Faucher G, Mahesh VB, Lunenfeld B, Rabau E, David A, Insler V. Ovulation and pregnancy in the Chiari-Frommel syndrome: report of 10 cases. Fertil Steril 17: 742–756, 1966. Greenblatt RB, Mahesh VB, Gambrell RD. Arrhenoblastoma—three case reports. Obstet Gynecol 39: 567–576, 1972. Greenblatt RB, Roy S, Mahesh VB, Barfield WE, Jungck EC. Induction of ovulation. Am J Obstet Gynecol 84: 900 –912, 1962. Greenblatt RB, Zarate A, Mahesh VB. Methods available for the induction of ovulation. Proc VI Pan American Cong Endocrinol Excerpta Med Int Congr Ser 112: 129 –133, 1965. Hendry LB, Bransome ED, Lehner AF, Muldoon TG, Hutson MS, Mahesh VB. The stereochemical complementarity of DNA and reproductive steroid hormones correlates with biological activity. J Steroid Biochem Mol Biol 24: 843–852, 1986. Hendry LB, Bransome ED, Mahesh VB. The ligand insertion hypothesis in the genomic action of steroid hormones. J Steroid Biochem Mol Biol 65: 75–89, 1998. Hendry LB, Mahesh VB. Stereochemical complementarity of progesterone and cavities between base pairs in partially unwound double stranded DNA using computer modeling and energy calculations to determine degree of fit. J Steroid Biochem Mol Biol 39: 133–146, 1991. Hendry LB, Mahesh VB. A putative step in steroid hormone action involves insertion of steroid ligands into DNA facilitated by receptor proteins. J Steroid Biochem Mol Biol 55: 173–183, 1995. Herbison AE, Robinson JE, Skinner DC. Distribution of estrogenimmunoreactive cells in the preoptic area of the ewe: co-localization with glutamic acid decarboxylase but not luteinizing hormone-releasing hormone. Neuroendocrinology 57: 751–759, 1993. Hoff JD, Quigley ME, Yen SSC. Hormonal dynamics in mid cycle: a reevaluation. J Clin Endocrinol Metab 57: 792–796, 1983. Jensen EV. On the mechanism of estrogen action. Perspect Biol Med 6: 47–59, 1962. Jungck EC, Roy S, Greenblatt RB, Mahesh VB. Effect of Clomiphene Citrate on spermatogenesis in the human. Fertil Steril 15: 40 –43, 1964. Khan MM, De Sevilla L, Mahesh VB, Brann DW. Enhanced glutamatergic and decreased GABAergic synaptic appositions to GnRH neurons on proestrus in the rat: modulatory effect of aging. PLoS One 5: e10172, 2010. Khan MM, Hadman M, Wakade C, DeSevilla L, Dhandapani KM, Mahesh VB, Vadlamudi RK, Brann DW. Cloning, expression and localization of MNAR/PELP1 in rodent brain: colocalization in estrogen receptor-␣ but not in gonadotropin-releasing hormone-positive neurons. Endocrinology 146: 5215–5227, 2005. Knudsen JF, Costoff A, Mahesh VB. Dehydroepiandrosterone-induced polycystic ovaries and acyclicity in the rat. Fertil Steril 26: 807–817, 1975. Knudsen JF, Mahesh VB. Initiation of precocious sexual maturation in the immature rat treated with dehydroepiandrosterone. Endocrinology 97: 458 –468, 1975. Lamar CA, Bhat GK, Mahesh VB, Brann DW. Evidence that neuronal nitric oxide synthase but not heme oxygenase increases in the hypothalamus on proestrus afternoon. Neuroendocrinology 70: 360 –367, 1999. Lamar CA, Mahesh VB, Brann DW. Regulation of gonadotropinreleasing hormone (GnRH) secretion by heme molecules: a regulatory role for carbon monoxide? Endocrinology 137: 790 –793, 1996. Legan SJ, Karsch FJ. A daily signal for LH surge in the rat. Endocrinology 96: 57–62, 1975. Lehman MN, Karsch FJ. Do gonadotropin-releasing hormone, tyrosine hydroxylase, and beta endorphin immunoreacive neurons contain estrogen receptors? A double-label immunocytochemical study in the Suffolk ewe. Endocrinology 133: 887–895, 1993. Lesoon LA, Mahesh VB. Stimulatory and inhibitory effects of progesterone on FSH secretion by the anterior pituitary. J Steroid Biochem Mol Biol 42: 479 –491, 1992. AJP-Endocrinol Metab • doi:10.1152/ajpendo.00488.2011 • www.ajpendo.org Downloaded from http://ajpendo.physiology.org/ by 10.220.33.3 on June 18, 2017 66. Dhandapani KM, Hadman M, De Seville L, Wade MF, Mahesh VB, Brann DW. Astrocyte protection of neurons: role of transforming growth factor-beta signaling via c-Jun-AP-1 protective pathway. J Biol Chem 278: 44329 –44339, 2003. 