Survey
* Your assessment is very important for improving the work of artificial intelligence, which forms the content of this project
Transgenic Research 7, 105±112 (1998)
Maternally expressed PGK-Cre transgene as a tool for
early and uniform activation of the Cre site-specific
recombinase
Y VA N L A L L E M A N D { } , V I C TO R L U R I A } , R E B E C C A H A F F N E R K R AU S Z and P E T E R L O NA I Department of Molecular Genetics, The Weizmann Institute of Science, Rehovot 76100, Israel (Fax: 972 8
934 4108)
Received 16 June 1997; revised 8 September 1997; accepted 14 October 1997
A transgenic mouse strain with early and uniform expression of the Cre site-specific recombinase is described. In
this strain, PGK-Crem , Cre is driven by the early acting PGK-1 promoter, but, probably due to cis effects at the
integration site, the recombinase is under dominant maternal control. When Cre is transmitted by PGK-Crem
females mated to males that carry a reporter transgene flanked by loxP sites, even offspring that do not inherit
PGK-Cre delete the target gene. It follows that in the PGK-Crem female Cre activity commences in the diploid
phase of oogenesis. In PGK-Crem crosses complete recombination was observed in all organs, including testis and
ovary. We prepared a mouse stock that is homozygous for PGK-Crem and at the albino (c) locus. This strain will
be useful for the early and uniform induction of ectopic and dominant negative mutations, for the in vivo removal
of selective elements from targeted mutations and in connection with the manipulation of targeted loci in `knock
in' and related technologies.
Keywords: Cre recombinase; maternal expression; gametogenesis; unifrorm Cre activity; early development
Introduction
The Cre-loxP system is becoming an important tool of
modern mouse genetics. Cre, a recombinase of the P1
bacteriophage, catalyses DNA recombination at loxP
recognition sites that can be introduced at will to specific
locations of the mammalian genome. Cre transferred in
vitro by transfection, or in vivo by genetic crosses, excises
genome fragments flanked by two loxP sites with great
efficiency (Sauer and Henderson, 1988). This principle
has been successfully used to activate `dormant' transgenes (Lakso et al., 1992), for cell- and tissue-specific
transgene expression (Orban et al., 1992), for site-specific
activation of targeted mutations (Gu et al., 1994), for the
exchange of gene sequences by the `knock in' method
(Hanks et al., 1995), for the introduction of point
To whom correspondence should be addressed.
{
Present address: Unite d'Embryologie MoleÂculaire, Institut Pasteur, 25, rue
du Dr Roux, Paris, France.
}
These two authors contributed equally to this study.
0962±8819 # 1998 Chapman & Hall
mutations (Gu et al., 1993), as well as to create major
genome changes, including chromosomal rearrangements
(Ramirez-Solis et al., 1995).
The primary application of Cre is in cell- and tissuespecific gene targeting, where the recombinase is placed
under the control of specific promoter elements and the
recombination induced mutation is restricted to the
promoter's spatio-temporal domain. Several important
molecular problems, however, require early and ubiquitous Cre activity. Ubiquitous Cre activity can be generated by promoters that are active either in the zygote, or
in the oocyte, as a result of maternal activation. Such
transgenic Cre lines could be used to activate dominant
mutations represented as dormant transgenes (Lakso et
al., 1992) and to investigate events that take place during
the earliest stages of embryogenesis. Lines with ubiquitous Cre activity will be also useful to process transgenic
or targeted mutations by genetic crosses instead of in
vitro transfection. This is an advantage, because protracted culture endangers the pluripotentiality of recombinant ES cells and decreases germline transmission.
106
Transgenic animals that express Cre in all lineages in a
uniform manner can be also employed to remove `floxed'
selection cassettes, such as the neo gene that may
influence the transcription of neighboring loci and
interfere with the targeted phenotype (Olson et al., 1996).
