Survey
* Your assessment is very important for improving the workof artificial intelligence, which forms the content of this project
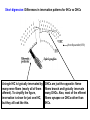
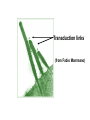
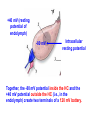
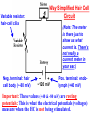
Hair Cell Transduction Electron micrograph of 3 OHCs (from Kimura, 1966) spiral ligament modiolus When the basilar membrane vibrates up and down in response to the sound wave, the hair cell cilia are displaced 1st in one direction then in the other; i.e., 1st away from the modiolus, then toward it. To state it differently, the cilia move 1st in the direction of the taller hairs, then in the direction of the shorter hairs. (Note: Shown here are OHCs, which is not inaccurate, but it is exactly this behavior in the IHCs that is of the greatest interest since the IHCs rather than the OHCs are the site of hair cell transduction.) Transduction Links: Thin filaments that connect adjacent cilia. Note that the simplified drawing here shows a single afferent fiber and a single efferent fiber. This is misleading. For the all-important IHCs, there are very few efferent fibers and many afferent fibers synapsing on a single cell (on average ~10 per IHC). Short digression: Differences in innervation patterns for IHCs vs OHCs (from Spoendlin,1979) A single IHC is typically innervated by many nerve fibers (nearly all of them afferent). To simplify the figure, innervation is show for just one IHC, but they all look like this. OHCs are just the opposite: Nerve fibers branch and typically innervate many OHCs. Also, most of the efferent fibers synapse on OHCs rather than IHCs. Another view of transduction links. They are shown in this schematic figure as tiny springs – which is how they behave (though not how they look). modiolus spiral ligament Transduction links (from Fabio Mammano) +40 mV (resting potential of endolymph) -80 mV Intracellular resting potential Together, the -80 mV potential inside the HC and the +40 mV potential outside the HC (i.e., in the endolymph) create two terminals of a 120 mV battery. When the short cilia move in the direction of the tall cilia, the transduction links are stretched. When the tall cilia move in the direction of the short cilia, the transduction links are compressed. Compressing the link squeezes the ion channel closed, inhibiting current flow. Stretching the link opens an ion channel, allowing current to flow. Note that the ion channel (also called a molecular gate) is not either open or closed. Large movement of the cilia pulls the channel way open; small movement pulls it just a little open. This is called a graded response or a continuous response – not all or none. 1. The flow of positive ions into the hair cell body is called the receptor current. 2. The receptor current stimulates the release of neurotransmitter chemicals from the hair cell into the tiny synaptic junction at the base of the hair cell. 3. The uptake of neurotransmitter chemicals by the adjacent nerve fibers stimulates the firing of the fibers – though this process is probabilistic rather than deterministic (more later). Another view of ion flow being regulated by the movement of cilia in just one direction: Short hairs moving in the direction of long hairs, opening the molecular gate (not shown) and allowing positively charged ions (the yellow stuff in the animation below) to flow into the hair cell. The receptor current is graded or continuous, completely unlike the all-or-none behavior of neurons. What does that mean? Just this: when the instantaneous amplitude of the signal is large the displacement of the basilar membrane will be large the displacement of the hair cell cilia will be large the receptor current will be large (because the molecular gate or ion channel will be way open) the quantity of neurotransmitter chemicals dumped into the synaptic junction will be large and finally, the probability of a pulse on the 8th N will be high (not certain) BUT: The 8th N fiber will either fire or not, in its usual all-or-none fashion Similarly: when the instantaneous amplitude of the signal is small the displacement of the basilar membrane will be small the displacement of the hair cell cilia will be small the receptor current will be small (because the molecular gate or ion channel will be just slightly open) the quantity of neurotransmitter chemicals dumped into the synaptic junction will be small and finally, the probability of a pulse on the 8th N will be low (not zero) BUT once again: The 8th N fiber will either fire or not, in its usual all-or-none fashion Variable resistor: hair-cell cilia Neg. terminal: hair cell body (~-80 mV) Way Simplified Hair Cell Circuit (Note: The meter is there just to show us what current is. There’s not really a current meter in your ear.) ~120 mV Pos. terminal: endolymph (+40 mV) Important: These values (+40 & -80 mV) are resting potentials: This is what the electrical potentials (voltages) measure when the HC is not being stimulated. (Brief Digression) Hallowell Davis Hallowell Davis, former director of the Central Institute for the Deaf in St. Louis, and a pioneer in hearing research. In 1939 Hallowell Davis and Robert Galambos made the 1st recordings of the electrical activity of individual neurons. That’s 1939 – the year in which, among many other things, WWII started and The Wizard of Oz (a nearly perfect movie) and Gone with the Wind (a terrible movie, according to at least one crackpot) were released. Our understanding of hair cell physiology has advanced enormously since Davis’ early work, but current models of hair cell function are very closely related to a model described by Davis in 1963. Relationship between the input signal and the receptor current for a sinusoid (left) and a complex periodic signal (right). Note that the 2 receptor current and the input signal look similar, except that the bottom half is missing. This is called half-wave rectification. Why is the bottom half of the signal missing in the receptor current? Input Signal Input Signal Receptor Current Receptor Current OHCs: Why do we have them at all? Transduction is carried out by the IHCs, right? What’s the point of having OHCs? watch this cell Phenomenon is called hair-cell motility (motility=movement). Only mammals have evolved HCs that exhibit this kind of motility. Why? Frequency ranges for a few mammals (with OHCs) Whales: Bats: Humans: 20 - 100,000 Hz 1,500 - 100,000 Hz 20-20,000 Hz Frequency ranges for a few non-mammals (without OHCs) Frogs: Fish: Crickets: Birds: 600 – 3,000 Hz 20 – 3,000 Hz 500 – 5,000 Hz Variable across species, but top end usually well below 10,000 Hz What’s the big difference? (A: Much improved high freq hearing) For what purpose did mammals evolve OHCs?