67. Dhandapani KM, Wade MF, Mahesh VB, Brann DW. Basic fibroblast growth factor induces TGF-beta release in an isoform and gliomaspecific manner. Neuroreport 13: 239 –41, 2002. 68. Dhandapani KM, Wade MF, Mahesh VB, Brann DW. Astrocyte derived transforming growth factor- mediates the neuroprotective effects of 17-estradiol: involvement of non-classical genomic signaling pathways. Endocrinology 146: 2749 –2759, 2005. 69. Dhandapani KM, Wade MF, Mahesh VB, Brann DW. Neuroprotection by stem cell factor in rat cortical neurons involves Akt and NFB. J Neurochem 95: 9 –19, 2005. 70. Dulin WE, Bowman BJ, Stafford RO. Effects of 2-methylation on glucocorticoid activity of various C-21. Proc Soc Exp Biol Med 94: 303–305, 1957. 71. El Ayat AAB, Mahesh VB. Stimulation of 17-hydroxy steroid dehydrogenase in the rat anterior pituitary gland by progesterone. J Steroid Biochem Mol Biol 20: 1141–1145, 1984. 72. Eldridge JC, Dmowski WP, Mahesh VB. Effects of castration of immature rats on serum FSH and LH, and of various steroid treatments after castration. Biol Reprod 10: 438 –446, 1974. 73. Eldridge JC, Mahesh VB. The pituitary-gonadal axis before puberty: evaluation of testicular steroids in the male rat. Biol Reprod 11: 385–397, 1974. 74. Eldridge JC, McPherson JC, Mahesh VB. Maturation of the negative feedback control of gonadotropin secretion in the female rat. Endocrinology 94: 1536 –1540, 1974. 75. Fuentes MA, Muldoon TG, Mahesh VB. Inhibitory effect of progesterone on occupied estrogen receptors of anterior pituitary and uterus in adult rats. J Neuroendocrinol 2: 517–522, 1990. 76. Fuentes MA, Muldoon TG, Mahesh VB. Role of 17-hydroxysteroid dehydrogenase in the modulation of nuclear estradiol receptor binding by progesterone in the rat anterior pituitary gland and the uterus. J Steroid Biochem Mol Biol 37: 57–63, 1990. 77. Fuentes MA, Muldoon TG, Mahesh VB. Inhibitory effect of 5-alphadihydroprogesterone on nuclear estrogen receptor binding of the anterior pituitary and uterus in the rat. Neuroendocrinology 52: 213–220, 1990. 78. Gambrell RD, Greenblatt RB, Mahesh VB. Serum gonadotropins levels and ancillary studies in Stein-Leventhal syndrome treated with Clomiphene Citrate. Obstet Gynecol 38: 850 –862, 1971. 79. Gambrell RD, Greenblatt RB, Mahesh VB. Serum gonadotropins in a variety of anovulatory women treated with Clomiphene Citrate. Int J Gynecol Obstet 11: 90 –95, 1973. 80. Gambrell RD, Greenblatt RB, Mahesh VB. Inappropriate secretion of LH in the Stein-Leventhal syndrome. Obstet Gynecol 42: 429 –440, 1973. 81. Gao X, Greenbaum L, Mahesh VB, Brann DW. Characterization of the kinin system in the ovary during ovulation in the rat. Biol Reprod 47: 945–951, 1992. 82. Glenn EM, Stafford RO, Lyster SC, Bowman BJ. Relationship between biological activity of hydrocortisone analogues and their rates of inactivation by rat liver enzymes. Endocrinology 61: 128 –142, 1957. 83. Goldman BD, Kamberi IA, Siiteri K, Porter JC. Temporal relationship of progesterone secretion, LH release and ovulation in rats. Endocrinology 85: 1137–1143, 1979. 84. Goldman BD, Mahesh VB. Fluctuations in pituitary FSH during the ovulatory cycle in the rat and a possible role of FSH in the induction of ovulation. Endocrinology 83: 97–106, 1968. 85. Goldman BD, Mahesh VB. A possible role of acute FSH-release in ovulation in the hamster, as demonstrated by utilization of antibodies to LH and FSH. Endocrinology 84: 236 –243, 1969. 