Survey
* Your assessment is very important for improving the work of artificial intelligence, which forms the content of this project
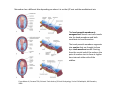
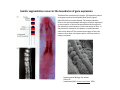
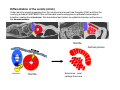
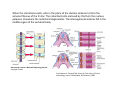
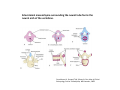
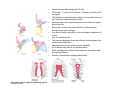
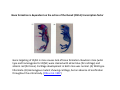
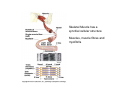
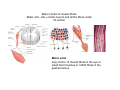
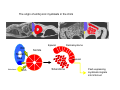
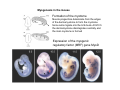
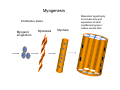
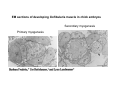
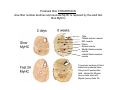
Musculoskeletal Development Objectives • • • • • • • • • Review of the subdivisions of mesoderm development. Differentiation of somites Development of the axial skeleton – skull and vertebral column. Resegmentation of the sclerotome Development of the vertebrae and their specializations. Development of the skull. Cartilage formation Bone formation Development of skeletal muscle Citation: The Developing Human: clinically oriented embryology 9th ed. Keith L. Moore, T.V.N. Persaud, Mark G. Torchia. Philadelphia, PA: Saunders, 2011. Chapter 14 ‐ Skeletal System Chapter 15 ‐ Muscular System Citation: Larsen's human embryology 4th ed. Schoenwolf, Gary C; Larsen, William J, (William James). Philadelphia, PA : Elsevier/Churchill Livingstone, c2009. Chapter 8 ‐ Development of the Musculoskeletal System Mesoderm has a different fate depending on where it is on the A/P axis and the mediolateral axis The head paraxial mesoderm is unsegmented. Neural crest cells invade into the head mesoderm and both contribute to head formation The trunk paraxial mesoderm segments into somites that are thought to form by a clock‐wavefront model. Starting from the cranial end of the embryo, the pairs of somites start to form at regular time intervals either side of the midline. From Moore KL, Persaud TVN, Shiota K: Color Atlas of Clinical Embryology, 2nd ed. Philadelphia, WB Saunders, 2000. The formation of muscle and bone in the trunk region the products of the paraxial>somitic mesoderm Developmental Biology. 6th edition. Gilbert SF. Sunderland (MA): Sinauer Associates; 2000. Somitic segmentation occurs at the boundaries of gene expression Transition from somitomere to somite. (A) Expression pattern of receptor tyrosine kinase EphA4 (blue) and its ligand, ephrinB2 (red) as somites develop. The somite boundary forms at the junction between the region of ephrin expression on the posterior of the last formed somite and the region of Eph expression on the anterior of the next somite to form. In the presomitic mesoderm, the pattern is created anew as each somite buds off. The posteriormost region of the next somite to form does not express ephrin until that somite is ready to separate. Developmental Biology. 6th edition. Gilbert SF. Sunderland (MA): Sinauer Associates; 2000. Differentiation of the somite (chick) Under inductive signals emanating from the notochord and neural tube floorplate (SHH) and from the overlying ectoderm (WNT/BMP4) the ventromedial somite undergoes an epithelial>mesenchymal transition creating the sclerotome. The dorsolateral part retains an epithelial character and becomes the dermamyotome. Somite Dermamyotome Neural tube SHH SHH Notochord Somite Sclerotome – axial cartilage then bone (A) The somites divide into sclerotome cells and dermamyotome cells. (B) The sclerotome cells lose their adhesion to one another and migrate toward the neural tube. (C) The remaining dermamyotome cells divide. The medial cells form an epaxial myotome beneath the dermamyotome, while the lateral cells form a hypaxial myotome. (D) A layer of muscle cell precursors (the myotome) forms beneath the epithelial dermamyotome. Developmental Biology. 