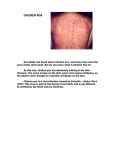
Survey
* Your assessment is very important for improving the workof artificial intelligence, which forms the content of this project
Botanical Journal o f t h e Linnean Society. 74: 131.162 . With 7 figures February 1977 The delimitation of the tribe Vicieae (Legurninosae) and the relationships of Cicer L . FRANCES K . KUPICHA. F.L.S. Department o f Botany. British Museum (Natural History). Cromwell Road. London SW7 5BD Accepted f o r publication September 1976 A survey of morphological. anatomical. karyological and chemical characters has been carried out. centred o n the Vicieae b u t extending t o the neighbouring tribes Trifolieae and Ononideae . The results show that Cicer. traditionally a member of the Vicieae. has more in common with genera of the Trifolieae and Ononideae than with the rest of the Vicieae . I t is proposed that Cicer should be removed from the Vicieae and recognized as t h e monogeneric tribe Cicereae Alef . The tribe Vicieae sensu stricto. a well-defined natural group. is delimited and described . Phylogenetic relationships of the Cicereae are discussed . KEY WORDS:-Cicer-Cicereae-Vicieae-Trifolieae-Ononideae-Leguminosae-morphology-cytology-chemotaxonomy . CONTENTS Introduction . . . . . . . . Historical background . . . . . Taxonomic characters . . . . . Habit . . . . . . . . The seedling . . . . . . Germination . . . . Position of first leaf . . Ontogeny . . . . Vascular anatomy . . . . In the mature plant . . In theseedling . . . The leaf . . . . . . . Leaflet shape and venation . . . . . Stipules Form of the mature leaf . . Number of leaflets Ontogeny . . . . Evolution of the leaves . lndumentum . . . . . . Inflorescence . . . . . Theflower . . . . . . Corolla . . . . . Androecium . . . . Pollen . . . . . . Gynoecium . . . . The fruit . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 131 132 132 135 135 135 136 136 138 139 139 140 140 140 141 141 141 141 143 143 146 146 146 146 147 147 147 132 F. K. KUPICHA The seed . . . . . . . . . . . Seed shape . . . . . . . . Milurn length . . . . . . . . Endosperm . . . . . . . . Vascular supply . . . . . . . The liarotype: chromosome number . . . Chcniosysternatics . . . . . . . . Anthocyanins . . . . . . . Isoflavonoids . . . . . . . . Nowprotein amino acids: canavanine Seed proteins . . . . . . . Discussion . . . . . . . . . . . . T h e delimitation of the Vicieae . . . . Description of t h e tribe Vicieae sensu strict0 T h e taxonomic position of Cicer . . . . Phylogenetic relationships of Cicer . . . Acknowledgements . . . . . . . . . . Rcferences . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 148 148 149 149 149 . . . . . . . . . . 150 . . . . . . . . . . . . . . . . . . . . 152 152 152 153 154 155 155 157 157 158 159 160 . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . INTRODUCTION The tribe Vicieae contains several economically important species ( Vicia faba L.-broad bean; Lathyrus odoratus L.-sweet pea; Pisum sativum-garden pea; Lens culinaris-lentil and Cicer arietinum-chickpea) and consequently has been much studied. Its members are found throughout the north temperate zone and to a lesser extent in South America, and modern taxonomic accounts of most of its genera have been published in Floras covering the chief centres of distribution, as well as in monographic accounts. Despite this concentrated activity, no-one has recently taken a broad view of the tribe, whose delimitation is problematic. This work sets out to define the problem, survey the available taxonomic evidence, and achieve a more natural tribal classification. Authorities for species are given on first mention in the text except for species listed in Tables 3 and 5 . HISTORICAL BACKGROUND The first post-Linnean taxonomist to recognize the Vicieae was Adanson (1763), who called this entity a ‘section’, and included in it Tournefort’s genera Aphaca, Cicer, Clymenum, Ervum, Lathyrus, Lens, Nissolia, Orobus, Pisurn and Vicia. De Candolle was the first to give the Vicieae tribal rank. He stated (1825a and 182513: 353) that the essential characters of the tribe are 1, germination hypogeal; 2, first plumular leaves alternate, never opposite; 3 , leaves epulvinate; 4, leaves paripinnate, with the rachis prolonged into a tendril, or (in some species of Cicer) leaves imparipinnate. The genera which De Candolle accepted as members of the tribe were Cicer L., Ervum L., Faba L., Lathyrus L., Orobus L., Pisum L. and Vicia L. The number of genera into which the species of the Vicieae are grouped has tended to decrease with time, at least until very recently. Bentham (1865: 524) recognized six, viz. Cicer, Vicia, Lens Miller, Lathyrus and Pisum (comprising the Vicieae sensu DC.), and Abrus L. He was uncertain whether Abrus was correctly placed in the Vicieae, as some of its characters suggested affinity with DELIMITATION O F THE VICIEAE-LLEGUMINOSAE 133 the Phaseoleae or Dalbergieae. Bentham’s decision to include Abrus was followed by several taxonomists, e.g. Taubert (1894: 350), Ascherson & Graebner (1909: 898) and Gams (1924: 1497);but on the other hand, various workers who made comparative morphological, anatomical and karyological studies in the Leguminosae, e.g. Streicher (1902: 493), Popov (1928-29: 236), Senn (1938: 273) and Dormer (1946: 161), advocated that Abrus should be excluded from the Vicieae. In the most recent generic and tribal revision of the family Hutchinson (1964: 451) placed Abrus in its own tribe, the Abreae, immediately before the Vicieae. Table 1 shows that the decision to separate Abrus from the Vicieae is amply supported by the evidence of many differential characters. Table 1. Characters separating Abrus from the Vicieae Abrus 1. plants usually woody 2. stems usually twining 3. first two seedling leaves opposite 4. stems not containing cortical vascular bundles 5. phyllotaxy polymerous 6. leaves with pulvinus at base of petiole 7. stipels present 8. leaves not tendrillous 9. flowers borne in ‘pseudoracemes’ (panicles with very contracted branches), these terminal o n leafy branches 10. vexillum united with staminal tube 11. androecium with 9 stamens 12. pollen grains with unthickened endoapertures and microreticulate ornamentation (provisional, based o n one specimen of A. precatorius L; cf. Clarke & Kupicha, 1976: 43) 1 3 . style glabrous 14. germination epigeal 15. haploid chro’mosome number: n=ll 16. chromosomes relatively small 17. geographical distribution: pantropical * Vicieae 1. 2. 3. 4. 5. 6. 7. 8. 9. 10. 11. 12. plants herbaceous, rarely woody stems not twining first two seedling leaves alternate stems containing cortical vascular bundles (except in Cicer) phyllotaxy dimerous leaves with epulvinate petiole stipels absent leaves often tendrillous flowers in axillary racemes, very rarely in panicles vexillum free from staminal tube androecium with 10 stamens pollen grains with thickened endoapertures and a widely reticulate ornamentation (except in Cicer) 13. 14. 15. 16. 17. style pubescent (except in Cicer) germination hypogeal haploid chromosome number: n = 5, 6 , 7, 8 chromosomes relatively large geographical distribution: temperate Eurasia and North America, temperate South America Generic limits in the Vicieae are still a matter of debate. The genera which I recognize as constituting the Vicieae sensu DC. are Cicer (whose position is discussed in this paper), Vicia, Lathyrus, Lens, Anatropostylia (Plitm.) Kupicha, Pisum and Vavilovia Feodorov. Apart from the controversy over Abrus, the boundaries of the Vicieae have remained constant since the early 19th century. At the same time, many authors have commented o n the isolated position of Cicer within the tribe, and have speculated on its possible relationship with other parts of the Papilionoideae. De Candolle (1825a) himself pointed out the apparent affinity of Cicer and Ononis L.: “The aristate peduncle is not the only similarity between these two genera. The sharp teeth of the leaflets and stipules of Cicer, F. K. KUPICHA 134 the form of its hairs, the arrangement of the flowers, the shape of the calyx and corolla and the appearance of the fruit, all recall certain species of Ononis, especially those with pinnate leaves. The swollen cotyledons and mode of germination are the best characters to distinguish Cicer from Ononis.” (translated). Ononis has had a very unstable taxonomic history. In the earliest classifications, e.g. that of Bronn (1822: 132), it was associated with members of the Genisteae. Bentham (1865: 485) considered that Ononis belongs somewhere between the Genisteae and Trifolieae, because it resembles the former in its monadelphous androecium and dimorphic anthers, and the latter in its leaflets, inflorescence and filament apices; on balance, he decided to place Ononis in the Trifolieae. Gams (1924: 1216) believed that Ononis has no true connections with either the Trifolieae or the Genisteae, but is an advanced genus related to Cicer and thus indirectly to the rest of the Vicieae. However, although he expressed this opinion, he placed Ononis in its traditional taxonomic position. In his monograph of Ononis, Sirjaev (1932: 414) concluded that the intermediate characteristics of this genus justified the creation of a separate tribe, the Ononideae. Hutchinson (1964: 454) also recognized the Ononideae; in his system it comprises Ononis and the monospecific segregate genus Passaea Adanson (P. ornithopodioides (L.) Adanson differs from other members of the Ononideae only in having internally septate fruits, and it is treated here as a species of Ononis). According to Hutchinson, the Trifolieae include six genera: Parochetus Buch.-Ham., Melilotus Miller, Trigonella L., Factorovskya Eig, Medicago L. and Trifolium L. The features which distinguish the Ononideae and the Trifolieae are listed in Table 2. I doubt whether these differences are really sufficient t o maintain the two tribes. However, in the present context, where I am considering the relationships of Cicer, it is most convenient to recognize the Ononideae as a taxon parallelling the Trifolieae sensu stricto. Popov (1928-29: 107, 236) was impressed by the phenetically intermediate position of Cicer between the Vicieae and Ononis, and proposed the eccentric hypothesis that in the Paleocene epoch, Ononis appeared as a hybrid between members of the Genisteae and Trifolieae. At this time Ononis and Vicia were vicarious genera, separated by the Tethys sea and belonging, respectively, to a xerophytic African and a mesophytic Asian flora. Geological and climatic Table 2. Characters separating the Trifolieae and Ononideae Trifolieae androecium diadelphous anthers monomorphic 3. anthers all versatile 4. plants never spiny 5 . inflorescence usually capitate or spicate 6 . legume not inflated, but of various characteristic forms 7. pollen grains oblong-ellipsoid 3. 4. 5. 6. 8. 8. 1. 2. haploid chromosome number: n=8 1. 2. 