86. Goldman BD, Mahesh VB, Porter JC. The role of the ovary in control of cyclic LH release in the hamster, Mesocricetus auratus. Biol Reprod 4: 57–65, 1971. 87. Greeley GH Jr, Allen MB, Mahesh VB. FSH and LH rsponse in the rat after intravenous, intracarotid or subcutaneous administration of LH-RH. Proc Soc Expl Biol Med 147: 859 –862, 1974. 88. Greeley GH Jr, Allen MB, Mahesh VB. Potentiation of luteinizing hormone release by estradiol at the level of the pituitary. Neuroendocrinology 18: 233–241, 1975. 89. Greeley GH Jr, Muldoon TG, Mahesh VB. Correlative aspects of luteinizing hormone-releasing hormone sensitivity and cytoplasmic es- E15 Perspectives E16 STEROID-GONADOTROPIN-FEEDBACK SYSTEM: CAREER RETROSPECTIVE 138. Mahesh VB, Toledo SPA, Mattar E. Hormone levels following wedge resection in polycystic ovary syndrome. Obstet Gynecol 51: 64 –69, 1978. 139. Mahesh VB, Ulrich F. Distribution of enzyme systems responsible for steroid metabolism in different tissues and subcellular particles. Nature 184, Suppl 15: 1147–1148, 1959. 140. Mahesh VB, Ulrich F. Metabolism of cortisol and cortisone by various tissues and subcellular particles. J Biol Chem 235: 356 –360, 1960. 141. Mahesh VB, Zamorano P, De Sevilla L, Lewis D, Brann DW. Characterization of ionotropic glutamate receptors in rat hypothalamus, pituitary and immortalized gonadotropin-releasing hormone (GnRH) neurons (GT1–7 Cells). Neuroendocrinology 69: 397–407, 1999. 142. Mahesh VB, Zarate A, Roper BK, Greenblatt RB. Studies on the action of 17␣-methyl-B-nortestosterone as an antiandrogen. Steroids 8: 297–308, 1966. 143. McDonough PC, Ellegood JO, Byrd JR, Mahesh VB. Ovarian and peripheral venous steroids in XY gonadal dysgenesis and gonadoblastoma. Obstet Gynecol 47: 351–355, 1976. 144. McPherson JC III, Costoff A, Eldridge JC, Mahesh VB. Effect of various progestational preparations on gonadotropin secretion in ovariectomized immature female rats. Fertil Steril 25: 1063–1070, 1974. 145. McPherson JC III, Costoff A, Mahesh VB. The pituitary-gonadal axis before puberty: effects of various estrogenic steroids in the ovariectomized rat. Steroids 24: 41–56, 1974. 146. McPherson JC III, Costoff A, Mahesh VB. Influence of estrogenprogesterone combinations on gonadotropin secretion in castrate female rats. Endocrinology 97: 771–779, 1975. 147. McPherson JC III, Costoff A, Mahesh VB. Effects of aging on the hypothalamic-hypophyseal-gonadal axis in female rats. Fertil Steril 28: 1365–1370, 1977. 148. McPherson JC III, Mahesh VB. Dose related effect of a single injection of progesterone on gonadotropin secretion and pituitary sensitivity to LHRH in estrogen-primed castrate female rats. Biol Reprod 20: 763–772, 1979. 149. McPherson JC III, Mahesh VB. Induction of LH and FSH surge in the estrogen-primed castrated male rat by progesterone. Biol Reprod 27: 1222–1229, 1982. 150. Mehta SH, Dhandapani KM, De Seville LM, Webb RC, Mahesh VB, Brann DW. Tamoxifen, a selective estrogen receptor modulator, reduces ischemic damage caused by middle cerebral artery occlusion in the ovariectomized female rat. Neuroendocrinology 77: 44 –50, 2003. 151. Mills TM, Lin TJ, Braselton WE Jr, Ellegood JO, Mahesh VB. Metabolism of oral contraceptive drugs. The formation and disappearance of metabolites of Norethindrone and Mestranol after intravenous and oral administration. Am J Obstet Gynecol 126: 987–992, 1976. 152. Mills TM, Lin TJ, Hernandez-Ayup S, Greenblatt RB, Ellegood JO, Mahesh VB. The metabolic clearance rate (MCR) and urinary excretion of oral contraceptive drugs. I. Norethindrone. Am J Obstet Gynecol 120: 764 –772, 1974. 153. Mills TM, Lin TJ, Hernandez-Ayup S, Greenblatt RB, Ellegood JO, Mahesh VB. The metabolic clearance rate (MCR) and urinary excretion of oral contraceptive drugs. II. Mestranol. Am J Obstet Gynecol 120: 773–778, 1974. 