Survey
* Your assessment is very important for improving the workof artificial intelligence, which forms the content of this project
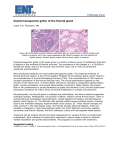
0013-7227/01/$03.00/0 Printed in U.S.A. The Journal of Clinical Endocrinology & Metabolism 86(10):4843– 4848 Copyright © 2001 by The Endocrine Society Goiter and Hypothyroidism in Two Siblings due to Impaired Caⴙ2/NAD(P)H-Dependent H2O2-Generating Activity MARCIA D. L. FIGUEIREDO, LUCIENE C. CARDOSO, ANDREA C. F. FERREIRA, DENISE V. B. CAMPOS, MANOEL DA CRUZ DOMINGOS, ROSSANA CORBO, LUIZ EURICO NASCIUTTI, MARIO VAISMAN, AND DENISE P. CARVALHO Instituto de Biofı́sica Carlos Chagas Filho (M.D.L.F., L.C.C., A.C.F.F., D.V.B.C., D.P.C.), Departamento de Histologia e Embriologia (L.E.N.), and Serviços de Endocrinologia (M.V.), Cirurgia (M.d.C.D.), and Medicina Nuclear (R.C.), Hospital Universitário Clementino Fraga Filho, Universidade Federal do Rio de Janeiro, 21949-900 Rio de Janeiro, Brazil We report herein the study of two siblings (DESM and DSM) with hypothyroidism, goiter, and positive perchlorate discharge tests (50% and 70%) in a family (M) with no history of consanguinity. Thyroid gland histology showed a predominance of hyperactive follicles, with high epithelial cells and variable colloid content. Thyroid peroxidase iodide oxidation (DESM, 1034; DSM, 1064 U/g protein) and albumin iodination (DESM, 16; DSM, 8 nmol I/mg protein) activities were within the normal range. Tg content was normal in both glands compared with that in diffuse toxic goiter (DESM, 28; DSM, 17; diffuse toxic goiter, 19 mg/g tissue), and Tg could be normally iodinated by thyroid peroxidase in vitro (DESM, 3.4; DSM, 4.3; diffuse toxic goiter, 6.3 nmol I/mg Tg). Thyroid cytochrome c reductase activities in these goiters were higher than that in paranodular tissues (DESM, 473; DSM, 567; paranodular tissues, 78 nmol NADPⴙ/h/mg protein). However, thyroid NADPH oxidase activities were very low both in the particulate 3,000 ⴛ g (DESM, 4.8; DSM, 44; paranodular tissues, 224 nmol H2O2/ h/mg protein) and in the particulate 100,000 ⴛ g fractions (DESM, 40; DSM, 47; paranodular tissues, 200 nmol H2O2/h/mg protein). Thus, a decreased Ca2ⴙ/NAD(P)H-dependent H2O2 generation is the probable cause of the organification defect in these goiters. (J Clin Endocrinol Metab 86: 4843– 4848, 2001) I In human thyroid tissues the presence of a Ca2⫹/ NAD(P)H-dependent H2O2 generator similar to the porcine thyroid NADPH oxidase has only recently been characterized (13), and the thyroid oxidase (ThOx) cDNA has been cloned (10, 11). However, further molecular identification of the thyroid enzyme has not yet been achieved, and the cDNAs cloned probably correspond only to the flavoprotein, a component of the enzymatic system (14). We report herein the study of two siblings with goiter and hypothyroidism due to iodine organification defect. In these goiters we found normal TPO activity and Tg content, but thyroid NADPH oxidase activity was almost undetectable in both the particulate 3,000 ⫻ g (P3,000 g) and 100,000 ⫻ g (P100,000 g) fractions compared with the activity found in paranodular to cold thyroid nodule tissues. ODIDE IS RAPIDLY transported into thyroid follicular cells and in the cellular apical surface is covalently bound to Tg in an enzymatic reaction called iodine organification (1, 2). Failure of iodine organification results in impaired thyroid hormone biosynthesis, increased TSH stimulation, and goiter (3, 4). In patients with goiter and hypothyroidism, a thyroid iodine organification defect is suspected when intrathyroidal radiolabeled iodide is displaced by perchlorate or thiocyanate administration, resulting in a positive perchlorate discharge test (3, 4). Thyroid iodine organification depends on thyroid peroxidase (TPO) activity, which is modulated by the concentration of substrates (Tg and