Survey
* Your assessment is very important for improving the workof artificial intelligence, which forms the content of this project
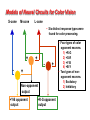
OPTO 5320 VISION SCIENCE I Monocular Sensory Processes of Vision: Color Vision Color Vision Theories Color Vision Theories Modern research has described many aspects of color vision processing. Two major theories have guided the research on color vision. 1. the Trichromatic Theory (Young-Helmholtz theory) 2. the Opponent-Processes Theory (Hering’s theory). These complementary theories are important in explaining color vision at different levels of the visual system. Color Vision Theories A. Requirements for a theory of color vision B. Young-Helmholtz’s Trichromatic Theory C. Hering’s Opponent Colors Theory Color Vision Theories • A viable color vision theory should be able to explain, in a quantitative way, all of the phenomena of color. • Especially, a color vision theory must be able to account for the following. 1) Color mixture and Grassmann’s laws 2) Discrimination of hue, saturation, and brightness 3) The perception of spectral hues 4) Dichromatic and trichromatic color vision anomalies 5) The perception of violet 6) The appearance of after-images 7) The Bezold-Brucke effect History – Isaac Newton http://www.crystalinks.com/ • 1672 - Newton demonstrated the prismatic dispersion of white light to obtain the spectral colors. He assumed that when light struck the retina and it started to vibrate at a frequency proportional to the wavelength, much like the cochlea of the inner ear responds to sound. http://www.ucolick.org/ History – Thomas Young http://www.eoht.info/ 1801 - Young’s Bakerian Lecture, “it is almost impossible to conceive each sensitive point on the retina to contain an infinite number of particles each capable of vibrating in perfect unison with every possible undulation, it becomes necessary to suppose the number is limited, for instance to the three principal colors - red, yellow and blue.” Later Young corrected the primaries to red, green and violet. Young also described the color blindness of his friend, John Dalton, as missing the particle vibrating in response to red (protanopia). Color blindness became known as “Daltonism.” Young-Helmholtz’s Trichromatic Color Vision Theory http://120years.net/ 1866 - Helmholtz presented a relatively complete description of color vision based on Young’s original proposal of three fundamental retinal mechanisms. The essence of the theory is that three sets of sensory mechanisms determine the quantitative basis for the various aspects of color vision. Each of the mechanisms is sensitive across the entire visible spectrum, but they each differ in their spectra response properties. Young-Helmholtz’s Trichromatic Color Vision Theory The basic elements of the Young-Helmholtz Trichromatic Color Theory. 1) Each sensory mechanism has a specific sensitivity curve at each spectral wavelength for each of the three mechanisms 2) Hue is a function of the relative responses of the three mechanisms at each wavelength. 3) Brightness is a function of the total of the responses of the three mechanisms at each wavelength. Hecht’s Trichromatic Color Vision Theory http://www.nap.edu/ The elementary hypotheses of a trichromatic theory do not restrict the shape or locations of the cone fundamentals. Hecht developed a mathematical account of the physiological processes of color vision based on the three fundamental cone response curves shown in the figure. Enduring Principles of Trichromatic Color Theory Color information is initially encoded by three cone mechanisms. The fundamental phenomena of color vision are a result of neural processes which combine the signals from the three cone mechanisms in various ways. 1) Color mixture and Grassman’s laws 2) Discrimination of hue, saturation, and brightness. 3) The perception of spectral hues, except yellow 4) Dichromatic, but not trichromatic, color vision anomalies Hering’s Opponent-Colors Theory 1872 - Hering proposed a theory of color vision to account for Luminance intensity perceptual or psychological aspects of color vision. 1. The perception of yellow in the 560 - 580 nm range. 2. Negative after-images are complements of induction stimuli. 3. The Bezold-Brucke effect: The perceived hues of monochromatic stimuli change with intensity, except at specific wavelengths (unique hues). 