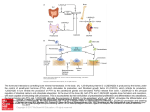
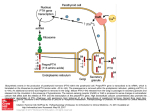
Survey
* Your assessment is very important for improving the workof artificial intelligence, which forms the content of this project
The American Society for Bone and Mineral Research 28th Annual Meeting Webcast Featured Basic Symposium: Osteoclast Signaling and Function Sunday, September 17, 2006 Co-Chairs: Steven L. Teitelbaum, M.D. Anna Teti, Ph.D. Speakers: F. Patrick Ross, Ph.D. Erwin F. Wagner , Ph.D. Laurie K. McCauley, D.D.S., Ph.D. 00:00:00 Anna Teti: Good morning ladies and gentleman. I welcome you to this "Featured Basic Symposium on Osteoclast Signaling and Function", and I am very pleased to co-chair the session with my friend and mentor and osteoclast expert, Steve Teitelbaum. Before we start, I have to make an announcement 00:00:23 just to remind you that the use of the cameras and audio taping devices is prohibited and also to tell you that you will receive an e-mail immediately after the meeting for the online meeting evaluation and we encourage you to fill the form. So as you know, the osteoclast field has recently made a 00:00:45 tremendous progress and our first speaker has largely contributed to the field of osteoclast signaling. Dr. Patrick Ross is Professor of Pathology, Immunology, Cell Biology, and Physiology at the Washington University in St. Louis with the main interest in the signaling induced by the alpha (v), 00:01:09 beta (3) integrin, and he would talk to you today about the mechanisms of M-CSF and alpha (v), beta (3) signaling in osteoclasts. Patrick? 00:01:37 F. Patrick Ross: So it's a great pleasure for me to be here and I thank the organizers for giving me the opportunity to talk about our recent work on M-CSF and the osteoclast cytoskeleton. 00:01:48 So, just very briefly to remind you how the osteoclast works. The osteoclast is a multinucleated polarized cell, which attaches to bone via the integrin alpha (v) beta (3) and that's a critical player that we are going to talk about again later today. 00:02:08 And having polarized, the cell then generates its unique organelle, the ruffled border, and from that ruffled border are secreted hydrochloric acid and proteolytic enzymes such as cathepsin K and MMPs, which together degrade the bone. So how osteoclast is made? 00:02:27 Well, we all know today that osteoclasts are cells of a myeloid lineage and that osteoclast precursors have on their surface two important receptors c-Fms and RANK, which are bound respectively by M-CSF and RANK ligand, and you will see that the M-CSF and RANK ligand can come from cells such as 00:02:47 mesenchymal cells or in other circumstances, some of them come from T cells, and that you can also see that there are two forms of M-CSF. There is a membrane bound form and there is a soluble form, and both of those forms are thought to contribute to osteoclastogenesis. 00:03:00 So the signaling is induced when M-CSF and RANK ligand bind Fms and RANK respectively is what gives rise to the osteoclast. But the osteoclast just because it's multinucleated doesn't mean that it's necessarily active. And it's a step that has to be taken in which the osteoclast is activated as 00:03:21 shown here and what you are seeing again is the cell adhering specifically by alpha (v) beta (3) forming the ruffled border and then degrading bone as I have talked about just a moment ago. And there are two important features that identify this resorptive cell. 00:03:39 The first one is the ruffled border and the second one is the actin cytoskeleton, and I draw your attention particularly to point number four, which represents an actively resolving osteoclast with this actin cytoskeleton organized in a unique fashion. So what organizes the actin cytoskeleton? 00:04:00 Well, a number of signals can do that, but as you can see here, one of the signals that does it potently is the integrin alpha (v) beta (3). And what you are looking at are osteoclasts generated on bone, in which at least one copy of the beta (3) integrin is present or you have the null mouse. 00:04:20 And you can see that in the beta (3) containing osteoclast, there is good organization of the actin ring whereas in the null cell, there is no organization of the actin ring comparably. So, another signal that also regulates the osteoclast cytoskeleton, and that's what I am going to spend a lot of my time talking about today, is M-CSF. 00:04:46 And people usually think as M-CSF is a cytokine that's involved in the proliferation and survival of osteoclast precursors in their regulation of differentiation and also for the inhibition of apoptosis of mature osteoclasts and their precursors, but as Karl and Sonia and Barone group and others 00:05:04 have shown M-CSF is an important regulator on the osteoclast cytoskeleton. And how does it do that? Well, actually the first insight is to how it did it, came from this seminal paper published 15 years ago now by Phil Soriano in which he showed that in the targeted disruption of c-Src, the major 00:05:26 phenotype was severe osteopetrosis and was not because the cells did not have osteoclasts, in fact they had more osteoclasts as Brendan Boyce subsequently showed. It was because the osteoclasts just didn't work. So, another recent development is a realization that another kinase often interacts in 00:05:49 a signaling modality with Src and that kinase are the other kinases of the Syk Zap-70 family. And what is Syk? Syk is Spleen Tyrosine Kinase and as you can see here from its domain structure, it's got an amino terminal or carboxy-terminal SH2 domains. 