Survey
* Your assessment is very important for improving the workof artificial intelligence, which forms the content of this project
zoological Journal ofthe Linnean Society (1987), 91: 107-135. With 7 figures
The eutherian stapedial artery: character
analysis and implications for superordinal
relationships
JOHN R. WIBLE
flepartment of Anatomy, Uniuersity of Chicago, Chicago, ~ l l ~ 60637,
~ ~ i s U S .A .
Received May 1.986, accepted for publication Septelnber 1986
Evidence from outgroups, ontogeny, neontology, and fossils is used to distinguish primitive and
derived rharacter states for the major components of the eutherian stapedial artery in I 7 modern
orders. Derived states support the following higher-level phylogenetic hypotheses: ( 1)
Microchiroptera and Mcgachiroptera are monophylrtir; and (2) within Ungulata, Tubulidentata is
the outgroup to the remaining modern orders, followcd in succession by Artiodactyla and then
Cetacca. Three branches of the stapedial artery, the a. diploetica magna, ramus temporalis, and
ramus posterior, all but neglerted in previous syntheses, are shown to be primitive for Eutheria and
Amniota.
KEY WORDS:-Stapedial
rclationships rrania.
artery
-
Eutheria
-
mammals
-
character analysis
superordinal
-
~
CONTENTS
Introduction .
,
. . . . . , ,
Material and methods . , , . . .
,
Ramus superior and outgroup comparisons.
Character analysis . .
.
. . . . .
Proximal stapedial artery . . . .
.
Ramus posterior .
.
.
.
.
. .
Ramus superior/ramus inferior bifurcation .
Ramus superior . .
.
. . . .
Meningeal rami . .
.
.
.
. .
Ramus temporalis.
.
. . . . .
Arteria diploetira magna . .
. . .
Ramus inferior
.
.
. . . . .
Maxillary artery . . .
. . . .
Ramus infraorbitalis . .
. . . .
Ramus orbitalis . .
. . . . .
Other anastomoses
. . . . . .
Discussion . .
. . . . . . . .
Lagomorpha, Rodentia, and Macroscelidea
Microchiroptera and Megachiroptera . .
Ungulata.
.
. . . . . . .
Acknowledgements
.
. . . . . .
References.
.
. . . . . . . .
List of abbreviations . . . .
. . .
+
0024-4082/87/100107 29 $03.00/0
,
.
.
,
.
,
.
.
.
,
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
,
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
,
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
108
.
.
.
109
110
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
,
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
111
112
I15
1 16
116
1 17
118
120
I21
123
I24
125
125
I26
126
I28
129
130
I3 I
135
107
0 1987 T h e Linnean
Society of London
.I. K. WlHLE
108
1N'I'KOL) U C'I'ION
The stapedial artery (also called thc orbital, temporal, or orbitoteniporal
artery by some authors) can be traced within Vertebrata from selachians to
mammals (de Beer, 1926; Goodrich, 1930). It is dcrived from the artery of the
second branchial or hyoid arch (the second aortic arch) and arises from the
internal carotid near the hyomaridibula in fish or the stapes (columella auris) in
tetrapods (de Beer, 1926; Goodrich, 1930). Within Mammalia, the stapedial
artery is the original supplicr of blood to the supraorbital, infraorbital, and
mandibular regions in most embryos and also the major supplier to those
regions in many adults (Wible, 1984). For students of mammalian systematics,
the stapedial artery, its branches, arid associated grooves, canals, and foramina
are widely recognized as important sources of traits for rcconstruc.ting
phylogenetic relationships (e.g. Bugge, 1974; Wahlert, 1974; MacPhee &
Cartmill, 1986).
Despite the importance of the stapedial artery in mammalian oritogeriy arid
phylogeny, knowledge of this vessel is in many regards rather limited. Ontogeny
of the stapedial artery has been studied in detail in only 13 forms, representing
nine of the 20 modern orders (Tablc 1). Although the stapedial artery and its
branchcs have been described in a wide assortment of adult mammals, the
homologies of some segments are uncertain. For instance, in most mammals thc
branch of the stapedial artery that supplies the infraorbital region (the ramus
infraorbitalis) runs through the orhitotemporal region ventral to the skull base
(Wible, 1984); however, in dipodoid rodents the supposedly homologous vessel
lies dorsal to the alisphenoid and leaves the cranial cavity with the maxillary
nerve (Bugge, 1971). A number of models, or morphotypes, of the cranial
arteries in ancestral cuthcrians have appeared in recent years (Szalay, 1972,
1975; Bugge, 1974; Archibald, 1977), but these generally have been based on
Table 1. References on Stapedial Artery Ontogeny
in Mammals
Marsupialia
D f l $ y U T U S quo//
Carnivora
canis ,familiaris
Lagomorpha
O~yrtolaguscunicu1u.c.
Rodentia
RauuJ
Erethizon dorsnlum
Chiroptera
Rhinolophus hipposideroj
Vespertilio murinus
Primates
Gnla,<o s~nigalensis
Tarsius spectrum
Homo sapiens
Insec tivora
Talpa europaea
Artiodactyla
.?US Sl'T!fa
Hyraroidea
Procauia capensis
Wiblc (1984)
Wible (1984)
Fuchs (1905)
Tandlrr (1902)
Struthers (1930)
Grosser ( I90 I )
Grosser (1901)
H. Butler (1983)
Hafferl (1916)
Tandler (1902), Padget (1948)
Sicher (1913)
von Hofrnann (1914)
Lindahl & Lundberg (1946)
EU 1 HERIAN STAPEDIAL ARTERY
109
analyses of only a few eutherian orders and have not included data from
relevant outgroups (marsupials, monotremes, and sauropsids, i.e. reptiles, plus
birds). Absence of a broad comparative approach has handicapped these
reconstructions. For example, the ramus temporalis, a branch of the stapedial
artery that supplies the temporalis muscle in some modern eutherians (e.g.
microchiropterans, Grosser, 1901; Buchanan & Arata, 1969; xenarthrans,
Bugge, 1979), has not been included in these morphotypes, yet the antiquity of
this vessel is supported by the occurrence of apparently homologous vessels in
living reptiles ( Cartmill & MacPhee, 1980; Wible, 1984).
This paper synthesizes current ontogenetic, neontological,
and
palaeontological evidence concerning the eutherian stapedial system with
consideration of the pattern in non-eutherian amniotes. A detailed character
analysis of the stapedial artery, its branches, and associated grooves, canals, and
foramina is presented for 17 of the 18 modern eutherian orders (detailed
descriptions are not currently available for Sirenia). This character analysis is
the basis for a reconstruction of the stapedial system in ancestral eutherians and
a discussion of superordinal relationships of certain eutherian groups.
MATERIAL AND ME'I'HODS
T h e method of phylogenetic analysis employed here is a two-step process: ( 1 )
distinguishing homologous and homoplastic features and (2) arranging the
character states of homologous features in a transformation series (morphocline)
to show the direction of evolutionary change (see Bock, 1977).
Positional relationship is used as the major criterion for determining
homology in the amniote stapedial system. However, analyses of positional
relationships are not confined to the adult. Ontogenetic data are included
wherever possiblc, because the process of development provides a level of
resolution to analyses of similarities and differences that is unattainable in the
study of a single stage such as the adult. Embryonic origin is a second criterion
of homology only for the most proximal portion of the stapedial artery, because
of its derivation from the second aortic arch. The remaining portions of tha
stapedial system are secondary outgrowths unrelated to the embryonic aortic
arches, and the timing of their ontogenetic formation varies considerably
between species.
Criteria commonly used for identifying the most primitive (plesiomorphous)
character state in a transformation series include: "( 1) commonality or frequency
of distribution of character states; (2) outgroup comparisons; ( 3 ) analysis of
character covariance in relation to a 'form-function complex'; (4) study of
ontogenetic transformations; and (5) reference to the relative geochronologic
age of taxa with certain character states" (Novacek, 1980: 37). While debate
concerning the validity of certain criteria continues (e.g. Bishop, 1982; Alberch,
1985), I advocate multiple testing with as many relevant criteria as is possible.
Of the five criteria cited above, I rely most on outgroup comparison, ontogeny,
and commonality. Evidence from the fossil record is of value, but only for those
portions of the stapedial system that leave imprints in bone (grooves, canals, or
foramina). Although several factors, including body size and auditory
specializations, are thought to account for changes in the distributional pattern
110
-1 R WlBLE
of the stapedial system in mammals (Fleischer, 1978; Packer, 1983), current
understanding of causation (and resultant predictive power) is very limited.
Therefore, I avoid discussion of 'form function complexes' here.
Most of the data for the character analysis are taken from a comprehensive
study of the ontogeny and phylogeny of the cranial arterial pattern in ten
modern mammalian orders (Monotremata, Marsupialia, Xenarthra, Pholidota,
Carnivora, Lagomorpha, Rodentia, Chiroptera, Artiodactyla, and Cetacea)
and in several reptiles and birds (Wible, 1984). Data for the remaining
manimalian orders are from the following sources: Macroscelidea (Bugge, 1972,
1971; MacPhee, 1977, 1981), Dermoptera (Cartmill & MacPhee, 1980; pers.
obs.), Scandentia (Spatz, 1964; Steuerwald, 1969; Bugge, 1972, 1974; MacPhee,
1977, 1981; Cartmill & MacPhee, 1980), Primates (Bugge, 1972, 1974;
MacPhee & Cartmill, 1986), Insectivora (Bugge, 1972, 1974; MacPhee,
1977, 1981), 'l'ubulidentata (Patterson, 1975; Thewissen, 1985), Perissodactyla
(Tandler, 1899; Sisson, 1914; Radinsky, 1965; Savage, Russell & Louis, 1965;
Cifelli, 1982), Hyracoidea (Lindahl & Lundberg, 1946), and Proboscidea (M.
Watson, 1875; Tassy, 1981).
~
Ramus superior and outgroup comparisons
Determining the homologies of one component of the eutherian stapedial
system, the ramus superior, has proven somewhat difficult. This vessel, one of
the two ma.jor end-branches of the stapedial artery, accompanies the ophthalmic
nerve into the supraorbital region; the ramus inferior, the other end-branch,
bifurcates and gives off arteries that accompany the maxillary and mandibular
nerves into the infraorbital and mandibular regions respectively (Fig. 1). This
distributional pattern - proximal stapedial artery and superior and inferior
rami - is very constant within Eutheria and is known to appear during
ontogeny in some representatives of all orders except Tubulidentata, Cetacea,
Perissodactyla, Proboscidea, and Sirenia.
A similar distributional pattern develops and is retained in the adults of most
reptiles and birds (Hafferl, 1933). T h e only significant difference concerns the
course of the artery accompanying the ophthalmic nerve. I n reptiles and birds,
this vessel follows a wholly extracranial course, lateral to the processus ascendens
of the epipterygoid. I n contrast, the eutherian ramus superior pierces the
tympanic roof and runs forward along the intracranial surface of the alisphenoid
(which either wholly or i n part is the homologue of the processus ascendens of
the epipterygoid). Either: ( I ) the eutherian ramus superior has a homologue in
the sauropsid stapedial system and somehow has been incorporated into the
cranial cavity or (2) the eutherian ramus superior is a neomorphic structure.
The evidence from the cranial arteries in the remaining relevant outgroups
(monotremes and marsupials) is equivocal. 'The stapedial system is greatly
reduced and its end-branches are annexed to the external carotid system in the
adults of all forms except the platypus Ornithorhynchus (Fig. 2). No rnonotreme or
marsupial has a vessel wholly equivalent to the eutherian ramus superior at any
point during ontogeny, supporting no. 2 above. (Ornithorhynchus has a somewhat
similar vessel, but it is wholly extracranial (Fig. 2A).) Yet, some of the endbranches of the stapedial systems in monotremes and marsupials pass through
the cranial cavity and correspond to portions of the eutherian rarnus superior,
EUTHERIAN STAPEDIAL ARTERY
r. sup.
mr
n
1
/
1.-0.
oph. n.
max. n.
ic’a
r.\mand.