154. Morris JM, Mahesh VB. Further observations on the syndrome, “Testicular Feminization.” Am J Obstet Gynecol 87: 731–745, 1963. 155. Murphy LL, Mahesh VB. Selective release of FSH by 5␣-dihydroprogesterone in immature ovariectomized estrogen-primed rats. Biol Reprod 30: 594 –602, 1984. 156. Murphy LL, Mahesh VB. Selective release of luteinizing hormone by 3␣-hydroxy-5␣-pregnan-20-one in immature ovariectomized estrogenprimed rats. Biol Reprod 30: 795–803, 1984. 157. Murphy LL, Mahesh VB. Selective release of FSH and LH by 5␣,dihydroprogesterone, and 3␣, 5␣-tetrahydroprogesterone in pregnant mare’s serum gonadotropin-primed immature rats exposed to constant light. Biol Reprod 32: 795–803, 1985. 158. Natrajan PK, Muldoon TG, Greenblatt RB, Mahesh VB. Estradiol and progesterone receptors in estrogen-primed endometrium. Am J Obstet Gynecol 140: 387–392, 1981. 159. Natrajan PK. On estrogen and progesterone receptors of the human endometrium. J Reprod Med 27: 227–230, 1982. 160. Nazian SJ, Mahesh VB. Alterations in end organ responsiveness to testosterone during the sexual maturation of the male rat. Biol Reprod 21: 465–471, 1979. AJP-Endocrinol Metab • doi:10.1152/ajpendo.00488.2011 • www.ajpendo.org Downloaded from http://ajpendo.physiology.org/ by 10.220.33.3 on June 18, 2017 113. Mahesh VB. The dynamic interaction between steroids and gonadotropins in the mammalian ovulatory cycle. Neurosci Biobehav Rev 9: 245–260, 1985. 114. Mahesh VB, Brann DW. Interaction between ovarian and adrenal steroids in the regulation of gonadotropin secretion. J Steroid Biochem Mol Biol 41: 495–513, 1992. 115. Mahesh VB, Brann DW. Neuroendocrine mechanisms underlying the control of gonadotropin secretion by steroids. Steroids 63: 252–256, 1998. 116. Mahesh VB, Dhandapani KM, Brann DW. Role of astrocytes in reproduction and neuroprotection. Mol Cell Endocrinol 246: 1–9, 2006. 117. Mahesh VB, Ewing DE, Hendry LB. Assessing activity and toxicity of drugs in silico based on DNA structure. Med Chem Res 17: 159 –168, 2008. 118. Mahesh VB, Ewing DE, Sidell N, Bransome ED Jr, Hendry LB. New technology for drug discovery based upon insertion of ligands into gene sequences by nuclear receptor proteins. Med Chem Res 13: 497–508, 2004. 119. Mahesh VB, Greenblatt RB. Physiology and pathogenesis of the Stein-Leventhal syndrome. Nature 191: 888 –890, 1961. 120. Mahesh VB, Greenblatt RB. Isolation of dehydroepiandrosterone and 17␣-hydroxy-delta-5-pregnenolone from the polycystic ovaries of the Stein-Leventhal syndrome. J Clin Endocrinol Metab 22: 441–448, 1962. 121. Mahesh VB, Greenblatt RB. The in vivo conversion of dehydroepiandrosterone and androstenedione to testosterone in the human. Acta Endocrinol 41: 400 –406, 1962. 122. Mahesh VB, Greenblatt RB. Steroid secretions of the normal and polycystic ovary. Recent Prog Horm Res 20: 341–394, 1964. 123. Mahesh VB, Greenblatt RB. Urinary steroid excretion patterns in hirsutism. II. Effect of ovarian stimulation with human pituitary FSH on urinary 17-ketosteroids. J Clin Endocrinol Metab 24: 1293–1302, 1964. 124. Mahesh VB, Greenblatt RB. Hormonal studies in females with virilizing adrenal tumors. Res Steroids 3: 221–231, 1967. 125. Mahesh VB, Greenblatt RB, Aydar CK, Roy S. Secretion of androgens by the polycystic ovary and its significance. Fertil Steril 13: 513–530, 1962. 126. Mahesh VB, Greenblatt RB, Aydar CK, Roy S, Puebla RA, Ellegood JO. Urinary steroid excretion patterns in hirsutism. I. Use of adrenal and ovarian suppression tests in the study of hirsutism. J Clin Endocrinol Metab 24: 1283–1292, 1964. 127. Mahesh VB, Greenblatt RB, Coniff RF. Urinary steroid excretion before and after dexamethasone administration and steroid content of adrenal tissue and venous blood in virilizing adrenal tumors. Am J Obstet Gynecol 100: 1043–1054, 1968. 