00:06:12 It's got a kinase domain of course and then it has a number of tyrosines scattered throughout the molecule, which are targets of kinases themselves. Importantly for this talk, these two SH2 domains acting in tandem bind an immunoreceptor tyrosine activation motif abbreviated to ITAM, which 00:06:34 consists of two tyrosines, which are phosphorylated and they are separated by somewhere between 8 and 10 amino acids and they have flanking sequences, which define them as well. So, we looked at the mice that regenerated, not when in which there was no Syk in their osteoclasts. 00:06:53 And as you can see here, the number of osteoclasts per unit surface is the same in the wildtype and the knockout mouse, and when you treat both sets of animals with PTH, the numbers go up, but again, there is no difference between the wild-type and the null animal. 00:07:13 On the other hand, if you look at resorptive capacity on the osteoclasts, and here you are looking our measure is serum CTX, which is the best index we think today of global bone resorption. You can see that the wild-type animals undergo a robust increase in CTX and this is blunted significantly in mice incidental lacking Syk. 00:07:36 Consistent with this, if you look at the histology of these mice, you can see in the wild-type animals, there it is well spread, big osteoclasts, and when you go to higher resolution, you can see ruffled borders and you can see many resorption lacunae scattered around the bone. 00:07:52 And again, in contrast, if you look at the knockouts and it's best seen here in this blowup of this area. Most of the osteoclasts are off bone suggesting that their cytoskeleton is abnormal and if you look closely, you can see very few resorption lacunae. 00:08:09 So, consistent with the in vivo data, we also just generated osteoclasts on dentin slices and we stained them for actin. And the wild-type animal or animal containing one copy of the gene, the actin cytoskeleton is quite normal as you can see over here, and the cells resorb, the dentin shown here with staining. 00:08:33 In contrast, the Syk-null osteoclasts don't organize their cytoskeleton and not surprisingly, they don't resolve the dentin. So those data both in vivo and in vitro led us to conclude that Syk is a key regulator of the osteoclast cytoskeleton. And I remind you again of the interaction and other circumstances between Src and Syk kinases. 00:08:58 And when we took this into account and we also took into account the fact we know that Src is activated by M-CSF, we were led to the question, does the Src-Syk signaling access mediate M-CSF regulation of the osteoclast cytoskeleton? So to do that, we undertook a series of signaling studies. 00:09:23 And the first thing we wanted to show was in fact did M-CSF activate Syk and you can see here this is a Western blot and you are going to see a number of these, so let me take you through this first one relatively slowly. What we did is we made osteoclasts, we removed all cytokines, and then, we 00:09:41 treated them for a short period of time either with vehicle or M-CSF, we did an immunoprecipitation for total Syk, and then we did a Western blot for phosphorylated Syk. And what you can see here clearly is that M-CSF activates Syk by phosphorylation, and consistent with the potential role for 00:09:59 c-Src in that process, if we pretreated the cells with a potent Src family kinase inhibitor, this signaling was totally abrogated. So, what we had established at this stage was in fact c-Fms activates Src, that's well established, but that Src in turn activates Syk. 00:10:19 So, which proteins downstream of Syk, maybe organizing the osteoclast cytoskeleton is the next question we asked. And the protein we focused on, based on largely the literature, is SLAP-76. SLAP-76, as you can see here, in a cartoon from Gary Koretzky's recent review, it's an adaptive protein that binds multiple partners. 00:10:44 And in particular, we wanted to know whether SLAP-76 and Syk were in the same pathway. So once again, we carried out signaling studies, and in this instance, you are looking at osteoclasts that had been generated, washed, treated with M-CSF. We did an immunoprecipitation for SLAP-76 and then we 00:11:05 did a phospho block for the phosphorylated form of the protein. As you can see here, in animals, which are wild type, i.e., they contain Syk, there is robust phosphorylation of SLAP76, but in the knockout animals, there is no phosphorylation, even though the protein is present in the same concentration. 00:11:24 So we've now established an extent of that pathway from Fms all the way through SLAP-76. What could be SLAP-76 binding that may regulate the osteoclast cytoskeleton? And as I pointed out to you, there are a whole host of proteins, but the only one I want you to focus your attention on today in fact is Vav, and why would that be? 00:11:49 Well, that's because last year, we published a paper in which we showed that Vav3 regulates osteoclast function and bone mass and in particular, the cytoskeleton of those Vav3 knockout mice was abnormal. So the obvious question was, does M-CSF activate Vav3, and is SLAP-76 upstream of that? 00:12:14 So again, we have another Western blot where we make osteoclasts, treated them or not with cytokine, immunoprecipitated here in separate experiments, Syk and then Vav3, and you can see that in the wild-type animals, M-CSF induces phosphorylation of Syk, which I have shown you previously, but it also produces phorylation of Vav3. 