4. Unique hues represent the locations of cone photopigments with approximately equal sensitivity to a single wavelength Wavelength (nm) Hering’s Opponent-Colors Theory 1. http://www.orden-pourlemerite.de/ Hering’s (1872) theory was based on 2. 3. fundamental color vision response functions with negative as well as positive values. Three retinal substrates which respond to light by opposite anabolic and catabolic reactions. 1. catabolic by light - anabolic by dark (bk/wh opponency). 2. catabolic by red light - anabolic by green light (R/G opponency). 3. catabolic by yellow light - anabolic by blue light (B/Y opponency). Hering’s Opponent-Colors Theory • Photopigments of the type postulated by Hering do not exist. • Light activated changes in chromophores are identical in all photopigments. • Opponent color responses can occur from excitatory and inhibitory synaptic substances of neural responses. Opponent-Colors Theory • There is considerable psychophysical evidence for opponent color responses. • One line of evidence has been obtained by hue cancellation measurements the intensity of a primary of one opponent pair to cancel the perception of hue in the other opponent pair, i.e., the amount of yellow (570 nm) light to cancel the blueness of lights from 400 - 500 nm. Enduring Principles of OpponentColors Theory • Neural mechanisms at some level of the visual pathway must combine information from three cone types to form two opponent color vision channels, the R/G & B/Y channels, and one non-opponent channel, the luminance or bk/wh channel. • Color perception is better explained by the information in opponent channels than trichromatic color vision theories. Multi-stage Color Vision Theories • The Young-Helmholtz Trichromatic theory and the Hering Opponent-colors theory are one-stage models - all of color information processing is accomplished by the initial photopic sensory mechanisms. • The data from hue-cancellation experiments demonstrated that a two-stage model is necessary - a first stage for the initial encoding and a second stage to develop spectrallyopponent responses. • More recently, chromatic contrast experiments have indicated the necessity for a third stage which responds to hue without being confounded by brightness. • Additional stages may be required to explain color perception that is not correlated to wavelength (e.g., A Land Mondrian). A Three-stage Color Vision Theory (DeValois & DeValois) • Stage 1 is the absorption of light by three cone photopigments - an elementary requirement for trichromatic vision. DeValois, R & DeValois K. Vis Res 33:1053-1065, 1993 A Three-stage Color Vision Theory (DeValois & DeValois) Stage 2 is the formation of spectrally opponent responses. This model proposes three color opponent mechanisms, one for each cone type, rather than two from the Hering Opponentcolors theory. Because the output is also proportional to intensity, wavelength and intensity can be confused. DeValois, R & DeValois K. Vis Res 33:1053-1065, 1993 Synaptic Interactions in the Retina Second stage mechanisms Feed-forward and feed-back synapses of retinal neurons in the inner and outer plexiform layers provide connections for spatial interactions between cone types. Receptors (R) drive both bipolar (B) and horizontal (H) cells. Bipolar cells terminate on both retinal ganglion (G) and amacrine (A) cells. The receptive fields of retinal ganglion cells are described as center-surround opponent mechanisms, with the center and surround being luminancenonopponent or color-opponent. http://medical-dictionary.thefreedictionary.com/ Color Vision Theories and Models Color vision theories were originally proposed to explain normal trichromacy and the alterations caused by genetic color vision anomalies. Now there are considerable psychophysical and physiological research data to serve as the foundation for color vision models. The original concepts and terminology have been retained Interactions at second and third order must be proportional to the relative absorption of light by the three cone photopigments. http://retina.umh.es/webvision/ Simple models of retinal circuits for Lresponses M-cone L-cone + + M-cone + L-cone output = L-response • The outputs of M- and L-cones are combined by neurons with identical synapse types from both cone types. • The output of the neuron is the linear sum of the responses of the two types of cones. Receptive Field Organization of Non-Opponent RGCs +L • The spatial arrangement of receptive fields of RGC and LGN cells is a centersurround organization of inputs from the various cone types. +M • Monochromatic stimuli produce the characteristic nonopponent (summation) responses, and white light is a very effective stimulus. -M -L M+L>0 M-cone Non-Opponent response L-cone Cone inputs to the L-response The L-response represents the linear, additive combination of the responses of M- and Lcones. SS = M + L at each wavelength. 