128. Mahesh VB, Greenblatt RB, Coniff RF. Adrenal hyperplasia—a case report of delayed onset of the congential form or an acquired form. J Clin Endocrinol Metab 28: 619 –623, 1968. 129. Mahesh VB, Goldman BD. Secretion of FSH and LH during the ovulatory cycle and their role in the induction of ovulation and corpus luteum function. In: “Hormonal Steroids”. Excerpta Med Int Congr Ser 219: 662–669, 1972. 130. Mahesh VB, Herrmann W. Isolation of estrone and 11-hydroxy estrone from a feminizing adrenal carcinoma. Steroids 1: 51–61, 1962. 131. Mahesh VB, McDonough PG. Urinary steroid excretion in a hirsute patient with ovarian cortical stromal hyperplasia and endometrial carcinoma. Am J Obstet Gynecol 104: 602–603, 1969. 132. Mahesh VB, McDonough PG, Deleo CA. Endocrine studies in the arrhenoblastoma. Am J Obstet Gynecol 107: 183–187, 1970. 133. Mahesh VB, McDonough PG, Greenblatt RB. Endocrine studies in a patient with virilism, ovarian stromal hyperplasia, and endometrial carcinoma. Obstet Gynecol 30: 584 –590, 1967. 134. Mahesh VB, Muldoon TG. Integration of the effects of estradiol and progesterone in the modulation of gonadotropin secretion. J Steroid Biochem Mol Biol 27: 665–675, 1987. 135. Mahesh VB, Nazian SJ. Role of sex steroids in the initiation of puberty. J Steroid Biochem Mol Biol 11: 587–591, 1979. 136. Mahesh VB, Roach LW, Cannady WE, Ogle TF, Brann DW, Bransome ED Jr, Hendry LB. Gene based pharmacophores for drug design. Med Chem Res 10: 440 –455, 2001. 137. Mahesh VB, Sidell N, Ewing DE, Bransome ED Jr, Hendry LB. A molecular guidance system based upon target genes, nuclear receptors and ligands applied to drug discovery and prediction of toxicity. MiniReviews Med Chem 8: 1252–1264, 2008. Perspectives STEROID-GONADOTROPIN-FEEDBACK SYSTEM: CAREER RETROSPECTIVE 183. Peterson RE, Pierce CE, Wyngaarden JB, Bunim JJ, Brodie BB. The physiological disposition and metabolic fate of cortisone in man. J Clin Invest 36: 1301–1312, 1957. 184. Ping L, Mahesh VB, Bhat GK, Brann DW. Regulation of gonadotropin-releasing hormone and luteinizing hormone secretion by AMPA receptors. Neuroendocrinology 66: 246 –253, 1997. 185. Ping L, Mahesh VB, Brann DW. A physiological role for NMDA and non-NMDA receptors in pulsatile gonadotropin secretion in the adult female rat. Endocrinology 135: 113–118, 1993. 186. Ping L, Mahesh VB, Brann DW. Effect of NMDA and non-NMDA recptor antagonists on pulsatile luteinizing hormone secretion in the adult male rat. Neuroendocrinology 61: 226 –234, 1995. 187. Ping L, Mahesh VB, Wiedmeier VT, Brann DW. Release of glutamate and aspartate from the preoptic area during the progesterone-induced LH surge: in vivo microdialysis studies. Neuroendocrinology 59: 318 –324, 1994. 188. Piva F, Maggi R, Limonta P, Motta M, Martini L. Effect of naloxone on luteinizing hormone, follicle stimulating hormone and prolactin sevretion in the different phases of the estrous cycle. Endocrinology 117: 766 –772, 1985. 189. Putnam CD, Brann DW, Kolbeck RC, Mahesh VB. Inhibition of uterine contractility by progesterone and progesterone metabolites: mediation by progesterone and gamma amino butyric acid A receptor systems. Biol Reprod 45: 246 –272, 1991. 190. Putnam CD, Brann DW, Mahesh VB. Acute activation of the ACTHadrenal axis: effect upon gonadotropin and prolactin secretion in the female rat. Endocrinology 128: 2558 –2566, 1991. 191. Putnam-Roberts CD, Brann DW, Mahesh VB. Role of 5␣-reduction in progesterone’s ability to release FSH in estrogen-primed ovariectomized rats. J Steroid Biochem Mol Biol 42: 875–882, 1992. 192. Rao IM, Allsbrook WC Jr, Conway BA, Martinez JE, Beck JR, Pantazis CG, Mills TM, Anderson E, Mahesh VB. Flow cytometric analysis of granulosa cells from developing rat follicles. J Reprod Fertil 91: 521–530, 1991. 