Cre-transgenic mouse lines that are active in the early
zygote have already been reported. Schwenk et al. (1995)
placed Cre under the control of the human cytomegalovirus minimal promoter, whereas Lakso et al. (1996)
used the EIIa promoter of adenovirus. More recently,
Lewandoski et al. (1997) reported a Zp3-Cre transgenic
strain, which allows the investigation of developmental
events that take place during the later stages of
oogenesis. We prepared transgenic mouse lines carrying
Cre controlled by the PGK-1 promoter (Adra et al.,
1987). Here we report the establishment and analysis of a
unique PGK-Cre mouse strain that, in females, expresses
Cre as early as the diploid primordial germ cell and
displays early and ubiquitous expression. The use of this
strain in the study of early development will be
discussed.
Materials and methods
Mouse strains
(BALB/c 3 C57B1/B6)F1 female mice were superovulated
at 3±4 weeks of age (Hogan et al., 1994) and mated to
(BALB/c 3 C57B1/B6)F1 males to generate one-cell
embryos. Pseudopregnant MF1 females served as recipients for embryo transfer.
Plasmids for microinjection
As a first step to provide a polyadenylation signal to the
different transgenes the pSKTnlac Z plasmid (Tajbakhsh et
al., 1996) was modified by eliminating the Eco RI-Lac Z
fragment to obtain pPA. pPA contains only, in the multiple
cloning site of Bluescript, a small, inactive Eco RI-Xba I
59 fragment of lac Z and the Xba I-Bam HI fragment
containing the SV40 polyadenylation signal.
The Cre coding sequence was obtained by PCR, using
the pMC-Cre (Gu et al., 1993) as template. The primers
used were: GCAAGCTTTCGACCATGCCCAAGAAGAAG (59 sense) and GCGAATTCCGTTAATGGCTAATCGCCATCn (39 antisense). They contained a Hind III
(59) and a Eco RI (39) site (underlined), respectively,
which allowed the direct cloning of the Cre fragment into
the pPA plasmid. The Cre sequence was confirmed by
sequencing. The PGK-1 promoter (Adra et al., 1987) was
introduced 59 to the Cre gene to obtain the definitive
construct pPGK-Cre.
Our GLD reporter strain is derived from the
pSKTnlac Z plasmid (Tajbakhsh et al., 1996). A loxP
site, derived from the pGZM30 plasmid (a gift from Dr
K. Rajewsky, Cologne), was ligated 39 to the SV40
Lallemand et al.
polyA-signal and a second loxP site, in the same
transcriptional orientation, was inserted 59 to lac Z. To
the 59 end of this construct the PGK-1 promoter was
ligated to obtain the pYS3 plasmid.
A Not I-Eco RI genomic fragment of the murine Dlx2
gene, containing its complete coding sequence, was
derived from a phage kindly provided by E. Boncinelli
(Milano). This fragment was cloned into the Sal I (blunt)Eco RI sites of pUC19. The Not I site was placed at
about 380 bp 59 to the ATG initiating codon of Dlx2,
whereas Eco RI was placed 537 bp downstream to the
TAA stop codon (Porteus et al., 1991). The Hind IIIEco RI fragment of pUC-Dlx2 was then cloned in the
Hind III-Eco RI sites of pPA to give the pDlx-PA plasmid.
The Sal I-Not I fragment (Dlx2 + polyA) was cloned into
the Xho I-Not I sites of pYS3, downstream to the more 59
loxP site to give the pYSD2 plasmid. The Xho I-Xho I
fragment of the 59 non-coding sequence (Porteus et al.,
1991) was eliminated by cutting with Xho I and
religating, to obtain the definitive construct pGLD.
Generation of transgenic mice
For microinjection, pGLD was cut by Xmn I and Not I.
pPGK-Cre was cut by Xho I and Not I. The DNA
fragments were isolated by electrophoresis in 1% agarose,
then electroeluted and concentrated using Elutip-D
columns (Shleicher & Schuell). The DNA was diluted to
a final concentration of 2 ìg mlÿ1 in injection buffer and
microinjected as described (Hogan et al., 1994).