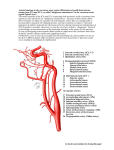
Figure 1. Reconstruction of the internal carotid and stapedial arteries in a hypothetical ancestral
eutherian. A, Ventral view of left basicranium. B, Lateral view of right braincase. The auditory
bulla (the protective shell composed of membrane, cartilage, and bone in varying proportions that
covers the tympanic cavity ventrally) and two of the middle-ear ossicles, the malleus and incus,
have been excluded to expose the arteries. Dashed arteries run within the cranial cavity, and the
stippled portion of the a. diplortica magna lies in a canal between the squamosal and pars
canalicularis of the auditory capsule.
which could be taken as support for no. 1. Because the homologies of the ramus
superior outside Eutheria are unresolved at present, outgroup comparison is not
used as a criterion of morphocline polarity for that vessel.
CHARACTER ANALYSIS
Character state distributions for the various components of the stapedial
system in 17 modern eutherian orders are presented in Table 2, and a
reconstruction of the stapedial system in a hypothetical ancestral eutherian
appears in Fig. 1. Several derived (apomorphous) anastomotic linkages to the
plesiomorphous stapedial system are shown in Fig. 3. Character states are
identified and morphocline polarities are assessed below.
J. R. WIBLE
A
r.
-r.
prox. stap. a.
s.-0.
orb.
u
r. i.-0.
ica
1
CCh
max. a.
\ r . mand.
r. s.-o
r.
\\
ko.
L-
postgl. a.-
max. a.
cca
Figure 2. 'l'he major cranial arteries in monotrrmes and marsupials. A, 'Ihc monotrcme
Ornilhor/ynchu.c (after 'l'andlcr, 1899, 1901; D. M. S. Watson, 1916; Wible, 1984). B, The marsupial
Diddphis (after 'l'andler, 1899; Wible, 1984). Tnchy,&ssuJ differs from Orniihorhiynchzts in that: thc
proximal stapedial, ramus superior (?), and ramus inferior are lacking; the occipital artery
originatcs from the rxterrial carotid (arid not the proximal stapedial); and the meningeal ramus of
thc a. diploetica magna reaches forward to the orbit to supply the ramus supraorbitalis ('l'andler,
1899, 1901; LVible, 1984). Other marsupials that have been studied resemble Didelphis (Tandler,
1899; Archer, 1976; Wible, 1984); the most significant difference is the occurrence of an a.
diploetica magna in some forms. Striped vessels are neomorphic anastomoses (thc stripcd portion of
the ramus supraorbitalis of the opossum represents its origin from the ophthalmic branch of the
intcrual carotid artery). I n the platypus, the stippled portions of the a. diploetica tnagna and ramus
infraorbitalis lie within the posttemporal and alispherioid canals respectively, arid portions of the
proximal stapedial and ramus superior (?) are covered by the expanded mastoid. In the opossum,
the postglenoid artery enters thc cranial cavity via the postglenoid foramen (not visible here) and
divides into a mcriirigcal rarrius arid a temporal ramus that passes through the subsquamosal
foramen.
Proximal stapedial artery
The proximal stapedial artery, the stem of the stapedial, extends from the
internal carotid beyond the stapes to the bifurcation of the superior and inferior
rami. Character states of the proximal stapedial a. concern its site of origin from
the internal carotid and its relationships to the stapes and the surface of the
petrosal bone.
I w o distinct patterns of origin for the proximal stapedial a. occur among
r 7
(3)
absent
absent
absent
absent
absent
INTRATY MP.
?INTRATYMP.
INTRATYMP.
?INTRATYMP.
absent
absent
absent
intrarran.
intracran. *
intracran.
intrarran.
intracran.
absent
r.sup.
origin
no
no
?YES*
sof
YES*
YES*
YES*
?YES*
YES*
?YES*
YES*
COF*
COF*
COF*
COF
COF*
COF*
COF
COF*
COF
COF
COF*
COF*
COF*
no
no
YES*
no
?YES*
?YES*
YES*
no
YES*
YES
YES*
no
no
YES
?YES*
no
no
YES
YES*
YES
YES*
YES*
YES*
YES
YES
YES
solsolsof
COF
(6)
a.d.
mag.
(5)
r.
temp.
(4)
r.sup.
orb.for.
yes
yes2
yes'
yes '
Yes:
Yes
absent
absent
absent
absent
INTRATYMP. *
INTRATYMP.*
NO
INTRATYMP.;
yes 1 .
yes'
INTRATYMP. * yes',
INTRATYMP. * yes '
?INTRATYMP.* yes'
3'
2s 3
yes"5
NO
yes', 2'
yes2
yes'.'
ves
INTRATYMP. *
INTRATYMP.*
INTRATYMP.*
INTRACRAN.
intracran.
intracran.
intracran.*
intracran.
(8)
max
a.
(7)
r.inf.
course
(9)
as canal
as canal
as canal
EXTRACRAN.
EXTRACRAN.
EXTRACRAN.
EXTRACRAN.
EXTRACRAN.
as canal
EXTRACRAN.
as canal
as canal
as canal
intracran.
intracran.
EXTRACRAN.
EXTRACRAN.
EXTRACRAN.
r.i.-o.
course
yes
Yes
Yes
NO**
NO
Yes
Yes
NO
NO?**
NO
NO
NO
NO**
NO
NO**
NO
NO
NO
(11)
r.
anast.
yes
NO
NO
NO
Yes
NO
NO
NO
NO
NO**
NO**
NO**
NO**
NO
NO**
NO
NO
NO
Table 2. Character analysis of the adult stapedial artery in the modern euthrian orders (except Sirenia). Micro- and
Megachiroptera are listed separately to address the issue of bat monophyly or diphyly (see Discussion). Character states that are
plesiomorphous for Eutheria are in upper case; apomorphous states are in lower case. The superscripts on the maxillary artery
refer to the different sorts of maxillary arteries described in the text. * Denotes the loss of that component of the stapedial artery
in some representatives of ar, order; ** denotes the secondary addition of that anastomosis in some representatives of an order
no
no
no
no
no
no
YES*
no
no
no
no
YES*
no
YES*
no
no
no
no
no
XENARTHRA
no
PHOLIDOTA
CARNIVORA
YES*
no
LAGOMORPHA
YES*
RODENTIA
MACROSCELIDEA
YES
MICROCHIROPTERA YES
MEGACHIROPTERA
YES*
no
DERMOPTERA
YES
SCANDENTIA
YES*
PRIMATES
INSECTIVORA
YES
TUBULIDENTATA
YES
YES*
ARTIODACTYLA
CETACEA
no
PERISSODACTYLA
no
no
HYRACOIDEA
PROBOSCIDEA
no
(2)
r.
post
(1)
prox.
stap.a.
Table 2. Character Analysis of the Eutherian Stapedial Artery
I14
J . R. WIBLE
r. anast.
r. orb.
Figure 3. ‘l‘he major cranial arteries in a hypothetical ancestral ruthcrian showing four of the
anastomoses (striped vessels) that occnr among modern forms. (‘l‘hc connection between the a.
diploctica m q n a and orcipital artery is considered plesiomorphous for Eutheria.)
modern mammals. In nearly all euthcrians in which this vessel appears (and in
reptiles, birds, and embryonic marsupials), the proximal stapedial arises from
the internal carotid within the tympanic cavity (middle-ear space) on the
ventral surface of the promontorium of the petrosal bone (the cochlear housing).
‘ l h e only exceptions to this pattern within Mammalia (and Amniota) where the
proximal stapedial artery occurs are found in the monotreme Ornithorhynchu.r and
some muroid rodents. I n these forms, the proximal stapedial leaves the internal
carotid posterior to the auditory bulla. As a consequence, the proximal stapedial
has its own foramen of entrance into the tympanic cavity. This pattern results
from a relative posteromedial movement in the internal carotid and proximal
stapedial arteries during ontogeny in the muroid rodents (and possibly in the
platypus) (Fig. 4) and is treated as an apomorphous character state in these
forms.
The proximal stapedial exhibits two character states in its relationship to the
stapes among modern mammals. I n nearly all forms in which the artery is
present, it passes through an opening (the obturator foramen) in the stapes, but
in the monotreme Ornithorhynchus that opening is absent and the artcry runs
behind the stapes. Some authors (e.g. Gregory, 1910; Segall, 1970) argue that
an imperforate stapes is primitive for Mammalia (and Eutheria) and that an
obturator foramen in the stapes is derived. Others (e.g. Goodrich, 1930;
Archibald, 1979) claim the reverse. T h e latter view is followed here because: ( 1 )
an obturator foramen is found in the stapes of Mesozoic mammals where known
(the Early Jurassic “triconodont” Morganucodon, Kermack, Mussett & Rigney,
1981; and a Late Cretaceous “unguiculate” eutherian, Archibald, 1979) and
many advanced therapsids, including ictidosaurs, the probable sister-group of
Mammalia (Hopson & Barghusen, 1986); and (2) an obturator foramen
appears in the stapes in early ontogenetic stages of the monotreme
Omithorhynchus, but is later lost (Goodrich, 1915, 1930). (See Novacek & Wyss,
1986b, for a different view of character transformation of the eutherian stapes.)
-
EU'IHERIAN STAPEDIAL AKTERY
Early
r.
I I5
Late stages
int.
aud. bulla
Figure 4. Schematic representation of the development of the internal carotid and stapedial arteries
in muroid rodents. In early embryonic stages, the internal carotid runs ventral to the cochlear
capsule (the cartilaginous precursor of the promontorium of the petrosal), and the proximal
stapedial originates in front of the fenestra cochleae. In later stages, the internal carotid and
proximal stapedial shift somewhat posteromedially relative to their prior positions. T h e presumptive
auditory bulla (composed of connctive-tissue fihrrs and thr rctotympanic bone) is added in still
later stages, excluding the origin of the proximal stapedial from the tympanic cavity and producing
separate foramina for the internal carotid and stapedial arteries in the adult. As in other rodents,
the proximal stapedial leaves the tympanic cavity through an opening in the tympanic roof and the
ramus superior/rarnus inferior bifurcation is within the cranial cavity (not visible here).
Ventrally open grooves or closed canals for the proximal stapedial appear in
two places on the surface of the petrosal bone among many eutherians. The first
is on the promontorium between the internal carotid artery and the fenestra
vestibuli (the opening within which the footplate of the stapes sits). T h e second
is on the roof of the tympanic cavity lateral to (or sometimes in common with)
the facial sulcus or canal (the channel which accommodates the facial nerve in
its course through the tympanic cavity). Facial sulci are widely distributed
among monotremes, marsupials, and eutherians (van Kampen, 1905) and are
surely plesiomorphous for Eutheria (and possibly for Mammalia because sulci
appear in some triconodontids, Kermack, 1963; and multituberculates,
Simpson, 1937; Kielan-Jaworowska, Presley & Poplin, 1986). However, the
facial sulcus need not contain a proximal stapedial artery. Separate grooves and
canals on the promontorium and tympanic roof specifically for the proximal
stapedial are absent from non-therian mammals (except some multituberculates:
Sloan, 1979; Kielan-Jaworowska, Presley & Poplin, 1986) and variably
present among Late Cretaceous and Early Tertiary eutherians. Apparently,
osseous channels for the proximal stapedial have been acquired independently
numerous times within Eutheria. Those modern forms in which this vessel leaves
no imprint on the petrosal bone (e.g. some insectivorans and microchiropterans)
have retained the plesiomorphous state.
Ramus posterior
The ramus posterior is a small branch of the proximal stapedial that
originates just before its parent vessel reaches the stapes (Fig. 1A). It runs
posteriorly with the facial nerve to supply the stapedius muscle, the back of the
tympanic cavity, and (sometimes) the mastoid region. I n most adult mammals,
I16
-1. R . W1BI.E
thc ramus posterior loses its connection to the proximal stapedial and
anastomoses with a branch of the occipital or posterior auricular artery (forming
what is termed a stylomastoid artery) (Fig. 3 ) . However, because a proximal
stapedial origin for the ramus posterior is ontogenetically primary in
Monotremata, Xenarthra,
Lagomorpha, Kodentia,
Microchiroptera,
Artiodactyla, and Hyracoidea and a homologous vessel appears in a number of
reptiles (Wible, 1984), I accept i t as plesiornorphous for Eutheria. The ramus
posterior seldom leaves any impression on the petrosal bone among modern
fbrms and has not yet been identified in any fossil skull.