193. Rao IM, Mahesh VB. Role of progesterone in the modulation of the preovulatory surge of gonadotropins and ovulation in the pregnant mare’s serum gonadotropin-primed immature rat and the adult rat. Biol Reprod 35: 1154 –1161, 1986. 194. Rao IM, Mills TM, Anderson E, Mahesh VB. Heterogeneity in granulosa cells of developing rat follicles. Anat Rec 229: 177–185, 1991. 195. Rapkin AJ, Morgan M, Goldman L, Brann DW, Simone D, Mahesh VB. Progesterone metabolite allopregnanolone in women with premenstrual syndrome. Obstet Gynecol 90: 709 –714, 1997. 196. Rapkin AJ, Shoupe D, Reading A, Bohn Y, Brann DW, Mahesh VB. Decreased central opioid activity in premenstrual syndrome: luteinizing hormone response to naloxone. J Soc Gynecol Invest 3: 93–98, 1996. 197. Rosser M, Chorich L, Howard E, Zamorano P, Mahesh VB. Changes in rat uterine estrogen receptor mRNA levels during estrogen and progesterone induced estrogen receptor depletion and subsequent replenishment. Biol Reprod 48: 89 –98, 1993. 198. Roy S, Greenblatt RB, Mahesh VB. Effects of clomiphene on the physiology of reproduction in the rat. II. Its oestrogenic and antioestrogenic actions. Acta Endocrinol 47: 657–668, 1964. 199. Roy S, Greenblatt RB, Mahesh VB, Jungck EC. Clomiphene citrate: further observations on its use in induction of ovulation in the human and on its mode of action. Fertil Steril 14: 575–595, 1963. 200. Roy S, Mahesh VB, Greenblatt RB. Effects of clomiphene on the physiology of reproduction in the rat. I. Changes in the hypophysealgonadal axis. Acta Endocrinol 47: 645–656, 1964. 201. Roy S, Mahesh VB, Greenblatt RB. Effects of clomiphene on the physiology of reproduction in the rat. III. Inhibition of uptake of radioactive oestradiol by the uterus and the pituitary gland of immature rat. Acta Endocrinol 47: 669 –675, 1964. 202. Roy S, Mahesh VB, Greenblatt RB. Effect of dehydroepiandrosterone and delta-4-androstenedione on the reproductive organs of female rats: production of cystic change in the ovary. Nature 196: 42–43, 1962. 203. Sato T, Greenblatt RB, Mahesh VB. Levels of luteinizing hormone during the menstrual cycle determined by immunologic techniques. Fertil Steril 16: 223–228, 1965. 204. Schenken RS, Werlin LB, Williams RF, Prihods TJ, Hodgen GD. Periovulatory hormonal dynamics: relationships of immunoassayable gonadotropins and ovarian steroids to the bioassayable LH surge in Rhesus monkey. J Clin Endocrinol Metab 60: 886 –890, 1985. AJP-Endocrinol Metab • doi:10.1152/ajpendo.00488.2011 • www.ajpendo.org Downloaded from http://ajpendo.physiology.org/ by 10.220.33.3 on June 18, 2017 161. Nazian SJ, Mahesh VB. Sexual maturation of the male rat. Influence of androgens on the pituitary response to single or multiple injections of LH-RH. Neuroendocrinology 29: 313–322, 1979. 162. Nazian SJ, Mahesh VB. Delay in some aspects of the sexual maturation of the male rat induced by androstenedione. Biol Reprod 22: 451–458, 1980. 163. Nazian SJ, Mahesh VB. Hypothalamic, pituitary, testicular, and secondary organ functions and interactions during the sexual maturation of the male rat. Arch Androl 4: 283–303, 1980. 164. Nazian SJ, Mahesh VB. LH-induced testicular steroid secretion after FSH treatment in immature and young adult hypophysectomized male rats. Arch Androl 6: 33–38, 1981. 165. Nequin LG, Alvarez J, Schwartz NB. Measurement of serum steroid and gonadotropin levels and uterine and ovarian variables throughout 4 day and 5 day estrus cycles in the rat. Biol Reprod 20: 659 –670, 1979. 166. O’Conner JL, Lapp CA, Mahesh VB. Peptidase activity in the hypothalamus and pituitary of the rat: fluctuations and possible regulatory role of luteinizing hormone releasing hormone-degrading activity during the estrous cycle. Biol Reprod 30: 855–862, 1984. 