Survey
* Your assessment is very important for improving the workof artificial intelligence, which forms the content of this project
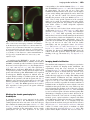
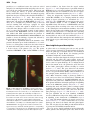
Journal of Experimental Botany, Vol. 62, No. 5, pp. 1651–1658, 2011 doi:10.1093/jxb/erq305 Advance Access publication 15 October, 2010 REVIEW PAPER Imaging fertilization in flowering plants, not so abominable after all Frédéric Berger1,2,* 1 2 Temasek LifeScience Laboratory, 1 Research Link, National University of Singapore, Singapore 117604 Department of Biological Sciences, 14 Science Drive 4, National University of Singapore, Singapore 117543 * E-mail: [email protected] Received 2 August 2010; Revised 7 September 2010; Accepted 9 September 2010 Abstract Although the discovery of double fertilization in flowering plants took place at the end of the nineteenth century little progress had been made in understanding the cellular and molecular mechanisms involved until the end of the twentieth century. After attempts to study fertilization with isolated male and female gametes, researchers turned to Arabidopsis thaliana as a model for genetic analysis and in vivo imaging. The development of confocal imaging and fluorescent proteins, coupled with new molecular insights into cell fate specification of plant gametes, allowed the development of robust markers for cells participating in double fertilization. These markers enabled the imaging of double fertilization in vivo in Arabidopsis. These studies have been coupled with the identification and molecular characterization of genes controlling fertilization in Arabidopsis. Live imaging has already provided new insights on sperm cell delivery, the equivalence of the fate of the sperm cells, gamete fusion, and re-initiation of the zygotic life. This review covers these topics and outlines many important aspects of double fertilization that remain unknown. Key words: Arabidopsis, fertilization, gametes, imaging. Introduction Microscopic descriptions of fertilization in flowering plants marked the end of the nineteenth century (Guignard, 1899; Nawaschin, 1898). Two parallel ‘fertilization events’ are required to produce a viable embryo, a trait unique to flowering plants. Hence the term ‘double-fertilization’ is used to describe the sexual reproductive process in flowering plants. Double-fertilization involves two sperm cells delivered by the pollen tube. The discharge of the sperm cells by the pollen tube is triggered at the entrance of the embryo sac, which contains the egg cell and the central cell, usually referred to as female gametes. The embryo sac is the haploid female gametophyte that also contains three antipodals of unknown function and two synergids involved in pollen tube guidance and in sperm cell discharge. Fertilization of the egg cell produces a zygote, which further develops as the plant embryo. Fertilization of the central cell triggers the development of the endosperm, which nurtures embryo development but does not participate in the next plant generation. The history of the discovery of double-fertilization in angiosperms has been reviewed (Faure and Dumas, 2001; Friedman, 2001) and this review will focus on the experimental investigations of doublefertilization using live imaging. Gametogenesis, pollen tube growth, and pollen tube attraction precede the sperm cells discharge that initiates double-fertilization and these events are not reviewed here. Isolation of the male and female gametes from maize (Zea mays) allowed imaging of gamete fusion in vitro (Kranz et al., 1991; Faure et al., 1994; Kranz and Lörz, 1994; Kranz and Dresselhaus, 1996) and recording of early electrical events (membrane potential and calcium signal) that are triggered in the egg cell by fusion with a sperm cell (Antoine et al., 2000; Digonnet et al., 1997). However, these early studies used plant gametes isolated before full maturity and took place outside of the physiological context (in vitro fertilization was performed in liquid medium). Thus researchers attempted to achieve live imaging of fertilization in planta. Torenia fournieri produces ‘naked’ embryo sacs devoid of ovule integuments at the ª The Author [2010]. Published by Oxford University Press [on behalf of the Society for Experimental Biology]. All rights reserved. For Permissions, please e-mail: [email protected] 1652 | Berger micropylar pole where the pollen tube delivers the sperm cells. An experimental set-up including isolated embryo sacs on a medium favourable for in vitro pollen tube germination allowed the recording of the release of the pollen tube content (including sperm cells and cytoplasm) and of the first series of syncytial nuclei division in the endosperm after fertilization of the central cell (Higashiyama et al., 1997, 2000). However, DIC microscopy applied to the imaging of fertilization in isolated Torenia fournieri ovules did not allow clear images of the fusion of the sperm cells with the egg or the central cell and subsequent karyogamy to be obtained (gamete nuclei fusion). New confocal