iodide) and cofactor (hydrogen peroxide) (1). H2O2 generation is a limiting step in thyroid hormone biosynthesis, as previously shown in dog thyroid slice (5). In porcine thyroid glands previous biochemical studies have reported that H2O2 is generated by a calciumdependent NAD(P)H:O2 oxidoreductase, the thyroid NADPH oxidase (6 – 8). Some researchers have previously shown that thyroid NADPH oxidase activity (9) and gene expression (10, 11) are induced by TSH, as are TPO and Tg. In dog thyrocytes, TSH also induces the expression of the H2O2 generator via the cAMP cascade (12). NADPH oxidase, therefore, seems to be the enzyme responsible for H2O2 generation linked to thyroid hormonogenesis. Abbreviations: DTG, Diffuse toxic goiter; HRP, horseradish peroxidase; P3,000 g, particulate 3,000 ⫻ g; P100,000 g, particulate 100,000 ⫻ g; PAS, periodic acid-Schiff; PN, paranodular tissues; ThOx, thyroid oxidase; TPO, thyroid peroxidase. Materials and Methods Materials NADPH and lyophilized horseradish peroxidase (HRP; grade 1) were purchased from Roche (Mannheim, Germany); BSA, scopoletin, cytochrome c, and FAD were obtained from Sigma (St. Louis, MO). Patients Family M, with no history of consanguinity, is composed of three siblings; two of them are affected (Fig. 1), indicating that the patients can be heterozygous for the mutation. The index patient (DSM), a 19 yr-old male, underwent total thyroidectomy for mechanical neck compression caused by the presence of a very large goiter. At 3 yr of age he started thyroid hormone replacement due to thyroid enlargement associated with clinical and laboratory findings of hypothyroidism. Hearing and 4843 4844 J Clin Endocrinol Metab, October 2001, 86(10):4843– 4848 mental development were normal. During adolescence his goiter increased in size. At the time of surgery a large multinodular asymmetrical goiter with a fiber-elastic texture was noted. Thyroid autoantibodies (anti-TPO and anti-Tg) were negative. Two-hour thyroid radioiodine uptake was 63% (normal, 4 –12%), and potassium perchlorate (1.5 g, orally) administration produced a discharge of 70% of the thyroid radioiodine content. Ultrasonographic studies of the thyroid confirmed multinodular goiter, with several anechoic and hypoechoic areas. DSM had been off thyroid medication for 30 d at the time of surgery. His sister (DESM) had hypothyroidism diagnosed at 3 months of age, when thyroid hormone replacement was started. She also had no mental retardation or hearing loss, and did not maintain adequate adherence to the prescribed medication. Goiter was first noted when she was 11 yr old, and her thyroid enlarged considerably during adolescence. At admission to the university hospital at the age of 17 yr, a large soft multinodular asymmetrical goiter was noted. Thyroid autoantibodies (antiTPO and anti-Tg) were also negative. Thyroid radioiodine uptake (2 h) was increased (77%; normal, 4 –12%), and she also had a positive perchlorate discharge test (50% of thyroid-accumulated radioiodine). Thyroidectomy was performed due to compressive symptoms, and the patient was off thyroid hormone replacement for 30 d before surgery. The patients gave their informed consent and the study was approved by the institutional human research committee. Serum total and free T4, T3, and TSH were assayed using commercial kits (Euro/Diagnostic Products, Los Angels, CA). Anti-TPO and anti-Tg autoantibodies and serum Tg were also assayed using commercial kits. Laboratory data from both patients are described in Table 1. No cochlear malformation was detected by computer axial tomography. Thyroid tissue Thyroid tissue samples from the dyshormonogenetic goiters were obtained during thyroidectomy. Samples from paranodular tissues (PN) surrounding cold thyroid nodules (n ⫽ 5) or a diffuse toxic goiter (DTG) were used as controls. Analyses in the DTG sample were performed in