Ranius tuperiorlRamu.5 infirior biJurmtzon
The bifurcation of the proximal stapedial artery into the ramus superior and
ramus inferior exhibits two principal patterns among recent euthcrians. In some
fbrms, thc bifiircation is ventral to the tympanic roof within the middle-ear
cavity (Fig. 1A): the ramus inferior then runs forward into the orbitotemporal
region vcntral to the tympanic roof and the ramus superior moves dorsally into
the cranial cavity through the tympanic roof: In some others (lagoniorphs,
rodents, elephant shrews, and bats), the proximal stapedial pierces the tympanic
roof and the ramus superior/ranius inferior bifurcation is within the cranial
cavity: the ramus inferior then reaches the orbitotemporal region via a course
dorsal to the tympanic roof (Fig. 4). A variant on this latter state is found in the
tree shrew Tufiaia: the ramus superior/ramus inferior bifurcation is within the
cranial cavity, but thc ramus inferior then runs forward vcntral to the tympanic
roof (MacPhee, 1977, 1981, personal communication). However, this pattern is
not found i n all tree shrews; in Piitocercus the bifurcation is ventral to the
tympanic roof (Zeller, 1986). The intratympanic bifurcation is accepted as
pleisomorphous for Eutlieria chiefly because that is the pattern in the
monotreme Ornithorhynchus, the only non-eutherian mammal in which a
complcte ramus inferior appears in the adult (Fig. 2A). Also, the lesser petrosal
nerve, a branch of the glossopharyngeal nerve that runs with (or near) the
ramus inferior in the platypus and eutherians (whether the artery is ventral or
dorsal to the tympanic roof), lies ventral to the tympanic roof in the monotreme
Tachyglossur and in marsupials.
‘ l w o sorts of openings transmit the proximal stapedial or ramus superior
through the tympanic roof in modern eutherians. Some forms have a discrete
foramen in the osseous tympanic roof, while in others there is merely a gap (the
piriform fenestra of McDowell, 1958) between the various bony elements
contributing to thc tympanic roof. As MacPhee (1981) suggests, a small piriform
fenestra may be the plesiomorphous state for Eutheria, but the incidence of the
various tympanic-roof openings among fossil forms is not well known.
Ramus superior
The eutherian ramus superior irrigates a very large area with branches that
supply structures on both the intra- and extracranial surfaces of the occipital,
orbitotemporal, and ethmoidal regions. From its origin on the proximal
stapedial a., the ramus superior stretches dorsally along the intracranial surface
o f the squamosal. I t moves medial to the petrosquamous sinus (the dural sinus
EV I HERIAN STAPEDIAL ARTERY
117
that drains into the external jugular vein via the postglenoid foramen) and sends
a large branch off posteriorly. This posterior branch of the ramus superior in
turn supplies meningeal rami, one or more temporal rami, and an arteria
diploetica magna that are discussed separately below. The main (or anterior)
portion of the ramus superior runs forward medial to the squamosal, parietal,
and alisphenoid (or sometimes frontal), supplying additional meningeal rami en
route. Entering the back of the orbit, the ramus superior (more correctly now,
the ramus supraorbitalis) ends in vessels that travcl with the lacrimal, frontal,
and ethmoidal branches of the ophthalmic nerve.
‘The only character states of the eutherian ramus superior considered here
concern its foramen of entrance into the orbit. The ramus superior either passes
through the superior orbital fissure between the alisphenoid and orbitosphenoid
or through a separate, more dorsally situated foramen lying near the juncture of
the alisphenoid, orbitosphenoid, parietal, and frontal. A separate foramen
(generally called the cranio-orbital or sinus canal foramen) is accepted as
plesiomorphous for Eutheria, because that opening is more widespread among
the modern orders and also appears in many Late Cretaceous and Early
Tertiary forms (e.g. kennalestids, Kielan-Jaworowska, 1981; leptictids, P. M.
Butler, 1956; arctocyonids, Russell, 1964).
T h e intracranial portions of the ramus superior generally occupy grooves on
the medial surfaces of the bones in the side wall of the braincase. The most
prominent of these vascular grooves, the one for the anterior portion of the
ramus superior, has been called variously the sinus canal (Parker, 1885), the
lateral cerebral sinus (P. M. Butler, 1948), and the cranio-orbital sinus
(Cartmill & MacPhee, 1980). Each of these terms implies that the major
occupant of this vascular groove is the venous channel connecting the orbital
and petrosquamous sinuses. Veins accompany the anterior portion of the ramus
superior in the eutherians (and monotremes) investigated here, but the major
occupant of the vascular groove is the artery. Similar grooves appear in extinct
non-therian mammals (e.g. Morganucodon, Kermack et al., 1981; multituberculates, Kielan-Jaworowska et al., 1986) and advanced therapsids (Fourie,
1974) and, as Kielan-Jaworowska et al. (1986) suggest, may have bcen chiefly
arterial in nature in these forms.
Meningeal rami
One or more meningeal rami originating from the intracranial portions of the
ramus superior have been reported in representatives of all extant mammalian
orders (except Sirenia) and are surely plesiomorphous for Eutheria and
Mammalia. These rami course dorsally along the intracranial surfaces of the
squamosal and parietal within the dura mater. In general, these vessels have
been termed collectively the ‘middle meningeal artery’, but this usage is avoided
here for the following reason. ‘Middle meningeal artery’ is a term of human
anatomy, and the vessel in Homo may not be wholly homologous with the
meningeal rami of the ramus superior. MacPhee & Cartmill (1986) suggest
from positional relationships that the middle meningeal artery of man and other
catarrhine primates is actually a compound vessel: its intracranial meningeal
rami derive from the ramus superior, and its extracranial stem is a neomorph (a
ramus anastomoticus) that grows out from the ramus inferior to annex the
I18
J. R. WIBLE
meningeal rami (Fig. 3 ) (which thereafter loses any connection with stapedialartery remnants within the tympanic cavity). Until the homologies of the
human middle meningeal artery are resolved through detailed reanalyses of
relevant ontogenetic stages, I recommend the term ‘meningeal rami’ [or the
meningeal branches of the eutherian ramus superior.
Ramu.r temporalis
The eutherian ramus temporalis arises from the posterior branch of the ramus
superior along the intracranial surface of the squamosal. It leaves the cranial
cavity via foramina within or between the squamosal and parietal (dorsal to the
external acoustic meatus) and supplies the posterior portion of the temporalis
muscle. (The remaining portions of the temporalis muscle are supplied through
branches of the external carotid artery.) One or more rami temporales have
been reported for representatives of nine modern orders, but may be even more
widespread within Euthcria (including Late Cretaceous and Early Tertiary
forms) judging from the frequent occurrence of foramina along or near the
parietosquamous juncture (see Cope, 1880; Russell, 1964; Kielan-Jaworowska,
1981). Such foramina are generally thought (e.g. Savage el al., 1965; Tassy,
1981) to be entirely venous in fossil forms, but this view must be amended in
light of the widespread distribution of temporal rami among modern eutherians.
Temporal rami are absent in all recent Dermoptera, Tubulidentata, Cetacea,
Hyracoidea, and Proboscidea investigated to date, and the temporalis muscle in
these forms is supplied exclusively through branches of the external carotid
artery. The loss of the temporal rami is presumably the result of the decreased
importance of the stapedial system in these groups.
A stapedial origin for the temporal supply is accepted as plesiomorphous for
Eutheria in part because a similar pattern appears in other amniotes. However,
there is a difference in the course of this vasculature between modern therians
and non-therian amniotes. In the latter forms, the vascular supply to the
adductor mandibulae externus (in monotremes, the temporalis muscle) follows a
wholly extracranial course, arising outside the braincase and running directly
into the temporal region (Fig. 2A). O n the other hand, in most therians, the
ramus temporalis originates within the cranial cavity and must pierce the side
wall of the braincase (squamosal and/or parietal bones) to reach the temporalis
muscle. (The eutherian and marsupial temporal rami have different vessels of
origin within the cranial cavity: the posterior branch of the ramus superior in
eutherians and, in marsupials, a branch of the external carotid system that
enters the cranial cavity via the postglenoid foramen with the petrosquamous
sinus (Fig. 2B).)
The intracranial origin of the therian ramus temporalis appears to result from
the expansion of the squamosal bone medial to the temporalis muscle. In
advanced therapsids (Barghusen, 1968), extinct non-therian mammals (KielanJaworowska, 1971; Kermack et al., 1981; Crompton & Sun, 1985), and modern
monotremes, the adductor mandibulae externus (or temporalis muscle) originates in part on the extracranial surface of the auditory capsule (Fig. 5A). In
contrast, in therians, a portion of the squamosal bone (the squama) is interposed
between the temporalis muscle on the one hand and the auditory capsule and
ramus superior (or the postglenoid artery, in marsupials) on the other (Fig. 5B).
EUTHERIAN STAPEDIAL ARTERY
A
P a r s canallcularis
119
,squamosal
brain
cochlear
capsule
\
r. temp.
incus
ica
ectopterygoid
I . “-----1
/
t e n s o r t y m p a n i m.
dorsal
ectotympanlc
t
-lateral
parietal
p r o x . s t a p . a.
B
I
brain
r . sup./r. inf.
bifurcation
cochlear
capsule
t e n s o r tympani m:
I
entotympanic
U
ectotympanic
Figure 5. Schematic cross sections through the middle portions of developing tympanic regions,
showing the varying relationship between the ramus temporalis, temporalis muscle, and the side
wall of the braincase. A, 92 mm crown-rump length pouch young of the monotreme 7uchyglossus
aculeatus (Duke University Comparative Embryological Collection no. 8327: section 622). B,
93.5 mm crown-rump length foetus of the megachiropteran Pleropus (Duke University Comparative
Embryological Collection no. 83 1: scrtiori 1451). Shading by open circles indicates elements
prrformed in cartilage; cross hatching denotes dermal bone. Note in B that ( I ) the proximal
stapedial and ramus superior/ramus inferior bifurcation are dorsal to the tympanic roof (tegmen
tympani), an apomorphous state found in lagomorphs, rodents, elephant shrews, and bats and (2)
the tegmen tympani has a process jutting ventrally into the middle ear that forms continuous with
the cartilage of the auditory tube, a bat autapomorphy.
120
.J. R. WIBLE
Consequently, the ramus temporalis pierces this neomorphic portion of the
squamosal bone to reac,h the temporalis muscle.
Arteria diploetica magna
‘l’iie arteria diploetica magna is a little-known branch of the mammalian
stapedial system that connects the posterior branch of the ramus superior and
the occipital (or posterior auricular) branch of the external carotid system. T h e
a. diploetica magna was first described in an adult monotreme, the Tasmanian
echidna 7~achyglossu.rsetosus (“Echidna setasus”), by Hyrtl in 1853. In that form,
the artery originates from the posterior branch of the ramus superior on the
intracranial surface of the parietal. It leaves the cranial cavity between the
parietal and the canalicular part of the petrosal (the portion that houses the
u triclc and semicircular canals) and runs posteriorly in an extracranial space
between the petrosal and squamosal that also includes part of the temporalis
m u d e (Fig. 5A; Kuhn, 1971). This space, the posttemporal canal, opens onto
the occiput via the posttemporal foramen between the petrosal and squamosal,
and there the a. diploetica magna joins the occipital artery. Hyrtl (1854) next
described this vessel in several adult xenarthrans, Tamandua tetradacgla
(“M-Yrmecophaga tamandua”) and Dag~pusnovemcinctus. The a. diploetica magna of
these forms differs from that of the echidna in that it no longer runs with the
temporalis muscle in the posttemporal canal, but is separated from that muscle
because of the expansion of the squama of the squamosal bone discussed above
(Fig. 5B).
Until recently, the a. diploetica magna has been thought (e.g. Tandler, 1899,
1901; Bugge, 1979) to be a peculiarity of these few forms. However, my studies
(Wible, 1984) reveal that this vessel also appears in the monotreme
Ornilhurhynchus (Fig. 2A) and in foetal and/or adult representatives of
Marsupialia, Pholidota, Lagomorpha, Megachiroptera, Dermoptera, and
Artiodactyla. In addition, an a. diploetica magna is figured (but not labelled)
for a neonatal tree shrew Tupnia by Spatz (1964: figs 25, 26), and the caudal
meningeal artery of the adult horse, which originates from the occipital artery
and runs forward through a canal between the petrosal and squamosal (Sisson,
19 14), probably represents an a. diploetica magna. Finally, a probable
homologue for the mammalian a. diploetica magna appears in a diverse lot of
reptiles and birds. This branch of the stapedial system runs posteriorly along the
auditory capsule through the region equivalent to the monotreme posttemporal
canal - that is, medial to the adductor mandibulae externus and squamosal and ends in the occipital region. Such a vessel has been reported in crocodilians
(Shiino, 1914), chelonians (Shindo, 1914; McDowell, 1961; Albrecht, 1976),
lacertilians (Bhatia & Dayal, 1933; Oelrich, 1956), and birds (Wible, 1984).