167. O’Conner JL, Mahesh VB. A possible role for progesterone in the preovulatory gonadotropin surge through modulation of LHRH degrading activity. J Steroid Biochem Mol Biol 29: 257–264, 1988. 168. O’Conner JL, Wade MF, Brann DW, Mahesh VB. Direct anterior pituitary modulation of gonadotropin secretion by neuropeptide Y (NPY): role of gonadal steroids. Neuroendocrinology 58: 129 –135, 1993. 169. O’Conner JL, Wade MF, Brann DW, Mahesh VB. Evidence that progesterone modulates anterior pituitary neuropeptide Y levels during the progesterone-induced gonadotropin surge in the estrogen-primed intact immature female rat. J Steroid Biochem Mol Biol 52: 497–504, 1995. 170. O’Conner JL, Wade MF, Edwards DP, Mahesh VB. Progesterone and regulation of the follicle-stimulating hormone (FSH-beta) gene. Steroids 64: 592–597, 1999. 171. O’Conner JL, Wade MF, Prendergast P, Edwards DP, Boonyaratanakornkit V, Mahesh VB. A 361 base pair region of the rat FSH-b promoter contains multiple progesterone receptor binding sequences and confers progesterone responsiveness. Mol Cell Endocrinol 136: 67–78, 1997. 172. Ogle TF, Dai D, George P, Mahesh VB. Stromal cell progesterone and estrogen receptors during proliferation and regression of the decidua basalis in the pregnant rat. Biol Reprod 57: 495–506, 1997. 173. Ogle TF, Da D, George P, Mahesh VB. Regulation of the progesterone receptor and estrogen receptor in decidua basalis by progesterone and estradiol during pregnancy. Biol Reprod 58: 1188 –1198, 1998. 174. Ogle TF, Soares MJ, Zhou Y, Chorich L, Mahesh VB. Regulation of the estrogen receptor and estrogen receptor mRNA in rat decidua basalis and placenta during pregnancy. Steroid Biochem (Life Sci Adv) 12: 1–12, 1993. 175. Ondo JG, Pass KA, Baldwin R. The effect of neurally active aminoacids on the pituitary gonadotropin secretion. Neuroendocrinology 21: 79 –89, 1976. 176. Parker CR, Bruneteau DW, Greenblatt RB, Mahesh VB. Peripheral, ovarian, and adrenal vein steroids in hirsute women: acute effects of human chorionic gonadotropin and adrenocorticotrophic hormone. Fertil Steril 26: 877–888, 1975. 177. Parker CR, Costoff A, Muldoon TG, Mahesh VB. Actions of pregnant mare serum gonadotropin in the immature female rat: correlative changes in steroids, gonadotropins, and cytoplasmic estradiol receptors of the anterior pituitary and hypothalamus. Endocrinology 98: 129 –138, 1976. 178. Parker CR, Mahesh VB. Interrelationship between excessive levels of circulating androgens in blood and ovulatory failure. J Reprod Med 17: 75–90, 1976. 179. Parker CR, Mahesh VB. Hormonal events surrounding the natural onset of puberty in female rats. Biol Reprod 14: 347–353, 1976. 180. Parker CR, Mahesh VB. Dehydroepiandrosterone (DHA) induced precocious ovulation: correlative changes in blood steroids, gonadotropins and cytosol estradiol receptors of anterior pituitary gland and hypothalamus. J Steroid Biochem Mol Biol 8: 173–177, 1977. 181. Parker CR, Servy E, McDonough PG, Mahesh VB. In vivo endocrine studies in adrenal rest tumor of ovary. Obstet Gynecol 44: 327–332, 1974. 182. Peduto JC, Mahesh VB. Effects of progesterone on hypothalamic and plasma LHRH. Neuroendocrinology 40: 238 –245, 1985. E17 Perspectives E18 STEROID-GONADOTROPIN-FEEDBACK SYSTEM: CAREER RETROSPECTIVE 216. Wakade C, Khan MM, DeSevilla LM, Zhang QG, Mahesh VB, Brann DW. Tamoxifen neuroprotection in cerebral ischemia involves attenuation of kinase activation and superoxide production and potentiation of mitochondrial superoxide dismutase. Endocrinology 149: 367– 79, 2008. 217. Ward RC, Costoff A, Mahesh VB. The induction of polycystic ovaries in mature cycling rats by the administration of dehydroepiandrosterone (DHA). Biol Reprod 18: 614 –623, 1978. 