00:12:36 In contrast, in the SLAP-76 knockout mice, you continue to phosphorylate Syk indicating that Syk is indeed upstream of SLAP-76, but there is now no phosphorylation at all of Vav3. And so, that allowed us to extend the pathway yet again from Fms to Src to Syk to SLAP-76 to Vav3. 00:12:59 Well, what about Vav3 and its downstream effectors? So, this cartoon shows you that when receptor tyrosine kinases or integrins initiate signaling that leads to changes in the osteoclast cytoskeleton, they do it largely by activating a family of proteins called guanosine exchange factors. 00:13:22 And what they are doing is they are taking a small GTPase such as Rac, Rho or Cdc42, which is in its inactive form, GDP-bound, and to change from the GDP for GTP and that activates Rac, and then Rac can then organize the actin cytoskeleton. So to ask the question, does Rac play a role in the osteoclast cytoskeleton? 00:13:48 We generated Rac1, Rac2, double knockout mice. We bred these mice and as you can see here, both histologically and histomorphometrically, the double knockouts have considerably more bone mass. So, now the pathway goes all the way from Fms down to activated Rac. 00:14:08 I want to come back now to spleen tyrosine kinase and in particular, to its ITAM binding SH2 sequences. So, in many cases, the binding of Syk to an ITAM is a necessity for signaling through that pathway. So as Hiroshi Takayanagi and I apologize, I noted this misspelling on it after I put my slides in. 00:14:34 He has shown elegantly as we heard yesterday that DAP12 and Fc gamma receptor, which are two ITAM proteins, together mediate osteoclast differentiation signaling in response to RANK ligand. Well, I am going to show you these ITAM proteins also have a role in the osteoclast cytoskeleton in response to M-CSF. 00:14:57 So, the question or the hypothesis was that the ITAM is involved for M-CSF induced osteoclast cytoskeletal organization and you can see that in the cartoon here. Here, you have Fms and I've shown you all of these players. I've shown you Src, Syk, SLAP-76, Vav3, Rac, and the osteoclast cytoskeleton. 00:15:17 What I haven't shown you is whether or not this putative ITAM plays a role. I am going to focus on one ITAM and that is DAP12. And the question I am going to ask is does DAP12 play a role in regulation of the osteoclast cytoskeleton by M-CSF. And to do that, we took DAP12 knockout, osteoclast 00:15:37 precursors, made osteoclasts, washed them, treated them with M-CSF with time, immunoprecipitated DAP12, and did a Western blot for phosphotyrosine. And as you can see here, there is a time-dependent increase in phosphorylation of DAP12 in the wild-type cells and consistent again with the role of an 00:15:59 Scr, which you would have predicted from other studies, when we used an Scr inhibitor, we were able to totally block this signal. So having established that M-CSF does activate DAP12, we wanted to know does it also participate in the downstream signaling from MCSF? 00:16:17 So here we took wild-type osteoclasts, treated them with M-CSF, and you can see that again, with time, we got phosphorylation of Syk, and this is just the amount of DAP12 protein. Wherever when we take the DAP12 knockout mice, make osteoclasts and do the exactly the same experiment, there is no phosphorylation at all or Syk. 00:16:38 Furthermore, going downstream now, if you take wild-type mice and you treat them with MCSF, I've shown you this previously, but this is the time dependency. In this particular experiment, you activate Vav3, but in the DAP12 knockout mouse, you completely fail to do so. 00:16:56 And finally, and the most distal signal that we really have our hands on right now is Rac. And what you are looking at here is a Rac activation assay and what you do is you use a GST CRIB domain protein for a pull-down, and that GST CRIB domain recognizes only the activated form of Rac. 00:17:17 And then, you do total Western blot and you can see here that M-CSF activates Rac, but in the absence of DAP12, it completely fails to do so. So, we do now know that that question mark has gone away. And I want to finish up in a few minutes by just going back to beta (3). 00:17:35 And we pointed out to you that the beta (3) integrin is a major regulator of the osteoclast cytoskeleton. And the fact that we had to establish the signaling pathway that I have laid out for you, let us ask the question, is DAP12 essential for alpha (v) beta (3) induced Syk phosphorylation. 00:17:52 And the answer is, yes, and I am not going to show you any of these data, but this cartoon, which is very similar to the cartoon that you have seen for M-CSF shows that Src is constitutively on alpha (v) beta (3) even when it's activated, it phosphorylates Syk in a DAP12 dependent manner and then that activates DAP3 Rac and the cytoskeleton. 00:18:15 The only thing we haven't established at this moment for alpha (v) beta (3) is whether or not SLAP-76 is in the middle of it as well. So, if indeed M-CSF and alpha (v) beta (3) are using very similar pathways to activate the osteoclast cytoskeleton, one would predict that their signaling may in some way be additive. 00:18:39 And in fact, we published the paper three years ago now, in which we demonstrated exactly that fact, that c-Fms and alpha (v) beta (3) integrate their signaling during osteoclast differentiation and function. So our model now is as follows, that c-Fms and alpha (v) beta (3) by different 00:18:58 mechanisms activate c-Src, which phosphorylates DAP12, which recruits Syk, which phosphorylates SLAP-76, which recruits Vav3 and it is activated, which activates Rac, and that Rac in turn then converts nonpolarized osteoclasts to polarized osteoclasts. 