7. androecium monadelphous anthers of two sizes anthers alternately basifixed and versatile plants often spiny inflorescence often 1-few-flowered legume inflated pollen grains subspherical (Clarke & Kupicha, 1976) haploid chromosome number: n = 1 5 , 16 (see Table 4) DELIMITATION O F THE VICIEAE-LEGUMINOSAE 135 changes then allowed these floras to come into contact, and crossing took place between Vicia and Ononis; the first generation hybrid, ‘Onono-Vicia’ underwent selfing and produced a number of segregants comprising the two parental types and four intermediates, each of which was the ancestor of one of Popov’s four sections of Cicer. Popov decided that, in spite of its equal affinity with Ononis, Cicer should be included in the Vicieae according to the usual taxonomic practice. In the light of modern evolutionary theory, this story cannot be accepted as a serious explanation of the origin of Cicer, or any other genus; if this kind of wide and free mixing of genetic material were possible, genera would inevitably lose their distinctness and their status. Popov’s theory also implies that hybridization at generic level must be common in the Papilionoideae, but this is not the case (McComb, 1975). Several authors (e.g. Senn, 1938: 249, 273; Simola, 1969: 655) have quoted Popov’s conclusions about the origin and taxonomic relationships of the genus which he monographed, apparently accepting his authority without questioning the reasoning behind these ideas. Having examined and rejected the hypothesis which underlies his systematic treatment of Cicer, one is left with the unsolved problem which Popov indicated so clearly: what are the relationships of Cicer? This is the most critical issue in the Vicieae, as presently defined. TAXONOMIC CHARACTERS The following paragraphs discuss all the known characters whose variation provides evidence about the relationship of Cicer with the Vicieae, Trifolieae and Ononideae. As there is no doubt that Cicer’s closest affinities lie within these groups, other tribes are not considered here. Habit All members of the Vicieae, Trifolieae and Ononideae are herbs, ranging from delicate, short-lived annuals to perennials with welldeveloped secondary thickening. Cicer and Ononis are remarkable for their tendency towards spininess, a trait not found in the other genera. The spines are variously formed from modified stipules, leaflet-teeth, tendrils (in Cicer), bracteoles and the sterile apex of peduncles. The seedling During the early part of this century many papers reported observations on the germination of legumes, and members of the Vicieae were often represented. It is regrettable that very little of this new information was noted by taxonomists, and the interest in seedling characters apparently died away leaving a residue of data about unevenly surveyed taxa, some of the findings contradictory but some indicating that seedlings can yield significant taxonomic characters. The following paragraphs summarize most of the variable features found so far in the seedlings of the Vicieae and neighbouring tribes, and show that certain patterns have emerged which are relevant to the classification of Cicer. The recent study of Baudet (1974) in the Phaseoleae F. K. KUPICHA 136 demonstrates that comparable variation-patterns are present in other parts of the Papilionoideae. Germination All members of the Vicieae, including Cicer, have hypogeal germination, while the Trifolieae and Ononideae are exclusively epigeal. Compton (1912-1913: 91), who surveyed the whole of the Leguminosae for this character, found that within the Papilionoideae the Sophoreae are mostly hypogeal; the Podalyrieae, Genisteae, Galegeae, Loteae and Dalbergieae are uniformly epigeal; the Hedysareae are almost entirely epigeal, and the Phaseoleae show both types in abundance. The Vicieae are thus distinctive in having uniformly hypogeal germination, and it is not surprising that this character has been a heavily-weighted tribal feature. Having observed the germination of about 20 species of the Vicieae (Table 3 ) , I found that Cicer differs from the other genera in the direction of growth of the radicle. In Vicia, Lathyrus, Lens and Pisum the radicle always grows straight down past the hilum and the plumule emerges from between the cotyledons on the side of the cotyledonary petioles away from the hilum (Fig. l A , B). In Cicer the radicle has a variable behaviour: in some seeds it grows past the hilum (Fig. lC, D), but in others it turns in the opposite direction and then the plumule emerges on the hilar side of the cotyledonary petioles (Fig. l E , F). As only two species of Cicer have been studied for this character it is not known whether the variable direction of radiclegrowth is a constant generic feature. It would be interesting to complete the survey and to extend it to the neighbouring tribes. Position of first leaf My observation of the species listed in Table 3 showed that in Vicia, Luthyrus, Lens and Pisum the first leaf of the plumule, which is a small scaIe-leaf, is always borne on the side of the epicotyl away from the cotyledons (the ‘convex’ side of the shoot). This fact was noted by van Tieghem (1871: 220) and Compton (1912-13: 86, 107). The illustrations of seedlings of the Table 3. Species of the Vicieae in which the first-leaf position in the seedling has been observed Cicer arietinum L. CC, CV C. pinnatifidum Jaub. & Spach Lathyrus cicera L. CV L . inconspicuus L. CV L. nissolia L. CV L . tingitanus L. CV L. sphaericus Ketz. CV Lens culinaris Medicus Pisum sativum L. CV CV CC, CV Vicia americana Miihl. e x Willd. V. amurensis Oettel CV V . anatolica T u r d CV V . biennis L. CV V. bithynica (L.) L. CV V. cracca L. CV V. dumetorum L. CV V. ervilia (L.) Willd. CV V . hybrida L. CV V. lutea L. CV V. peregrina L. CV V. pubescens (DC.) Link CV V . sativa L. CV CC, first scale-leaf on concave side of plumule; CV, on convex side. CV DELIMITATION O F THE VICIEAE--LEGUMINOSAE 137 Vicieae in Csapody (1968: 261-265) show most with the first scale in this ‘convex’ position; in the remaining cases, it may be that the plumules were twisted or drawn from a misleading angle. There is disagreement in the literature about the situation in Cicer: van Tieghem (1871: 219) said that in C. arietinum the first leaf is inserted on the side of the plumule nearest the cotyledons (the ‘concave’ side), while Muller (1937: 203) stated that it is produced on the convex side. I found that in both C. arietinum and C. pinnatifidum the first leaf varies in position; in some seedlings it is on the convex side of the plumule, in others it is on the concave side. As the radicle may grow either past or away from the hilum, there are four possible kinds of seedling, and these are illustrated in Fig. 1C-F. Compton apparently did not observe the germination of any species of Cicer, for he concluded that the Vicieae have a very uniform pattern of germination, characterized by the constant insertion of the first leaf on the convex side of the plumule. He emphasized this, and contrasted the Vicieae with the Hedysareae, Galegeae, Trifolieae and Loteae, whose members, with few Figure 1. Germinating seeds; in each seedling the first plumular leaf is indicated by an arrow. A , Vicia f a b ~ B, ; Lens culinaris: in A and B the first leaf is borne on the “convex” side of the shoot. C-F, Cicer pinnatifidum: in C and D the radicle grows past the hilum, in E and F it grows away from the hilum; in C and F the first leaf is borne towards the cotyledons, in D and E it is borne away from the cotyledons. G-I, Trigonella foenum-guuecum: G , diagrammatic longisection of seed (endosperm not drawn), with the ‘convex’ side of the cotyledon marked with a dot; H and I , seedlings with the first leaf appearing in opposite positions. Not to scale. 138 F. K . KUPICHA exceptions, were believed to have the first leaf inserted on the concave side of the epicotyl (Compton, 1912-13: 107). This character is difficult to observe in epigeal seedlings, because once the testa has been shed from the cotyledons these straighten up and become completely symmetrical, leaving no indication of their original orientation in the seed. In order t o discover the spatial relationship of the cotyledons and the first leaf one must mark the cotyledons during germination to preserve their initial asymmetry. I have followed the germination of only one species outside the Vicieae, Trigonella foenumgraecum L.; it was found that the first leaf is sometimes borne on the concave side of the plumule, sometimes on the convex side (Fig. l H , I). This suggests that the Trifolieae may be characterized by a variable first-leaf position rather than a constant concave-side insertion as Compton postulated, and in this case Cicer would resemble the Trifolieae in this character. The invariable position of the first leaf in the Vicieae (apart from Cicer) is correlated with a very uniform type of vascular symmetry in seedlings of this group (Compton, 1912-13: 86). The root and hypocotyl are triarch; two of the protoxylem strands supply the cotyledons, which are borne at an angle of 120°, while the third continues in the plumule to become the median trace to the first scale-leaf. Cicer arietinum, however, is tetrarch, and the cotyledons are inserted at 180" (Muller, 1937: 203). The presence of two intercotyledonary protoxylem strands in Cicer may explain why the first leaf is able to appear on either the concave or convex side of the plumule. This relationship between primary vasculature and leaf-insertion is not absolute, however; Vicia faba, which always has the first leaf on the convex side of the plumule, is exceptional in having tetrarch seedlings (fide Compton and Muller). The situation in the Trifolieae and Ononideae may not be strictly comparable with that in the Vicieae; according t o Compton (1912-13: l O l ) , in their slender epigeal seedlings the protoxylem of the cotyledons, hypocotyl and radicle forms a self-contained unit and is only connected by metaxylem with protoxylem of the plumule (Erith (1924: 14) disagreed with Compton in the case of Trifolium repens L. and stated that in this species there is a protoxylem connection between radicle and plumule). Compton found that in the Trifolieae and Ononideae the seedling vasculature has an unstable symmetry, varying from tetrarch to triarch or diarch often within a species or even at different levels in the axis of a single specimen. If there is a causal relationship between the vascular symmetry of the embryo and the position of early leaves on the plumule, these data would be consistent with a variable first-leaf position in the Trifolieae and Ononideae. No firm taxonomic conclusions can be drawn from the information available on this subject at present; a more thorough survey in these tribes is needed to elucidate the situation. Ontogeny In seedlings of Vicia, Luthyrus, Lens and Pisurn the primary shoot rarely attains more than a few inches in height before its growth ceases, and the bulk of the plant is formed from vigorous secondary branches arising from the axils of the scale leaves at the lowest nodes (Vicia faba and its close relatives are exceptional in having indeterminate primary stems). My observations on Cicer arietinum, C. pinnatifidum and C. anatolicum Alef. show that in these species the plumular shoot continues to develop throughout the growing season, DELIMITATION O F THE VICIEAE-LEGUMINOSAE 139 although the basal nodes often give rise to lateral branches which may overtop the primary stem. This aspect of ontogeny is difficult to measure in herbarium specimens, and without further studies on living material I cannot tell whether the apparent