Given this wide distribution among reptiles, birds, and mammals, the a.
diploetica magna is considered to be a plesiomorphous branch of the stapedial
system in Amniota, Mammalia, and Eutheria.
The a. diploetica magna has generally been overlooked by comparative
anatomists and may be even more widespread within Amniota than is apparent
from published sources. Large gaps (posttemporal foramina) connect the
occipital and temporal regions in many non-therian amniotes, including
advanced therapsids (e.g. Thrinaxodun, Fourie, 1974) and extinct nontherian
EC'THERIAN STAPEDIAL AR r m y
121
niammals (e.g. Morganucodon, Kermack el al., 1981). These openings are thought
by these and other authors to have transmitted only venous channels in these
forms, similar to what has been reported for the colubrid snake Nalrix (Bruner,
1907) and the sand lizard Lacerta ngilis (Shindo, 1915). However, in light of the
broad distribution of the a. diploetica magna among extant amniotes, the
posttemporal foramen of advanced therapsids and extinct non-therian mammals
probably contained an artery as well and, as Kielan-Jaworowska el al. (1986)
have recently pointed out, may have been chiefly arterial in nature. T h e same
holds for the smaller openings between the posterior border of the squamosal
and canalicular part of the petrosal (postsquamosal foramina) that frequently
appear in extinct and extant therian skulls (see Cope, 1880; Russell, 1964).
Because the a . diploetica magna has received such little attention, its
character states are not well understood. T h e therian vessel differs from that of
monotremes in its relationships to the temporalis muscle and squamosal bone.
The mammalian vessel is distinguished from that of other amniotes by the
formation of an anastomosis to the external carotid system (Figs 2A, 3), and this
pattern may be plesiomorphous for Mammalia and Eutheria. Within
Mammalia, differences in the completeness of the anastomotic link between the
stapedial and external carotid systems are as follows: ( 1 ) the a. diploetica magna
forms a complete anastomotic link in the monotreme TnchygloJsus, dasypodid
xenarthrans, and some marsupials; (2) it fails to reach the external carotid
system in the marsupial Petrogale, the lagomorph O~c1olagus cuniculus, the
megachiropteran Pteropuf, and the pholidotan Manis javanica; and (3) it loses its
connection to the ramus superior in the monotreme Ornithorhynchus, bradypodid
and myrmecophagid xenarthrans, dermopterans, the lagomorph Ochotona
princeps, the artiodacty Tragulus, and the perissodactyl Equus.
Rnmus inferior
From its origin on the proximal stapedial artery, the ramus inferior runs
forward into the temporal fossa and bifurcates into infraorbital and mandibular
rami behind the mandibular nerve. In most eutherians and the monotreme
Orni/horhynchus, the ramus inferior arises within the middle-ear cavity and travels
ventral to the osseous tympanic roof next to the lesser petrosal nerve (or between
the lesser petrosal and chorda tympani nerves). (As stated above, the tree shrew
Tupaia exhibits a variant on this: the ramus inferior originates within the cranial
cavity, but then exits that space and runs forward ventral to the tympanic roof.)
T h e artery's passage beneath the tympanic roof is sometimes marked by a
groove (e.g. some insectivorans) or canal (e.g. some scandentians), but these are
certainly derived features because they are absent in extinct non-therian
mammals (except some multituberculates; Kielan-Jaworowska el al., 1986)
and most early eutherians. It is often said (e.g. van Kampen, 1905; van der
Klaauw, 1931; MacPhee, 1981) that the ramus inferior sometimes runs in the
petrotympanic (Glaserian) fissure, the groove in the tympanic roof for the
chorda tympani nerve that is well developed among modern mammals only in
insectivorans, tubulidentates, hyracoids, and some primates (Novacek, 1986).
However, the only specific examples of a ramus inferior in the Glaserian fissure
are in some primates (MacPhee & Cartmill, 1986), and the homologies of
that vessel are controversial (see below). I n several eutherian orders (i.e.
122
J R WlBLE
Xenarthra, Pholidota, Carnivora, Dermoptera, Artiodactyla, and Perissodactyla), the greatly reduced ramus inferior loses its attachment to the proximal
stapedial and is annexed to the external carotid system as a tympanic branch of
the maxillary artery (Fig. 3 ) . It is not certain whether this represents the
plesiomorphous condition for the ramus inferior in these groups or whether the
artery is secondarily reduced.
The ramus inferior exhibits a unique pattern in Lagomorpha, Rodentia,
Macroscelidea, and Chiroptera: it arises from the proximal stapedial within the
cranial cavity and runs forward dorsal to the O S S ~ O U S tympanic roof (Fig. 4).
After this intracranial course, the ramus inferior enters the temporal fossa eithcr
through the piriform fenestra in lagomorphs, chiropterans, and most rodents or
adjacent to the foramen ovale in macroscelideans. In dipodoid rodents, the
ramus inferior continues forward within the cranial cavity and enters the
orbit, as the ramus infraorbitalis, through the confluent superior orbital
fissure/foramen rotundum. The ramus inferior in one extinct eutherian group,
the Leptictidae, is thought by Novacek (1980, 1986) to have passed dorsal to the
tympanic roof, but he remarks that the direct evidence for that pathway is
somewhat ambiguous.
The intracranial course of the ramus inferior can be interpreted in several
ways:
( 1 ) The intracranial ramus inferior is not the homologue of the ramus inferior
of other mammals, but a new anastomotic channel that forms dorsal to the
tympanic roof.
(2) It is the homologue of the ramus inferior of other mammals, and its course
has been shifted dorsally relative to the tympanic roof.
(3) It is the homologue of the ramus inferior of other mammals, but the
tympanic roof has been shifted ventrally relative t o the artery.
The first alternative is rejected, because the artery in question clearly is the
homologue of the intratyrnpanic ramus inferior: both vessels run with the lesser
petrosal nerve. If the true ramus inferior and lesser petrosal nerve have shifted
dorsal to the tympanic roof in lagomorphs, rodents, macroscelideans, and
chiropterans (no. 2), the developmental process involved may be similar to the
relative posteromedial shift of the internal carotid and proximal stapedial
arteries that occurs during ontogeny in some tnuroid rodents (Fig. 4).However,
no apparent relative movement of the ramus inferior and lesser petrosal nerve
during development has been found in the lagomorphs, rodents, and
chiropterans studied by the author (Wible, 1984). At this time, the third
alternative receives the most support, because there is some controversy about
the homologies of one of the major tympanic-roof elements, the tegmcn
tympani. This element, a mammalian neomorph (van der Klaauw, 1931; d e
Brcr, 1937), forms in diverse ways in different groups of mammals. Based on
these differences in ontogenetic formation, Kuhn (197 1 ) suggests that the
tegmen tympani (or portions of it) may have been added to the tympanic roof
independently numerous times within Mammalia. T h e variable relationship
between the tegmen tympani and lesser petrosal nerve described above is strong
support for Kuhn’s hypothesis, because nerve courses through the tympanic
region have been found to be very conservative among eutherians (MacPhee,
1977, 1981).
EU’I’HERIAN STAPEDIAL ARTERY
I23
The fate of the ramus inferior in adult humans (and in other primates) is a
matter of some debate. According to most investigators (e.g. Tandler, 1902;
Padget, 1948; Bugge, 1974), the ramus inferior is the extracranial stem of the
human middle meningeal artery, which enters the cranial cavity immediately
posterior to the mandibular nerve through the foramen ovale or its own foramen
spinosum. MacPhee & Cartmill (1986) point out that this vessel, unlike the
ramus inferior of other mammals, has no course along the tympanic roof. They
suggest that the stem of the human middle meningeal artery is actually a ramus
anastomoticus, a neomorphic vessel that enters the cranial cavity posterior to
the mandibular nerve and anatomoses with the meningeal rami of the ramus
superior (Fig. 3) in, for example, many carnivorans and ungulates. MacPhee &
Cartmill further suggest that the ramus inferior is represented in adult humans
by the anterior tympanic branch of the maxillary artery, a small vessel that runs
with the chorda tympani nerve in the Glaserian fissure on the ventral surface of
the tympanic roof. T h e controversy concerning the fate of the ramus inferior in
adult humans (and in other primates) awaits detailed investigations of relevant
ontogenetic stages.
Maxillary artery
In all recent mammals except some insectivorans and rodents, an anastomosis
forms between the distal portions of the external carotid system and ramus
inferior (or one of its major end-branches, the ramus infraorbitalis or ramus
mandibularis) near the exit of the mandibular nerve from the cranial cavity
(Fig. 3 ) . These anastomotic links, all of which are generally called maxillary
arteries, exhibit a remarkable degree of variability in their origins, courses, and
relationships to other structures. In addition, some forms have more than one
type of anastomotic channel (e.g. dermopterans), I interpret this variability as
evidence for the independent acquisition of anastomotic links between the
stapedial and external carotid systems many times during mammalian
phylogeny. Included under the category ‘maxillary artery’ are the following
sorts of vessels:
( 1 ) An artery leaves the external carotid stem and moves forward medial to
the mandible. It bends medially dorsal to Meckel’s cartilage and passes
posterior to the lateral pterygoid muscle. After supplying the ramus
mandibularis, it turns forward medial to the mandibular nerve beneath the
foramen ovale. This sort of maxillary artery appears in the monotreme
Omithorhynchus, megachiropterans, scandentians, perissodactyls, hyracoids, and
some xenarthrans, carnivorans, rodents, microchiropterans, and insectivorans. It
also appears in macroscelideans, but supplies only the ramus mandibularis in
the adult (Bugge, 1972, 1974). Apparently, the connection between the
infraorbital and mandibular rami is secondarily lost in elephant shrews.
(2) An artery runs lateral to the mandibular nerve beneath the foramen
ovale, but is otherwise similar to the first type (no. 1 above). This sort of maxillary artery appears in the monotreme Tachyglossus, pholidotans, artiodactyls,
cetaceans, and some xenarthrans, carnivorans, and microchiropterans.
( 3 ) An artery sends channels both medial and lateral to the mandibular nerve
beneath the foramen ovale. This sort of maxillary artery occurs in the
124
J. K. \.YIBLl<
xenarthran Dasypus villosus (Tandler, 190 1) and the microchiropteran Artibeus
l i t u r ~ ~ u(Buchanan
s
8r Arata, 1969).
(4) An artery runs forward between the mandible and Meckel’s cartilage,
passing lateral to the mandibular nerve, but separated from i t by the lateral
pterygoid muscle. It supplies a ramus mandibularis and turns dorsally in front
of the lateral pterygoid muscle. This maxillary artery appears in marsupials,
dermopterans, primates, and the xenarthran Tamandua letradacpla.
(5) An artery leaves the external carotid stem and moves forward directly
beneath the tympanic region. It then passes dorsally in front of the auditory
bulla, well medial to Meckel’s cartilage. After supplying the ramus
mandibularis, it turns forward medial to the mandibular nerve beneath the
foramen ovale. This vessel, the pterygoorbital artery of some authors, appears in
lagomorphs, dermopterans, and some rodents.
The taxonomic significance of these various anastomoses is uncertain. Because
the same anastomotic patterns are repeated within certain taxa, it would
appear, at least in some cases, that these ‘maxillary arteries’ may be reliable
indicators of phyletic relationship. I suspect that this is in particular relevant
with regard to nos 4 arid 5 above, vessels which have a very restricted
distribution within Eutheria. However, until more is known about intrataxon
variation, I suggest caution. In the form that is known best, Homo .vapien.s, the
maxillary artery varies considerably both in its relationship to the inferior head
of the lateral pterygoid muscle and to some of the branches of the rnandibular
nerve (Hollinshead, 1968).