00:19:15 I would like to thank our collaborators who have given us animals. David Williams, who gave us the Rac1, 2 knockout mice, Victor Tybulewicz, who gave us the Syk knockouts, Wojciech Swat who gave us Vav3. Gary Koretzky, who gave us the SLAP-76 knockouts, and finally, Marco Colonna, who gave us the DAP12 knockouts from Japan. 00:19:35 We need to acknowledge everybody who did the work. Most of the work you see here was done by Wei Zou, Sunao Takeshita, and Xu Feng have set up the retroviral system that allows us to transduce null cells with a great efficiency and they need great acknowledgment. 00:19:48 Roberta Faccio was involved in some of the studies, so was Hideki Kitaura. Jennifer Reeve is a new MD, Ph.D. in the lab looking at SLAP-76, and Jean Chappell is a senior person in the lab who does everything. And as all of you would know, this has been a long-term collaboration with my great friend and colleague, Steven Teitelbaum. Thank you very much. 00:20:15 Anna Teti: Thank you very much for this wonderful presentation and the discussion is open. Please introduce yourself. 00:20:28 Female Speaker: Patrick, that was a beautiful presentation. I was just wondering whether you looked at alternative ITAM containing domain proteins such as, you know, SIRP beta and also the (inaudible), the ITIMs? 00:20:43 F. Patrick Ross: Sorry I could...could you repeat the question please, Yanis? 00:20:45 Female Speaker: I was wondering whether you looked at alternative ITAM containing domains proteins such as sub-beta and also ITAM-containing protein? F. Patrick Ross: Yeah. Female Speaker: They can go into regular... 00:20:57 F. Patrick Ross: We don't know. We haven't looked at anything else. The obvious one we are in the middle of looking at is FcR gamma, and because we don't want to use a heterologous system, we are just going to...in fact, we've already got the virus ready to go. 00:21:09 We've put the tail of FcR gamma onto the rest of DAP12. We are going to see whether we can get that to signal. That's a minute experiment. 00:21:18 Dan Bickley: Bickley, San Francisco. Patty, that's a lovely presentation. We've gotten very interested in the interaction of the beta (3) interaction 00:21:27 with the IGF-1 receptor in the osteoblast and their direct connection, and I am wondering if the same thing is true in the osteoclast with respect to Fms and the alpha (v) beta (3). Do you have a direct interaction and if you eliminated beta (3), for example, would M-CSF work... 00:21:50 F. Patrick Ross: So... Dan Bickley: ...on your pathway? 00:21:52 F. Patrick Ross: Thank you Dan, an interesting question. If I get in Dan, the late Wong recently published, they found an interaction in cells between alpha (v) beta (3) and c-Fms, I have to tell you, we've tried very hard and we have put 00:22:09 in c-Fms with a flag tag on it, which enables us to pull it down with high affinity. We put in beta (3) with a human external domain. We have great antibodies to that and we have not just been able to find that association. I believe that it's there given the lays work, but we can't find it. 00:22:28 Dan Bickley: If you, in your beta (3) knockout, will M-CSF activate this pathway? That's...that's a key. 00:22:37 F. Patrick Ross: Absolutely and those experiments were ongoing as we left the lab. Anna Teti: Okay. Next. 00:22:43 Male Speaker 1: I am (inaudible) from Tokyo, Japan. What wasn't clear for me is interaction between the RANK ligand signaling and M-CSF signaling in terms of DAP12 pathway? So, do you have any comments on that? 00:22:59 F. Patrick Ross: Sure. So, I think as is the case in signaling biology in general, things are very complex and we like to packet them in individual little clusters because that's how we can figure things out. So I think as for many receptors, there are multiple interacting, signaling clusters, which are independent of one another. 00:23:21 So I think in some circumstances, DAP12 is mediating RANK signaling and it's doing that in a way which activates PLC-gamma and in the calcium transients and that's for differentiation and NFAT, and so on. And I think in other circumstances, there is a pool of DAP12, which is associated either 00:23:40 with the integrin or with Fms and in that circumstance, that DAP12 is regulating the osteoclast cytoskeleton. 00:23:47 Male Speaker 1: The other thing is your data clearly shows that the DAP12 exists...presence of DAP12 is important, but... F. Patrick Ross: Right. 00:23:54 Male Speaker 1: ...do you think that activation of DAP12, I mean the ligand-dependent activation of DAP12 is required for the signaling of the M-CSF? 00:24:04 F. Patrick Ross: So as you know there a number of ligands identified and there are probably some that have not been identified. All I can tell you is two things. Number one, if we mutate the tyrosine residues on the tail of DAP12 to phenylalanine, signaling goes away and we can do that in the 00:24:20 DAP12 null cells, which has got all of their accessory receptors that they need because they are primary microphages and just mutating Y to F kills the signals, all the signals, that's number one. And number two, we've done some experiments that I haven't talked about, where in heterologous system, 00:24:41 we wanted to look at interactions between domains of DAP12 and other proteins. And to do that, we co-expressed TREM2, one of the activators of DAP12, and in the presence of TREM2, we could get signals that we liked and were meaningful, and in the absence of TREM2, we didn't. 00:25:00 So that...and again, I don't like heterologous systems. As you may know, our lab works exclusively in primary cells and we don't overexpress genes either. So, with that reservation, I think the answer is yes. 00:25:12 Male Speaker 1: Thank you. Anna Teti: Okay, right. Next. 00:25:15 Roger Sabbadini: Thanks Patty for great overview. Would you just care to speculate on what the downstream effectors of Rac might be in osteoclasts? 