parallel with dyshormonogenetic goiters. Patients with cold thyroid nodules received no medication before surgery, and the patient with diffuse toxic goiter used propylthiouracil until the day before surgery and saturated iodine solution (Lugol, five drops, three times per day) 14 d before surgery. When the DTG patient achieved euthyroidism, thyroidectomy was performed. Thyroid tissue samples were either freshly processed for H2O2 generation measurements or stored at ⫺20 C for other experiments. Figueiredo et al. • Goiter due to Thyroid NADPH Oxidase Defect For light microscopy, thyroid tissue samples were immediately fixed in 10% buffered formalin. Sections of paraffin-embedded tissue were stained with hematoxylin-eosin or periodic acid-Schiff (PAS). Thyroid sample processing For NADPH oxidase and cytochrome c reductase preparations, fresh human thyroid tissue samples (1 g) were cleaned from fibrous tissue or hemorrhagic areas, minced, and homogenized in sodium phosphate buffer, pH 7.2, containing 0.25 m sucrose, 0.5 mm dithiothreitol, and 1 mm EGTA using an Ultra-Turrax (IKA, Staufen, Germany). The homogenate was filtered through cheesecloth. The particulate fraction was collected by centrifugation at 3,000 ⫻ g for 15 min at 4 C and resuspended in 3 ml 50 mm sodium phosphate buffer, pH 7.2, containing 0.25 m sucrose and 2 mm MgCl2 (buffer A). The pellet was washed twice with 3 ml buffer A and centrifuged at 3,000 ⫻ g for 15 min at 4 C. The last pellet (P3,000 g) was gently resuspended in 1 ml buffer A. The supernatant of the first centrifugation was centrifuged at 100,000 ⫻ g for 1 h at 4 C. The pellet (microsomal fraction, P100,000 g) was washed twice in 2 ml buffer A and gently resuspended in 0.5 ml buffer A. Protein concentrations were measured by the method of Bradford (15), using BSA as standard. The particulate fractions (P3,000 g and P100,000 g) were incubated with 1 n NaOH (30 min, 20 C) to dissolve particulate before protein determination. For TPO preparation, thyroid tissue samples (1 g) were cleaned from fibrous tissue or hemorrhagic areas, minced, and homogenized in 50 mm Tris-HCl buffer, pH 7.2, containing 1 mm potassium iodide, using an Ultra-Turrax. The homogenate was centrifuged at 100,000 ⫻ g for 1 h at 4 C, and the pellet was resuspended in 2 ml digitonin (1%, wt/vol). The mixture was incubated at 4 C for 24 h and then centrifuged at 100,000 ⫻ g for 1 h at 4 C. The supernatant containing solubilized TPO was used for the iodide oxidation and albumin iodination assays, as previously described (16 –18). Ca2⫹- and NADPH-dependent H2O2 generating system: NADPH oxidase activity H2O2 formation was measured by incubating samples of the thyroid particulate fractions (P3,000 g and P100,000 g) at 30 C in 1 ml 170 mm sodium phosphate buffer, pH 7.4, containing 1 mm sodium azide, 1 mm EGTA, 1 m FAD, and 1.5 mm CaCl2, as previously described (8, 9, 13). The reaction was started by adding 0.2 mm NADPH; aliquots of 100 l were collected at intervals up to 20 min and mixed with 10 l 3 n HCl to stop the reaction and destroy the remaining NADPH. Initial rates of H2O2 formation were determined from eight aliquots of each assay by following the decrease in 0.4 m scopoletin fluorescence in the presence of HRP (0.5 g/ml) in 200 mm phosphate buffer, pH 7.8, in a spectrofluorometer (F 4000, Hitachi, Hialeah, FL), as previously described (13). The excitation and emission wavelengths were 360 and 460 nm, respectively. All measurements were performed on at least three samples from each particulate preparation and are expressed as nanomoles of H2O2 per h. Specific activities were expressed per mg protein (nanomoles of H2O2 per h/mg protein) in the thyroid P3,000 g and P100,000 g fractions. Thyroid NADPH-cytochrome c reductase activity FIG. 1. Pedigree of the patient’s family. F, Female subjects studied; f, male subjects studied; 䡺, normal