The only forms for which some sort of maxillary artery does not occur are in
the orders Insectivora (chrysochlorids, soricids, and erinaccids) and Rodentia
(dipodoids and some muroids and geomyoids). In each case in which the course
of the ramus inferior has been described or figured (except the dipodoid
rodents) , that vessel after supplying the ramus mandibularis continues forward
medial to the mandibular nerve beneath the foramen ovale (Tandler, 1899;
ROUX,1947; P. M. Butler, 1948; Wible, 1984). This course probably is the
plesiomorphous one for the eutherian ramus inferior. ’The ramus inferior and
ramus infraorbitalis in dipodoid rodents run dorsal to the skull base within the
cranial cavity (Bugge, 1971).
Ramus infraorbilalis
From its origin on the ramus inferior (or maxillary artery), the ramus
infraorbitalis runs forward through the orbitotcmporal region of the skull (Fig.
1B). In some forms (for example, the monotreme Tachyglossus, marsupials, and
xenarthrans), the ramus infraorbitalis runs directly ventral to the skull base. In
others (for example, the monotreme OrnilhorhynchuJ, lagomorphs, and
scandentians), the ramus infraorbitalis passes through a canal (or opening) on
the ventral surface of the skull base, generally termed an alisphenoid canal (Fig.
2A). Some authors (e.g. Novacek, 1980; Thewissen, 1985) support a n
alisphenoid canal as plesiomorphous for Eutheria and its absence as an
apomorphous state. However, because an alisphenoid canal is absent in extinct
and extant non-eutherian mammals (except the platypus) and because i t
exhibits a somewhat spotty distribution among extinct eutherians and some
modern groups (e.g. insectivorans, carnivorans, and rodents), I and others (e.g.
EU‘I’HERIAN S‘IAPEDIAL AK’I’ERY
125
Novacek, 1986) consider that this channel has been acquired independently
numerous times within Mammalia. Also, alisphenoid canals do not develop in
the same way in all forms. In some, these channels form within portions of the
chondrocranium (for example, in the ala temporalis in the monotreme
Ornithorhynchus; D. M. S. Watson, 1916; in the pterygoid process of the ala
temporalis in the lagomorph Lepus, de Beer, 1937). In others (for example, the
scandentian Tupaia; Spatz, 1964), the alisphenoid canal lies between the
chondrocranial and dermal portions of the alisphenoid.
Additional apomorphous states of the ramus infraorbitalis occur in dipodoid
rodents and chiropterans. In these forms, the ramus infraorbitalis runs forward
into the back of the orbit dorsal to the alisphenoid. ‘The chiropteran vessel enters
and exits the cranial cavity through foramen ovale (or a separate opening in
front of foramen ovale) and foramen rotundum respectively. I n dipodoid
rodents, the ramus infraorbitalis originates from the ramus inferior within the
cranial cavity and enters the back of the orbit through the confluent superior
orbital fissure/foramen rotundum. Because these two variants exhibit different
relationships to the skull base, they are probably not wholly homologous, nor
are they homologous with the extracranial ramus infraorbitalis of other
mammals. This is supported by the concurrent incidence of both intra- and
extracranial infraorbital rami in the microchiropteran Artibeus lituratus
(Buchanan & Arata, 1969). (The intracranial ramus infraorbitalis is absent in
two microchiropterans, the rhinolophid Rhinolophus (Grosser, 1901 ; Kallen,
1977) and the phyllostomatid Desmodus (Kallen, 1977). I view these as derived
conditions, because a separate foramen in front of the foramen ovale appears in
other representatives of these families.)
Ramus orbitalis
Anastomoses frequently occur between the ramus infraorbitalis and the
supraorbital branches of the ramus superior in extant mammals. T h e most
frequent pattern for such anastomotic channels is a course through the back of
the orbit outside the periorbita (Fig. 3 ) . However, in dipodoid rodents,
megachiropterans, and some microchiropterans, this vessel arises from the ramus
infraorbitalis within the cranial cavity and enters the back of the orbit through
the foramen rotundum (or the confluent superior orbital fissure/foramen
rotundum). Also, in some forms (e.g. pholidotans, dermopterans) the
supraorbital vessels originate directly from the ramus infraorbitalis and not from
a separate anastomotic trunk. Because these vessels, which fall under the
category ‘ramus orbitalis’, are absent from some extant forms and because they
tend to be somewhat variable in their courses and relationships, I treat them as
apomorphous states. (The term ‘ramus orbitalis’ is equivalent to the ‘external
ophthalmic artery’ of the Nomzna anatomica ueterinaria, 1973, but is preferred here
to avoid confusion with the ophthalmic branch of the internal carotid artery.)
Other anastomoses
Two other sorts of anastomotic channels join intracranial portions of the
ramus superior. These vessels exhibit somewhat spotty distributions among
modern mammals and apparently have arisen numerous times:
126
J . R. WIBLE
( 1 ) The ramus anastomoticus connects the maxillary artery to meningeal
branches of the ramus superior through the foramen ovale (Fig. 3) in the
Tachyglossus, dermoptcrans, tubulidentates,
perissodactyls,
monotrcmc
hyracoids, proboscideans, and some carnivorans, rodents, artiodactyls, and
cetaceans (and possibly some primates; MacPhee & Cartmill, 1986).
(2) The arteria anastomotica, which joins intracranial portions of the ramus
superior and internal carotid artery, appears in cetaceans, dermopterans, and
some xenarthrans, pholidotans, carnivorans, rodents, and artiodactyls.
D 1SC U SS10 N
As is apparent in the foregoing character analysis (Table 2 ) , no modern
eutherian retains all the primary components of the plesiomorphous stapedial
system. The forms that most closely approximate the morphotypical pattern are
certain insectivorans, which lack only an arteria diploetica magna. (This artery,
in fact, may be present in some insectivorans, for instance, Eb1omy.r and
Echinosorex, given the occurrence of well-developed postsquamosal foramina in
macerated skulls.) In all modern eutherians studied to date, at least one major
component of the stapedial system is lost or reduced and its end-branches are
annexed to the external carotid system.
Thc character analysis presented in Table 2 also reveals that many of the
apomorphous states of the stapedial system are shared among the modern
orders. These shared apomorphous states deserve further analysis, in particular
because few synapomorphies of orders have been identified to date in syntheses
of fossil and anatomical evidence on higher eutherian phylogeny (e.g. Novacek,
1980, 1982, 1986; MacPhee & Cartmill, 1986). T h e distribution of these
apomorphous states of the stapedial system is such that some apparently have
been acquired independently several times within Eutheria and are not reliable
evidence of any superordinal affinity (for example, the loss of the ramus
temporalis or the addition of an a. anastomotica). Other states, however, are
consonant with superordinal groupings that have been proposed or supported
by other researchers (e.g. McKenna, 1975; Szalay, 1977; Novacek, 1982, 1986).
These groupings and apomorphous states are discussed below.
Lagomorpha, Rodentia, and Macroscelidea
There is a general consensus of late among paleontologists (e.g. McKenna,
1975; Szalay, 1977; Novacek, 1982, 1986) that the diverse assortment of extinct
pseudictopids, zalambdalestids, eurymylids, and anagalids from the Late
Cretaceous and Early Tertiary of Asia forms a closely knit group, which is in or
near the ancestry of several modern eutherian orders
in particular,
Lagomorpha and Macroscelidea. Other groups that have been added to this
grouping include the extinct palaeoryctines and leptictids (McKenna, 1975)
and Rodentia (Novacek, 1982); i n fact, Novacek (1982) has revived Linnaeus’s
original concept of Glires for lagomorphs and rodents and placed Macroscelidea
as the sister group to Glires (Fig. 6). The grouping of these very diverse extinct
and extant forms, which is equivalent to McKenna’s (1975) magnorder
Ernotheria plus Rodentia, is supported by numerous cranioskeletal and dental
features (discussed in the publications cited above and others cited therein), and
~
EUTHERIAN STAPEDIAL ARTERY
127
Figure 6. Phylogenetic relationships for the major groups of Recent and fossil (*) eutherians
suggested by Novacek (1982: fig. I ) . Dashed linrs indicate highly tentative relationships. (Following
Engelmann (l978), I use the term “Xenarthra” instead of “Edentata” for the monophyktir
grouping of New World rdrntates.)
the monophyly of Glires is supported by several derived features of foetalmembrane development and placental morphology (Luckett, 1977, 1985) and a
wide assortment of cranioskeletal and dental features (Li & Ting, 1985; Luckett,
1985; Novacek, 1985).
In the character analysis presented here, a derived pattern for the closure of
the tympanic roof with the tegmen tympani ventral to the ramus superior/ramus
inferior bifurcation and ramus inferior course is shared by Lagomorpha,
Rodentia, and Macroscelidea and possibly by some of the extinct members of
the ernothere-rodent clade (i.e. leptictids, Novacek, 1980; Pulueoryctes, pers.
obs.). (Novacek’s (1986) proposal that leptictids are more closely related to
insectivorans requires independent acquisition of the derived pattern of the
ramus superiorlramus inferior bifurcation in these fossils.) The incidence of this
feature among other extinct forms in this clade is not known, because the
tympanic regions are either covered by a complete osseous bulla (e.g. Anugule;
McKenna, 1963) or have rock matrix covering the tegmen tympani (e.g.
Kennalestes and Asioryctes; pers. obs.). This apomorphous state is not unique to
ernotheres and rodents, but appears in one other group, Chiroptera. Bats either
have independently acquired this pattern of tympanic-roof closure or they may
be the sister group of the ernothere-rodent clade. (The sub- and superordinal
affinities of bats are discussed further below.)
Another apomorphous feature that appears in many extinct and extant
members of the ernothere-rodent clade (as well as in a number of other
see Table 2) is the enclosure of the ramus infraorbitalis within a
mammals
canal in the alisphenoid bone. An alisphenoid canal is known for lagomorphs,
~
I28
J R WlBLE
rodents, macroscelideans (Saban, 1956), leptictids (Novacek, 1980), anagalids
(McKenna, 1963), and Pnlaeoryctes (Van Valen, 1966). It develops in a similar
position within the cartilaginous ala temporalis in modern representatives of this
clade, such as the lagomorph Lepus (de Beer, 1937), the rodents Rattus (Youssef,
1966) and Tntpra (Kadam, 1973), and the macroscelideari Elephunlulu~ ( ROUX,
1947), and may be a synapomorphy of the modern orders. Howcver, because an
alisphenoid canal is absent in the earliest representatives of the ernothere-rodent
clade, the kennalestids (Kielan-Jaworowska, 1981) and zalambdalestids
(Kielan-Jaworowska & Trofimov, 1980), it is not a synapomorphy of this group.
Microchiroptern and Megachiroptera
Bats have traditionally becn thought to be monophyletic (Gregory, 1910;
Simpson, 1945), but separate origins for micro- and megachiropterans within
Eutheria recently have been supported by several investigatorq. Smith &
Madkour (1980) use two features of the penis (absence of accessory cavernous
tissue and presence of distally expanded, vascular corpus spongiosum) to suggest
“a closer phylogenetic relationship between Megachiroptera, Dermoptera, and
Primates than was previously supposed between Mega and Microchiroptera
[ 1980: 3631.” Pettigrew (1986) and Pettigrew & Cooper (1986) report derived
features of the visual pathways unique to Megachiroptera, Dermoptera, and
Primates among vertebrates and suggest that megachiropterans and flying
lemurs may be an early branch of the primate line (“aerial primates”).
Pettigrew (1986) dismisses the 16 shared derived features employed by Novacek
( 1982) to link micro- and megachiropterans, because they are either associatcd
with the adaptation to flight (possible convergences), not unique to bats within
Eutheria, or possible plesiomorphies.
Diphyletic origins for micro- and inegachiropterans are not supported by my
character analysis of the eutherian stapedial artery. No shared derived features
separately unite Micro- and Megachiroptera with any other modern order. In
contrast, two unique apomorphous states strongly support the monophyly of
bats. The first is the course of the ramus infraorhitalis. This artery enters the
cranial cavity in front of the mandibular nerve, runs forward along the
intracranial surface of the alisphenoid, and passes into the orbit with the
maxillary nerve via the confluent superior orbital fissure/foramen rotundum.
The second concerns the pattern of tympanic-roof closure. Along with
lagomorphs, rodents, and macroscelideans, bats have the tegmen tympani
ventral to the ramus superior/ramus inferior bifurcation and ramus inferior
course. However, bats are distinguished from the other forms by the unique
form of the tegmen tympani. In general, the mammalian tegmen tympani lies in
a horizontal plane lateral to the promontorium of the petrosal arid forms a roof
for the tympanic cavity. In chiropterans, however, the tegmen tympani is a
slender, more vertically-orientated rod that projects ventrally into the tympanic
cavity (Fig. 5B) and forms in continuity with the cartilage of the auditory tube
(Wible, 1984).