microscopy protocols adapted to the Arabidopsis female gametophyte (Christensen et al., 1997) led to the first detailed description of double-fertilization including the documentation of karyogamy in this species (Faure et al., 2002). These studies performed with fixed material indicated the time-course of the events taking place during double-fertilization and paved the way for further studies in vivo. With the identification of promoters that express fluorescent markers in male and female gametes it became possible to image in vivo double-fertilization in Arabidopsis. These recent research developments are reviewed here together with the results of combining in vivo imaging with genetic studies. Although significant progress has been made, new tools are required to assess how sperm cells are discharged and fuse, the timing of cell cycle events, how a second fertilization is prevented, how gamete nuclei migration take place before karyogamy, the details of mechanisms leading to karyogamy, and the events leading to reactivation of the zygotic life. Marking Arabidopsis male gametes The discovery of fluorescent proteins from marine invertebrates and their derivative has been essential for research in developmental and cell biology (Chalfie et al., 1994; Tsien, 1998; Haseloff, 1999; Campbell et al., 2002; Giepmans et al., 2006). Identification of suitable promoters enabling a robust and specific expression in plant gametes or associated cells was achieved in recent years as a result of a large community effort (Table 1). The isolation of sperm cells from different plant species provided access to the transcriptome of this cell type. cDNA and EST sequences have been reported from generative cells of Lilium longiflorum (Okada et al., 2006), and from sperm cells of Nicotiana tabacum (Weterings et al., 1992) Zea mays (Engel et al., 2003, 2005) and Plumbago zeylanica (Gou et al., 2009) and, eventually, an Arabidopsis thaliana sperm cell transcriptome was obtained (Borges et al., 2008). The identification of the promoter LAT52 from tomato pollen was the first step towards imaging the discharge of the pollen tube during plant fertilization (Twell et al., 1991). In Arabidopsis the promoter LAT52 drives expression in the microspores before the first pollen division and later in the vegetative cell (Eady et al., 1994). After the microspore unequal division, pLAT52-GFP expression becomes confined to the vegetative cell (Eady et al., 1994) (Fig. 1A). In mature pollen pLAT52-GFP expression is very high in the mature pollen vegetative cell (Cheung, 2001) (Fig. 1B). When pollen germinates and produces a pollen tube the bright fluorescence from GFP expression driven by pLAT52 enables observation of pollen tube growth and gamete delivery in vivo (Rotman et al., 2003; Feijo and Moreno, 2004). Identification of the first gene to be expressed only in sperm cells originated from studies of the male gametophytic mutant duo1 that produces a single sperm cell (Durbarry et al., 2005). This led to the identification of the gene DUO1 that is expressed only in the male germ lineage in Arabidopsis (Rotman et al., 2005). DUO1 encodes a MYB domain protein, which activates the expression of genes involved in cell cycle control and sperm cell fate specification (Rotman et al., 2005; Brownfield et al., 2009). Studies of genes expressed in lily pollen led to the identification of the Arabidopsis gene GENERATIVE CELL SPECIFIC 1 (GCS1) (Mori et al., 2006) (also named HAPLESS2; von Besser et al., 2006), which is expressed only in the sperm cell lineage including the generative cell and sperm cells (Brownfield et al., 2009). Although DUO1 and GCS1 promoters drive the expression of fluorescent markers in the sperm cells, the transgenic lines expressing fluorescent proteins under the control of these promoters did not provide sufficient signal to perform live imaging of fertilization. Table 1. Genes used for live imaging of double-fertilization Gene name AGI ID number HTR10/AtGMH3 AT1G19890 LAT52 Promoter derived from Tomato LAT52 promoter EC1 AT1G76750 FWA AT4G25530 FIE AT3G20740 MYB98 CENH3 ATLIGASE1 AT4G18770 AT1G0130 AT1G08130 Cell type labelled Step of fertilization studied Sperm cells Pollen vegetative cell and pollen tube Egg cell Central cell Egg cell and central cell (non-specific) Synergids All All Sperm cell discharge, migration and karyogamy Ingouff et al., 2007, 2009; Aw et al., 2010 Pollen tube guidance and discharge Eady et al., 1994; Rotman et al., 2003; Sandaklie-Nikolova et al., 2007 Karyogamy Ingouff et al., 2009 Karyogamy Kinoshita et al., 2004; Aw et al., 2010 Karyogamy Ingouff et al., 2007 Pollen tube discharge Activation of transcription and translation Paternal genome expression Reference Sandaklie-Nikolova et al., 2007 Aw et al., 2010 Ingouff et al., 2009 Imaging double fertilization | 1653 Fig. 1. Fluorescent markers of the male and female gametes, which can be used for live imaging of fertilization in Arabidopsis. (A) Bicellular pollen grain with expression of pLAT52-GFP in the vegetative cell (v) and pHTR10-HTR10-RFP in the