Results
Detection of PGK-Cre activity
Transgenic mice were produced carrying the pPGK-Cre
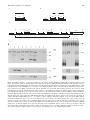
construct (Fig. 1A). The transgene was detected by
Southern blotting as a 1.3 kb Eco RI fragment hybridizing
to the Cre probe (Fig. 1D). Three of five PGK-Cre
transgenic lines displayed Cre activity. They deleted Lac Z
in crosses with a reporter strain that carries the PGK-loxPlac Z-loxP-Dlx2 transgene designated as GLD (Fig. 1B).
This transgenic line was devised as a `dormant' transgene
where Dlx-2 (Porteus et al., 1991) could be brought under
the control of the PGK-1 promoter by deleting the
intermediate floxed lac Z (Fig. 1B). In the GLD strain
however, Dlx2 was truncated and inactive, hence it was
used only as a reporter of Cre activity.
Cre-mediated recombination was detected in embryos
of GLD 3 PGK-Cre crosses as the loss of a 3.0 kb
Eco RI fragment through the deletion of lac Z. Deletion
of this fragment, in embryos that inherited Cre via their
PGK-Crem transgenic father, is shown in Fig. 1D.
Hybridization with Dlx-2 detected three Eco RI fragments. A large fragment (, 20 kb) represented the wildtype locus, a 2.7 kb multiple-copy fragment representing
Maternally regulated Cre transgene
107
A
B
E
pgk
E
E
1.3
pgk
Cre
3.0
E
E
LacZ
loxP
2.7
E
Dlx2
loxP
C
E
E
E
2.7
E
E
E
E
2.7
E
E
E
E
5.6
E
Fig. 1. The PGK-Cre transgene, preparation and detection of its activity. (A) The pPGK-Cre plasmid. PGK: Eco RI-Taq I fragment of the
mouse pgk1 promoter (Adra et al. 1987). Hatched box: SV40 pA site. (B) The pGLD plasmid. Dark hatching: lac Z. Triangles: loxP sites.
Light hatching: SV40 polyadenylation site. Heavy lines above Fig. 1A and B: DNA probes. (C) Organization of the GLD transgene, and its
rearrangement in crosses with PGK-Cre. A concatenate of three GLD monomers is shown. As in Fig. 1B, the unshaded box represents the
pgk-1 promoter, heavy shading represents lac Z and the unshaded box, joined with light hatching represents Dlx-2 with the SV-40
polyadenylation site and the arrow-heads represent the loxP sites. A two-headed arrow brackets the deletion and the broken unshaded box
represents the truncated Dlx-2 gene at the end of the concatenate. (D) Southern blot analysis of GLD 3 PGK-Cre embryos. Embryonic
DNA was digested with Eco RI. The three panels of both autoradiogramms correspond to individual Southern blots hybridized to three
different probes. The upper panels show hybridization with Dlx2, the middle panels with Cre and the lower with lac Z. Left picture: In
embryos that did not inherit GLD (lanes 1, 3, 5, 8 and 9), only the wild-type, 20 kb Dlx2 fragment can be detected. In GLD embryos,
which did not inherit Cre, (lanes 4, 6 and 7) two other bands are present. They correspond to multiple Dlx2 copies (2.7 kb) and to the
truncated 39 single copy (5.6 kb). In the recombinant double transgenic embryo (lane 2) only the truncated (5.6 kb) Dlx-2 copy is present,
whereas the internal copies (2.7 kb) are deleted. Recombination is confirmed by the absence of the lac Z signal in the double recombinant
embryo (lane 2), whereas multiple lac Z copies were detectable in non-recombinants (lanes 4, 6 and 7). Picture on the right of Fig. 1D
shows characteristic hybridization patterns from a number of experiments. Lanes 1 & 2, GLD 3 PGK-Cre, that did not inherit Cre; lanes 3
& 4 double transgenic, complete deletion of Lac Z; lanes 5 & 6 wild-type; lanes 7 & 8 GLD transgenic DNA.