difference in growth-pattern between Cicer and the rest of the Vicieae has any significance. I have no parallel information about ontogeny in the Ononideae and Trifolieae. I t would be very interesting to know if there is systematic variation in this character within or between these tribes. Vascular anatomy In the mature plant In Cicer and members of the Trifolieae and Ononideae the three leaf-traces which supply each petiole are all emitted from the main stele at the same level (Fig. 2C, D; see also Winter, 1932). The remaining genera of the Vicieae are M M Figure 2. Cauline vascular systems in the Vicieae. A-B, Viciu f a b u : A, diagrammatic reconstruction of nodal vascular anatomy, showing supply to stipules and petiole (for simplicity the main stele is drawn as a solid cylinder); B, transverse section through internode, showing the stele as a ring of primary bundles with two additional cortical bundles. C-D, Cicer ariefinum: C, reconstruction of nodal vascular anatomy; D, transverse section through internode, showing absence of cortical bundles. L, Lateral leaf-trace; M, median leaf-trace; S, traces to stipules; Bud, traces t o the axillary bud. 140 F. K. KUPICHA characterized by an unusual type of cauline vasculature in which the lateral leaf-traces are present in the internode below the insertion of each leaf as a pair of cortical bundles (Fig. 2A, B; see also Kupicha, 1975). This important difference between Cicer and the rest of the Vicieae has been pointed out by several authors, e.g. van Tieghem (1884a: 1 3 3 ) , Hkrail (1885: 223, 306) and Dormer (1946: 161), but as yet it has had no influence on the tribal classification. In the seedling The seedlings of Vicia, Latlzyrus, Lens and Pisum are remarkable in having a very prolonged transition region between root and stem. The first two plumular internodes contain a solid core of xylem which is exarch in the first internode above the cotyledons; the construction of the shoot becomes progressively more stem-like with the emission of successive endarch median leaf-traces to the first, second and third scale-leaves (Compton, 1912-13 ; Gourley, 1931; Muller, 1937). (The lateral leaf-traces are supplied from the two cortical bundles which are given off from the cotyledonary leafgaps, as described by Kupicha, 1975 .) This situation is anomalous within the Leguminosae and among dicotyledons in general. Compton suggested that the rootlike structure of the young epicotyl may be an adaptation enabling this region to bend, in relation to the climbing habit of the Vicieae. His hypothesis may be supported by the fact that Vicia faba, which has-a medullated and therefore stiffer transition region, is a non-climber with etendrillous leaves. According to Muller (1937), in Cicer ilrietinum the transition region between root and stem is in the normal hypocotyledonary position and there is neither a solid core of xylem nor any exarch protoxylem in the epicotyl; in this respect Cicer resembles the Trifolieae and Ononideae, as far as is known. The leaf Leaflet shape and venation Cicer, and genera of the Trifolieae and Ononideae, typically have serrate leaflets in which the lateral veins end blindly in the marginal teeth (Fig. 3B). This kind of venation is termed craspedodromous (Stearn, 1966: 541). Many Figure 3. Leaflet shape and venation. A, Vicia sepium L.: entire leaflet with brochidodromous venation. B, Cicer anatolicum: serrate leaflet with craspedodromous venation. DELIMITATION O F T H E VICIEAE-LEGUMINOSAE 141 species of the Trifolieae have entire leaflets, but the venation type is constant throughout the tribe. Craspedodromous venation is rare in the Papilionoideae; as far as I know, it is confined to these groups. In contrast, the remaining genera of the Vicieae are characterized by entire leaflets with the very common brochidodromous venation pattern in which each lateral nerve curves towards the apex as it approaches the margin and unites with its neighbour (Fig. 3.4). A very few taxa have serrate leaflets (e.g. Vicia grandiflora var. dissecta, V. serratifolia Jacq.), but here the teeth are not provided with a main vein as in Cicer. Stipu lcs The Trifolieae and Ononideae have leaves with the stipules adnate to the petiole; in all members of the Vicieae, including Cicer, the stipules are free. Form of the mature leaf The most striking vegetative feature of the Vicieae is their tendrillous leaves. Tendrils are absent from the rest of the Papilionoideae, and their occurrence in this group, coupled with the almost equally unusual hypogeal type of germination, has been thought of overriding significance in considerations about tribal limits in this area of the subfamily. Although tendrils are so characteristic of the tribe Vicieae, many species are etendrillous. In Vicia, Lathyrus and Vavilovia, species with etendrillous adult leaves almost always have paripinnate leaves in which the rachis ends in a mucro; only a very few species of Vicia (V. abbreviata Fischer ex Sprengel, V. argentea Lapeyr., V. serniglabra Rupr. ex Boiss. and V. subvillosa (Ledeb.) Trautv.) have leaves with a terminal leaflet. By contrast, in Cicer there are no species with paripinnate leaves; here the leaves are either tendrillous or imparipinnate. As will be shown, this difference between Cicer and the rest of the Vicieae is also reflected in disparate patterns of foliar ontogeny, and the latter in turn suggests that the two groups are at different stages with respect to the evolution of tendrils. Number of leaflets Most species of Cicer have multijugate leaves, although C. incisum (Willd.) K. Maly has leaves with 3-7 leaflets and C. chorassanicum (Bunge) Popov is uniformly trifoliolate. The rest of the Vicieae vary widely from unijugate to multijugate. Apart from a few species in Trifolium sect. Lupinaster with palmately 5-9-foliolate leaves, the Trifolieae are all trifoliolate. Ononis includes a few species with pinnate leaves (e.g. 0.fruticosa L.), but most of its members have leaves with one or three leaflets. Ontogeny Having observed seedlings of some 70 species of the Vicieae, I found that ontogeny in the tribe is very uniform; all genera, except for Cicer, are characterized by the same pattern of development. The first two or three nodes of the seedling bear a trifid scale-leaf. The next leaf comprises a pair of stipules, a petiole and a pair of leaflets, the rachis ending in a minute point (Fig. 4A: J l ) . In species with unijugate etendrillous adult leaves (e.g. Lathyrus roseus Phil., Vicia unijuga A. Braun and Vavilovia formosa (Steven) Fedorov) further F. K . KUPICHA 142 P Figure 4. Ontogenetic leaf-series. A, Vicia melnnops Sibth. & Sm. B , Cicernnarolicum. J1, First juvenile leaf of the plumule (which follows two or three scale-leaves); 54 and J5, fourth and fifth juvenile leaves; M, mature leaf. Scale: x * j 3 . development towards the mature leaf-form simply involves an increase in size. In other species successive juvenile leaves have additional pairs of leaflets, while the rachis is extended past the distal pair and becomes tendril-like, until the mature multijugate tendrillous leaf is achieved (Fig. 4A: J5,M). There are two exceptions to this normal pattern of ontogeny. Firstly, Lathyrus includes four species (L. clymenum, L. gloeospermus Warb. & Eig, L. ochrus (L.) DC. and L. nissolia) in which the juvenile leaves are phyllodes; in the first three, adult leaves develop leaflets, but in L. nissolia the phyllodic condition persists throughout. Secondly, as mentioned above, there are a few species of Vicia with imparipinnate leaves. I have seen the development of only one of these, V. semiglabra. Among a small sample of seedlings it was found that the first true leaf varied in construction from trifoliolate to more rarely unifoliolate or unijugate. In every case successive leaves were imparipinnate, but, especially in the lowest ones, the terminal leaflet was slightly asymmetrically placed, suggesting that the leaf might be secondarily imparipinnate, the distal leaflet having been displaced from a lateral position. There was, however, no sign of a mucro at the end of the rachis, and the distal leaflet was apparently truly terminal. In Cicer, the first leaf to appear after the scales of the earliest nodes always has two or more pairs of lateral leaflets and a terminal leaflet (Fig. 4B: J 1). (I have observed this in C. anatolicum, C. arietinum and C. pinnatifidum, while L.J.G. van der Maesen (pers. comm.) has very kindly supplied information DELIMITATION OF THE VICIEAE-LEGUMINOSAE 143 about C. bijugum K. H. Rechinger, C. chorassanicum, C. judaicum Boiss., C, pungens Boiss. and C. microphyllum Royle.) In tendrillous species of Cicer the terminal leaflet is replaced by a tendril in leaves of the adult plant (Fig. 4B: M). In several species, e.g. C. anatolicum, C. baldskuanicum (Popov) Lincz., C. graecum Orph. ex Boiss. and C. songaricum Jaub. & Spach, this transition may be represented in intermediate leaves by the presence of leaflets whose midribs, and sometimes even the lateral veins, are produced into rudimentary tendrils. Evolution of the leaves I t is generally agreed that the ancestral form of leaf in the Papilionoideae was imparipinnate, with many pairs of lateral leaflets, and this has been stated specifically in connection with the Vicieae by van Tieghem (1884b: 339) and Reinke (1897: 553), who said that the tendrils in this tribe evolved by modification of the terminal and lateral leaflets. Simola (1968: fig. 22) and Bassler (1973: fig. 3) both believe that the appearance of paripinnate adult leaves in Lathyrus is an advanced feature, being due to loss of the terminal tendril. I concur with these authors, but I suggest that if, in addition, patterns of ontogeny are used to interpret the evolution of leaves in the Vicieae, the conclusions are more interesting and have a bearing on the relationship of Cicer with the other genera. Since the supporting function of tendrils is much more relevant to the adult plant than the seedling, it seems reasonable to suppose that the selection of tendrils was focused on the adult stage of the life-cycle and that the occurrence of reduced, paripinnate leaves in seedlings of Vicia, Lathyrus etc. is a secondary effect consequent on the evolution of tendrils in mature leaves. I t seems apparent that in all genera of the Vicieae apart from Cicer the evolutionary reduction sequence ‘leaves imparipinnate -+ tendrillous +.paripinnate’ has influenced the entire ontogenetic development of the plants so that the juvenile leaves show the most reduced and specialized form (Fig. 4A: J1). In species with unijugate mucronate adult leaves, paedomorphosis maintains this simple state throughout the life of the plant. In species with multijugate and/or tendrillous adult leaves, the more primitive, more elaborate type of leaf develops towards maturity. In Ciceu, however, the evolutionary specialization of adult leaves is curtailed at the tendrillous stage (i.e. ‘leaves irnparipinnate tendrillous’), and this trend has only influenced the later part of the ontogenetic pathway; in tendrillous species of Cicer the juvenile leaves are more primitive in form than the adult leaves (Fig. 4B). The occurrence of a few species of Vicia with imparipinnate leaves and a Cicer-like ontogeny is interesting