generative cell (g). (B) Tricellular pollen grain with expression of pLAT52-GFP in the vegetative cell (v) and pHTR10-HTR10-RFP in the sperm cells (s). (C) Embryo sac with expression of pEC1-HISTONE2B-RFP in the egg cell (ec) and pFWA-GFP in the central cell (cc). Bars represent 10 lm. A germline-specific HISTONE 3 encoded by the gene HISTONE THREE RELATED 10 (HTR10; also known as AtGMH3) eventually provided a marker of sperm cells that fluoresces at levels required for in vivo live imaging (Okada et al., 2005, 2006; Ingouff et al., 2007). The expression of HTR10 fused to monomeric RFP under the control of its own promoter provides a strong fluorescent signal only in sperm cells in mature pollen. HTR10-RFP is not expressed in microspores. HTR10 expression is initiated after the unequal division leading to bicellular pollen and remains confined in the generative cell and the sperm cells (Fig. 1). HTR10 is only expressed in pollen and not in other cells than the generative cell and sperm cells (Okada et al., 2005; Ingouff et al., 2007). The HTR10 promoter can be used to drive expression of other genes in the sperm cell lineage (Brownfield et al., 2009). Marking the female gametophyte in Arabidopsis A systematic search for genes expressed in the female gametophyte has led to the identification of promoters causing expression in all or only some of the four cell types that constitute the female gametophyte (Steffen et al., 2007; Wang et al., 2010). Most promoters identified do not cause a specific expression in the gametophyte, nor in a single gametophytic cell type. The promoters of the genes corresponding to the AGI At5G01860 (Wang et al., 2010) and At5G40260 (Pagnussat et al., 2007) allow expression in all gametophytic cell types and are not active after fertilization. Such promoters can be used to test the impact of gene activity during gametophytic life and to label both the egg cell and the central cell. Several promoters, including the promoter of the gene MYB98 (Steffen et al., 2007), cause a fairly specific expression in the synergids, which can be useful for monitoring pollen tube arrival. Amongst several promoters, the promoter of the imprinted genes FWA confers a central cell-specific expression (Kinoshita et al., 2004). Egg cells have been isolated from various species including maize (Cordts et al., 2001; Yang et al., 2006), wheat (Triticum cereale) (Sprunck et al., 2005), and Arabidopsis (Wuest et al., 2010). EST sequencing and microarray analyses have been used to identify transcripts from isolated egg cells and have yielded some insights into the biology of the angiosperm female gamete. Surprisingly, only a few genes have been identified as being expressed specifically in the egg cell and not in other cells from the female gametophyte and the sporophytic tissues (Gross-Hardt et al., 2007; Steffen et al., 2007; Alandete-Saez et al., 2008; Ingouff et al., 2009; Wang et al., 2010). Transgenic lines expressing the fluorescent HISTONE2B-RFP under the control of the EC1 promoter enables specific and robust labelling of the egg cell nucleus (Fig. 1C) (Ingouff et al., 2009). Imaging double fertilization The small size and transparency of Arabidopsis reproductive organs are suitable for imaging double fertilization. An experimental set-up has been developed for imaging in planta (Rotman et al., 2003). Four hours after pollination, the pistils are placed on double-sided tape and one ovary wall is removed. A joint of silicon grease around the double-sided tape supports a coverslip, thus defining a small humid chamber. Fertilization and early endosperm and zygote development can be observed using confocal microscopy in vivo for up to 24 h after dissection. This set-up preserves as accurately as possible the normal conditions of fertilization since no liquid is in contact with the ovule and fertilization takes places inside the ovary. This set-up was used for imaging the discharge of the pollen tube contents in the female gametophyte (Rotman et al., 2003) and might be used to monitor sperm cell fusion and karyogamy. One disadvantage of imaging in planta is that it involves an air interface between the ovule and the coverslip, leading to relatively poorer optical resolution compared with when water immersion is employed in another in vivo imaging setup described hereafter. The in vivo imaging set-up was adapted from the method developed with Torenia fournieri (Higashiyama et al., 1998; Palanivelu and Preuss, 2006) and consists of excised ovules placed on medium on which pollen tubes grow (Sandaklie-Nikolova et al., 2007). This system enables higher resolution but does not reflect so 1654 | Berger precisely in vivo conditions because the ovules are excised and placed on semi-liquid medium and pollen tubes do not grow all the way inside maternal tissues. This set-up was used to image the delivery of sperm cell mitochondria labelled with GFP tagged with the N-terminal targeting signal from the gene encoding a mitochondrial F1-ATPase d subunit (Matsushima et al., 2008). This method also allowed imaging of sperm cell