0 The L-response has the major characteristics of the photopic luminosity function. L-cone M-cone Simple models of neural circuits for +G-R opponent responses The responses of R/G + - opponent cells are similar to the C-response form of Spotentials. The output of the opponent +G-R opponent output neuron is the linear difference of the responses of the two types of cones. 535 nm 565 nm SS = M – L The null point occurs at the M-cone wavelength of equal chromatic sensitivity for the two photopigments. L-cone 520 nm 620 nm 0 Cone inputs to +G-R RGC cells The M - L operation causes the peak response points to be displaced to a wavelength shorter than the M-cone response for the “green” response and to a wavelength longer than the L-cone for the “red” response. Receptive Field Organization of R/G Opponent RGC cells +L • The spatial arrangement of receptive fields of opponent cells is a center-surround organization of antagonistic inputs from M- and L-cone types. -M • Monochromatic stimuli produce the characteristic opponent responses, but white light is not an effective stimulus. +M -L S-cone M-cone L-cone Simple Models of Neural Circuits for +Y-B Opponent Responses The yellow component of the + - + + Y/B opponent cells is the additive combination of the M- and L-cone responses (the luminosity response). The output of the opponent +Y-B opponent output neuron is the linear difference of the responses of S-cone’s output and the sum of the M- and L-cones’ outputs. Receptive Field Organization of Y/B Opponent RGC Cells • The scheme of the spatial +L+M organization of Y/B neurons is similar to the R/G opponent cells except for the difference in the specific cone inputs. -S +S -L-M Models of Neural Circuits for Color Vision S-cone M-cone L-cone • Six distinct response types were found for color processing. + - + - + + Non-opponent output +Y-B opponent output +R-G opponent output • Four types of color opponent neurons. 1) +R-G 2) +G-R 3) +Y-B 4) +B-Y • Two types of nonopponent neurons. 1) Excitatory 2) Inhibitory Non-opponent and Opponent Mechanisms of Color Vision Opponent mechanisms Non-Opponent mechanisms http://web.mit.edu/ Opponent and non-opponent channels are represented in different channels of the afferent visual pathway. • Opponent channels – parvocellular pathway • Non-opponent channels – magnocellular pathway A Three-stage Color Vision Theory (DeValois & DeValois) Stage 3 is the formation of pure chromatic contrast responses, by further processing of the responses from stage 2 spectral opponency – called doubleopponent cells in the visual cortex. Stage 3 responses are still wavelength dependent, but their responses are related, specifically, to chromatic contrast and they are not driven by luminance contrast. DeValois, R & DeValois K. Vis Res 33:1053-1065, 1993 Receptive Field Organization DoubleOpponent Cortical Cells +L-M -M+L Optimal Stimulus The scheme of the spatial organization of double-opponent cells combines opponent cell responses of opposite types in the center and surround areas of the receptive fields. The optimal stimulus has spatial and spatial frequency properties – important for identifying edges of colored objects. Non-Optimal Stimulus Segregation of Cortical Areas for Specialized Visual Functions 1. Color blobs Inter-blobs Image from: read.uconn.edu/PSYC3501/Lecture07/ Schematic representation of the modular segregation of the brain. 2. The largest modules are ocular dominance (OD) columns. 3. Superimposed on the OD organization are the interblobs, blobs, and orientation columns. Color Vision Theories Recap Young-Helmholtz Trichromatic Theory • Perception of hue and brightness Hering’s Opponent Colors Theory • Perception of yellow • Perception of negative afterimages Multi-stage Color Vision Theories • Receptive field organization of opponent and non-opponent channels. Opponent and Non-Opponent Channels • Parvo- and Magno-cellular Pathways Double Opponent cells in V1