and problematic. These species are not all closely related, and they are not particularly primitive within the genus (Kupicha, 1976). I t is conceivable that they are secondarily imparipinnate. On the other hand, they may represent isolated examples of the ancestral condition in the Vicieae. --+ Indumentum The different kinds of hair found in the Vicieae have been described by Shah & Kothari (1973), and I agree with their observations. The following account 144 F. K . KUPICIIA amplifies their data, which were based on only a few species, and broadens the survey to include the neighbouring tribes. I have examined most species of the Vicieae, and many of the Ononideae and Trifolieae. Most members of Vicia, Lathyrus, Lens, Pisum, Vavilovia and Anatropostylia have sparsely hairy leaves, the hairs usually being most abundant on the lower epidermis and along the midvein of the leaflets. The indumentum in these genera is of three types, comprising unicellular hairs (Fig. 5C) and short-stalked glands (Fig. 5A, 13). The simple hair varies considerably in length but is otherwise very uniform, having a thin wall which is always smooth. I t arises from an isodiametric, straight-walled cell which projects slightly above the general level of the epidermis. At the base of the hair is often found a granular cytoplasmic body which can falsely give the impression of a separate cell. Pisum and Lathyrus tend to have nearly glabrous leaves; in Vicia and Lens they are more densely hairy. Occasional species (e.g. Lathyrus macropus Gill., Vicia canescem Labill., Lens montbretii Fischer & Meyer) have villous leaves. Vicia lutea, and a few species of Lathyrus (L. trachycarpus Boiss., etc.) have legumes with tuberculate-based hairs. The hairs themselves are of the same type as that shown in Fig. 5C, but the epidermal cell subtending each hair is produced into a prominent tubercle. The glandular hair illustrated in Fig. 5A is as widely distributed as the simple hair, occurring sparsely on the leaves of most species, and on the young legumes of some members of Lathyrus. The cells of the head, which contain dense, brownish, granular cytoplasm, are usually eight, and there is always a single stalk cell. The glandular hair shown in Fig. 5B was found in only two species, both in Lathyrus: on the young shoots of L. odoratus, and on the fruits of L. cassius Boiss. Figure 5 . The different types of indumentum found in the Vicieae, Ononideae and Trifolieae. A and B, Short-stalked glandular hiars; C, unicellular, smooth-walled hair; D, unicellular, papillate-walled hair; E, long-stalked glandular hair; F, multicellutar hair. DELIMITATION OF THE VICIEAE-LEGUMINOSAE 145 The remaining genus, Cicer, is very distinct within the tribe in possessing glands with multicellular stalks (Fig. 5E). These trichomes have been studied in detail by Schnepf (1965). Their exact form and dimensions vary considerably, due both t o specific differences and to their stage of growth. It was observed that they grow acropetally, beginning as a simple filament; the distal cell then starts to divide both transversely and longitudinally, until the mature head is formed. The hair arises from a large, convex cell which protrudes above the neighbouring cells of the epidermis, and the cuticle is wrinkled up around it. While this type of hair is confined to Cicer within the Vicieae, it is found also in the Ononideae and some species of Medicago and Trigonella. This distribution pattern (Table 4) is one of the strongest lines of evidence from vegetative characters suggesting that Cicer is related to the Ononideae and Trifolieae. It is interesting that this unusual glandular hair is associated with the equally rare craspedodromous type of venation. Unfortunately, however, the taxonomic evidence provided by indumentum is not straight-forward. Besides the long-stalked glands, Cicer also has simple, smooth-walled hairs like those of the rest of the Vicieae (sometimes these have very thick walls, as in C. incisurn). Members of the Trifolieae also have simple hairs, but my survey has shown that these always have papillate walls (Fig. 5D), and the basal cell does not project above the surrounding epidermis as in the Vicieae. In Ononis there are no simple hairs, only multicellular hairs (Fig. 5F) which are obviously related t o the long-stalked glands. Multicellular hairs are not found in Cicer, but occur in some species of Medicago. The short-stalked gland (Fig. SA) occurs throughout the Vicieae, Trifolieae and Ononideae, and is of no systematic value within them. Table 4 shows that the distribution patterns of these indumentum types are taxonomically very interesting. On one hand, Cicer is linked with the Ononideae and Trifolieae by the long-stalked glands; on the other, the microcharacter involving the wall-structure of the unicellular hair (smooth or papillate) supports the present systematic arrangement. The absence of simple Table 4. Distribution of indumentum types Genus A Vavilo via Pisum L a thyrus A nu tropostylia Lens Vicia Cicer + Ononis Medicago Trigonella Melilotus Parochetus Trifolium -, Indicates hair-type absent; members of the genus. 11 B Types of hair (as in Fig. 5) C D E F + + + + + + - + + + + (+) + + + + +, hair-type present in all species; (+), hair-type present in only some 146 F. K. KUPICHA hairs in the Ononideae and the limited distribution of the multicellular hairs are further complicating factors. Inflorescence The inflorescence in the Vicieae, Trifolieae and Ononideae is basically racemose. In each tribe there is a complete range from few- t o many-flowered peduncles, and this variation does not provide any guide to Cicer’s affinities, despite the statement of De Candolle (1825a) that Cicer and Ononis have similar inflorescences. This is true, but the same type occurs in the rest of the Vicieae and in some members of the Trifolieae. The flower Corolla The flowers of Cicer have petals of simpler shape than those of the rest of the Vicieae, apart from the smallest-flowered autogamous species of Lathyrus and Vicia (e.g. L. gloeospermus, V. hirsuta (L.) S . F . Gray). In Cicer the wing petals are completely free from the keel (Popov, 1928-29: 22; van der Maesen, 1972), whereas in the other genera these parts are interlocked, as described by Kupicha (1973, 1976). The Ononideae and Trifolieae resemble Cicer in this respect; many members of these tribes, however, have flowers of more specialized construction than Cicer, as exemplified by the ‘medicaginoid’ corolla found in Medicago and parts of Trigonella and Ononis and by the situation in Trifolium where the petals are often adnate to the staminal tube (Townsend, 1974). Androecium Although the androecium is a relatively inconspicuous part of the plant, its variation was noted and emphasized by early taxonomists such as Bentham (1865) in their comments on the delimitation of leguminous tribes. The aspects which vary are: the staminal tube (monadelphous, completely diadelphous or with vexillary stamen lightly adnate to the filament tube); the filaments (tapering or dilated at the apex); anther size (monomorphir or dimorphic) and anther attachment (basifixed or dorsifixed). The distribution of these different character-states has been described by many authors, especially Bentham (1865), Popov (1928-29), Hutchinson (1964), van der Maesen (1972), Townsend (1974) and Gunn & Kluve (1976), although none has considered them all together in this context. The Vicieae are all diadelphous. In Cicer the vexillary stamen has a terete filament which is completely free from the other nine, but in the remaining genera this adaxial filament is flanged and lightly adnate to the others so that the androecium is functionally monadelphous, at least until the vexillary filament is pushed away by the expanding fruit (Kupicha, 1976). The Ononideae are remarkable in being truly monadelphous, with the partial exception of some species of Ononis (e.g. 0 . rotundifoliu L.) in which the vexillary filament is united with its neighbours only at the base. The Trifolieae are diadelphous (Townsend, 1974), although according t o Hutchinson (1964) DELIMITATION O F T H E VICIEAE-LECUMINOSAE 147 the vexillary filament may be united with others in the middle, presumably as in Vicia, etc. In Vicia, Lathyrus, Lens and Anatropostylia the filaments are all slender, and taper towards the apex. I found that Pisum and Vavilovia have alternate filaments dilated at the apex, although other workers have stated that they are all more or less dilated (Townsend, 1974) or linear except for the adaxial one, which is dilated (Gunn & Kluve, 1976). Cicer typically has all or most of the filaments swollen (van der Maesen, 1972; Gunn & Kluve, 1976). In the Ononideae all or alternate filaments are swollen (Popov, 1928-1929). Trifolium often has dilated filaments, but in the rest of the Trifolieae they are slender (Townsend). The Vicieae, including Cicer, and the Trifolieae have anthers of uniform size. The Ononideae, however, have alternately large and small anthers (Bentham, 1865; Townsend, 1974). The Vicieae, apart from Cicer, and the Trifolieae have versatile anthers, while the Ononideae characteristically have alternate anthers basifixed and versatile. Cicer seems to be variable in this respect, some species having uniformly versatile anthers and others having them alternately basi- and dorsifixed (van der Maesen, 1972). Although the significance of these androecial details is obscure, the fact that genera tend to be constant in their possession of each state suggests that they may have some systematic value. Pollen A paper describing the pollen of the Vicieae, Trifolieae and Ononideae has recently been published (Clarke & Kupicha, 1976). It was concluded that the pollen of Cicer is quite distinct from that of the rest of the Vicieae and the Trifolieae but is like that of the Ononideae. Unfortunately, we overlooked the earlier paper of Gapotchka (1974) which covered very similar ground. This author argues that the distinctive pollen of Cicer is evidence for recognizing a separate tribe, the Cicereae. Gynoecium Except for members of Cicer, all species of the Vicieae have pubescent styles which are usually flattened either dorsally or laterally and are held approximately at right angles to the ovary (Kupicha, 1973, 1976). By contrast, in Cicer the style is terete and glabrous and curves gently away from the end of the ovary (van der Maesen, 1972). In all of these characteristics the gynoecium of Cicer resembles those of the Ononideae and Trifolieae. The fruit Cicer has inflated fruits very similar to those of Ononis. Vicia and the rest of the Vicieae have fruits of rather uniform shape which are typically laterally compressed; they may be subtorulose (as in V. ervilia) but are never as inflated as in Cicer. The Trifolieae are remarkable in the diversity of their legumes, whose variation is useful for generic identification, but none of the fruits is like those of Cicer and Ononis. Illustrations of these fruits can be seen in van der Maesen (1972), Townsend (1974) and Kupicha (1976). 148 F. K. KUPICHA The seed The leguminous seed has been described in detail by Corner (195 l ) , and the terms used here for the details of seed morphology and anatomy are taken from his paper. Seed shape ?