discharge, and karyogamy using sperm cells tagged with HTR10-RFP (Ingouff et al., 2007). In vivo imaging was performed using a disc-scan confocal scanning laser microscope equipped for rapid recording of z-stacks, and a prism to monitor two colours at the same time. Images could be recorded at a rate of one per 15 s or 30 s. The chromatin of nuclei of male gametes was labelled with HTR10-RFP and the female gametes were labelled with GFP expressed under the promoter of FERTILIZATION INDEPENDENT ENDOSPERM. It was possible to observe the discharge of sperm cells, followed by the observation of karyogamy (Fig. 2A). These series of experiments allowed the dynamics of the paternal chromatin upon double-fertilization to be monitored in vivo. Karyogamy results from the fusion between the male and female gamete nuclei and takes place about 1 h after release of the gametes (Fig. 2A). The spermcontributed HISTONE 3 (H3) variant HTR10 is actively Fig. 2. Double fertilization in Arabidopsis. (A) Karyogamy upon fertilization of an embryo sac marked with GFP expression under the FWA promoter in the central cell (cc) 7 h after pollination with tricellular pollen with sperm cells marked by the expression of pHTR10-HTR10-RFP. The paternal chromatin in red mixes with the maternal chromatin in the central cell nucleus and in the egg cell nucleus (ec). (B) Activation of transcription and translation marked by the expression of CENH3-GFP from the paternal allele after fertilization of an embryo sac marked by expression of pEC1HISTONE2B-RFP in the egg cell. The maternal chromatin in the zygote nucleus (z) is marked in red with green dots corresponding to centromeres. Centromeres are also marked by de novo synthesis of CENH3-GFP in endosperm (ed) nuclei. Bars represent 10 lm. removed within a few hours from the zygotic nucleus (Ingouff et al., 2007). Replacement of paternally inherited HTR10 does not occur after fertilization of the central cell, suggesting that the two fertilization events are not equivalent in terms of chromatin dynamics (Ingouff et al., 2007). Using the ubiquitously expressed centromeric histone variant H3 (CENH3), in vivo imaging enabled the earliest timing of zygotic activation to be determined (Aw et al., 2010) (Fig. 2B). CENH3-GFP is removed from the zygote with dynamics similar to that of HTR10 (F Berger, unpublished data). The absence of CENH3-GFP at the end of karyogamy allows recording of the onset of the activity of the transcription and translation machinery after fertilization. In developing seeds resulting from crosses between wild type pollen and ovules of a transgenic line expressing CENH3-GFP, CENH3 is expressed in the zygote from the maternal allele as early as 8 h after fertilization (Aw et al., 2010). This indicates that transcription and translation are intiated in the zygote earlier than previously thought (Pillot et al., 2009). New insights beyond description In planta and in vivo imaging protocols not only provide a more accurate description of double-fertilization, but also allows the phenotype of mutants defective for fertilization to be analysed in depth. Hence, live imaging provides mechanistic insights into double-fertilization. Studies of mutants that impair the dialogue between the synergids and the pollen tube and regulates sperm cell release have identified a new signalling pathway responsible for the control of sperm discharge. In vivo imaging showed that, in the mutant sire`ne, the pollen tube grows and coils in the female gametophyte without releasing its content (Rotman et al., 2003). The gene affected in the mutant sire`ne was later cloned from the allele feronia and shown to encode a receptor-like kinase (RLK), which accumulates on the synergid plasma membrane (Escobar-Restrepo et al., 2007). Other mutations causing a phenotype similar to sire`ne and feronia have been characterized. ANXUR1 and ANXUR2 are paralogues of the RLK FERONIA and are expressed in the pollen tube. ANXUR proteins are present in the plasma membrane and might participate in signalling events in the FERONIA pathway (Miyazaki et al., 2009) indicating that male and female gametes exchange signals regulating pollen tube arrest and gamete release. The female gametophytic mutants lorelei, scylla, and sire`ne share the same phenotype (Capron et al., 2008; Rotman et al., 2008). The gene SCYLLA has not been identified. LORELEI encodes a GPI-anchored G protein, which could be involved in signalling events downstream of FERONIA (Capron et al., 2008). The mutant abstinence by mutual consent (amc) (Boisson-Dernier et al., 2008) is characterized by a feronia phenotype but shows both female and male gametophytic effect. The role played by the AMC peroxin and the function of peroxysomes in fertilization remains unclear. In a few species double fertilization involves dimorphic female gametes and dimorphic sperm cells, while in most Imaging double fertilization | 1655 species sperm cells are not distinguished by their morphology (Lord and Russell, 2002). Still it was proposed that molecular functions are distinct between the two sperm