male; E, normal female; L, female with goiter. Aliquots of human thyroid particulate fractions (P3,000 g and P100,000 g) were incubated at 30 C in 1 ml 50 mm sodium phosphate buffer (pH 7.2) containing 1 mm sodium azide, 1.2 mm EGTA, and 0.1 mm cytochrome c. The reaction was started by adding 0.1 mm NADPH. The initial NADPH concentration was measured at 340 nm in a U-3300 (Hitachi) double beam spectrophotometer, using a molar absorption TABLE 1. Laboratorial data from DESM and DSM patients 2-h DESM DSM Normal range 131 I uptake (%) 63 77 4 –12 Perchlorate test (% of 131I discharged) 50 70 ⬍15 Total T3 (ng/dl) Total T4 (g/dl) Free T4 (ng/dl) TSH (U/ml) Serum Tg (ng/ml) 64.4 72.6 1.2 1.8 0.23 0.22 694.0 12.5 ⬎500 ⬎500 80 –180 4.5–12.5 0.9 –1.9 0.47–5.0 ⬍55 All data were obtained after suspension of T4 replacement for 30 days. Figueiredo et al. • Goiter due to Thyroid NADPH Oxidase Defect J Clin Endocrinol Metab, October 2001, 86(10):4843– 4848 4845 coefficient of 6.2 ⫻ 103 m/cm. Aliquots of 100 l were taken at intervals and mixed with 10 l 2 mm dithiothreitol and 100 l 2% SDS to stop the reaction. The initial rates of cytochrome c-dependent NADPH oxidation were determined from eight aliquots of each assay by following the decrease in NADPH fluorescence at pH 8.0 in a Hitachi spectrofluorometer (F 4000), as previously described (9). The excitation and emission wavelengths were 340 and 453 nm, respectively. NADPH-cytochrome c reductase activity was expressed as nanomoles of NADPH oxidized per h and milligrams of protein in the thyroid P3,000 g and P100,000 g fractions (nanomoles of NADP⫹ per h/mg protein). TPO iodide oxidation and albumin iodination activities TPO iodide oxidation assays were performed using 12 mm potassium iodide in 50 mm phosphate buffer (pH 7.4) and glucose-glucose oxidase as the hydrogen peroxide (H2O2)-generating system, as previously described (16, 17). The increase in absorbency at 353 nm (⌬A353) was followed for 4 min on a U-3300 Hitachi double beam spectrophotometer. The TPO activity was estimated from the ⌬A353/min determined from the linear portion of the reaction curve. One unit of iodide oxidation activity is defined as ⌬A353/min, and activity was related to the protein concentration in the enzyme preparation (units per g protein). The TPO albumin iodination activity was determined using BSA as iodine acceptor, and trichloroacetic acid precipitation of iodine bound to BSA, as previously described (18). TPO iodination activity was expressed as nanomoles of I bound per mg protein/30 min. Tg extraction and purification Thyroid tissue samples (1 g) were homogenized and centrifuged at 100,000 ⫻ g at 4 C for 1 h. The supernatant containing soluble proteins were submitted to salting out with 50% ammonium sulfate (4 m, pH 7.2), to precipitate Tg. After a 24-h incubation period samples were centrifuged (10,000 ⫻ g, 4 C, 30 min), and the pellets were resuspended in 1 ml 50 mm sodium phosphate buffer, pH 7.4. Purified Tg was submitted to dialysis (4 C, overnight), followed by protein measurement by the method of Bradford (15). Tg aliquots were either used for TPO iodination assays or SDS-PAGE gel electrophoresis (6.0% polyacrylamide), using the method of Laemmli (19). Equal amounts of Tg protein (25 g) were applied and then stained by Coomassie blue. Results TPO iodide oxidation and albumin iodination activities in the goiters of hypothyroid patients were higher than the normal range found in previous reports from our laboratory (16, 17) and did not differ significantly from the TPO activity found in the DTG sample studied in parallel (Table 2). Normal amounts of Tg could be extracted from the goitrous tissues (DESM, 28; DSM, 17; DTG, 19 mg Tg/g tissue), and their purified Tg could be normally iodinated in vitro (DESM, 3.4; DSM, 4.3; DTG, 6.3 nmol I/mg Tg). Also, SDS-PAGE analysis showed that Tg extracted from these