‘he seeds of the Vicieae, with the exception of Cicer, are compressed, spherical or oblong (though often slightly angled through contact with neighbouring seeds in the ovary), with only a slight ridge marking the position of the radicle. The latter is fairly long and curved and lies closely against the cotyledons (Fig. 6A). Seeds of Cicer, on the other hand, are grooved between the two cotyledons and the short conical radicle causes a beak above the micropyle (Fig. 6B). Corner described the radicle of Cicer as “short and straight”, and he likened it t o the median radicle of the mimosoid-caesalpinioid seed in contrast t o the “long curved radicle typical of the bean seed”. I consider, however, that the Cicer radicle is not truly of the mimosoid type, but E x t e r n a l appearance o f seed V a s c u l a r supply o f seed Seed w i t h testa removed Figure 6. Secds of the Vicieae, Ononideae and Trifolicae. A, Vicia sativu; B, Cicer ariefinurn;C , Ononis repens; D, Medicago rugosa. c, Cotyledon; ch, chalaza; e, endosperm; h, hilum; 1, lens; m, micropyle; pvb, post-chalaza1 vascular bundle; rvb, recurrent vascular bundle. Not to scale. Note: the lens is not invariably above the chalaza; in members of Vicia sect. Hypechusa the chalaza is in the normal position near the hilum, b u t the lens is situated on the other side of the seed (Kupicha, 1976: fig. 6b). DELIMITATION O F T H E VICIEAE-LEGUMINOSAE 149 is similar to that of the rest of the Vicieae, because its axis is turned at right angles to that of the cotyledons rather than being in line with them. The seeds of the Ononideae and 'l'rifolieae are strongly beaked and have long, curved radicles which are not appressed to the cotyledons (Fig. 6C, D). Hilum length The genera of the Vicieae, apart from Cicer, have more or less linear hila which range from & to 3 of the seed circumference; I found that, in general, perennial species tend to have longer hila than annuals (Kupicha, 1976; this is also shown by Gunn, 1970). By contrast, Cicer and genera of the Ononideae and Trifolieae have seeds with uniformly small elliptic or suborbicular hila (Fig. 6B, C, D), and this appears to be the common condition in the Papilionoideae. Endosperm Although members of the Vicieae have endosperm in the developing seed, as shown by Maheshwari (1950: 257), this tissue is broken down as the seeds reach maturity and the cavity within the testa becomes entirely filled by the cotyledons and radicle (Fig. 6A, B). The absence of endosperm in seeds of the Vicieae is related to their hypogeal mode of germination in which the cotyledons remain in the testa and are modified to act as food stores. There is some disagreement in the literature about the situation in the Ononideae and Trifolieae. Lubbock (1892: 411) and Erith (1924: 4) stated, respectively, that seeds of Ononis ultissirna Colm. and Trifolium repens are exalbuminous. I have observed, however, that seeds of the Ononideae and Trifolieae contain a tough hyaline endospermous sheath round the embryo, which readily imbibes water, becoming swollen and mucilaginous (Fig. 6C, D). The survey of Reid & Meier (1970) confirms that this tissue is present in species of Medicugo, Melilotus, Trifoliurn and Trigonella. Reid (1971) found that the endosperm of Trigonella foenum-graecurn is composed of large, thin-walled cells completely filled with the polysaccharide galactomannan, which is degraded during germination. I t is this storage carbohydrate which gives the seeds of the Trifolieae (and presumably the Ononideae) their characteristic mucilaginous property. Vascular supply According to my investigations in some 30 species of the Vicieae (Table 5 ) , in members of this tribe there is a long post-chalaza1 vascular bundle in the antiraphe. In Cicer the trace dichotomizes at a point about opposite the chalaza (i.e. on the other side of the seed), and its branches run laterally in the testa, often dividing again (Fig. 6B). This pattern of seed vasculature was observed by Kuhn (1928: 347). In the other genera of the tribe there is always a simple, unbranched vascular bundle forming a hoop round the periphery of the seed (Fig. 6A). I d o not agree with Kuhn, who stated that in Vicia faba this trace divides weakly at its extremity. In the Ononideae and Trifolieae (judging from the small sample in my survey) the seeds have a very short vascular bundle which ends at the chalaza (Fig. 6C, D). F. K. KUPICHA 150 Table 5 . Species in which the seed vasculature has been observed Cicer arietinum L. F C. pinnatifidum Jaub. & Spach F Lathyrus annuus L. S L. aphaca L. S L. blepharicarpus Boiss. S I,. clymenum L. S L. gorgoni Parl. S L . hirsutus L. S L . niger (L.) Bernh. S L . nissolia L. S L. rotundifolius Willd. S I,. sariwus L. S L. sphaericus Retz. S L . tingitanus L. S Lens culinaris Medicus S Vicia anatolica Turrill V. articulata Hornem. V. benghalensis L. S S S A, Post-chalaza1 vascular bundle absent; S, simple. Vicia bithynica (L.) L. S V. cuspidata Boiss. S V.graminea Smith S V . grandiflora Scop. S V. hyrcanica Pischer & C . A. Meyer V. meianops Sibth. & Sm. S V. michauxii Sprengel S V. pannonica Crantz S V . peregrina L. S V. sativa L. S V. villosa Roth S S Ononis repens L. A 0. spinosa L. A Medicago rugosa Desr. A Melilotus alba Medicus A Trifolium constan tinopolitanum Ser. A F , S , post-chalaza1 bundle present: F, forked; The karo type: chromosome number The distribution of chromosome numbers throughout the Leguminosae was discussed by Senn (1 9 3 8), and subsequently Turner & Fearing ( 1 95 9) provided a modified phyletic interpretation of chromosome number relationships within the Caesalpinioideae and Papilionoideae. These authors suggested that the Papilionoideae evolved from caesalpinioid ancestors along two main lines: one, characterized by the base number 1 1 , led to the Phaseoleae and Dalbergieae and parts of the Galegeae and Hedysareae; the other, with primitive base number 8, gave rise t o the ‘Galegeae complex’: Genisteae, Loteae, Trifolieae and Vicieae, together with the rest of the Hedysareae and Galegeae. The Podalyrieae and Sophoreae, with x = 9, formed a third branch connected to the Galegeae complex. Many authors have noted the usefulness of chromosome numbers as a simple taxonomic character. Darlington (1956: 102) said that where the numbers are constant they can reveal ancestry or phyletic lines: “they label whole tribes, families or orders”. The situation in the region of the Vicieae is not as straightforward as this but even so the pattern of variation of chromosome numbers does provide useful taxonomic evidence. Table 6 shows the haploid chromosome numbers ( n ) found in genera of the Vicieae, Ononideae and Trifolieae. It was compiled from data given by Darlington & Wylie (1955), Cave (1958-60 and 1961-651, Ornduff (1967 and 1968), Moore (1973 and 1974), van der Maesen (1972) and Contandriopoulos e l al. ( 1 972). Several interesting facts emerge from the table. Vicia, Lathyrus, Lens and Pisum are characterized by n = 7 ; some lower numbers are found but n = 8 is completely absent. The genera of the Trifolieae are characterized by y1 = 8, with only a small spread of lower and higher numbers. Polyploidy is 28 2 8 3 1 1 6 11 53 2 51 1 41 5 8 14 23 113 20 8 7 1 4 9 10 1 1 I1 4 4 1 1 12 13 5 1 1 5 1 3 I4 16 1 18 1 9 1 2 14 15 1 17 1 18 19 1 20 21 2 22 23 5 2 24 25 26 27 1 1 28 29 1 30 31 2 4 32 N o t e . The scores indicate the approximate number of species in each genus having a particular value for n , as reported in the literature. When a species was recorded as having two or more values for n , each of these values was scored as if equivalent to the single value of a species yielding only one haploid chromosome number. The report of 2n = 16 for Vicia faba in Moore (1973)is erroneous. Pisum La f hyrus Lens Vicia Cicer Ononis Medicago Trigonella Melilo tus Trifolium n=5 Table 6. Cliromosome numbers in the Vicieae, Ononideae and Trifolieae 152 F. K. KUPICHA relatively infrequent in these two groups. The indices for Cicer are striking, as they show this genus with a strong representation of both YZ = 7 and IZ = 8, typical of neither the rest of the Vicieae nor the Trifolieae, but intermediate between them! Here again there is little polyploidy. The Ononideae prove t o be quite distinct; they are entirely polyploid, predominantly with ii = 15 and 16. According to the hypothesis of Turner & Fearing, the Vicieae, belonging to the ‘Calegeae complex’, have x = 8 as the ancestral base number. This number, however, is apparently unrepresented in Vicia, Lathyrus, Lens and Pisum,and so i suggest that these genera probably had a more recent common ancestor with n = 7. Ciccr, on the other hand, has both ii = 7 and n = 8. If Ciccr is closely related to the rest of the Vicieae, it must either have diverged from them before the appearance of the y1 = 7 ancestor, or, if it was descended from this ancestor, the presence of 11 = 8 must be due to recent ascending aneuploidy and be merely a parallelism imitating the situation in the Trifolieae. The former alternative is more parsimonious and seems to me the more likely. C/icmosy st ema tics Anthocyanins The important plant pigments, anthocyanins, which are largely responsible for flower colour, are glycosidic derivatives of anthocyanidins (cf. Harborne, 1967). Variation in the composition of anthocyanin sugar residues within the Papilionoideae may be of systematic value. A recent review showed that whereas many members of the subfamily have glucose residues, the Vicieae (including Cicev), which have been quite thoroughly surveyed, are characterized by anthocyanins with rhamnose residues (Harborne, 19712: 34). At the time of this review the Trifolieae and Ononideae had been very sparsely screened, and found to possess anthocyanidin glucosides. The published data thus imply that the Vicieae can be separated from their neighbours by anthocyanin chemistry. However, J . L. Ingham (Botany Ilept., University of Reading) has informed me that malvidin-3-rhaninoside has been provisionally identified from one member of the Trifolieae, Parochetus cornmuPiis Buch.