00:25:24 F. Patrick Ross: Speculate on what...sorry? Roger Sabbadini: The downstream effectors of Rac might be? 00:25:27 F. Patrick Ross: Of Rac? Well, again, if you believe the literature, Racs activate members of the WASP/WAVE family and Racs would presumably actually activate waves and waves would in turn go downstream in a way that I think is not clearly defined to Arp2/3. 00:25:44 We've shown by the way, we had a paper in MBC, earlier this year that cortactin when you knock it down in the osteoclast, doesn't organize your cytoskeleton, and we were able to rescue that with wild-type cortactin, but if we put in the triple cortactin (ph), Y to F mutant, we did not rescue. 00:26:02 So, I am sure that cortactin is in that pathway not surprisingly. So I think WAVE, Arp2/3, cortactin watch forever. 00:26:11 Roger Sabbadini: Thanks. Anna Teti: Last question. 00:26:13 Male Speaker 2: (inaudible). I would like to ask you something about the Rho GTPase (Rho) and not Rac. It has been shown previously that Rho was essential for actin or organization of different level, and where do you put Rho in the pathway that you described now? 00:26:28 F. Patrick Ross: So the question is, where is Rho in this, and so, as you know very well that the family of those proteins, more GTPases that are cytoskeletal organizing on Rhos, Racs, and Cdc42s. And I think, and we have data that we haven't talked about was about Cdc42, it plays a role as well. 00:26:48 And yeah, there is a lot of data out there where people have blocked Rho with exonuclease and so on, and those data look convincing, but I think one of the issues which is confounding much of this is that Rho, Rac, and Cdc42 crosstalk to one another. 00:27:00 And I don't know whether that crosstalk changes the pattern of signaling downstream of all of them. I acknowledge that Rho plays a role, but I think we have the genetic data now that Rac 1 and 2 certainly are players in that field. 00:27:14 Anna Teti: Okay thank you, Patty. We will go to the next speaker. It will be presented by Dr. Teitelbaum. 00:27:27 Steven L. Teitelbaum: Good morning. It's my pleasure to introduce my buddy, Erwin Wagner, who is the Deputy Director of the Research Institute of Molecular Pathology in Vienna. 00:27:39 Erwin has made enormous contributions and varied contributions to understanding of the osteoclasts, and in particular, he has really defined the role of AP1 proteins in the osteoclastogenic process, and Erwin will talk to us about "Signaling of FOS Proteins in Osteoclasts." 00:27:58 Erwin F. Wagner: Yeah, many thanks for this nice introductory words. And obviously, I am delighted and honored to be here, particularly since I got my most important scientific training here in Philadelphia more than 25 yeas ago on the generation of genetically manipulated mice. 00:28:33 Now I am also delighted and very thankful to Dr. Andy Arnold who gave me a very strict briefing on how to do this presentation today. Numerous e-mails and also telephone conversations pointed to the direction that I really have to be very basic and in one of the emails, he even said that the 00:28:58 committee considers that it's especially important to have my talk tailored to the needs of a broad, general audience. Most of them, I saw it, have little background or no specific knowledge on the topic I talk about. So now, I mean the dilemma on one hand, do I start with bone and transcription 00:29:20 factors or do we have to start with the fertilized egg? Now, he also pointed out very clearly that the first 5 to 10 minutes have to be on the technology, how to generate and why to generate genetically manipulated mice. So, for those you who are in the audience and who really know these better than me, I apologize. 00:29:41 So, the first 5 to 7 minutes will be very basic and we will not talk about Fos proteins in osteoclasts, but we will talk about some general things on the technology. And then, I will come and illustrate to you how important these transcription factors are in determining the bone remodeling unit 00:30:00 and also the lineages, particularly in osteoclasts. So the mouse is a model organism, why is that so important? I think we all agree most of us in the field clearly that we need this to study the functional regulation of genes and to understand embryonic and organ development, but also if you 00:30:20 consider tumors as organs that you like to illustrate how important this is for tumor development. For this talk here, it's most important that we use this mice to study the pathogenesis of disease and possibly then apply these mouse models for pre-clinical studies to either prevent or cure diseases. 