goiters was of normal mol wt. The routine histological staining showed a similar general appearance of thyroid architecture in both patients (Fig. 2). Single layers of epithelial cells form spherical structures, the TABLE 2. Thyroperoxidase activity found in DESM and DSM goiters compared with TPO obtained from diffuse toxic goiters (DTG) and paranodular tissue (PN) TPO activity DESM DSM PN DTG Iodide oxidation (U/g protein) Albumin iodination (nmol I/mg protein) 1034 1064 232 ⫾ 121 2267 16 8 24a 66 a Mean of two normal tissues previously published by our group in Ref. 16. thyroid follicles, and delimit the follicle lumens, which contain Tg. Although the follicles exhibit variable size and shape, there is a predominance of hyperactive follicles, with high epithelial cells and variable colloid quantity, especially in the tissue sample from DESM. In PN thyroid tissues, the NAPDH oxidase H2O2-generating levels found in the P3,000 g fraction (224 ⫾ 38 nmol H2O2/h/mg protein) were similar to those described for porcine thyroid particulate (6, 8). On the other hand, H2O2 generation was either undetectable (DESM) or very low (DSM) in the P3,000 g (Fig. 3A) and low in the P100,000 g (Fig. 3B) obtained from DESM and DSM. Cytochrome c reductase is another enzymatic system that was proposed as capable of generating H2O2 in the thyroid gland. We found high cytochrome c reductase activity compared with PN tissues in either the P3,000 g or P100,000 g fraction (Fig. 4, A and B). Discussion In family M, the early onset of hypothyroidism together with goiter development and a positive perchlorate discharge test suggest an inherited defect in thyroid iodine organification, as previously reported for other families (20 – 22). The mode of inheritance remains unclear. By history, there is no suggestion of a common ancestor, and this is supported by genotyping several loci using microsatellite markers (Carvalho, D. P., and P. Kopp, unpublished results). A recessive mode of inheritance with homozygosity or compound heterozygosity for a defective gene cannot be excluded. Alternatively, the two affected siblings could carry only one defective allele, exerting a dominant negative effect or resulting in haploinsufficiency. Lastly, although less likely, one should also consider the possibility that inactivation of one allele in two (or several) distinct genes could be involved in the pathogenesis of the disorder. Usually, Tg defects are described as the cause of dyshormonogenesis with a negative perchlorate discharge test. In our patients a Tg defect can be excluded, because, apart from the finding of positive perchlorate discharge, their Tg was normally produced and could be normally iodinated by TPO in vitro. Furthermore, the histological analysis shows normal follicle structures with Tg detected by PAS in the lumen, also excluding the possibility of a Tg transport defect as previously reported (23–25). In Pendred’s syndrome it is believed that the cause of the iodine organification defect is impaired iodide transport through pendrin, a chloride/iodide transporter found in the apical thyroid plasma membrane (26, 27). However, in Pendred’s syndrome perchlorate is usually only partially discharged, the patients have congenital sensorineural deafness, and most of them are euthyroid (28). In contrast, our patients are hypothyroid, have a complete organification defect, as an almost complete perchlorate discharge (66% and 77%) was detected, and have no signs of cochlear malformation, indicating that pendrin might be normal in these two goitrous patients. Defective TPO is believed to be the most common cause of a positive perchlorate discharge test in thyroid dyshormonogenesis (22). However, in some patients with iodine or- 4846 J Clin Endocrinol Metab, October 2001, 86(10):4843– 4848 Figueiredo et al. • Goiter due to Thyroid NADPH Oxidase Defect FIG. 2. Light