cells and that one sperm cell is fated to fuse with the central cell while the other is fated to fuse with the egg cell (Lord and Russell, 2002). This hypothesis was tested in Arabidopsis using ovules of the mutants eostre (Pagnussat et al., 2007) and retinoblastoma related (rbr) (Ingouff et al., 2006), which produce two egg cells. It was possible to monitor directly the fusion of each sperm cell with each egg cell of the rbr mutant ovule (Ingouff et al., 2009) and to show that two embryos are produced (Ingouff et al., 2009), thus leading to the conclusion that each sperm cell is able to fuse with an egg cell. It was also shown, using live markers, that mutants in the CHROMATIN ASSEMBLY FACTOR pathway (Chen et al., 2008) and for the CYCLIN DEPENDENT KINASE A;1 (CDKA;1) (Aw et al., 2010) produce a fraction of pollen with a single sperm cell. The cdka;1 single sperm cell is able to fertilize either the central cell or the egg cell, which supports the idea that both sperm cells are equivalent. All the above results have been obtained using either mutant male or mutant female gametes. Such mutations may suppress the hypothetical mechanisms that may distinguish each sperm cell within the male germ unit. To know whether the two sperm cells are identical in Arabidopsis will ultimately require a protocol involving both wild-type male and female gametes. Initial studies of the cdka;1 phenotype had concluded that the mutant pollen produces a single sperm cell, that fertilizes the egg cell preferentially, causing the development of an embryo, which would produce a signal activating the central cell division (Iwakawa et al., 2006; Nowack et al., 2006). Live imaging applied to the analysis of the impact of cdka;1 on sexual reproduction has showed that, in a large fraction of cdka;1 pollen, the single sperm cell divides during pollen tube growth. Hence a majority of cdka;1 pollen tubes deliver two sperm cells. When cdka;1 pollen fertilizes wild-type ovules, one sperm cell fuses with the egg cell and the other sperm cell fuses with the central cell. Karyogamy is successful in the egg cell but not in the central cell. Sperm entry triggers cell division in the central cell, but after a few nuclei divisions, the endosperm-like structure aborts (Aw et al., 2010). These results indicate that the initiation of development of an endosperm-like structure by pollination with cdka;1 pollen does not result from a signal from the embryo as initially proposed (Nowack et al., 2006), but from an event associated with sperm entry. In addition, the absence of paternal genome expression in the endosperm leads to early abortion, which does not support the functional significance of a potential delay in paternal genome expression (Vielle-Calzada et al., 2000). Unanswered questions How exactly does sperm cell discharge take place? From the live imaging of pollen tube growth to the point of gamete release it is now clear that the pollen tube discharge does not cause synergid death (Rotman et al., 2003; Sandaklie-Nikolova et al., 2007). Still it remains unclear what causes synergid death and whether male gametes are delivered inside a degenerated synergid or if the gametes are discharged between one synergid and the egg cell. Sperm cell discharge occurs in less than 30 s. To answer these questions it will be necessary to develop imaging technology that allows sampling high definition images every 5 s, which should be possible with the latest generation of confocal microscopes and transgenic plant lines that combines several markers for the male and female gametes. How do sperm cells migrate? Dynamic observations suggest that after gamete fusion, the sperm cell nuclei migrate actively toward the nucleus of each female gametes. This migration may involve the actin cytoskeleton as suggested by immunolocalizations in tobacco and Torenia (Huang and Russell, 1994; Fu et al., 2000). Live imaging using newly developed markers of actin Lifeact-GFP (Era et al., 2009) and GFP-TUBULINa6 microtubules (Oh et al., 2010) will allow the dynamic aspects of the cytoskeleton after gamete fusion and its potential association with nuclei migration to be precisely determined. What prevents polyspermy? If the two sperm cells have an equivalent capacity to fuse with the egg cell and the central cell, there must be a mechanism preventing both sperm cells fusing with only one of the two female gametes. A limited number of studies in maize and Arabidopsis have suggested that the fertilization of the central cell causes a polyspermy block that is relatively inefficient in contrast to the fertilization of the egg cell (Scott et al., 2008; Spielman and Scott, 2008). These studies were based on mutant pollen that delivers additional sperm cells and it is still unknown what happens in the wildtype background. The lack of fluorescent markers for the plasma membrane hampers the establishment of the precise timing of gamete fusion. The site of gamete fusion is also not well defined. Live imaging with faster acquisition rates and higher optical resolution will be required to know whether