00:30:37 And the genetic modifications, which we introduce in the germline of mice, this is being done by either transgenic or knockout technologies. And so very...the next five minutes, very briefly, there are several methods to do this. For those of you who have never heard of it, there is microinjection of DNA into fertilized eggs. 00:30:56 So we start with the egg. We introduce the genes there. We can do it also by viruses or most importantly, and this is what you really have to remember, if you hear about genetically modified mice, knockout mice, this is all referred so that you do gene transfer into ES cells, chimera formation, 00:31:13 and then you end up with genetically modified mice. So now, to illustrate how this works, I'll give you a few examples. So the microinjection of DNA into the pronucleus of the fertilized eggs looks as follows. You see here, the fertilized egg, injection pipette, and here, you see the successful introduction into the pronucleus of the egg. 00:31:33 The frequency of getting these transgenic founders lies anywhere between 5% and 20% of the injected eggs, and the disadvantage of this technology is that you have random integration of the injected DNA or the injected genes. You cannot predict the outcome of these experiments, which is very 00:31:50 different, and all of us nowadays, mostly resume to the ES cell technology for generating specifically designed mutant mice. And here, the outstanding features of embryonic stem cells, you have the in vitro phase where the embryonic stem cells are being genetically altered by gene transfer. 00:32:10 And any mutation you wish to introduce, you can introduce and characterize at the level of the embryonic stem cells, whether this is a point mutation, whether this is a gene knockout, whether this is a conditional allele where the gene can be switched on or off in the time and cell type specific manner. 00:32:27 This you can or basically pre-select or pre-characterize at the level of the ES cells. And then, the magic technology emerged, how to convert this ES cells into ES mice, and this can be done either by blastocyst injections or you can compromise the embryos that had only the ES cells develop as a full ES mouse. 00:32:50 There are many new technologies emerging these days and this is then like you are going one step from the genetically modified cell to the mouse. Since maybe some of you have never looked through the microscope and seen how this works, I have taken out an old movie. 00:33:07 And while you will see this movie running, you should be able to see a holding pipette and an injection pipette, and then, you will see your favorite object of interest, this is the blastocyst and the ES cells. So there are single cells, suspensions of ES cells, which are being picked up here by the microinjection pipette. 00:33:25 And then, the skills of the experiment has to be quite high in order to now, as you will see, immobilize the blastocyst, which comprises about 80 cells to only the trophectoderm cells and the inner cell mass cells, which give rise to the embryo proper. You need to position this in the right way. 00:33:46 You see, this is sometimes a bit tricky, so that you can nicely then inject between these breaches of these trophectoderm cells. So now, we managed to see, this will be immobilized. The injection pipette will come right here between the trophectoderm cells and you pierce through here and then 00:34:03 you deliver the ES cells in front of the inner cell muscles in the range of 8 to 10 cells, little bit dependent on how good your ES cells are. Then, you withdraw the pipette and you have to assure yourself that the breaches are sealed here afterwards so that the cells don't go out again because then, you will not have a successful experiment. 00:34:27 And then, what you see here is that indeed, cells stay all within the zona, within the (inaudible), and you can see very nicely that the ES cells are part of this egg. Now, the next step is then to convert the ES cells into, sorry, it's on the right side. I just want to go further forward. 00:34:53 Press here? I did it. Now, we don't want to see this again for sure. No, this is backward. Okay, now here. Yes. So, if the experiment is successful, you will get a mouse being born. The donor strain is often a Black Six mouse and genetic background from the ES cell is...coat color brown. 00:35:16 Here, you can see the Agouti mouse here. If the experiment is very successful, the Agouti mouse or the ES mouse is, like you see here, this example is completely comprised of the color of the embryonic stem cells. This is also a very good chimera, and the most important thing is now that the 00:35:33 genetic mutation, which we have introduced in the ES cells and which is present in all these mice, will be transferred through the germline to offspring and then you end up with your mutant mouse. So, this is about it what I wanted to show you on the generation. 00:35:51 I have to go outside here with the movie. Why doesn't this work? So anyway, now...now, we are in the right position. So what this should have illustrated to you in this first 8 minutes is that we have powerful tools via ES cells to generate basically any mouse. 00:36:15 It will, with a genetic makeup, like you have outlined in the first place. And now, let's see how important Fos protein, transcription factors, how they are for bone biology, and I have listed here the three Fos proteins. We are mainly working on c-Fos, 51, 52. There is also fourth one, FosB. 00:36:36 And now, either if you generate transgenic mice by overexpression, like I have shown you by the microinjection port, you can get animals which develop consistently and inheritable for decades, osteosarcomas whereas if you resume to the ES cell technology and make lots of function mutations for 00:36:56 c-Fos for example and the other Fos proteins, you can see you get either osteopetrosis or very severe osteopenia in the case of 51 and 52. And what I will do now in the main part of the 15 minutes left is I would like to show you three stories from these experiments. 