micrographs of thyroid tissue. Thyroid tissue samples from the goiters of two hypothyroid siblings, DSM (B and C) and DESM (A and D), were fixed in 10% buffered formalin. Sections of paraffin-embedded tissue were stained with hematoxylin eosin (A) or PAS (B–C). A and B, Typical thyroid follicular structure and stroma (S) with irregular collagenous connective tissue. Follicular size and shape are heterogeneous. The colloid (C) is PAS positive. C and D, Some hyperactive follicles with hyperplasia of the follicular epithelial cells (FC) and regions of colloid endocytosis (*). The basal lamina (BL) that unsheathes each follicle is also PAS positive. A, ⫻100; B, ⫻40; C and D, ⫻400. ganification defects and normal TPO, impaired H2O2 generation has been proposed (16, 29, 30). Kusakabe (30) has described a defect in thyroid cytochrome b5 reductase as the cause of a goiter with H2O2 generation defect, but the participation of this enzyme in thyroid H2O2 generation has not been confirmed. The nature of the enzymatic system involved in thyroid H2O2 production has only recently been determined (10, 11, 13), and current data strongly indicate that H2O2 linked to thyroid hormonogenesis is generated by Ca2⫹- and NAD(P)H- dependent ThOx (NADPH oxidase or ThOx). In the present study we found normal TPO and decreased NADPH oxidase activity in both siblings, reinforcing the possibility that their goiters might be caused by a defect in H2O2 generation. Recently, two cDNAs (ThOx 1 and ThOx 2) that encode ThOx have been cloned (10, 11), but they correspond to the flavoprotein, a component of the H2O2-generating enzymatic system that is responsible for NADPH oxidation, but not for the electron transfer necessary for H2O2 production (14). Hence, as biochemical data suggest that thyroid H2O2 is Figueiredo et al. • Goiter due to Thyroid NADPH Oxidase Defect J Clin Endocrinol Metab, October 2001, 86(10):4843– 4848 4847 NAD(P)H:O2 oxidoreductase is involved in H2O2 generation associated with TPO and participates in the iodine organification reaction. Acknowledgments Received March 22, 2001. Accepted June 26, 2001. Address all correspondence and requests for reprints to: Dr. Denise Pires de Carvalho, Instituto de Biofı́sica Carlos Chagas Filho, CCS-Bloco G, Cidade Universitária, Ilha do Fundão, 21949-900 Rio de Janeiro, Brazil. E-mail: [email protected]. FIG. 3. Thyroid NADPH oxidase activity. Ca2⫹- and NADPH-dependent H2O2 generation was measured in human thyroid samples obtained from five paranodular to cold nodule tissues (PN) and from two siblings with goiter and hypothyroidism (DESM and DSM). H2O2 generation was measured in 170 mM sodium phosphate buffer, pH 7.4, containing 1 mM sodium azide, 1 mM EGTA, 1 M FAD, and 1.5 mM CaCl2. The reaction was started by adding 0.2 mM NADPH; aliquots of 100 l were collected at intervals up to 20 min and mixed with 10 l 3 N HCl to stop the reaction. Initial rates of H2O2 formation were determined from eight aliquots of each assay by following the decrease in 0.4 M scopoletin fluorescence in the presence of HRP (0.5 g/ml) in 200 mM phosphate buffer, pH 7.8, in a Hitachi spectrofluorometer (F 4000; excitation, 360 nm; emission, 460 nm). A, P3,000 g fraction; B, microsomal fraction (P100,000 g). Enzyme activity is expressed as mean of at least two measurements in each particulate preparation. FIG. 4. Thyroid cytochrome c reductase. NADPH oxidation by cytochrome c reductase and TPO iodide oxidation activities were measured in human thyroid samples obtained from five paranodular to cold nodule tissues (PN) and from two siblings with goiter and hypothyroidism (DESM and DSM). Cytochrome c reductase activity was measured in the presence of 1 mM sodium azide, 1.2 mM EGTA, and 0.1 mM cytochrome c. The reaction was started by adding 0.1 mM NADPH; aliquots of 100 l were taken at intervals and mixed with 10 l 2 mM dithiothreitol