Bats have been placed in the superorder Archonta along with dermopterans,
scandentians, and primates (McKenna, 1975; Szalay, 1977; Novacek & Wyss,
1986a). Novacek (1982) supports a grouping of Chiroptera and Dermoptera
EU’I’HERIAN SlAPEDIAL ARTERY
129
(Fig. 6) with 12 synapomorphies. Only five bat-colugo synapomorphies are
specializations of the flight apparatus (and therefore possible convergences,
according to Pettigrew), and included in these five are the humeropatagialis and
coracocutaneous muscles, unique elements not found in other gliding mammals.
Because a bat-colugo clade seems reasonably well established (see also Novacek,
1986), the apomorphous pattern of the ramus superior/ramus inferior
bifurcation and ramus inferior course that bats share with lagomorphs, rodents
and macroscelideans apparently has been independently acquired. This is
supported by the unique morphology of the chiropteran tegmeri tympani
described above, which differs considerably from the horizontally-orientated
element of lagomorphs, rodents, macroscelideans and most other mammals. Bats
and colugos do not share any apomorphous features of the stapedial system or
tympanic roof: in fact, the tegmen tympani is absent in colugos (Halbsguth,
1973) and the reduced ramus inferior and lesser petrosal nerve run ventral to
the membranous tympanic roof (personal observation).
Ungulata
The ungulate radiation
including the modern orders Tubulidentata,
Artiodactyla, Cetacea, Perissodactyla, Hyracoidea, Proboscidea and Sirenia,
is generally (e.g. Van Valen, 1971, 1978;
and a number of extinct orders
McKenna, 1975; Szalay, 1977; Novacek, 1982) held to be a monophyletic
assemblage originating in or near the Arctocyonidae, one of several Late
Cretaceous and Early Tertiary groups that are informally termed ‘condylarths’.
Special relationships between certain condylarth families and modern orders
have been demonstrated and generally accepted (for example, mesonychid
condylarths and cetaceans; Van Valen, 1966, 1968), but the origins and
interrelationships of most modern orders within Ungulata are the subject of
considerable debate.
Table 2 reveals that modern ungulates share a number of apomorphous states
of the stapedial artery (losses and de nouo anastomotic linkages). T h e
superordinal relationships suggested by this character distribution are depicted
in Figure 7. Of particular interest to the interrelationships within Ungulata are
the following:
~
( 1 ) Tubulidentates are distinguished from the remaining modern orders by
the retention of the stem of the ramus superior and a distinct foramen for that
vessel in the suture between the petrosal (tegmen tympani) and squamosal. In
fact, a foramen for the ramus superior distinguishes tubulidentates from all
extinct ungulates except some Palaeocene condylarths (i.e. Arctogon,
Arctocyonidev, and Pleuraspidotherium; Russell, 1964), and apparently in these forms
the foramen is entirely within the tegmen tympani. (Thewissen (1985) reports
that a fissure in the petrosquamous suture of two Middle Eocene artiodactyls
may have transmitted a ramus superior, but such openings are absent from
other Eocene artiodactyls, including the Early Eocene Diacodexis (Coombs &
Coombs, 1982).) Tubulidentata either had a unique origin within
Rrctocyonidae or, as has been suggested recently by Novacek (1982, 1986) and
Thewissen (1985), an origin outside Ungulata.
I30
J. R . WIBLE
P
Figure 7. Phylogenetic relationships of ungulate ordrrs modifird from Novacrk (1982: fig. 1 ) i n light
of thr character analysis of the staprdial artcry. Nunibcrs r e h [(I thc following apomorphous
charactri- states listed in ’I‘aldc 2: I , proximal stapcdial artery absent. 3, ramus superior ahsrnt. 9.
ramus infraorbitalis enclosrd within an alisphrnoid canal. 1 1 ramus anastoinoticus pr(~scnt.
~
(2) Cetaceans, perissodactyls, hyracoids, and proboscideans are distinguished
from artiodactyls by the loss of the proximal stapedial artery. Although this
vessel is lacking in all modern representatives of these five orders, a distinct
groove for the proximal stapedial appears on the promontorium of the petrosal
in most condylarths (Russell, 1964; Cifelli, 1982) and in Eocene and Oligocene
artiodactyls (Dechascaux, 1974; Webb & ‘Taylor, 1980; Coombs & Coombs,
1982). The absence of this groove in early cetaceans (Kellogg, 1936; Gingerich,
Wells, Russell & Shah, 1983), perissodactyls (Radinsky, 1965; Savage et al.,
1965; Cifelli, 1982), hyracoids (Whitworth, 1954), and proboscideans (Tassy,
1981) represents a derived state within Ungulata.
( 3 ) Perissodactyls, hyracoids, and proboscideans are distinguished from
cetaceans by the addition of an alisphenoid canal for the infraorbital ramus and
a ramus anastomoticus. The homologies of the alisphenoid canal of
perissodactyls (Equus, Muggia, 1931) and hyracoids (Procavia, Lindahl, 1948)
are supported by the ontogenetic formation of a canal within the cartilaginous
ala temporalis; the development in proboscideans is not known. A grouping of
pcrissodactyls, hyracoids, and proboscideans (along with Sirenia and two
extinct groups, Desrnostylia and Embrithopoda) has been proposed elsewhere
(Van Valcn, 1971, 1978), but the usefulness of the alisphenoid canal as a
synapomorphy for this clade is uncertain. An alisphenoid canal appears in some
condylarths including mesotiychids (Szalay & Gould, 1966), the probable sister
group ol’Cetacea (Van Valen, 1966, 1968), and may have been added and lost
several times within Ungulata. No other features of the stapedial system further
distinguish relationships among perissodactyls, hyracoids, and proboscideans.
ACKNOWLEDGEMENTS
I thank the following individuals and institutions for access to seriallysectioned specimens: Prof. D. Starck and Prof. W. Maier (Goethe-Univcrsitat,
Frankfurt am Main); Prof: H.-J. Kuhn (August-Universitat, G6ttingen); Prof.
W. Reinbach (Karl-Universitat, Heidelberg); Dr H. E. Evans (Cornell
EU'IHERIAN STAPEDIAL ARTERY
131
University, Ithaca); Drs M. Cartmill, R. D. E. MacPhee, E. L. Effmann, and
K. L. Duke (Duke University, Durham); Dr R. Presley (University College,
Cardiff); and Dr E. C. Boterenbrood (Hubrecht Laboratory, Utrecht). For
access to skulls of extinct and extant mammals, I thank Drs R . H. Tedford,
M. C. McKenna, and G. G. Musser (American Museum of Natural History, Ncw
York); Drs B. D. Patterson and R . M. Timm (Field Museum of Natural
History, Chicago); and Dr Z. Kielan-Jaworowska (Zaklad Paleobiologii,
Warsaw). Numerous individuals at various institutions have contributed in
significant ways towards the completion of this work. In particular, I a m
grateful to M. Cartmill and R. D. E. MacPhee for encouragement in the early
phases of'this study and to E. Allin, *J. A. Hopson, M. J. Novacek, J. M. Clark,
M. Diamond, and C. Vanderslice for comments and improvements on the
manuscript. I also thank M . Diamond for unpublished observations of a ramus
temporalis in some lemuriform primates. This work was supported in part by
National Science Foundation grant DEB-80 1 1634 for Doctoral Dissertation
Research in Systematic Biology, National Science Foundation grant DEB8208797 to M . Cartmill and R. D. E. MacPhee, the Duke University Graduate
School, and the German Academic Exchange Service (D.A.A.D.).
REFERENCES
ALBERCH, P., 1985. Problems with the interpretation of developmental sequences. Systematic z o o l ~ 34:
~ ~ ,
46-58.
ALBRECHT, P. W . , 1976. The cranial arteries of turtles and thcir evolutionary significance. Journal of
Morpholou, 149: 159.- 182.
ARCHER, M., 1976. T h e hasicranial region of marsupicarnivores (Marsupialia), interrelationships of
carnivorous marsupials, and affinities of the insectivorous peramelids. <oological Journal ?/the Linnean Surieg,
59 217-322.
ARCHIBALD, J. D., 1977. Ectotympanic bone and internal carotid circulation of eutherians in refercncr to
anthropoid origins. Journal of Human Euolution, 6 609-622.
ARCHIBALD, J. D., 1979. Oldest known euthcrian stapes and a marsupial pctrosal honc from the Late
Cretaceous of North America. Nature, London, 281: 669-670.
BARGHUSEN, H. R., 1968. T h e lower jaw of cynodonts (Krptilia, Therapsida) and the evolutionary origin
of mammal-like adductor jaw musrulaturc. Postilla, 116 1-49.
BHATIA, M. L. L? DAYAL, J., 1933. O n the arterial system of the lizard Hemidac~lus~apauiuirzdis
Kuppel (the
wall lizard). Analornzscher Anzeipr, 76: 4 17-437.
BISHOP, M. J,, 1982. Criteria for thc dctcrmination of thc dircction of charactcr state changes. zoological
Journal ofthe Linnean Socieg, 74: 197-206.
BOCK, W. J., 1977. Foundations and methods of cvolutionary rlassilication. I n M. K. Hecht, P. C. Goody 8:
B. M. Hecht (Eds), Major Paltern3 in Vertebrate Evolulton: 851-895. New York: Plrnum Press.
BRUNER, H. L., 1907. O n the cephalic veins and sinuses of reptiles, with description of a mechanism for
raising the venous blood-pressure in the head. American Journal of Anatomt, 7: 1-1 17.
BUCHANAN, G. D. L? ARATA, A. A,, 1969. Cranial vasculature of a neotropical fruit-eating Iiat, i1rtibeu.r
lituratw. Analomischer Anreiger, 124: 3 14-325.
BUGGE, J., 1971. 'I'he cephalic arterial systrm in mole-rats (Spalacidae), hamhoo rats (Rhizomyidac:),
jumping mice and jerboas (Dipodoidea) and dormice (Gliroidra) with spceial reference to the systematic
classification of rodcnts. Acla anatomica, 79: 165- 180.
BCGGE, J., 1972. The cephalic arterial system in the insectivores and the primates with special refrrerice to
the Macroscelidoidca and Tupaioidea and the insectivore-primate boundary. zeitschrifl Jur Anatornie und
Entwicklunp.s~eschichte,135 279-300.
BUGGE, J., 1974. The cephalic arterial system in insectivores, primates, rodents and lagomorphs, with special
reference to the systematic classification. Acts anatornica, 87, supplemenl 62 1-160.
BUGGE, J., 1979. Cephalic arterial pattern in New World edentates and Old World pangolins with special
rekrence to their phylogcnetic relationships arid taxonomy. A c l ~anatomica, 105 37-46.
19:
BUTLER, H., 1983. 'Ihe embryology of the lesser galago (Galago senegalensis). Contributions to Primatol(~g~,
1-156.
BIJTLER, P. M., 1948. On the evolution (if the skull and teeth in the Erinaceidae, with special reference to
fossil material in the British Museum. Proceedings Oftlie <oologiral Socieg of London, 1948 446-500.
132
J . K. W1BI.E
BL'I'LER, 1'. .M., 1956. I'hr skull of IclopJ arid the classiiicatiorr ol' tlrr Inscctivora. Procredirtgs o/ thr <iiologd
Sociely oJ London, 1956: 453 48 1 .
CAK'I'LMll~L,
M. & MACPHEE, K . L). E., 1980. 'I'upaiid affinities: 'I'hc evidence of thr cat-otid artrrics aud
cranial skrlcton. I n W. 1'. Luckrtt (Ed.), Comnpcrrcitiz~rBioloyy a7rd Euolutionary Krlatiun.hi/J.\ o/ 7 r u r Shrrw.t:
95-132. New York: Plenum Prrss.
G I FELLI, R . L.,1982. 'l'hc pctrosal structure of f!ynp.sodu.r with resprct to itiat of some o t h c r ungul;itcs, and
its phylogenetic itnplicat ions. Journal of l'aleontolo<y, 56': 7 9 5 ~ ~ 8 0 5 .