-Ham. ex D. Don. This suggests that the glycosidic variation-pattern, when fully exposed, might link the Vicieae and Trifolieae rather than separating them. A more intensive investigation in this area would be a worthwhile operation. Is0 fluvonoids The isoflavonoids are a group of compounds which have a very limited distribution among flowering plants in general but which occur in many members of the Papilionoideae (Harborne, 197 l a : 47); here, morcover, their diversity promises t o be of considerable taxonomic significance. in the Papilionoideae isoflavonoids exist as constitutive substances in healthy plant tissues but they also function as phytoalexins, being produced in response to stimulus by toxic chemicals or infecting micro-organisms (Ingham, 1972). Cicev appears to be distinct from the rest of the Vicieae in its isoflavonoids, but similar to the Trifolieae and Ononideae. The constitutive isoflavones biochanin A, formononetin, pratensein and daidzein have been found in C urietirzum; the first two, in particular, occur in large quantities in the stems, roots and leaves (Ingham, in press). None of these has been detected in other DELIMITATION O F THE VICIEAE-LEGUMINOSAE 153 genera of the Vicieae despite several detailed surveys (Ingham, personal communication), but all have been found in Trijulizim (Francis et al., 1967) and formononetin was reported from Ononis (Harborne, 1971a: 49 & 60). C. arietinum produces the isoflavonoid phytoalexins medicarpin and maackiain (Ingham, in press). Medicarpin has been found in small quantities in Vicia faba and its close relatives I? galilaea Plitm. & Zohary and V. narbonensis L. (Hargreaves et at., 1976) but in no other member of the Vicieae. I t is, however, a characteristic phytoalexin of Trigonella, Medicago and Melilotzts (Ingham & Harborne, 1976) and Trifolium (Higgins & Smith, 1972). Maackiain is produced by many members of the Trifolieae and was also reported from Pisum (Stoessl, 1972; here it is called inermin). On the other hand, two phytoalexins which have been isolated from members of the Vicieae, the isoflavonoid pisatin from Pisum and the polyacetylenic compound wyerone from Vicia (Harborne, 1971b: 276) are unknown in Cicer and the Trifolieae and Ononideae. Studies in this area of plant biochemistry are developing rapidly and yielding welcome taxonomic information. This brief account indicates the current feeling about Cicer's position with regard to its isoflavonoid chemistry; a proper appreciation of the taxonomic importance of presence or absence of the various compounds would, however, require a full understanding of their biosynthetic relationships. Non-pro t e in amino acids : cuna van in e The non-protein amino acid canavanine is apparently found only in the seeds of some members of the Papilionoideae, and its pattern of distribution within the subfamily is of taxonomic interest (Bell, 1971). Table 7 summarizes the data available in the area related to the present discussion. This shows that the Trifolieae and Ononideae are characterized by the constant presence of canavanine in their seeds. On the other hand, the Vicieae, apart from some members of Vicia subgenus Vicilla, lack this compound (Kupicha, 1976). The report of the situation in Cicer, which was based on only one species (C. arietinqm), should be confirmed before a firm conclusion can be drawn, but the present results suggest that the pattern of distribution of canavanine links Table 7. The distribution of canavanine in the Vicieae, Ononideae and Trifolieae Genus Pisum 1.u thyr1rs I,ens viciu Cicer Oilonis Medicago Trigonellu Melilotus Trifolium No. species examined 1 49 1 I17 , 30 1 4 57 3 3 17 Canavanine prescnt (+), absent ( -) ~ ~ - +: -I ~ + + + + + Authority Przybylska & Hurich (1961) Bell (1962) Birdsong, Alston & Turner (1960) Bell & Tirimanna (1965); results generally confirmed by Tschiersch & Hanelt (1967) Birdsong, Alston & Turner (1960) Turner & Harbornc (1967) Turner & Harborne (1967), Bell (1971) Turner & Harborne (1967) Turner & Harborne (1967) Turner & Harhorne (1967), Bell (1971) 154 F. K. KUPICHA Cicer with the rest of the Vicieae rather than with the other tribes. Outside these groups, canavanine occurs in most species of the Galegeae, and sporadically in the Podalyrieae, Genisteae, Hedysareae, Dalbergieae and Phaseoleae. Seed proteins The seed proteins of the Leguminosae have been investigated by many workers, using a variety of techniques. In a number of cases the findings are relevant to the present discussion, but while they show that variation in protein characters is systematic, the inadequate reporting of some results and the conflicting nature of others prevent a firm conclusion being drawn from the evidence. It is hoped that this brief survey will encourage further studies, focussed on the genus Cicer, which should be compared not only with the rest of the Vicieae but equally with members of the Ononideae and Trifolieae. The albumin fraction of legume seed protein contains enzymes, including formic and glutamic dehydrogenases (FDH and GDH). Thurman et al. (1967’) used gel electrophoresis to compare FDH and GDH in seeds of the Vicieae, Trifolieae and other tribes. The presence of the proteins in the gel after electrophoresis was detected by enzymatic production of formazan from tetrazolium compounds. It was found that all members of the Vicieae which were tested gave one FDH band, while there were five GDH bands in all genera of the tribe except Cicer, which had two. The Rp value of FDH in Cicer was higher than those of the rest and approached the Rp values of the Trifolieae FDH bands. The number of FDH and GDH bands in the Trifolieae was not mentioned. The globulin fraction (i.e. the storage protein) of legume seeds has been the object of particular interest. Boulter et al. (1967) made a wide survey of globulins of the Papilionoideae using gel electrophoresis. The 12 species of the Vicieae in their investigation (including Cicer arietinum) gave a very uniform band-pattern, and this differed from the equally uniform pattern of the Trifolieae. Jackson et al. (1967) analysed the globulin fraction of seeds of the Vicieae by ‘fingerprinting’ the peptides resulting from hydrolysis with trypsin. It was found, by comparing chromatograms, that the ‘fingerprints’ of the genera were quite similar. Those of Lathyrus and Lens differed from Vicia each by three spots, while those of Pisum and Cicer differed from Vicia by seven and thirteen spots, respectively. It was not stated by how many spots the genera Lens, Lathyrus, Pisum and Cicer differed from each other, or the distribution of each spot between the genera. I t would be interesting t o extend the range of the survey and compare globulins from the Trifolieae and Ononideae with those of the Vicieae. In many tribes of the Papilionoideae, including the Vicieae, Ononideae and Trifolieae, the globuIin fraction has been found to include two major components with sedimentation coefficients of about 7s and 12s: these are known, respectively, as vicilin and legumin (Danielsson, 1949; Boulter & Derbyshire, 1971). Jackson et al. (1968) separated the vicilin and legumin of Pisurn sativum, Vicia faba and Cicer arietinum, and compared them in terms of their amino acid composition, ‘fingerprint’ maps and gel electrophoresis band-patterns. The results showed that there is considerable similarity between these three species with respect to both vicilin and legumin. DEI.IMITATION O F THE VICIEAE-LEGUMINOSAE 155 Several workers have used the immunological properties of seed storage proteins to study the relationships of genera within the Vicieae and between this tribe and its neighbours. Kloz & Turkova (1963) separated the vicilin and legumin from seeds of Pisurn sativum and, having prepared antisera to each, used immunodiffusion and electrophoresis to test them against antigens from the other genera of the Vicieae and from other tribes. They concluded from these heterologous reactions that both vicilin and legumin are absent from the Genisteae and Phaseoleae; Trijbliurn and Medicago have no legumin but a protein of vicilin type; and all members of the Vicieae, including Cicer, have both legumin and vicilin, although the vicilin of Cicer is not the same as that in the other genera. Simola (1969) prepared antisera to seed proteins of several species of Lathyrus and used them to make a serological survey of the Vicieae and other tribes. She found that antigens of Pisum, Lens and Viciu produced heterologous reactions almost identical with the homologous reaction. Species of Abrus, Phaseolus, Lotus and Oxytropis gave reactions indicating distant relationship, and there was no reaction when antigen from Trifoliurn was used. Both Ononis and Cicer gave strong reactions; these indicated considerable homology with Lathyrus but a relationship more distant than that of the rest of the Vicieae. The recent paper of Dudman & Millerd (1975) reports an investigation carried out with particular care, based on antisera to the vicilin and legumin from seeds of Vicia faba. The results of immunodiffusion experiments showed that Pisum sativum and species of Lathyrus gave reactions of identity with both legumin and vicilin antisera; Lens culinuris gave a reaction of identity to legumin but of only partial identity to vicilin. Cicer arietinzcm, and members of the Trifolieae (ten species) and Ononideae (five species) all gave partial-identity reactions to legumin but had non-identity with vicilin. As Dudman & Millerd emphasized, differences in serological techniques can have a marked effect on the results and their interpretation, and this may account for the discrepancies between their findings and those of Kloz & Turkova (1963) and Simola (1969). Few generalizations can be made from the data summarized above. I t appears that as far as proteins are concerned one must be wary of accepting taxonomic evidence derived from any one method of analysis, because each technique seems to produce slightIy different results. In the present case, a11 the experiments lead to the conclusion that Vicia, Lathyrus, Pisum and Lens are very closely related. There is also considerable agreement that the Trifolieae (and Ononideae, although Ononis has rarely been studied) are closer to the Vicieae than any other tribe. The position of Cicer is equivocal: some studies decisively link this genus with the rest of the Vicieae, while others show it to be fairly closely related but set apart from the “core” of the tribe. In order to achieve a balanced view of the situation we need more experimental data about the Trifolieae. Ononideae and Cicer itself. DISCUSSION The delimitation of the Vicieae Table 8 is a list of the attributes of Cicer which link this genus with the rest of the Vicieae or with the Ononideae or Trifolieae; conversely, it also shows F. K. KUPICHA 156 Table 8. A comparison between Cicer and the rest of the Vicieae, the Ononideae and Trifolieae Characters of Cicer Vi. Ci. Ci. On. Tr. On. On. Tr. Ci. On. Tr. Vi. Vi. Vi. On. Tr. Vi, On. Tr. On. Tr. Tr. Vi. on. Tr. Vi. On. On. Tr. On. Tr. On. On. Tr. On. Tr. Vi. Vi. Ci. On. Tr. Vi. On. Tr. Tr. Vi. * * * 0 * 0 0 0 1. 2. 3. 4. 5. 6. 7. 8. 9. 0 10. 0 11. 12. 13. 14. 15. 16. 0 0 D * * * * U 0 0 0 * * * * * 0 0 17. 18. 19. 20. 21. 22. 23. 24. 25. 26. 27. 28. 29. 30. 