and 100 l 2% SDS to stop the reaction. NADPH oxidation was determined from eight aliquots of each assay by following the decrease in NADPH fluorescence at pH 8.0 in a Hitachi spectrofluorometer (F 4000; excitation, 340 nm; emission, 453 nm). A, P3,000 g; B, microsomal fraction (P100,000 g). All measurements were performed on at least two samples from each particulate preparation. generated by a multicomponent enzyme, it is possible that other components of the enzymatic system responsible for H2O2 production could be involved in the impaired hormonogenesis in these goiters. Moreno et al. (31) recently described possible inactivating mutations of the ThOx2 gene in some cases of congenital hypothyroidism; however, functional studies to determine the implications of these mutations on partial iodine organification defects are still lacking. In conclusion, this is the first report of a family with goiter and hypothyroidism due to impaired NADPH oxidase activity. We have shown that the thyroid calcium-dependent References 1. Taurog A 1991 Hormone synthesis: thyroid iodine metabolism. In: Braverman LE, Utiger RD, ed. The thyroid, 6th Ed. New York: Lippincott; 51–97 2. DeGroot LJ, Niepomniszcze H 1977 Biosynthesis of thyroid hormone: basic and clinical aspects. Metabolism 26:665–718 3. Refetoff S, Dumont J, Vassart G 2001 Thyroid disorders. In: Scriver CR, Beaudet AL, Sly WS, eds. The metabolic and molecular bases of inherited disease, 8th Ed, International Edition. New York: McGraw-Hill; 4029 – 4075 4. Medeiros-Neto G, Stanbury JB 1994 Defective organification of iodide. In: Medeiros-Neto G, Stanbury JB, eds. Inherited disorders of the thyroid system. Boca Raton: CRC Press; 53– 80 5. Corvilain B, Van Sande J, Laurent E, Dumont JE 1991 The H2O2 generating system modulates protein iodination and the activity of the pentose phosphate pathway in dog thyroid. Endocrinology 128:779 –785 6. Virion A, Michot JL, Dëme D, Kaniewski J, Pommier J 1984 NADPHdependent H2O2 generation and peroxidase activity in thyroid particulate fraction. Mol Cell Endocrinol 36:95–105 7. Dupuy C, Kaniewski J, Dème D, Pommier J, Virion A 1989 NADPH-dependent H2O2 generation catalyzed by thyroid plasma membranes. Studies with electron scavengers. Eur J Biochem 185:597– 603 8. Dème D, Virion A, Aôt-Hammou N, Pommier J 1985 NADPH-dependent generation of H2O2 in a thyroid particulate fraction requires Ca⫹2. FEBS Lett 186:107–110 9. Carvalho DP, Dupuy C, Gorin Y, et al. 1996 The Ca⫹2- and reduced nicotinamide adenine dinucleotide phosphate-dependent hydrogen peroxide generating system is induced by thyrotropin in porcine thyroid cells. Endocrinology 137:1007–1012 10. Dupuy C, Ohayon R, Valent A, Noel-Hudson MS, Dème D, Virion A 1999 Purification of a novel flavoprotein involved in the thyroid NADPH oxidase. Cloning of the porcine and human cDNAs. J Biol Chem 274:37265–37269 11. De Deken X, Wang D, Many MC, et al. 2000 Cloning of two human thyroid cDNAs encoding new members of the NADPH oxidase family. J Biol Chem 275:23227–23233 12. Raspé E, Dumont JE 1995 Tonic modulation of dog thyrocyte H2O2 generation and I⫺ uptake by thyrotropin through the cyclic adenosine 3⬘,5⬘-monophosphate cascade. Endocrinology 136:965–973 13. Leseney AM, Dème D, Dupuy C, et al. 1999 Biochemical characterization of a Ca2⫹/NAD(P)H-dependent H2O2 generator in human thyroid tissue. Biochimie 81:373–380 14. Gorin Y, Ohayon R, Carvalho DP, et al. 1996 Solubilization and characterization of a thyroid Ca2⫹-dependent and NADPH-dependent K3Fe(CN)6 reductase. Relationship with the NADPH-dependent H2O2 generating system. Eur J Biochem 240:807– 814 15. Bradford MM 1976 A rapid and sensitive method for he quantification of microgram quantities of proteins utilizing the protein-dye binding. Anal Biochem 72:248 –252 16. Carvalho DP, Rego KGM, Rosenthal D 1994 Thyroid peroxidase in dyshormonogenetic