the two sperm cells fuse simultaneously with each female gamete or if the fusions occurs sequentially, a prerequisite to understand the need for a polyspermy prevention mechanism. The regulation of the cell cycle during double-fertilization and in the zygote We still do not know at what stage of the cell cycle fertilization takes place. Convincing evidence has indicated that sperm cells are discharged after the S phase (Durbarry et al., 2005). The fact that mitosis is initiated immediately after karyogamy with the central cell nucleus also suggests that the central cell has reached the G2 phase at the time of fertilization. However, the regulation of the cell cycle in the zygote remains unclear. The 12 h delay between karyogamy 1656 | Berger and zygotic mitosis suggests two scenarios. (i) The male and female gametes all undergo DNA replication and karyogamy takes place in the G2 phase. The requirement for zygotic activation of transcription and translation accounts for the 12 h that elapses between karyogamy and the first mitosis in the fertilized egg. The fertilized central cell is transcriptionaly active immediately after fertilization (Aw et al., 2010) and is thus able to undergo mitosis immediately. (ii) The sperm cell and the central cell fuse after DNA replication while the egg cell does not undergo DNA replication. However, it is difficult to conceive how, after gamete nuclei fusion, the maternal genome would undergo S phase while the male genome has already duplicated in the zygotic nucleus. Both male and female gametes may undergo the S phase prior to fertilization and both gamete fusion events take place in G2 phase. The delayed mitosis in the zygote (relative to the immediate mitosis onset in the central cell) might be associated with a slight delay in transcriptional activation (estimated to a few hours), the acquisition of polarity, and directional elongation of the zygote. Investigations of these questions require the development of fluorescent markers that accurately reflect the dynamics of the cell cycle regulators, which has not currently been achieved. Acknowledgement FB’s research is funded by Temasek Lifescience Laboratory. References Alandete-Saez M, Ron M, McCormick S. 2008. GEX3, expressed in the male gametophyte and in the egg cell of Arabidopsis thaliana, is essential for micropylar pollen tube guidance and plays a role during early embryogenesis. Molecular Plant 1, 586–598. Antoine AF, Faure J, Cordeiro S, Dumas C, Rougier M, Feijo JA. 2000. A calcium influx is triggered and propagates in the zygote as a wavefront during in vitro fertilization of flowering plants. Proceedings of the National Academy of Sciences, USA 97, 10643–10648. Aw SJ, Hamamura Y, Chen Z, Schnittger A, Berger F. 2010. Sperm entry is sufficient to trigger division of the central cell but the paternal genome is required for endosperm development in Arabidopsis. Development 137, 2683–2690. Borges F, Gomes G, Gardner R, Moreno N, McCormick S, Feijo JA, Becker JD. 2008. Comparative transcriptomics of Arabidopsis sperm cells. Plant Physiology 148, 1168–1181. Boisson-Dernier A, Frietsch S, Kim TH, Dizon MB, Schroeder JI. 2008. The peroxin loss-of-function mutation abstinence by mutual consent disrupts male–female gametophyte recognition. Current Biology 18, 63–68. Brownfield L, Hafidh S, Borg M, Sidorova A, Mori T, Twell D. 2009. A plant germline-specific integrator of sperm specification and cell cycle progression. PLoS Genetics 5, e1000430. Campbell RE, Tour O, Palmer AE, Steinbach PA, Baird GS, Zacharias DA, Tsien RY. 2002. A monomeric red fluorescent protein. Proceedings of the National Academy of Sciences, USA 99, 7877–7882. Capron A, Gourgues M, Neiva LS, et al. 2008. Maternal control of male-gamete delivery in Arabidopsis involves a putative GPI-anchored protein encoded by the LORELEI gene. The Plant Cell 20, 3038–3049. Chalfie M, Tu Y, Euskirchen G, Ward WW, Prasher DC. 1994. Green fluorescent protein as a marker for gene expression. Science 263, 802–805. Chen Z, Tan JL, Ingouff M, Sundaresan V, Berger F. 2008. Chromatin assembly factor 1 regulates the cell cycle but not cell fate during male gametogenesis in Arabidopsis thaliana. Development 135, 65–73. Cheung AY. 2001. Imaging elongating pollen tubes by green fluorescent protein. Sexual Plant Reproduction 14, 9–14. Christensen CA, King EJ, Jordan JR, Drews GN. 1997. Megagametogenesis in Arabidopsis wild type and the Gf mutant. Sexual Plant Reproduction 10, 49–64. Cordts S, Bantin J, Wittich PE, Kranz E, Lorz H, Dresselhaus T. 2001. ZmES genes encode peptides with structural homology to defensins and are specifically expressed in the female gametophyte of maize. The Plant Journal 25, 103–114. Digonnet C, Aldon D, Leduc N, Dumas C, Rougier M. 1997. First evidence of a calcium transient in flowering plants at fertilization. Development 124, 2867–2874. Durbarry A, Vizir I, Twell D. 2005. Male germ line development in Arabidopsis: duo pollen mutants reveal gametophytic regulators of generative cell cycle progression. Plant Physiology 137, 297–307. Eady C, Lindsey K, Twell