6th edition. Gilbert SF. Sunderland (MA): Sinauer Associates; 2000. The cranial‐caudal expression boundary created at initial segmentation is maintained in the sclerotome and correlates with the boundary between a loosely organized cranial region and a cell‐dense caudal region. The division between these domains is called Von Ebner’s fissure. Later the sclerotomes split along this line and they fuse to adjacent sclerotomal segments to create the vertebral bodies. This allows the spinal nerves to pass between the vertebral bodies to contact the muscles developing in the myotomal blocks The sclerotome undergoes a process of resegmentation that involves the fusion of the caudal and cranial regions of successive sclerotomal masses. This gives rise to the vertebral body and neural arch. The making of the somite: molecular events in vertebrate segmentation Yumiko Saga & Hiroyuki Takeda Nature Reviews Genetics 2, 835‐845 (November 2001) The first 5 somites contribute to the occipital bone of the skull. The atlas is formed from the caudal region of the 5th somite and the cranial region of the 6th. This mechanism creates the situation in which there are 8 cervical spinal nerves but only 7 cervical vertebrae. In the thoracic, lumbar and sacral regions, the number of spinal nerves matches the number of vertebrae. When the sclerotome splits, cells in the plane of the division coalesce to form the annulus fibrosus of the IV disc. The notochord cells enclosed by this form the nucleus pulposus. Elsewhere the notochord degenerates. The intersegmental arteries fall in the middle region of the vertebral body. Schoenwolf: Larsen's Human Embryology, 4th ed. Copyright © 2008 From Moore KL, Persaud TVN, Shiota K: Color Atlas of Clinical Embryology, 2nd ed. Philadelphia, WB Saunders, 2000. Sclerotomal mesenchyme surrounding the neural tube forms the neural arch of the vertebrae. From Moore KL, Persaud TVN, Shiota K: Color Atlas of Clinical Embryology, 2nd ed. Philadelphia, WB Saunders, 2000. • • • • • • • • • • • • Schoenwolf: Larsen's Human Embryology, 4th ed. Copyright © 2008 Vertebrae specialize along the A/P axis 33 in total – 7 cervical, 12 thoracic, 5 lumbar, 5 sacral and 5 coccygeal The bodies are represented in yellow, the vertebral arches in red, and the costal processes in blue Mesenchyme of the costal processes in the thoracic region forms the ribs Seven pairs of true ribs attach directly to the sternum through their own cartilage Five pairs of false ribs attach via the cartilage of another rib or ribs Last 2 are floating ribs The sternum develops from sternal bars that emerge in the ventrolateral body wall Chondrification occurs as they move medially At 10 weeks they fuse in the median plane Form cartilaginous models of manubrium, sternebrae and the xiphoid process. Centers of ossification appear before birth Cranium development Can be subdivided into 4 main centres and 2 origins 1. Neurocranium – bones enclosing the brain 2. Viscerocranium – bones of the face derived from pharyngeal arches Cartilaginous neurocranium Forms the base of the brain box from temporary cartilage models a. Parachordal cartilage forms at the cranial end of the notochord and fuses with the occipital sclerotomes b. Hypophysial cartilage forms around the pituitary and forms the sphenoid c. Trabeculae cranii fuse to form the body of the ethmoid d. Ala orbitalis forms the lesser wing of the sphenoid e. Otic capsules form around the otic vesicles and contribute to the temporal bone f. Nasal capsules form around the nasal sacs and contribute to the ethmoid Membranous neurocranium Forms bone directly from the mesenchyme at the sides and top of the brain to form the calvaria (skullcap). Parietal and frontal with 6 fibrous fontanelles > sutures Cartilaginous Viscerocranium Derived from neural crest in the 1st two pharyngeal arches and develops via the intermediate formation of a cartilaginous model a. 1st pharyngeal arch – malleus and incus b. 2nd pharyngeal arch – stapes and styloid process of temporal bone c. 3rd pharyngeal