108
a concatenate formed by the transgene and a third 5.6 kb
fragment representing the 39-most integration fragment
(Fig. 1D, lane 7). In the presence of Cre the concatenate
was entirely deleted, but the 5.6 kb Dlx-2 fragment
remained unchanged, even in the recombinant (Fig. 1D,
lane 2). These results and our additional restriction
analysis (not shown) revealed that Cre deletes all
elements of the concatenate, save the most 39 copy,
which is truncated and whose remnant form is detected
as a 5.6 kb single copy Eco RI fragment, as is shown
schematically in Fig. 1C.
Recombinase activity in various PGK-Cre 3 GLD crosses
The above results were obtained with GLD females mated
to PGK-Crem males. In the reciprocal cross, when PGKCrem females were mated to GLD males, an unexpected
observation was made. In this cross, lac Z was deleted in
all resultant embryos irrespective whether or not they
inherited the Cre transgene from their mother (Fig. 2A,
lane 4 and Fig. 2B, lane 4 and 5). This result was repeated
in numerous similar crosses and we found that the Cre
and Creÿ offspring displayed equal ratios of recombina-
Lallemand et al.
tion (Table 1). Significantly all Cre individuals displayed
complete recombination and 31 of the 33 Creÿ offspring
showed complete (and only two showed incomplete)
recombination. This suggested that the single Cre
transgenic allele of the mother must have been active
already in the diploid oogonium and during meiosis the
Cre transcript or protein was transmitted to both the Cre
and Creÿ daughter cells.
The experiments described above (Table 1, first two
lines) were performed with the offspring of the PGKCrem founder, or with its first generation offspring. Next
we investigated whether maternal behaviour is stably
transmitted. Table 2 demonstrates that embryos of the
third generation also display maternal regulation. It
follows that the characteristic regulation of PGK-Crem
is a heritable trait. Here again recombination was
obtained in a close to 1:1 ratio in the Cre and Creÿ
offspring. We have evidence also for germline transmission of the Cre-induced rearrangement of the GLD
reporter gene (data not shown).
PGK-1 is known to be expressed during early
development (McBurney et al., 1994). In the absence
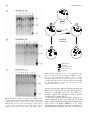
Fig. 2. Maternal regulation of recombination in a PGK-Crem GLD cross. A. PGK-Crem females mated to GLD males (lanes 1 to 7) are
compared to a control litter (lanes 8 to 11 in panel B) from a wild-type female crossed with the same male. Embryos in lanes 8 and 10 are
not transgenic whereas embryos 9 and 11 are non-recombinant pGLD transgenics. In the litter from the pGK-Cre female two embryos are
pGLD transgenics (lanes 4 and 6), both are recombinant although embryo number 4 does not carry Cre. B. Characteristic hybridization
patterns from an number of crosses. Lane 1 is wild-type, lane 3 & 4 are Cre double recombinants, whereas lane 4 & 5 are double
recombinants, that did not inherit Cre. Lane 6 is a GLD transgenic.
Maternally regulated Cre transgene
109
Table 1. Analysis of three PGK-Cre transgenic strains
Recombinants a
Genotypes
Cross
Total
GLD
GLD ;Cre
GLD ;Cre ÿ
Cre
Cre ÿ
Crem 3 GLD
GLD 3 Crem
Cref 3 GLD
GLD 3 Crefl
Creg 3 GLD
GLD 3 Creg
133
78
29
15
18
15
63
48
11
7
8
6
30
24
8
5
7
0
33
24
3
2
1
6
30
23
8
2
3
0
31
0
0
0
0
0
Only complete recombination is shown in the table. Recombination was observed in all Cre
animals and in the Creÿ offspring of Crem females.