D. 1994. Differential activation and conserved vegetative cell-specific activity of a late pollen promoter in species with bicellular and tricellular pollen. The Plant Journal 5, 543–550. Engel ML, Chaboud A, Dumas C, McCormick S. 2003. Sperm cells of Zea mays have a complex complement of mRNAs. The Plant Journal 34, 697–707. Engel ML, Holmes-Davis R, McCormick S. 2005. Green sperm: identification of male gamete promoters in Arabidopsis. Plant Physiology 138, 2124–2133. Era A, Tominaga M, Ebine K, Awai C, Saito C, Ishizaki K, Yamato KT, Kohchi T, Nakano A, Ueda T. 2009. Application of Lifeact reveals F-actin dynamics in Arabidopsis thaliana and the liverwort, Marchantia polymorpha. Plant and Cell Physiology 50, 1041–1048. Escobar-Restrepo JM, Huck N, Kessler S, Gagliardini V, Gheyselinck J, Yang WC, Grossniklaus U. 2007. The FERONIA receptor-like kinase mediates male–female interactions during pollen tube reception. Science 317, 656–660. Faure J-E, Digonnet C, Dumas C. 1994. An in vitro system for adhesion and fusion of maize gametes. Science 263, 1598–1600. Faure JE, Dumas C. 2001. Fertilization in flowering plants: new approaches for an old story. Plant Physiology 125, 102–104. Faure JE, Rotman N, Fortune P, Dumas C. 2002. Fertilization in Arabidopsis thaliana wild type: developmental stages and time course. The Plant Journal 30, 481–488. Imaging double fertilization | 1657 Feijo JA, Moreno N. 2004. Imaging plant cells by two-photon excitation. Protoplasma 223, 1–32. Friedman WE. 2001. Developmental and evolutionary hypotheses for the origin of double fertilization and endosperm. Comptes rendu hebdomadaire des séances de l’Academie des sciences 324, 559–567. Fu Y, Yuan M, Huang BQ, Yang HY, Zee SY, Tp OB. 2000. Changes in actin organization in the living egg apparatus of Torenia fournieri during fertilization. Sexual Plant Reproduction 12, 315–322. Giepmans BN, Adams SR, Ellisman MH, Tsien RY. 2006. The fluorescent toolbox for assessing protein location and function. Science 312, 217–224. Gou X, Yuan T, Wei X, Russell SD. 2009. Gene expression in the dimorphic sperm cells of Plumbago zeylanica: transcript profiling, diversity, and relationship to cell type. The Plant Journal 60, 33–47. Gross-Hardt R, Kagi C, Baumann N, Moore JM, Baskar R, Gagliano WB, Jurgens G, Grossniklaus U. 2007. LACHESIS restricts gametic cell fate in the female gametophyte of Arabidopsis. PLoS Biology 5, e47. Guignard ML. 1899. Sur les anthérozoı̈des et la double copulation sexuelle chez les végétaux angiospermes. Revue Génerale de Botanique 11, 129–135. Haseloff J. 1999. GFP variants for multispectral imaging of living cells. Methods in Cell Biology 58, 139–151. Higashiyama T, Kuroiwa H, Kawano S, Kuroiwa T. 1997. Kinetics of double fertilization in Torenia fournieri based on direct observations of the naked embryo sac. Planta 203, 101–110. Higashiyama T, Kuroiwa H, Kawano S, Kuroiwa T. 1998. Guidance in vitro of the pollen tube to the naked embryo sac of Torenia fournieri. The Plant Cell 10, 2019–2032. Kranz E, Bautor J, Lörz H. 1991. In vitro fertilization of single, isolated gametes of maize mediated by electrofusion. Sexual Plant Reproduction 4, 12–16. Kranz E, Dresselhaus T. 1996. In vitro fertilization with isolated higher plant gametes. Trends in Plant Science 1, 82–89. Kranz E, Lörz H. 1994. In vitro fertilisation of maize by single egg and sperm cell protoplast fusion mediated by high calcium and high pH. Zygote 2, 125–128. Lord EM, Russell SD. 2002. The mechanisms of pollination and fertilization in plants. Annual Review of Cell and Development Biology 18, 81–105. Matsushima R, Hamamura Y, Higashiyama T, Arimura S, Sodmergen Tsutsumi N, Sakamoto W. 2008. Mitochondrial dynamics in plant male gametophyte visualized by fluorescent live imaging. Plant and Cell Physiology 49, 1074–1083. Miyazaki S, Murata T, Sakurai-Ozato N, Kubo M, Demura T, Fukuda H, Hasebe M. 2009. ANXUR1 and 2, sister genes to FERONIA/SIRENE, are male factors for coordinated fertilization. Current Biology 19, 1327–1331. Mori T, Kuroiwa H, Higashiyama T, Kuroiwa T. 2006. GENERATIVE CELL SPECIFIC 1 is essential for angiosperm fertilization. Nature Cell Biology 8, 64–71. Nawaschin SG. 1898. Resultate einer revision der befruchtungsvorgaenge bei Lilium martagon und Fritillaria tenella. Bulletin de l#Académie Impériale des Sciences de Saint Pétersbourg 9, 377–382. Nowack MK, Grini PE, Jakoby MJ, Lafos M, Koncz C, Schnittger A. 2006. A positive signal from the fertilization of the egg cell sets off endosperm proliferation in angiosperm embryogenesis. Nature Genetics 38, 63–67. Higashiyama T, Kuroiwa H, Kawano S, Kuroiwa T. 2000. Explosive discharge of pollen tube contents in Torenia fournieri. Plant Physiology 122, 11–14. Oh SA, Pal MD, Park SK, Johnson JA, Twell D. 2010. The tobacco MAP215/Dis1-family protein TMBP200 is required for the functional organization of microtubule arrays during male germline establishment. Journal of Experimental Botany 61, 969–981. Huang B-Q, Russell SD. 1994. Fertilization in Nicotiana tabacum: cytoskeletal modifications in the embryo