arch – contribute to the hyoid d. 4th pharyngeal arch – laryngeal cartilage Membranous Viscerocranium Membranous ossification in the maxillary prominence of the1st arch – forms the squamous, temporal, maxillary and zygomatic bones The mandible is formed by membranous ossification around a cartilaginous model Formation of cartilage Cartilage development begins during the 5th week at sites of mesenchymal condensation called chondrification centres. Mesenchymal cells differentiate into prechondrocytes then chondoblasts, which secrete an extracellular matrix of ground substance (carbohydrates ‐hyaluronan, chondroitin sulfates and keratan sulfate) and the protein tropocollagen (type II) which polymerizes extracellularly to form collagen fibres. Types of cartilage Hyaline – the basic form Elastic cartilage – has elastic fibres mixed in. forms the epiglottic cartilage, the larynx, external ear and auditory tube Fibrocartilage – contains type I collagen as well as type II collagen. Flexible and tough. Forms the annulus fibrosus of the intervertebral discs and the pubic symphysis. Articular cartilage – lines the joint surfaces Bone formation ‐ Endochondrial Ossification • • • • • • • • Occurs in preexisting cartilaginous models Majority of skeleton formed by this process (vertebra, limb long bones) Osteoblasts replace cartilage matrix with a matrix rich in type I collagen Chondrocytes undergo hypertrophy then apoptosis Blood vessels enter bringing in osteoblasts which deposit bone matrix (osteoid) Ossification centres (primary and secondary) create an epiphyseal cartilaginous growth plate near the ends of long bones Some invading cells differentiate into haematopoietic cells – bone marrow Perichondrium converts into periosteum Developmental Biology. 6th edition. Gilbert SF. Sunderland (MA): Sinauer Associates; 2000. Intramembranous Ossification • • • • • • Forms in mesenchyme that has formed a membranous sheath Mesenchyme condenses and becomes highly vascular Precursor cells differentiate directly into osteoblasts and start to deposit bone matrix (osteoid). Calcium phosphate is deposited in the osteoid and the osteoblasts become trapped within the matrix to form osteocytes. Initially has no pattern – just spicules of bone which then organize into lamellae (layers). Bone van be continuously remodeled though resorption via osteoclasts and new deposition via osteoblasts Developmental Biology. 6th edition. Gilbert SF. Sunderland (MA): Sinauer Associates; 2000. Bone formation is dependent on the action of the Runx2 (Cbfa1) transcription factor Gene targeting of Cbfa1 in mice causes lack of bone formation. Newborn mice (wild‐ type and homozygotes for Cbfa1) were stained with alcian blue (for cartilage) and alizarin red (for bone). Cartilage development in both mice was normal. (A) Wild‐type littermate. (B) Homozygous mutant showing cartilage, but an absence of ossification throughout the entire body. (Otto et al. 1997) Muscle – Myogenesis • Smooth muscle – some cells originate from undifferentiated splanchnic mesoderm mesenchymal cells. Smooth muscle surrounding blood vessels originates from somatic mesoderm. Others (iris of the eye, myoepithelial cells in mammary and sweat glands) originate from ectoderm. Differentiate to express smooth muscle actin and myosin for contraction but remain mononuclear • Skeletal muscle ‐ cells originate from the paraxial mesoderm. Myoblasts undergo frequent divisions and then fuse to form multinucleated, syncytial myotubes that mature to form adult muscle fibres. • Cardiac muscle ‐ cells originate from the prechordal splanchnic mesoderm. Discussed later Skeletal Muscle has a syncitial cellular structure Muscles, muscle fibres and myofibrils Properties of Muscle Fiber Types Characteristic V (speed of shortening) Fast fibers IIb IIa Type I Intermediate Low High/moderate High Predominant energy system Anaerobic Combination Aerobic Myoglobin Low Medium High Capillary density Low