Table 2. Transmission of the maternal effect
Genotypes
Recombinants
Cross a
Total
GLD
GLD, Cre
GLD , Creÿ
Cre
Creÿ
Crem 3 GLD
GLD 3 Crem
49
19
21
15
12
15
9
0
12
15
9
0
a
3d generation PGK-Cre transgenic mice mated to the GLD reporter strain.
of contrary evidence, it was therefore possible that the
observed maternal effect could be a characteristic of the
PGK-1 promoter. An alternative possibility suggested that
PGK-Crem had integrated to a site under maternal
control. To test these hypothesis two other PGK-Cre
transgenic lines were investigated. Neither line showed
evidence of maternal activation. Only Cre offspring in
these lines carried a rearranged GLD transgene (Table 1,
lines 3±6).
To compare maternal and paternal inheritance in the
PGK-Crem line we investigated whether the Cre gene
product is transmitted by PGK-Crem males. To this end
GLD females were mated to PGK-Crem males. Repeated
experiments revealed that PGK-Crem males do not
transmit Cre activity to their Creÿ offspring, whereas
their Cre offspring displayed complete recombination
(Fig. 1D, lane 2 and Table 1, line 2). This suggested that
the Cre product is not retained during spermatogenesis,
although paternal Cre was transmitted and activated in
the zygote.
PGK-Cre mediated recombination is ubiquitous
We assumed that if PGK-Crem is active during oogenesis,
then in a cross with a transgene carrying loxP recognition
sites all cells should delete its `floxed' elements. To
investigate this possibility, a variety of tissues from both
Cre and Creÿ mice, derived from a PGK-Crem 3 GLD
cross (maternally transmitted) were subjected to Southern
analysis (Fig. 3AB) to test for the presence of the
rearranged transgene. A similar analysis was performed
with the reciprocal cross (see Fig. 3C) to study Cre
activity transmitted from the male parent. Results of all
three experiments showed that Cre-mediated recombination is complete in all tissues, including ovary and testis.
From these results we conclude that the PGK-Crem
transgenic strain fulfills stringent criteria of uniform and
early Cre activation. To make this genotype easy to
maintain, we bred the stock to homozygosity on the
albino, MFl, non-inbred background.
Discussion
This study describes a transgenic mouse strain that
expresses the Cre site specific recombinase under
maternal control. The strain produces site specific
recombination in most embryonic and adult cell lineages
when crossed to transgenes flanked by loxP recognition
sites.
Maternal transmission in the PGK-Crem 3 GLD cross
was demonstrated by Cre induced recombination of the
GLD transgene transmitted by the father. Recombination
took place with equal frequency in the Cre and the Creÿ
offspring of this cross. It follows that the recombinase was
already active in the early, diploid phase of oogenesis. We
assume that the diploid oocyte precursor transmitted its
single transgenic allele to every other daughter cell,
whereas the Cre gene product was transmitted, through the
cytoplasm, to both daughter cells. Whether Cre was
activated in primordial germ cells and segregated during
their mitotic division, or that it was activated in the
110
Lallemand et al.
2n
Meiosis
n
n
Fertilization
n
n
PGK-Cre gene
Cre protein
unrecombined GLD
recombined GLD
polar body
Fig. 4. Maternal transmission of Cre. The recombination takes
place in all zygotes derived from hemizygous PGK-Crem females.
Zygote on the right inherited PGK-Cre from its mother;
recombined GLD in the male pronucleus. Zygote on the left also
recombines the GLD transgene, although it did not inherit PGKCre. For abbreiviations see legend. The black squares in the
cytoplasm represent Cre activity.
Fig. 3. Uniform Cre induced recombination in multiple organs.
Southern analysis of tissue DNA hybridized with Dlx-2. (A) Lanes:
1, gut; 2 heart; 3, kidney; 4, liver; 4, spleen; 6, tail; 7, brain; 8, tail,
GLD. (B) Lane 1, brain; 2, ear; 4, heart; 4, kidney; 5, liver; 6, spleen;
7, tail; 8, ovary; 9, tail, GLD. (C) Lane 1, brain, 2 gut; 3, heart; 4,
kidney; 5, liver; 6, tail; 7, testis; 8, spleen; 9 tail, GLD as control.
oogonia and segregated during the first meiosis (Karp and
Berril, 1981) is however not clear. The equal frequency of
transgenic recombinants with Cre and with Creÿ
genotype, suggests that segregation occurs at meiosis.