sac during synergid degeneration. Planta 194, 200–214. Okada T, Endo M, Singh MB, Bhalla PL. 2005. Analysis of the histone H3 gene family in Arabidopsis and identification of the malegamete-specific variant AtMGH3. The Plant Journal 44, 557–568. Ingouff M, Hamamura Y, Gourgues M, Higashiyama T, Berger F. 2007. Distinct dynamics of HISTONE3 variants between the two fertilization products in plants. Current Biology 17, 1032–1037. Okada T, Singh MB, Bhalla PL. 2006. Histone H3 variants in male gametic cells of lily and H3 methylation in mature pollen. Plant Molecular Biology 62, 503–512. Ingouff M, Jullien PE, Berger F. 2006. The female gametophyte and the endosperm control cell proliferation and differentiation of the seed coat in Arabidopsis. The Plant Cell 18, 3491–3501. Pagnussat GC, Yu HJ, Sundaresan V. 2007. Cell-fate switch of synergid to egg cell in Arabidopsis eostre mutant embryo sacs arises from misexpression of the BEL1-Like homeodomain gene BLH1. The Plant Cell 19, 3578–3592. Ingouff M, Sakata T, Li J, Sprunck S, Dresselhaus T, Berger F. 2009. The two male gametes share equal ability to fertilize the egg cell in Arabidopsis thaliana. Current Biology 19, R19–R20. Palanivelu R, Preuss D. 2006. Distinct short-range ovule signals attract or repel Arabidopsis thaliana pollen tubes in vitro. BMC Plant Biology 6, 7. Iwakawa H, Shinmyo A, Sekine M. 2006. Arabidopsis CDKA;1, a cdc2 homologue, controls proliferation of generative cells in male gametogenesis. The Plant Journal 45, 819–831. Pillot M, Baroux C, Vazquez MA, Autran D, Leblanc O, Vielle-Calzada JP, Grossniklaus U, Grimanelli D . Embryo and Endosperm Inherit Distinct Chromatin and Transcriptional States from the Female Gametes in Arabidopsis. The Plant Cell 22, 307–320. Kinoshita T, Miura A, Choi Y, Kinoshita Y, Cao X, Jacobsen SE, Fischer RL, Kakutani T. 2004. One-way control of FWA imprinting in Arabidopsis endosperm by DNA methylation. Science 303, 521–523. Rotman N, Durbarry A, Wardle A, Yang WC, Chaboud A, Faure JE, Berger F, Twell D. 2005. A novel class of MYB factors controls sperm-cell formation in plants. Current Biology 15, 244–248. 1658 | Berger Rotman N, Gourgues M, Guitton AE, Faure J, Berger F. 2008. A dialogue between the sirene pathway in synergids and the fertilization independent seed pathway in the central cell controls male gamete release during double fertilization in Arabidopsis. Molecular Plant 4, 659–666. Rotman N, Rozier F, Boavida L, Dumas C, Berger F, Faure JE. 2003. Female control of male gamete delivery during fertilization in Arabidopsis thaliana. Current Biology 13, 432–436. Sandaklie-Nikolova L, Palanivelu R, King EJ, Copenhaver GP, Drews GN. 2007. Synergid cell death in Arabidopsis is triggered following direct interaction with the pollen tube. Plant Physiology 144, 1753–1762. Scott RJ, Armstrong SJ, Doughty J, Spielman M. 2008. Double fertilization in Arabidopsis thaliana involves a polyspermy block on the egg but not the central cell. Molecular Plant 1, 611–619. Spielman M, Scott R. 2008. Polyspermy barriers in plants: from preventing to promoting fertilization. Sexual Plant Reproduction 21, 53–65. Sprunck S, Baumann U, Edwards K, Langridge P, Dresselhaus T. 2005. The transcript composition of egg cells changes significantly following fertilization in wheat (Triticum aestivum L.). The Plant Journal 41, 660–672. Steffen JG, Kang IH, Macfarlane J, Drews GN. 2007. Identification of genes expressed in the Arabidopsis female gametophyte. The Plant Journal 51, 281–292. Tsien RY. 1998. The green fluorescent protein. Annual Review of Biochemistry 67, 509–544. Twell D, Yamaguchi J, Wing RA, Ushiba J, McCormick S. 1991. Promoter analysis of genes that are coordinately expressed during pollen development reveals pollen-specific enhancer sequences and shared regulatory elements. Genes and Development 5, 496–507. Vielle- Calzada JP, Baskar R, Grossniklaus U. 2000. Delayed activation of the paternal genome during seed development. Nature 404, 91–94. von Besser K, Frank AC, Johnson MA, Preuss D. 2006. Arabidopsis HAP2 (GCS1) is a sperm-specific gene required for pollen tube guidance and fertilization. Development 133, 4761–4769. Wang D, Zhang C, Hearn DJ, Kang IH, Punwani JA, Skaggs MI, Drews GN, Schumaker KS, Yadegari R. 2010. dentification of transcription-factor genes expressed in the Arabidopsis female gametophyte. BMC Plant Biology 10, 110. Weterings K, Reijnen W, van Aarssen R, Kortstee A, Spijkers J, van Herpen M, Schrauwen J, Wullems G. 1992. Characterization of a pollen-specific cDNA clone from Nicotiana tabacum expressed during microgametogenesis and germination. Plant Molecular Biology 18, 1101–1111. Wuest SE, Vijverberg K, Schmidt A, Weiss M, Gheyselinck J, Lohr M, Wellmer F, Rahnenfuhrer J, von Mering C, Grossniklaus U. 2010. Arabidopsis female gametophyte gene expression map reveals similarities between plant and animal gametes. Current Biology 20, 506–512. Yang H, Kaur N, Kiriakopolos S, McCormick S. 2006. EST generation and analyses towards identifying female gametophytespecific genes in Zea mays L. Planta 224, 1004–1014.