31. germination hypogcal radicle growing towards or away from hilum first scale-leaf of seedling having variable position transition region in hypocotyl plants often spiny no cortical vascular bundles in stem first juvenile leaf with 2 o r more pairs of lateral leaflets juvenile leaves with terminal leaflet mature leaves sometimes tendrillous mature leavcs usually multijugate stipules free from petiole leaflets serrate, with craspedodromous venation uniccllular hairs smooth-walled long-stalked glands present wing petals free from keel androecium diadelphous anthers alternately basifixed and versatile anthers of uniform size pollen grains subspherical, with relatively thin walls and large pori apertures of pollen grains unthickened style tcrete, glabrous legume inflated seeds beaked above hilum hilum small, subcircular endosperm absent in mature seed post-chalaza1 vascular bundle present in sccd post-chalaza1 vascular bundle dichotomizing isoflavonoid chemistry shows affinity with Ononideae and Trifolieae canavaninc absent from seeds base chromosome number x = 8 present chromosome complement usually diploid, not polyploid Vi. character-state typical of the Vicieae, On. of the Ononideae, Tr. of the Trifolieae; Ci. characterstate peculiar to Cicer, or, if corresponding character-state in Ononideae and Trifolieae is unknown, separating Cicer from the Vicieae. m, Charactcr-state specialized, within the context of these four tribes; 0, character-state primitive; *, character-state cannot be judged specialized or primitive. where there is discontinuity of character-variation between Cicer and its neighbours. The precise number of character-states which Cicer shares with each of the other groups is not significant, because several of the characters need confirmation, some do not hold true for every species, and some could be subdivided into more characters. However, the table is a convenient summary of the phenetic situation, as far as it is known to me. From the table, the following conclusions can be drawn. Firstly, the characters which link Cicer with the rest of the Vicieae are considerably outnumbered by those which separate these groups and instead show affinity between Cicer and the Trifolieae and/or Ononideae. Secondly, if all characters are given equal weighting, the present evidence indicates that the affinity between Cicer and the Ononideae is of the same order as that between Cicer and the Trifolieae. The claim of De Candolle (1825a), Popov (1828-29) and many others that Cicer has a special relationship with Ononis therefore cannot be upheld. DELIMITATION 0 I; THE VICIEAE-LEGUMINOSAE 157 The first of these statements confirms that Cicer is anomalous within the Vicieae. If it is removed, the range of variation within the tribe is significantly reduced, and its delimitation from other taxa of the same rank becomes much clearer. The effects of this alteration obviously result from the achievement of a ‘natural’ group which is held together by the total of its attributes, in contrast with the traditionally-defined Vicieae which were recognized merely by their possession of tendrils and hypogeal germination. I propose, therefore, that Cicer should be excluded from the Vicieae, which should now comprise Vicia, Luthyrzis, Lcns, Anutropostylia, Pisirm and Vavilovia. Description of the tribe Vicieae sensu stricto Perennial and annual herbs with erect or more usually climbing or sprawling habit; plants rarely tuberous. Indumentum of simple, smooth-walled hairs and short-stalked glandular hairs. Stems with cortical vascular bundles in the internodes, often winged; primary shoot almost always of limited growth, plants proliferating from basal nodes. Leaves epulvinate, alternate, distichous, hypostomatic, amphistomatic or epistomatic (stomata anomocytic), paripinnate with the rachis ending in a tendril or mucro, or very rarely imparipinnate; leaflets many-paired to unijugate; very rarely (in Lathyrus) leaves phyllodic or reduced to a tendril and stipules; stipules semisagittate or hastate or variously divided, sometimes with a nectariferous spot, never adnate to the petiole. Vernation of leaflets conduplicate or supervolute. Venation brochidodromous, veins pinnate or parallel. Inflorescence rarely a panicle, usually a secund, axillary raceme or sometimes 1-flowered; bracteoles almost always absent. Vexillum oblong, platonychioid or stenonychioid, occasionally pouched. Alae united with carina. Stamina1 tube diadelphous but with vexillary stamen lightly adhering to its neighbours, truncate or oblique at apex; anthers introrse, versatile, of equal size, rarely alternate filaments dilated at the apex. Style borne at right angles to ovary, usually compressed dorsally or laterally, always pubescent (distribution of pubescence various), sometimes spathulate, sometimes contorted; stigma terminal, very rarely double. Legume more or less linear, laterally compressed, 2-many-seeded, usually dehiscent, occasionally winged, sometimes with woolly or (very rarely) membranous partitions between the seeds. Seeds compressed-spherical, with long to short hilum; testa smooth or variously rough-textured; lens near hilum or rarely opposite; vascular bundle continuing past chalaza but not branched; endosperm absent. Germination hypogeal; radicle and hypocotyl triarch (very rarely tetrarch); transition region between root and stem in epicotyl; first scale-leaf borne on side of plumule away from cotyledons. The taxonomic position of Cicer Having taken Cicer out of the Vicieae, one must find a better place for this genus within the Papilionoideae. Cicer s systematic position should be influenced not only by the number of features which it has in common with various alternative groups, but also by the present delimitation and variability of those taxa and the effect which its inclusion within one of them would have on the hierarchy. 158 F. K. KUPICHA The Trifolieae form a very well-defined natural tribe comparable with the Vicieae sensu stricto, and the monogeneric Ononideae are similarly limited in their internal variability. In these circumstances, neither tribe could absorb Cicer without profoundly altering its definition. Moreover, the differential characters which distinguish between the Trifolieae and Ononideae (Table 2) are of less weight than those separating each of these tribes from Cicer, so that if the creation of the Ononideae is valid Cicer must also be excluded from both of them. Alternatively, if Ononis is regarded as a member of the Trifolieae (as has been done by most recent authorities), the latter becomes more polymorphic and could be further enlarged to encompass Cicer. Although it would be preferable to have Cicer in the Trifolieae sensu lato than in the Vicieae, it seems unwise to blur the definition of the Trifolieae to this extent. The most satisfactory course is to place Cicer in a tribe of its own. A suitable name in this category already exists because Alefeld (1859), with his peculiar concept of taxonomic ranking, made the Vicieae sensu lato a subfamily containing the tribes Ciceridae, Viciidae and Orobidae. I propose that the Ciceridae should be brought out of obscurity to form a new monogeneric tribe, the Cicereae Alef. Phylogenetic relationships of Cicer This discussion has so far been based on the phenetic method of classification, which gives all characters equal weight. I t is interesting to make an alternative interpretation of the same data, using the phylogenetic systematic method (Hennig, 1966), and to find that while the results are similar in practical terms, the evolutionary dimension becomes much clearer. The surest evidence that a group is monophyletic is the presence in all members of that group, and complete absence outside the group, of one or more uniquely derived characters. Each of the character-states of Cicer listed in Table 8 has been judged specialized, primitive or indeterminate, in the context of this part of the Papilionoideae (see symbols in second column). Two features of Cicer, both of which are shared with the Trifolieae and Ononideae, can confidently be assumed to be uniquely derived: the serrated leaflets with craspedodromous venation (character 12 in Table 8) and the long-stalked glandular hairs (ch. 14). The corresponding character-states in the Vicieae are relatively primitive. This evidence immediately indicates that Cicer is cladistically more closely related to the Trifolieae and Ononideae than t o the Vicieae (Fig. 7). It is theoretically impossible for Cicer to share different uniquely derived characters with the two mutually exclusive groups Trifolieae-Ononideae and Vicieae. This consideration throws doubt on the status of the two main characters, hypogeal germination (ch. 1) and the presence of tendrils (ch. 9), which link Cicer with the Vicieae. Hypogeal germination has already been shown to be widely distributed in the Papilionoideae and possibly reversible with epigeal germination, and is therefore of doubtful significance. Tendrils, on the other hand, are very rare and certainly specialized. Nevertheless, I am forced to conclude that they must have evolved separately in Cicer and the Vicieae sensu stricto. This conclusion is supported by the presence in both Cicer and Vicia of species which have the primitive, imparipinnate leaf-form DELIMITATION OF THE VICIEAE-LEGUMLNOSAE 159 W a Figure 7. Diagram of cladistic relationships. ch, Uniquely derived character, numbered as in Table 8. throughout their life history, as well as by the different patterns of leaf ontogeny in the two groups. The Ononideae and Trifolieae together possess one unique specialized character (ch. 11, the stipules adnate to the petiole) which suggests that they had a common ancestor subsequent to the isolation of the line leading to Cicer. This is shown in Fig. 7 at A. (Character 1 1 is expressed here in its derived state; Table 8 gives the corresponding primitive state for Cicer.) I do not have enough evidence to decide on cladistic relationships within the Trifolieae-Ononideae; this subject needs a full independent investigation. I t is apparent, however, that the Genisteae, which have often been proposed as close relatives of Ononis, are cladistically further from Ononis than both the Trifolieae and Cicereae, because they do not show the derived characters which are markers for these taxa. I t must be admitted that up till now I have assumed that the Cicereae, Ononideae, Trifolieae and Vicieae comprise a monophyletic group, but this may be wrong; I know of no uniquely derived character which delimits this group from the rest of the Papilionoideae. The only evidence which links the Vicieae with these other tribes is phytochemical, especially serological. A wider, very detailed assessment of character-variation within the Papilionoideae is required to determine the closest relatives of the Vicieae. Figure 7 expresses the present uncertain situation by the alternative branching at B and C. I suggest that in beginning a wider survey, one should look first at tribes which have the base chromosome number x = 8, viz. Galegeae p.p., Genisteae, Hedysareae p.p. and Loteae. ACKNOW I.8DGBML.NTS This paper presents part of the work carried out for the degree of Ph.D. in the University of Edinburgh. I thank the Science Research Council and the Senatus Academicus of Edinburgh University for their generous financial F. K. KUPICHA 160 support, and Profcssor R. Brown and Mr D. M. Henderson, the Kegius Keeper of the Edinburgh Royal Botanic Garden, for their permission to work in the University Botany Department and at the Botanic Garden, respectively. I also thank Dr Ir L. J , G. van der Maesen for information about Cicer seedlings, Mr 4. L. Ingham for helpful discussions about the chemosystematic part of the paper, and Dr C. J . Humphries for encouraging me to branch o u t into phylogenetics. I am most grateful to my two supervisors, Dr P. H. Davis and Ilr P. M. Smith, for their guidance, kindness and stimulating interest. ADANSON, M., 1 7 6 3 . Familles d e s plantes, 2. LD, I:., 1 8 5 9 . Ucbcr die Vicieen. Oesterreichische Botanische Zeitschrift, 9: 352-366. ASCIIEKSON, P . & GKAEBNER, P . , 1 9 0 9 . Synopsis der mitteleuropaischen Flora, 6 ( 2 ) . BASSLElR, M . , 1 9 7 3 . IIcvision tier eurasiatischcn Arten von Lathyrus I.. Sect. Ovobus (I>.) Gren. & Godr. Feddes Repertorium, 84: 329-447. B A U D I T , J . C., 1974. Signification taxonomique des charactkres b l a s t o g h i q u e s dans la tribu des Papilionaceae~l’liaseoleac. Bulletin d u Jardin Botaniyue Nutional d e BeZgique, 44: 259-293. BELL, Li. A., 1962. Associations of ninhyclrin-reacting c o m p o u n d s in t h e seeds of 49 species o f Luthyrus. Biochemical Journal, 83: 225-229. BELL, E. A., 1 9 7 1 . Comparative biochemistry of non-protein amino acids. I n J . B. Harborne, I>. Boulter & B. L. Turner (Kds), Chemotaxonomy of the Leguminosae: 179-206. London a n d New York: Acadcmic Press. BELL, E. A. & TIKIMANNA, A . S . L., 1965. Associations of amino acids and related c o m p o u n d s in t h e seeds o f 4 7 species of Viciu: their taxonomic and nutritional significance. Biochemical Journal, 97: 104-111. RENTIIAM, G.,1 8 6 5 . Lcguminosae. In Bentham & J . D. Hooker, Generu pluntarum, 1 ( 2 ) . BIRDSONG, B. A , , ALSTON, R. & T U R N E R , B. L., 1960. Distribution of canavanine in t h c faillily Lcguminosae as related to phyletic groupin@. Canadian Journal of Botany, 38: 499-505. BOU12Tl:K, D. & DEKBYSHIRE, E., 1 9 7 1 . Taxonomic aspects o f t h e structure of legume proteins. In J B. Ilarbornc, D. Boulter & 8. L. T u r n e r (Eds), Chemotuxonomy of the Leguminosae: 285.308. London & New York: Acadilemic Press. BOUI.TEK, D., TIIUKMAN, D. A. 6: I>I<RBYSHIKl, E., 1967. A disc electrophoretic study of globulin proteins of legume secds with reference t o their systematics. N e w Phytologist, 66: 27-36. BRONN, I I . (;,, 1822. De forrnis planturum leguminosurum primitivis e t derivatis. CAVE:, M. S . (Ed.), 1958-1960. Index to plant chromosome numbers, 1. University of North Carolina Press. CAVE, M. S. (Kd.), 1961-1965. Index to plant chromosome numbers. 