218. Zamorano PL, De Sevilla L, Mahesh VB, Brann DW. Production and refolding of recombinant leptin. BioTechniques 23, No.5, 1997. 219. Zamorano PL, Mahesh VB, De Sevilla L, Brann DW. Excitatory amino acid receptors and puberty. Steroids 63: 268 –270, 1998. 220. Zamorano P, Mahesh VB, De Sevilla LM, Chorich LP, Bhat GK, Brann DW. Expression and localization of the leptin receptor in endocrine and neuroendocrine tissues of the rat. Neuroendocrinology 65: 223–228, 1997. 221. Zarate A, Mahesh VB, Greenblatt RB. Effect of an antiandrogen, 17␣-methyl-B-nortestosterone, on acne and hirsutism. J Clin Endocrinol Metab 26: 1394 –1398, 1966. 222. Zhang QG, Wang R, Khan MM, Mahesh VB, Brann DW. Role of Dickkopf-1 (Dkk1), an antagonist of the Wnt--catenin signaling pathway, in estrogen-induced neuroprotection and attenuation of Tau phosphorylation. J Neurosci 28: 8430 –8441, 2008. 223. Zhou Y, Chorich LP, Mahesh VB, Ogle TF. Regulation of estrogen receptor protein and messenger ribonucleic acid by estradiol and progesterone in rat uterus. J Steroid Biochem Mol Biol 46: 687–698, 1993. 224. Zhou Y, Mahesh VB, Ogle TF. The role of rat uterine microsomal estrogen receptor in ovarian steroid hormone action. Endocr J 2: 53–62, 1994. 225. Zhou Z, Mahesh VB, Zamorano PL, Brann DW. Decreased gonadotropin-releasing hormone neurosecretory response to glutamate agonists in middle-aged female rats on proestrus afternoon: a possible role in reproductive aging. Endocrinology 137: 2334 –2338, 1996. AJP-Endocrinol Metab • doi:10.1152/ajpendo.00488.2011 • www.ajpendo.org Downloaded from http://ajpendo.physiology.org/ by 10.220.33.3 on June 18, 2017 205. Shi B, Bhat G, Mahesh VB, Brotto M, Nosek TM, Brann DW. Bradykinin receptor localization and cell signaling pathways used by bradykinin in the regulation of gonadotropin-releasing hormone secretion. Endocrinology 140: 4669 –4676, 1999. 206. Shi B, Mahesh VB, Bhat GK, Ping L, Brann DW. Evidence for a role of bradykinin neurons in the control of gonadotropin-hormone releasing hormone secretion. Neuroendocrinology 67: 209 –218, 1998. 207. Simone DA, Chorich LP, Mahesh VB. Mechanisms of action for an androgen-mediated autoregulatory process in rat thecal-interstitial cells. Biol Reprod 49: 1190 –1201, 1993. 208. Simone DA, Mahesh VB. An autoregulatory process for androgen production in rat thecal-interstitial cells. Biol Reprod 48: 46 –56, 1993. 209. Smanik E, Calderon J, Muldoon TG, Mahesh VB. Effect of progesterone on the activity of occupied nuclear estrogen receptor in vitro. Mol Cell Endocrinol 64: 111–117, 1989. 210. Smanik EJ, Young HK, Muldoon TG, Mahesh VB. Analysis of the effect of progesterone in vivo on estrogen receptor distribution in the rat anterior pituitary and hypothalamus. Endocrinology 113: 15–22, 1983. 211. Strott CA, Cargille CM, Ross GT, Lipsett MB. The short luteal phase. J Clin Endocrinol Metab 30: 246 –251, 1970. 212. Talalay P, Williams-Ashman HG. Activation of hydrogen transfer between pyridine nucleotides by steroid hormones. Proc Natl Acad Sci USA 44: 15–26, 1958. 213. Trott EA, Plouffe L Jr, Hansen K, Hines R, Brann DW, Mahesh VB. Ovulation induction in clomiphene-resistant anovulatory women with normal dehydroepiandrosterone sulfate levels: beneficial effects of the addition of dexamethasone during the follicular phase. Fertil Steril 66: 484 –486, 1996. 214. Uberoi NK, Mahesh VB, Black J, Meyer RK. Effect of 17␣-methylB-nortestosterone on ovulation and ovarian weight in female rats. Steroids 9: 245–255, 1967. 215. Unda R, Brann DW, Mahesh VB. Progesterone suppression of glutamic acid decarboxylase (GAD67) mRNA levels in the preoptic area: correlation to the luteinizing hormone surge. Neuroendocrinology 62: 562–570, 1995.