C. I
&.CKWX4BS, W. P., Jr., 1982. Anatomy of the car rcgitm of four Eoccnc ;irtiodactylr:
COOMBS, ,&
f'hhin/yt.r, ?Helnhyu.s, Iliacodexis and Homnrodon.j'ournal nf C'ertebrate l'aleonlolugv, 2 2 19 236.
COPE, t.;. D., 1880. On the roramina perforating the posterinr part of the squamosal boric of ttrc M;immalia.
Proceedings of the American PhiloJophical SocieQ, 1 8 452--46 1.
CROMPT'ON, A. LV. & SUN, A.-L., 1985. Ci-anial SIructurr and relationships o f the I.iassic- mammal
.Sirtocarrcicli~rr.~iio/o,qirnlJuurnal qf the Linnenn Sucie(y, 85 YY- 1 19.
DE BIIEK, G . K.,1926. Studirs o n thc vertebrate head. 11. 'l'hc orlritotrniporal region of the skull. Qr~urtt7/rj
ompicol .Suiit?v, 70: 263-370.
37. The I)~arliipmrn/of lhr C'utrhrate .Skull. Oxford: Clarcndon I'ress.
, 1974. Artiodactylcs
.
.primitits des I'hosphorites c l u Qucrcy.
. ,~Innnlr.s de I'alPontolo,~ie
( Vertibris), 60: 5% 100.
I:NC.EI.MANN, G. E'., 1978. 7 h r lo,gir nfplylogenetic ana!yJiJ and /hr phvloger!,' n f t h u Xcnarthra (dlarnmtilia). Ph.1).
Dissertation, Columbia L'nivcrsity
FLEISCHER, G , , 1978. Evolutionary principles or thc marnnialian middlc car. AduaricrJ in ,lnaton!y, Emb~vnlog~
and Cull Biologr, 55 1-69.
FOURIE, S., 1974. The cranial morphol(igy of Thrinaxodon liorhinu.\ Serlcy. :Innal,s of'/hr .South African .\IIL.WVJ,
I : 337-400.
HS, l i . , 1905. Zur Entwickelungsgrschic.hte des \I'irbclticraugcs. I . I!cl)cr die l<ntwickc.lung tlcr
Augeng&issr des Kaninchens. AncitomiJi
GINCEKICH, 1'. I)., WELLS, N. A , , R
L, U. E. & SHAH, S. M . I . , 1983. Origin of wli;ilcs i i i
rpicontincntal rrmnant seas: New evidence &om the Early Eoccnc of Pakistan. Scimre, 220: 403 406.
(;00L)RICH, E. S., 1915. T h e chorda tympani and middlr car in reptiles, birds, and m;trnni;ils. Quarterly
Journal u/'the Mirrosropiral .Socir!y, 61: 137-160.
GOODKICH, E. S., 1930. .Studie~on /hr Slructurc and llrvelopmrnt u/ l'ertebra/e.c. London: Macmillan.
GREGORY, W.K,, 1910. 'l'hc orders ofmamm;ils. Bulletin ofthe American .Wirsettm clJ.Vhtural Hislo
GROSSER, O., 1901. ZuI Anatomic und Entwickelungsgeseliichte des Gefisssystems der
.,lnatomisrhu Hyflu, 17: 203 424.
HALBS(;U'J'H, ,A, 1973. Das Craniim rinus Fiirten df.1 I;ln/termaki Cynocephalus volaiis (Galcopitherus volans)
Dernro/itera) oon 63 m m SchStly. Iriaugural Dissertation (Medizin) Johann Wolfgang Gocthc-
, 1916. Zur Entwicklungsgescliiclitr der Aortenbogen und der Koprartrrien von 'Tarsitis
enbtiim hftir$ro/ogiJches jJahrbud, 50: 19-48,
1933. Das Artcricnsystem. In L. Bolk, E. Goppert, E. Kallius & W. Lubosch (Eds), Handbuch
der z~ty/rIr/rundrtr;lna/onrie der Ct'irheltiere, 6: 563 684. Re-impression 1967; Amsterdam: Ascher).
HOI,I.INSHEAU, W.H., 1968. AnalonyJor Su~<ron.s: Volume I . The Head and Neck, 2nd edition. New York:
Harprr ?i Row.
HOPSON, J . A. & BARGHUSEN, H . K . , 1986. An analysis of therapsid relationships. I n N. Hotton 111,
o t l r Pr E. C. Roth (Eds), The Ecolu~uorid Biology qf Mammal-like Rrptiles: 83 106.
hsoniarr Institution Press.
zur vergleichendcn Arrgiologir. I\'. Ilas arterielle GcFiss-system der Monntrenien.
Dtnk.schr$en ilkadrmip der L2'i.ssensrhqftm, M'ien, ma~hemali~ch-natrrne!i.s.senschaftliche
Klasse, 5 1-20.
HYRI'L, J , , 1854. BeitrBgc zur verglcichenden Angiologie. V. Uas arterielle Gefass-system der Edentaten.
DrnkJrhrzftrn .~lkndernirdrr Il'issen.rrhaften, Il'ien, niathemn/isch-naturu~issenschaftliche
Klasse, 6 2 1-65,
KADASI, K. M., 1973. 'I'he clrvelopinrnt of the skull in tlrc Iiidiari gerbil, 7hlera indicn cuzieri (Waterhouse).
Part I I , Gesqmhaurs .\lorpho/iisqisches j'ahrhurh, 119: 47-7 1,
KALLEN, E'. C:., 1977. 'I'hr cardiovascular system (if bats: Structure and function. I n U'.A. LVinisatt (Ed.),
B i n l q y u/ BntJ, I'd. 3: 28!1 ~483.New York: Academic Prrss.
KkLLOGG, K.,1936. A review of Archaeoeeri. Carnegie Institute o / Ct'ashingtan Publirahn 482 1-366.
KERMACK, K. A,, 1963. 'l'he cranial structure of the triconodonts. I'hilosophica/ 7mn.iactzuns q f / h r Royal Socidy
of I,ondun, B246: 83 103.
K E R M . ~ K I(.
, A., mssE , F. & RIGNEY, H . W., 1981. T h e skull structurr of Morzanncodon. Ziiological
Journal uJthe Linnrnn .Sucie&, 71: 1-158.
KIELAN-JA~1.'(~)KO\.\'SKA,%., I97 I , Skull structurr and affinitics o f the Multituberculata. In Z. KiclanJaworowska (Ed.), Rrsult.~o/. thr Poli.sh-.~~lon~~olian
Palaeontoloyical ExpeditionJ-Part 111. Palaeontologia Polonicci,
2.5: 5 4 1.
N-JA\.VOKOWSKA. Z.,1981. Evolution d t h e thcri;in mammals in the Late Cretaceous dAsi;I. Part
Skull siructurr i n hennalestes and A , i o ~ v c t e s . 111 %. Kielan-J;iw~irowska (Ed.), Results U j thr Polzsh,\lun,yolian I'alnrun/ulu,qirol &pedi/ions-Part I X . Palaeonlolo~~ia
P d 0 7 1 7 ~4, 2 25-78.
EG'IHEKIAN SIAPEDIAL ARTERY
133
KIELAN-JAWOROWSKA, Z. & 'I'KOFIMOV, B. A,, 1980. Cranial morphology of the Cretaceous
eutherian mammal Harun1este.c. Acta Palaeontolo~~ica
Polonica, 25 167-185.
KIELAN-JAU'OROWSKA, Z., PRESLEY, R. & POPLIN, C., 1986. T h e cranial vascular system in
taeniolabidoid multituberculates. Philosophical Transactions of the Royal Sociely, B313: 525-602.
KUHN, H.-J., 197 I . Dic Entwirklung uiid Morphologie drs Schadels voti Tac/yglossus aculeatus. Abhandlungen
der senckrnbergbchen naturjimchrndm t;esell.rchuft, 528 1 -224.
LI, C.-K. & TING, S.-Y.> 1985. Possible phylogenctir relationships of Asiatic eurymylids and rodcnts, with
comments on m i m ~ o n i d s .I n W. 1'. 1,utkett & J.-L. Hartenberger (Eds), Euolutionary Rrlationshipx a m o q
Kodent.r: A Multidisciplinary Approach: 35-58. New York: Plenum Press.
LINDAHL, P. E., 1948. c h e r die Entwicklung urid Morphologie des Chondrocraniums von Prncavia capenst.7
Pall. Acta <oologica, Stockholm, 29 281-376.
LINDAHL, P. E. & LUNDBERG, M., 1946. On the arterics in the head of Prorczuia capensis Pall arid their
development. Acta <oologira, Stockholm, 27 1-53.
LC'CKETI', W. P., 1977. Ontogeny of amniote fetal membranes and their application to phylogeny. In
M. K. Hcrht, P. C. Goody & B. M. Hcrht (Eds), Major Patterns in Vertebrate Euolution: 439-516. New York:
Plenum Press.
LUCKETT, W. P., 1985. Supcrordinal and intraordinal affinities of rodents: Developmental evidence from
the dentition and plarentation. I n W. P. Luckett & J.-L. Hartenberger (Eds), Evolutionury Relationsh$s among
Rodents: A Multidisciplinary Approach: 2 1 1-276. New York: Plenum Press.
MACPHEE, R. D. J i . , 1977. Auditory regions of strepsirhinr primates, tree shrews, elephant shrews, and l$o[yphliius
insectzuoreJ: An ontogenetic perspectiue on character analysis. Ph.D. dissertation, University of Alberta.
MACPHEE, K. D. E., 1981. Auditory regions of primates and eutherian insectivores: Morphology, ontogeny,
and character analysis. Contributions to Primatology, 18 1-282.
MACPHEE, R. 1). E. & CARTMILL, M., 1986. Basicranial structures and primate systematics. In D. R.
Swindler & J. Erwin (Eds), Comparatiue Primate Biology, Vol. I . Qstematics, Evolution, and Anatomy: 219-275.
New York: Alan R. Liss.
bl, DOWELL, S. B., Jr., 1958. The Greater Antillcan insectivores. Bulletin of thr American Museum qf Natural
History, 11.5 1 13-2 14.
M ~ I I O W E L L S.
, B., Jr., 1961. O n the major arterial canals in the ear-region of testudinoid turtles and the
classification of thc 'l'estudinoidea. Bulletin nf the MuJeum of Comparaliue .Zoology, 125: 23-39.
14<KENNA,M . C., 1963. New evidence against the tupaioid affinities of the mammalian family Anagalidae.
American Museum Jouitates, 2/58: 1-16.
McKENNA, M .C., 1975. 'l'oward a phylogenetir rlassification of the Mammalia. In W. 1'. Luckett & E'. S.
Szalay (Eds), P/y/oseny of the Primates: A Multidisciplinary Approach: 21 -46. New York: Plenum Press.
MUGGIA, G., 1931. Der Knorpelsrhadel eines Pferdeembryos. <eitschrifl f u r Anatomie iind
Entwicklung~~eschichte,
95: 297-325.
NOVACEK, M. J , , 1980. Cranioskeletal features in tupaiids and selected Eutheria as phylogenetic evidence.
In W.P. Luckett (Ed.), Conipnratiue Biology and Euolutionary Relationsh$s of Tree Shrews: 35-93. New York:
Plenum Press.
NOVACEK, M . J . , 1982. Information for molecular studies from anatomical and fossil cvidericc 011 higher
euthcrian phylogeny. In M. Goodman (Ed.), hincromolecular Sequences in Systematic and Evolutionary Biology:
2-41. New York: Plenum Press.
NOVACEK, M. J , , 1985. Cranial evidcncr for rodent affinities. In W.P. Luckett & J.-L. Hartenberger (Eds),
Evolutionary Relationships among Rodents: A Multidisr$linary Annlysis: 59-81, New York: Plenum Press.
NOVACEK, hl. J , , 1986. The skull of Ieptictid insertivorans and thc higher-level classification of rutherian
mammals. Bulletin of'the Amerimn Museum of Naturai History, 183 1-1 12.
NOVACEK, M. J . & WYSS, A,, 1986a. Higher-level relationships of the recent eutherian orders:
morphotogical evidence. Cladistics, 2: 257-287.
NOVACEK, M. J . & WYSS, A,, 1986b. Origin and transformation of the mammalian stapes. Contributions to
Geology, University of LVyoming, Special Paper, 3: 35-53.
OELRICH, T. M., 1956. 'I'he anatomy of the head of Qenosaura pectinata (Iguanidae). Miscellaneous Publications
ofthe Museum o f < o o l o ~ ~liniiiersily
,
of Michigan, 94: 1-122.