2. University of North Carolina Press. CLARKE, C,. C. S. & KUI’ICIIA, t:. K., 1 9 7 6 . The relationships o f t h e genus Cicer I.. ( L c p m i n o s a c ) : t h e evidence froni pollen morphology. Botunicul .Journal of the Linnean Society, 72: 35-44. COMPTON, K. H., 1912-191 3. A n investigation of the seedling structure in t h e L c p m i n o s a e . Botanical Journal of the Linneun Society, 41: 1-122. CONTANL)llIOPOULOS, J . , PAMUKCUO6LU, A. & QUEXEL, P., 1 9 7 2 . A propos des Cicer vivaces d u t o u r M c d i t c r r d n k n oriental. Eiologiu Gallo-Hellenicu, 4 : 3-18. :I<, E. J. l l . , 1951. The leguminous seed. Phytomorphology. 1 : 117-150. DY, V., 1968. Keimlingsbestirnmurzysbucli der Dikotyledonen. Budapest: AkadPmiai Kiaclb. LSSON, C. E., 1949. Seed glotiulins of t h e Gramineae a n d Leguminosae. Biochemical Journal, 44: 387-400. DAKLING’L‘ON, C. D., 1 9 5 6 . Chromosome botany. London: Allen & Unwin. DARLIN(;TON, C. D. & WYLIli, A. P., 1 9 5 5 . Chromosome utlas offlowering plants, 2nd ed. Aberdeen: . Mkmoires sur la .famille des Lkgumineuses, 8: 347-51. . Prodromus systemutis nuturulis regni vegetabilis, 2. I)OIIMF.K, K. J . , 1946. Vegetative morphology as a guide t o t h e classification of t h e I’apilionatae. New Phyiologist, 45: 145-16 1. l)UDMAN, W. I’. & MIL.LL.:RI), A , 1975. Immunochcmical hehaviour o f legumin and vicilin f r o m Vicia fubu: a survey o f related proteins in t h e Leguniinosac subfarnily I:aboideae. Biochemical Systematics and f:‘coIogy. 3 : 25-33. RIIITH, A. (;., 1924. White clover, u monograph. London: Duckworth. DELIMITATION O F THE VICIEAE-LEGUMINOSAE 161 FRANCIS, C. M., MILLINGTON, A. J . & BAILEY, E . T., 1967. T h c distribution of ocstrogenic isoflavones in t h e genus Trifolium. A ustralian Journal of Agriculturul Research, 18: 47-54. GAMS, H., 1924. Leguminosae. In G . Hegi, Illustrierte Floru von Mittel-Europa, 4 ( 3 ) . GAPOTCHKA, G . P., 1974. O n t h e palynomorphology of the species of the tribe Vicicac (Fabaceae). Vestnik Moskovskogo Universiteta, Ser. 6 Biologiya, I Y 74, 3: 93-98. GOURLEY, J . I I . , 1931. Anatomy of the transition region of Pisum sativum. Botanical Gazette, 9 2 : 367-383. GUNN, C . R., 1970. A kcy and diagrams for thc seeds of 100 species of Vicia. International Seed Testing Association Proceedings, 3 5 : 773-790. GUNN, C. R. & KLUVE, J., 1976. Androecium and pistil characters for tribe Vicieae (Fabaceae). Taxon, 25: 563-575. HARBORNE. J . B., 1967. Comparative biochemistry o f the flavonoids London and New York: Academic Press. HARBORNE, J. B., 1971a. Distribution of flavonoids in the L.eguminosae. In J . B. Harborne. D. Boulter & B. L. Turncr (Kds), Chemotuxonomy of the Leguminosae: 31-71. London & New York: Acadcmic Press. HAKBOKNE, J . B., 1971b. Tcrpenoid and other low molecular weight substances of systematic interest in the Leguminosae. In J . B. I-larbornc, D. Boulter & B. I.. Turner (Eds), Chemotaxonomy o f the Leguminosae: 257-283. London & New York: Academic Press. HARGREAVBS, J . A,, MANSFIELD, J. W. & COXON, D. T., 1976. Identification of mcdicarpin as a phytoalexin in the broad bcan plant (Vicia faba L.). Nature, 2 6 2 : 318-319. HENNIG, W., 1966. Phylogenetic systematics. (Translated by D. D. Davis & R. Zangerl.) University of Illinois Press. HERAIL, M. J., 1885. Rechcrches sur I’anatomic c o m p a r k dc la dge dcs Dicotyltdones. Annules des Sciences Naturelles, scptikme shrie, Botanique, 2: 203-3 14. HIGGINS, V. J. & SMITH, D. G., 1972. Separation and identification of two pterocarpanoid phytoalcxins produced by red clover leaves. Phytopathology, 6 2 : 235-238. HUTCHINSON, J . , 1964. The genera offlowering plants, I . Oxford: Clarendon Press. INGHAM, J . L., 1972. Phytoalexins and other natural products as factors in plant disease resistance. Botanical Review, 3 8 : 343-424. INGHAM, J . L., in press. Induced and constitutive isoflavonoids from stems of chickpeas (Cicer arietinum L.) inoculated with spores of Nelminthosporium curbonum Ullstrup. Phytopathologische Zeitschrift. INGHAM, J . L. & HARBORNE, J . B., 1976. Phytoalexin induction as a new dynamic approach to t h e study of systematic relationships among higher plants. Nature, 260: 241-243. JACKSON, P., BOULTER, D. & THURMAN, D. A., 1968. A comparison of some properties of vicilin and legumin isolated from seeds of Pisum sutivum, Vicia faba and Cicer arietinum. N e w Pkytologist, 6 8 : 25-33. JACKSON, P., MILTON, J. M. & BOULTER, D., 1967. Fingerprint patterns of t h e globulin fraction obtained from seeds of various species of the Fabaceae. N e w Phytologisf, 66: 47-56. KLOZ, J . & TURKOVA, V., 1963. Legumin, vicilin and proteins similar to them in the seeds of some species of the Viciaceae family ( a comparative serological study). Biologia Plantarum, 5 : 29-40. KUHN, G., 1928. Beitrage zur Kenntnis der intraseminalen Lcitbiindel bei den Angiospermen. Botanische Jahrbucher f u r Systernatik, Pflanzengeschichte und Pjlanzengeographie, 6 1 : 325-379. KUPICHA, F . K., 1973. Studies in the Vicieae I: The new genus Anatropostylia. Notes f r o m the Royul Botanic Garden, Edinburgh, 32: 247-250. KUPICHA, F. K., 1975. Some observations on the vascular anatomy of the tribe Vicieae (Leguminosae). Botanical Journal o f the Linnean Society, 70: 231-242. KUPICHA, F. K., 1976. The infrageneric structure of Vicia. N o t e s f r o m the R o y a l Botanic Garden, Edinburgh, 34: 287-326. LUBBOCK, J . , 1892. A contribution to our knowledge of seedlings, I . MAHESHWARI, P., 1950. A n introduction t o the embryology o f Angiosperms. London: McCraw-Hill. McCOMB, J. A., 1975. Is intergcneric hybridisation in the Leguminosae possible? Euphytica, 24: 497-502. MOORE, R. J . (Ed.), 1973. Index t o plant chromosome numbers 1967-1971.Regnum Vegetabile, YO. MOORE, R. J. (Ed.), 1974. Index to plant chromosome numbers for 1972. Regnum Vegetabile, 91. MULLER, C., 1937. La tige feuillee et les cotylkdons des Vicites germination hypogte. La Cellule, 4 6 : 195-354. ORNDUFF, R. (Ed.), 1967. Index to plant chromosome numbers for 1965. Regnum Vegetabile, 50. ORNDUFF, R. (Ed.), 1968. Index to plant chromosome numbers for 1966. Regnum Vegetabile, 55. POPOV, M. G., 1928-29. The genus Cicer and its species. T r u d y P O Prikludnoi Botanike, Cenetike i Selektsii, 2 1 : 1-239. PRZYBLSKA, J. & HURICH, J., 1961. Primary research after various forms of nitrogen in seeds of certain leguminous plants. Cenetica Polonica, 2: 65-73. REID, J. S . G., 1971. Reserve carbohydrate metabolism in germinating seeds of Trigonella foenum-gvaecum L. (Leguminosae). Planta, 100: 13 1-142. 12 162 F. K. KUPICHA REID, J . S. G , & MEIEK, H . , 1970. Chemotaxonomic aspects of the reserve galactomannans in leguminous seeds. Zeitschrift fur Pflanzenphysiologie, 6 2 : 89-92. REINKE, J , , 1897. Untersuchungen iiber die Assimilationsorgane der Lcguminosen. Jahrbuch f u r Wissenschaftliche Botanik, 30: 5 29-6 14. SCHNEPF, E., 1965. Licht- und electronenmikroskopische Beobachtungen an den Trichom-Hydathoden von Cicer arietinum. Zeitschrift fur Pflanzenphysiologie, 5 3 : 245-254. SENN, H. A., 1938. Chromosome number relationships in the Leguminosae. Bibliographia Genetica, 13: 175-345. SHAH, G. L.. & KOTHARI, M. J., 1973. On the structure of stomata and hairs and its bearing on the systematics in the tribe Vicieae (Papilionoideae). Flora, 162: 533-548. SIMOLA, L. K., 1968. Comparative studies on number of leaflets, venation, and epidermal structure in the genus Lathyrus. Canadian Journal of Botany, 4 6 : 71-84. SIMOLA, L. K., 1969. A serological comparison of the seed proteins of the genus Lathyrus and certain other genera of the Papilionaceae. Flora, A b t . B, 158: 645-658. SIRJAEV, G . , 1932. Generis Ononis L. revisio critica. Beihefte zum Botanischen Centralblatt, 4 9 ( 2 ) : 381-665. STEARN, W. T., 1966. Botanical Latin. London: Nelson. STOESSL, A., 1972. Inermin associated with pisatin in peas inoculated with the fungus Monilinia fruticola. Canadian Journal o f Biochemistry, 50: 107-108. STREICIIER, O., 1902. Beitrage zur vergleichenden Anatomie der Vicicen. Beihefte z u m Botanischen Centralblatt, 12: 483-538. TAUBERT, P., 1894. Accounts of Icguminous genera in H. G. A. Engler & K. A. E. Pranrl (Eds), Die natiirlicheti Pflanzenfamilien, 3 ( 3 ) . THUKMAN, D. A., BOULTER, D., DERBYSHIRE, E. & TURNER, B. L., 1967. Electrophoretic mobilities of formic and glutamic dehydrogenases in the Fabaceae: a systematic survey. N e w Phytologist, 66: 37-45. TOWNSEND, C. C., 1974. Accounts of leguminous genera in C. C. Townsend & E. Guest (Eds), Flora of Iraq, 3. Baghdad. TSCHIERSCH, B. & HANELT, P., 1967. Dic freien Aminosauren der Samen von Vicia L. und die systematische Gliederung der Gattung. Flora, A b t . A , 15 7: 389-406. TURNER, B. L. & FEARING, 0. S., 1959. Chromosome numbers in the 1,eguminosae 11: African species, including phyletic interpretations. American Journal o f B o t a n y , 4 6 : 49-57. TURNER, B. L. & HARBORNE, J. B., 1967. Distribution of canavanine in the plant kingdom. Phytochemistry. 6: 863-866. VAN DER MAESEN, L. J . C . , 1972. Cicer L., a monograph of the genus, with special reference t o t h e chickpea (Cicer arietinurn), its ecology and cultivation. Mededelingen Landbouwhogeschool Wageningen, 72-10: 1-342. VAN TIEGMEM, P., 1871. Recherches sur la symktrie de structure des plantes vasculaires. Annates des Sciences Naturelles, cinquikme skrie. Botanique, 13: 1-3 14. VAN TIEGHEM, P., 1884a. Sur les faisceaux IibCro-ligneux corticaux des Vicikes. Bulletin de la Soci6t6 Botanique de France, 31: 133-135, VAN TIEGHEM, P., 1884b. Trait6 d e Botanique. Paris. WINTER, C. W., 1932. Vascular system of young plants of Medicago sativa. Botanical Gazette, 94: 1 5 2- 167.