00:37:12 And before it, I will give you a very brief introduction to the Fos proteins. So here, at the lower panel, you can see the classical Fos-mutant animal, which is like the Src-mutant animal in osteopetrotic mouse in contrast to the Src and many other osteopetrotic mutants, you have not 00:37:30 diagnosed...not a single osteoclast present, instead you have many more microphages. The implication of these functional microphages, I have no time to speculate in this presentation. So, what else do we know about these mutations? So, we know that clearly, in the signaling, which you have heard in 00:37:50 the first talk via M-CSF and RANK ligand, Fos is the determining factor because without the Fos, you will never make an osteoclast into a dark mouse. We know also that the Fos proteins in gain of function experiments, have very important roles in osteoblasts because Fos proteins can transform 00:38:11 osteoblasts, in particular c-Fos can lead to osteosarcoma formation. Now, the signaling of Fos is complex, and as you have heard before, it depends on the angle you look at. So, if my angle is from the transcription factor field, you looked the downstream of c-Fos, you have other transcription 00:38:27 factors, and they are important to forming osteoclasts, and you can genetically analyze this, and as you have heard before, you have upstream kinases which receive this signal, either from the osteoblasts or from soluble cytokines and they activate and downstream pathways. 00:38:42 In case of M-CSF receptor, we do believe that goes via PI3-kinase and ERKs via Ras (ph) down to c-Fos. And in terms of RANK ligand, there are multiple steps where you can activate either the AP-1 pathway or the NF-kappa B pathway, but this is not all, this is much more complex and if we look, 00:39:01 there is downstream signaling via interferons, which again can make your feedback loop to keep this lineage intact and functionally operating and many more targets, I guess to be discovered. But for this talk now, I would like to show you, based on mouse models, the genetics, how you can analyze, 00:39:23 how important are these transcriptional targets, for example of c-Fos, downstream of M-CSF and RANK ligand signaling. And so what do you want to do? Basically, you have four Fos proteins and you want to understand what is the function of these proteins in osteoclastogenesis. 00:39:39 Question #1, can 51, as an example, which is downstream of c-Fos, can be substitute for the function of c-Fos? You know, without c-Fos in the osteoclasts, if you replace c-Fos, can you get osteoclasts being formed? So, those of you who think about how can you genetically or experimentally prove 00:40:00 it, you would say, well, let's just replace one gene for the other and then see what happens. And this is exactly what we have done, and for the aficionados in transcription factology, Fra is different from Fos by virtue of having no transactivation domain essential for transformation. 00:40:18 So the question is then, well, can 51 really do it? And what you do here is now the technology. You have to design a vector on the sketch board, and then you convert it into a DNA, and then you transfer this DNA into ES cells, and make sure that you replace, here in this case the Fra-1 gene, you 00:40:36 put it in the c-Fos locus, you disrupt c-Fos coding sequence, but have Fra-1 under the control of the regulatory elements of c-Fos. And this, you have to characterize in vitro and then, when you have characterized it, as I said before, the ES cells in vitro, you then do the next step - you do your blastocyst injections. 00:40:54 You get your mice and then you breed them genetically, make sure that you have this allele c-Fos, Fra-1, meaning that Fra-1 is in the place of c-Fos, and then you see what happens to the mice, and this happens. So, if you have two copies of Fra-1 instead of c-Fos, the mouse at this level looks indistinguishable from a control mouse. 00:41:16 Yet, if you have only one Fra-1 in place of c-Fos and the other one is Fos minus, the mouse looks identical to a Fos minus mouse, which I have introduced to you, which is an osteopetrotic mouse, which is growth retarded, has no teeth, etc., etc. So, you can do now histology and you can convince 00:41:32 yourself indeed that you have bonafide osteoclast present in these knockin animals. You don't have any in the Fos minus and you don't have any...if you have one allele replaced. Then you can go on and study these cells in vitro and then you find out that if you culture the cells in vitro, things look very different. 00:41:50 But again, we are talking about models in vivo and I would like to carry on and conclude this part one of the mouse models. That from the data, which I have shown you that the transcriptional activation domain seems to be dispensable for Fos functions in vivo because as I have shown you, two 00:42:07 copies of Fra for Fos, look Mickey Mouse, which looks almost the same as a wild-type mouse. It doesn't mean that if you challenge this mouse with inflammatory insults and so forth, that this can cope to the same extent. So there, I think there will be differences or symptoms of transformation most likely. 