Medium High max Resistance to fatigue Highest IIx Slow fibers Low Motor control of muscle fibres Motor unit – the -motor neuron and all the fibres under its control Motor units may control <5 muscle fibres in the eye or small hand muscles or >2000 fibres in the gastrocnemius The origin of embryonic myoblasts in the chick Epaxial Wnt Somite Neural tube Notochord Shh Dermamyotome Hypaxial Sclerotome Pax3-expressing myoblasts migrate into limb-bud Myogenesis in the mouse Formation of the myotome Muscle progenitors delaminate from the edges of the dermamyotome to form the myotome. Some cells migrate into the limb buds. At E10.5 the dermamyotome disintegrates centrally and the main myotome is formed Expression of the myogenic regulatory factor (MRF) gene MyoD Epaxial and hypaxial components of the myotome E11.5 mouse embryos. Eloy-Trinquet S , Nicolas J Development 2002;129:111-122 Myogenesis Proliferative phase Myogenic progenitors specification Myoblasts determination differentiation Myotube maturation growth hypertrophy Maturation hypertrophy to increase size and expression of adult myofilament genes = mature muscle fiber Differentiation of primary myotubes in the mouse hind-limb (12-14 dpc) and the beginning of fibre type patterning Fusion of myoblasts is ordered and synchronous. Nerve is not required for fusion or Myosin Heavy Chain Slow expression Tendon formation from sclerotome-derived cells – marked by expression of Scleraxis (Scx). Induced by the myotome. MyHC expression 1. Embryonic 2. Neonatal 3. Slow Secondary myotube formation – mouse hindlimb 14dpc - birth and continuing fibre type patterning 14-16 dpc - Pioneer motor axons contact primary myotubes. Necessary for survival of myotube and secondary myotube cluster formation Secondary myotubes form in Clusters around primaries. MyHC gene expression 1. Embryonic 2. Neonatal Late fetal stage- clusters disperse. MyHC gene expression Primaries – slow MyHC Secondaries - neonatal MyHC EM sections of developing iliofibularis muscle in chick embryos Secondary myogenesis Primary myogenesis In situ hybridisation analysis of Troponin I isoforms in mouse crural sections G = Gastrocnemius S = Soleus E = EDL T = Anterior tibialis Tnni1 is the gene that encodes the inhibitory subunit of the Troponin complex that is found in slow-twitch fibres. Postnatal fibre CONVERSION: slow fiber number declines and neonatal MyHC is replaced by the adult fast fibre MyHCs A Tibia Tibialis anterior muscle EDL muscle Fibula Soleus muscle Medial Gastrocnemius muscle Lateral Gastrocnemius muscle Transverse sections of hindlimbs from postnatal mice 2days and 6 weeks after birth – stained for Myosin heavy chain slow and Myosin heavy chain 2A Plasticity and Regeneration of Adult Muscle Muscle Adaptation to Exercise Training Adaptations to exercise training, particularly elevation in oxidative capacity of exercised muscle but also some myosin isoform changes mainly in fast subtypes. Cross‐Reinnervation Buller et al. (1960) – Motor nerves supplying the (slow) soleus and (fast) FDL muscles swapped around. Contraction speed of soleus got faster, FDL slower. Chronic Low‐Frequency Stimulation (CLFS) Artificial electrical stimulation of a nerve supplying a fast muscle with a tonic pattern mimics the impulse pattern of a slow nerve and induces fast to slow transformation Pette et al. (1973). Regeneration Injured muscle can regenerate itself using a population of stem cells that are laid down during embryogenesis – called satellite cells. Satellite cells lie between the sarcolemma and the basal lamina of each muscle fibre and activated by injury. MATURATION HYPERTROPHY that occurs in mouse SOLEUS muscle fibres between birth and adulthood Images taken at same magnification – HIGH power BIRTH ADULT Muscle hypertrophy and hyperplasia – exercise induced hypertrophy and genetic control of hyperplasia via Myostatin. Mutation of this gene causes double muscling in the Belgian Blue breed.