goiters with organification and thyroglobulin defects. Thyroid 4:421– 426 17. Moura EG, Rosenthal D and Carvalho-Guimarães DP 1989 Thyroid peroxidase activity in human nodular goiters. Brazilian J Med Biol Res 22:31–39 18. Carvalho-Guimarães DP, Ramos CF, Rosenthal D 1989 A technical improvement for the thyroid peroxidase iodination assay. Brazilian J Med Biol Res 22:821– 823 19. Laemmli UK 1970 Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature 227:680 – 685 20. Lever EC, Medeiros-Neto GA, DeGroot L 1983 Inherited disorders of thyroid metabolism. Endocr Rev 4:213–239 21. Takeuchi K, Suzuki H, Yoshihiko H, Mashimo K 1970 Significance of iodideperchlorate discharge test for detection of iodine organification defect of the thyroid. J Clin Endocrinol Metab 31:144 –146 22. Medeiros-Neto GA, Billerbeck AEC, Wajchenberg BL, Targovnik HM 1993 Defective organification of iodide causing hereditary goitrous hypothyroidism. Thyroid 3:143–159 4848 J Clin Endocrinol Metab, October 2001, 86(10):4843– 4848 23. Ohyama Y, Hosoya T, Kameya T, et al. 1994 Congenital euthyroid goitre with impaired thyroglobulin transport. Clin Endocrinol (Oxf) 41:129 –135 24. Kim PS, Kwon OY, Arvan P 1996 An endoplasmic reticulum storage disease causing goiter with hypothyroidism. J Cell Biol 133:517–527 25. Medeiros-Neto G, Kim PS, Yoo SE, et al. 1996 Congenital hypothyroid goiter with deficient thyroglobulin. Identification of an endoplasmic reticulum storage disease with induction of molecular chaperones. J Clin Invest 98:2838 –2844 26. Bidart JM, Mian C, Lazar V, et al. 2000 Expression of pendrin and the Pendred syndrome (PDS) gene in human thyroid tissues. J Clin Endocrinol Metab 85:2028 –2033 27. Royaux IE, Suzuki K, Mori A, et al. 2000 Pendrin, the protein encoded by the Figueiredo et al. • Goiter due to Thyroid NADPH Oxidase Defect 28. 29. 30. 31. Pendred syndrome gene (PDS), is an apical porter of iodide in the thyroid and is regulated by thyroglobulin in FRTL-5 cells. Endocrinology 141:839 – 845 Kopp P 1999 Pendred’s syndrome: identification of the genetic defect a century after its recognition. Thyroid 9:65– 69 Niepomniszcze H, Targovnik HM, Gluzman BE, Curutchet P 1987 Abnormal H2O2 supply in the thyroid of a patient with goiter and iodine organification defect. J Clin Endocrinol Metab 65:344 –348 Kusakabe T 1975 Deficient cytochrome b5 reductase activity in nontoxic goiter with iodide organification defect. Metabolism 24:1103–1113 Moreno JC, Bikker H, de Randamie J, et al. 2000 Mutations in the ThOx2 gene in patients with congenital hypothyroidism due to iodide organification defects. Endocr J 47:107 Project Announcement Genetically Modified Animals in Endocrinology As a service to the endocrine community, Endocrine Reviews intends to publish bibliographies of papers describing knockout, transgenic, and mutant animals that may be useful in the study of endocrinology. In the print version of the journal, we will publish subject-limited bibliographies as the individual sections become available. We also intend to create a cumulative database to be made available on the web in a searchable format. At this time, we would like to hear what enhancements would be desirable on this web site. Readers are encouraged to contact the editorial office with bibliographic information about knockout, transgenic, and mutant animals that they would wish to have included in the database; please include the species and the citation for the article in which the original description appeared. In addition, suggestions regarding topics that we should consider adding to our bibliographies would be appreciated. Address your contributions to the database via e-mail or standard mail, using the following addresses: Dr. E. Brad Thompson/Endocrine Reviews, The University of Texas Medical Branch, Room 111C, Basic Science Building, Galveston, TX 77555-0628 USA. [email protected]