PAC:KER, D. J., 1983. Physiological noise, rarotid arteries, and auditory sensitiuily in mammals. 1'h.D. Dissertation,
New York University.
PADGET, D. H., 1948. 'I'hc dcvelopment of the cranial arteries in the human embryo. Contributions to
Embryology, Carnegie InJtitution, 32 205-261.
I'AKKER, W. K., 1885. On the structure and development of the skull in the Mamnialia. Part 3. Insectivora.
Philosophical 7ransar.tion.r ofthe Royal SocieQ, BI 76: 121-275.
PXl'TERSON, B., 1975. 'l'hc I'ossil aardvarks (Mamrnaliii: Tubulidentata). Bulletin of the Museum of
Comparatioe ~ o u l q y 147
,
185-237.
PE'ITIGREW, J . D., 1986. Flying primates? Megabats liavc the advanced pathway from eye to midbrain.
Science, 231: 1304-1306.
IGREW, J. I). & COOPER, H. M., 1986. Aerial primates: Advanced visual pathways in megabats and
gliding lemurs. Sociecy Jjr Neuroscience Abstracts, 12: 1035.
RAIIINSKY, L. B., 1965. Evolution of the tapiroid skclcton from Heptndon to Tapirus. Bulletin o/the Museum of
Comparatiue .Zoo"lqgy, 134: 69-1 06.
134
J. K. WlBLE
KOLX, C;. H . , 1947. ‘l‘hccranial development of ceriain Ethiopian insectivores and its bearing on the mi~iual
affinities of the group. il(.tn <uologica, Stockholm, 28: 165-397.
F,I.L, D. E,, 1964. Les MammiEres pali.oci.nes d’1’:urope. Mkmoires du MnsPum .Vational d’IIi.itoir?
turellu, Paris, . h i r C, 1 3 1 -324.
SABAN, R.,1956. Les athities du genre Tupaia Katfrs 1821, d’aprps Ics caractprcs morphologiques de la ttte
OSSCIISC. Annales de Pulc‘ontologi~ ( Vertkhrls), 42: I fi!) 224.
SAV.4GE, I). E., RUSSELL, D. E. & LOUIS, P., 1965. European Eocene Equidac (l’criss(rtlac-tylir).
C’nizjersily oJ CulzJurnia Puhlications in Geololical Sriencer, 56‘: 1-94.
SEGALI., W.,1970. Morphological parallelisms of the bulla and auditory ossicles in soinr insectivores and
marsupials. 1:ieldiana: ~ o o l o s y 51:
, 169-205.
SHIINO, ti., 1914. Studien zur Kcnntnis des Wirbeltierkopfcs. I. Das Chondrocranium von Crocodih mit
Heriicksiehtigung der Gehirnnervcri urid der Kop[qefasse. Anatomische Hefle, 50: 253-382.
SHINDC), ’I.,19 14. Zur vergleichenden Anatornie der artcrirllen Kop[qefasse der Reptilien. Anatumische Hub?,
50: 267-356.
SHINDO, T., 1915. (:her die Bedcutung des Siniis cavernosns der Siiuxcr mit vcralcii.hend-atiatomisrher
Bcrucksirhtigurig andere Kopfvenen. Anatomische HPftu, 52 3 1 9 ,495.
SICHER, H., 1913. Die Kntwirklungsgesrhirhte der Kopfarterien viin 7 d p a europaea. GegenbaurJ
Morphologisches Jahrbuch, 44; 465-487.
SIMI’SON, G . G . , 1937. Skull structure of the Multituherculata. Bulletin of the American Museum u/ ..l’a/urul
Hi,rto!v, 7.1: 727-763.
SIMI’SON, G. G., 1945. The principles of classiliration and a classification of mammals. Bulletin g f l h u Arnericati
AtfuseurnOJNaturul Histoyy, 8.9 1-350.
SISSON, S.,1914. The Anatomy ( ! / / h e Doniustic Animals, 2nd edition. Philadelphia: Saurrders.
SI,OAN, R. E., 1979. Multituberculata. I n K. W. Fairhridge & D . Jablonski (Eds), Encyclopedia of Eaith
IICPS, V11: hi:i2cyclopediuo J ~ P a l e o n t o / o ~ ~ y : 4 9Stroudsberg,
2~~8.
Pennsylvania: Dowdcn, Hutchinson & R(iss, Inc.
H , J . I). & MADKOUR, G., 1980. l’enial morphology and the qucstion olchiropteran phylogeny. I n
D. E. Wilson & A. L. Gardner (Eds), Proceedings o f t h e FiJlh Bat International Research Conference: 347-365.
Lubbock, Texas ‘Tech Press.
SI’AI‘Z, W., 1964. &ti-ag m r Kenntnis der Ontogrnese dcs C:rariittm win 7upaia g l i ~(Diard 1820). Gegenbaurs
Morphulo,<ische Jahrbuch, 106 32 I 416.
S‘I‘EIJERWALD, E. A,, 1969. Reuierei sf’ thr phylogerretic position of the trep shreu) (‘l‘upaia glis Diard), wi/h N P Z I ’
obserualiim 071 the arteria corotis interna. Ph.D. Dissertation, Michigan State University.
STRUTHERS, 1’. H., 1930. ‘Ihe aortir arches and their derivatives in the embryo porcupine !Ere/hiiorr
di~rsaluaj.~7nurnalof Morphology, 50: 36 1-392.
SZALAY, F. S . , 1972. Cranial morphology of the Early ‘l’ertiary Phrnarolrmur and its bearing on primate
phylogeny. .‘lmerican Journal oJ.Physical Anthropolu~g,36 59-76.
SZALAY, F. S . , 1975. Phylogeny of primate higher taxa: T h e hasicranial evidence. I n W. P. Luckett & F. S.
Szalay (Eds), Plylo,qeny q J & Primates: A Multidi.iciplinacy Approach: 91-125. New York: Plcnum Press.
SZALAY, F. S., 1977. I’hylogcrietic relationships and a classification of eutherian Mammalia. 111 M . K .
Hecht, P. C. Goody & B. M . Heeht (Edsj, Major Pat/urn.i in Vdehrate Euolution: 315-372. New York:
Plenum Prcw
SZALAY, F. S. & G O L J I J , S . J . , 1966. Asiatic Mesonychidae (Mammalia, Condylarthra). Hullvtin qf the
Anteriran Museum uJXatura1 Hisloly, 132: 127- 174.
TANDLER, J ., 1899. %ur vrrgleichenden Anatomic der Koplhrtericri Iici d e n Maniniali;i. Drnk.\chrifirn
Akademie der U’issenschaJen, W i r n , mathemati.ich-naturu~i.r.ren.tchaftliche
Klasse, 67 67 7-784.
‘I‘ANDLER, .J., 1901. Zur vergleichenden Anatomic dcr Kopfartericn hei den Mammalia. AnatomiJrhe Hejtu,
IH: 327-368.
‘IAXIILER, J., 1902. Zur E;rrtwicklungsgeschicte der Kopfartericn bei den Mammalia. Ge,qrnhaur.\
Morpholugisrhus jahrhuch, 30 2 75-3 7 3.
TASSY, l’., 1981. Le c r i n e dc Moeritherium (Proboscidca, Marnrnalia) de I’lhcPne de Dor el Talha (Lihyej et
IP probleme de la classification phylogkri6tique dii genre dans les ‘Iethytheria McKenna, 1975. Bulletin du
tional &Histoire Naturellt, Paris, 4 sc‘rie, .\ectiun C, 3: H7- 147.
, .I. G. M., 1985. Cephalic evidence lor the dfinities of ‘I‘ut~irlideritatii.Mamnralia, 49: 257-~284.
VAN DER KLAAUW, C..J., 1931. The auditory bulla in some fossil mammals, with a gcneral introducrion to
this region of the skull. Bulletin o f t h e American Museuni of.Nahmzl H i s / o y ~62
, 1-352.
VAN KAMPEN, 1’. N . , 1905. Die ’l‘ynipanalgrgend des Saugetierschadels. G‘exenbaurs Murpholqqisches j‘uhr/iuch,
34: 321-722.
VAN VALEN, I>., 1966. Deltatheridia, a new order of mammals. Bulletin oJ the American Museum UJ ,Vaturn1
History, 132: 1-126.
VAN VALEN, L., 1968. Morrophyly or diphyly in the origin of whales. Ei~ohtiun,22: 37 41,
VAN VALEN, I.., 1971. Adaptive zones and the i r d t m of mammals. Evolution, 25:420-428.
VAN VALEN, I.., 1978. The brginning of the age of mammals. Ei~olutiona:~Thuog~,4 45-80.
VON HOFMANN, L., 1914. Die Entwicklung der Kopfartericn hei SUJ ,mufa d(imusticus. Gegenbaurs
Murphulogi.trhes Jahrhuch, 48 645-67 I .
W A H L E R I , .]. H., 1974. T h e cranial foramina of protrogomorphous rodents: An anatomical and
phylogenrrir study. Bullutrn 0 $ 6 h Museum oJ Comparatiue <oology, 146: 3 6 3 4 1 0.
EUTHERIAN S'I'APEDIAL ARTERY
135
WA'I'SON, D. M. S., 19 16. T h e monotreme skull: A contribution to mammalian morphogencsis. Philo.rophir:al
Tiansactiom of the Royal Society of London, Series B, 207 31 1L374.
U'AI'SON, M., 1875. Contributions to the anatomy of the Indian elephant, Part IV. hlusclcs and hloodvessels of the face and head. Journal CJJ Anatomy, 9: 118-133.
WEBB, S. D. & TAYLOR, B. E., 1980. T h e phylogeny of hornless ruminants and a description of the cranium
of Archaeomeryx. Bulletin ofthe American Museum of Natural History,167 1 17-158.
WHITWORTH, T,, 1954. The Miocene hyraroids of East Africa. British Museum (~Vatural Hislory) Fossil
Mammals ofAfrica, 7 1-58.
WIBLE, J. R., 1984. T h e ontopny and phylogeny g t h e mammalian cranial arlerial pattern. Ph.D. Dissertation, Duke
Univcrsity.
YOUSSEF, E. H., 1966. The chondrocranium of the albino rat. Actn anatomica, 64 586-617.
ZELLER, U. A,, 1986. Ontogeny and cranial morphology of the tympanic region of the Tupaiidae, with
special reference to Ptilocercus. Folia prirnatologica, 47: 6 1-80.
LIST OF ABBREVIATIONS
A. anast.
a. d. mag.
acf
>Is
aud. bulla
bO
bs
cc
rca
cof
ct
cca
eo
ex tracran.
fi,
fin
lr
13.a
hf
ica
intracran.
i n t ratymp.
I11 v
lo
mand. n.
mas
max. a .
max. n.
mr
oa
arteria anastomotica
arteria diploetica magna
anterior carotid foramen
alisphenoid
auditory hulla
hasioccipi tal
basisphcnoid
cochlear capsule
common carotid artery
rranio-orhital foramen
cavum tympani
rxtrrnal carotid artrry
exoccipital
extracranial
fenestra cochlear (round
window)
foramen magnum
frontal
foramen for stapedial artery
hypoglossal foramen
internal carotid artery
intracranial
intratympanic
lateral head vein
lamina obturans
mandibular nerve
mastoid portion of petrosal
maxillary artery
maxillary nerve
meningeal ramus
occipital artery
of
orb. for.
oph. n.
0s
par
pcf
Pf
Pgf
plf
postgl. a.
P'
prox. stap. a.
r. anast.
r. I.-o.
r. inf.
r. mand.
r. post.
r. orb.
r. s.-o.
r. sup.
r. temp
rt
sma
sof
sq
temp. m.
tt
tYmP
V
VII
optic foramen
orbital foramen
ophthalmic nerve
orbitosphenoid
parietal
posterior carotid foramen
piriform fenestra
postglenoid foramen
posterior lacerate foramen
postglenoid artery
promontorium of petrosal
proximal stapedial artery
ramus anastomoticus
ramus infraorhitalis
ramus inferior
ramus mandibularis
ramus posterior
ramus orbitalis
ramus supraorhitalis
ramus superior
ramus temporalis
ramus temporalis
stylomastoid artery
superior orbital fissure
squamosal
temporalis muscle
tegmen tympani
tympanum
trigeminal nerve
facial nerve