Our hypothesis is shown in Fig. 4.
It has been reported that the PGK-1 promoter can
activate transgenes in the 3.5 day blastocyst and in the
adult it expresses the reporter ubiquitously, although not at
equal levels in all tissues (McBurney et al., 1994).
Nevertheless, no maternal Cre activity was observed in
two other PGK-Cre transgenic lines. We can suggest two
Maternally regulated Cre transgene
related alternative explanations for this phenomenon. It is
possible that the PGK-Crem transgene is controlled by a
maternal regulator in the vicinity of its integration site.
Alternatively the transgene may have induced a factor that
is responsible for high levels, or extended half life of Cre.
Whether Cre is transiently expressed during spermatogenesis in the paternal lineage remains to be determined.
Our results demonstrate that PGK-Crem males transmit
site-specific recombination only to their Cre offspring.
It is possible that, if the gene is expressed in male
primordial germ cells, its product is removed during
sperm head condensation (Fawcett, 1975). The contrasting behaviour of PGK-Crem in the maternal and paternal
lineages is likely to be connected to the mechanism that
transmits maternal information to the early embryo. A
detailed analysis of PGK-Crem transmission may help the
molecular analysis of mammalian gametogenesis and
maternal transmission of developmental genes.
Gene targeting will benefit from strains that express
Cre early and cause uniform site-specific recombination.
Two such strains have been reported previously. Lakso et
al. (1996) showed that EIIa-Cre is uniformly expressed in
multiple organs, although a considerable degree of
chimaerism was observed. A similar Cre strain was
obtained by Schwenk et al. (1995), who used the hCMV
promoter. Their transgene is localized on the X-chromosome. Because no mosaicism was found, these authors
suggested that recombination should have occurred before
X-inactivation, which takes place between 4.5 and 6 days
p.c. in the mouse (for review, see Chapman, 1986), that
is, during an interval when the embryonic ectoderm
numbers between 32 to 700 cells (Snow, 1977). Our
PGK-Crem strain causes complete recombination in most
cases and in multiple organs, including testis and ovary.
It, moreover, has the characteristic of dominant maternal
regulation, which places the activation of Cre earlier, to a
period before the first meiotic division: hence recombination in the zygote could take place as early as the
pronuclear fusion.
Another transgenic strain, which is active in the diploid
female gamete, was described by Lewandoski et al.
(1997). They used the promoter of Zp3, a major
structural gene of the zona pellucida, to drive the Cre
recombinase. Zp3-Cre is expressed exclusively during
oogenesis, hence this strain can be used to activate
`floxed' target genes in the maternal germ line to
investigate their maternal regulation. Lewandoski et al.
report that enough Cre activity remains in the mature
oocyte to recombine paternally transmitted transgenes of
the zygote. They however report considerable mosaicism
at this stage. It follows that our PGK-Crem strain, with its
high degree of uniform recombination and early expression, maybe more suitable for the manipulation of the
early zygote. A capability to manipulate the mouse
genome between the one cell stage and the early post
111
implantation eggcylinder stage is important because this
early period of mammalian embrygenesis is poorly
understood.
Acknowledgements
This study was supported by a grant from the Israel
Science Foundation and by the Infrastructure Laboratory
Program of the Ministry of Science and Arts. Y.L. was
supported by a long term EMBO fellowship.
References
Adra, C.N., Boer, P.H. and McBurney, M.W. (1987) Cloning and
expression of the mouse pgk-1 gene and the nucleotide
sequence of its promoter. Gene 65, 65±75.
Fawcett, D.W. (1975) The mammalian spermatozoon. Dev. Biol. 44,
394±436.
Grant, S.G. and Chapman, V.M. (1988) Mechanisms of Xchromosome regulation Ann Rev Genet 22, 199±233.
Gu, H., Zou, Y.R. and Rajewsky, K. (1993) Independent control of
immunoglobulin switch region recombination at individual
switch regions evidenced through Cre-loxP mediated gene
targeting. Cell 73, 1155±64.
Gu, H., Marth, J.H., Orban, P.C., Mossmann, H. and Rajewsky, K.
(1994) Deletion of a polymerase beta gene segment in T cells
using cell type specific gene targeting. Science 265, 26±8.
Hanks, M., Wurst, W., Anson-Cartwright, L., Auerbach, A.B. and
Joyner, A.L. (1995) Rescue of the En-1 mutant phenotype by
replacement of En-1 with En-2. Science 269, 679±82.
Hogan, B., Beddington, R., Costantini, F., Lacy, E. (1994)
Manipulating the Mouse Embryo: A Laboratory Manual. Cold
Spring Harbor, NY: Cold Spring Harbor Laboratory.
Karp, G. and Berril, N.J. (1981). In: Development. New York,
USA: McGraw-Hill, pp. 116±38.
Lakso, M., Sauer, B., Mosinger, B.J., Lee, E.J., Manning, R.W., Yu,
S.H., Mulder, K.L. and Westphal, H. (1992) Targeted
oncogene activation by site-specific recombination in transgenic mice. Proc. Natl Acad. Sci. USA 89, 6232±6.
Lakso, M., Pichel, J.G., Gorman, J.R., Sauer, B., Okamoto, Y., Lee,
E.J., Alt, F.W. and Westphal, H. (1996) Efficient in vivo
manipulation of mouse genomic sequences at the zygote
stage. Proc. Natl Acad. Sci. USA 93, 5860±5.
Lewandoski, M., Wassarman, K.M. and Martin, G.R. (1997) Zp3cre, a transgenic mouse line for the activation or inactivation
of LoxP-flanked target genes specifically in the female germ
line. Curr. Biol. 7, 148±51.
McBurney, M.W., Staines, W.A., Boekelheide, K., Parry, D.,
Jardine, K. and Pickavance, L. (1994) Murine PGK-1
promoter drives widespread but not uniform expression in
transgenic mice. Dev. Dynamics 200, 278±93.
Olson, E.L., Arnold, H.H., Rigby, P.W.J. and Wold, B.J. (1996)
Know your neighbours: three phenotypes in null mutants of
the myogenic bHLH gene MRF4. Cell 85, 1±4.
Orban, P.C., Chui, D. and Marth, J.D. (1992) Tissue- and sitespecific DNA recombination in transgenic mice. Proc. Natl
Acad. Sci. USA 89, 6861±5.
112
Porteus, M.H., Bulfone, A., Ciaranello, R.D. and Rubenstein, J.L.R.
(1991) Isolation of a cDNA clone encoding a homeodomain
that is developmentally regulated in the ventral forebrain.
Neuron 7, 221±9.
Ramirez-Solis, R., Liu, P. and Bradley, A. (1995) Chromosome
engineering in mice. Nature 381, 720±4.
Sauer, B., and Henderson, N. (1988) Site-specific DNA recombination: in mammalian cells by the Cre recombinase of
bacteriophage P1. Proc. Natl Acad. Sci. USA 85, 5166±70.
Lallemand et al.
Schwenk, F., Baron, U. and Rajewsky, K. (1995) A cre-transgenic
mouse strain for the ubiquitous deletion of loxP-flanked gene
segments including deletion in germ cells. Nucl. Acids Res.
23, 5080±1.
Snow, M.H.L. (1977) Gastrulation in the mouse: growth and
regionalization of the epiblast. J. Embryol. Exp. Morphol 42,
293±303.
Tajbakhsh, S., Bober, E., Babinet, C., Pourin, S., Arnold, H. and
Buckingham, M. (1996) Dev. Dyn. 206, 291±300.