00:42:24 Okay, so I have told you now about that Fra is almost as good as Fos, but not as good as you have seen. Now what do we know about Fra? In general, we know that Fra is an essential gene. So you cannot easily study now the role of Fra-1 in mice because if you knockout Fra-1 in a mouse model, the animal is dead, as you can see here. 00:42:45 There is some embryogenesis going on, but it is very much retarded and very much in bad shape. It will never make it to life. On the other hand, we know that over expression by DNA injections by transgenic technology, not via ES, gives rise to an osteosclerotic phenotype where increased 00:43:03 osteoblast differentiation in a cell autonomous manner seems to effect this lineage being responsible for this osteosclerosis. So what do you do with mouse technology? You can either knock it in, this I've shown you and we have discussed it or you can make a conditional allele. 00:43:19 And this is now quickly the second story I will tell you. In the conditional allele now, again, you need to make a DNA vector. This was done by Robert Eferl in the lab. Now, this may be a bit tricky, but what you should remember now, now you don't want to substitute gene A for gene B. 00:43:36 Now, you want to flank your gene of interest, in this case Fra-1, with recognition sequences, which can be recognized by a recombinase. And this recombinase can be activated when you want it to activate and then you can turn the gene off and then, we call it a conditional knockout animal. 00:43:51 And then, you can verify when you do so that this is a delta allele and then, you have DNA analysis, which convinces you that you have done it in the right way. And what you do normally is not to go immediately with a bone specific recombinase lane. 00:44:05 If you want to find out whether my survival of animals is compatible in the absence of Fra-1, then you employ Cre recombinase strain where the gene of interest is inactivated in the entire embryo, but not in the placenta because we know that in the Fra knockout mice, they most likely die from a placental phenotype. 00:44:26 So, in this case, you can then ask is life compatible without Fra-1 and you get a Fra rescued, viable, knockout, Fra-1, delta, delta animal in this case. And the question for bone biologists or people interested in osteoclast is clearly, do they have a bone phenotype and the answer is yes. 00:44:45 They lose bone, they are osteopenic, but they don't have a massive osteoclast phenotype unless you take these cells out in culture. So what is the mechanism for the osteopenic phenotype? We have shown and have published that Fra is basically dispensable for development of most organs, but it 00:45:05 controls the expression of matrix proteins, and thereby, it's most likely important for the maintenance of osteoblast activity and one good candidate gene is this matrix Gla protein, which we do believe is a transcriptional target of Fra-1. Part two. 00:45:20 So I've shown you how we do conditional mouse mutants and how we can put this in perspective on how Fra-1 is different from c-Fos. Now, last story has to do with the third Fos related protein, which is called Fra-2, and again, I will confine most of what I am saying in my last part now, again to osteoclasts. 00:45:39 And here again, you can either make overexpression studies with the transgenic technology by DNA injection or you can do loss of function experiments. And if you know very little about the function of a gene, you go and do a conditional allele and knock it out completely and see what happens, and this is exactly what we have done. 00:45:57 Before I briefly show you the data on these experiments, I mention the people who have mostly involved in this project lately, Aline Bozec, a postdoc in the lab, and Latifa Bakiri, and also Robert Eferl, and Hasselblatt, and the bone histomorphometric analysis was done in collaboration with (inaudible) School. 00:46:17 Now Fra-2 has been suspected to play a role in osteoblast as well as in osteoclast because in vitro experiments have pointed in the direction that this transcription factor does indeed is expressed in the cells of interest and may be functionally important. 00:46:33 And we don't have to go through all these statements. I think this, you can also find in the literature. So for a mouse model personnel you knock the gene out, either conditionally or constitutively and you ask, is Fra-2 too indeed needed in any of these bone cell compartments, from the chondrocytes to the osteoblasts over to the osteoclasts. 00:46:53 And we'll focus here now, mainly on the osteoclasts, and I will mention a bit the osteoblast activity. So again, targeting strategy, since I need to talk about technology a bit here. Again, you make a flox allele, flanked by loxP sites. You remove the gene of interest either in the ES cells to 00:47:10 get a delta allele, and then, you convert these ES cells into mice and ask, and you can also put in now GFP marker to visualize the activity of the protein. And when you do so, Aline and others found out that all these mice, which have a delta allele inherited and they are homozygote for this allele, they died between day one and five. 00:47:32 They are growth retarded. They are not growth retarded at P1 and P2, but at P5, you can see these animals really are very runted and very much growth retarded. They have several defects. For the bone biologists, I think the most important defect is illustrated on this slide. 00:47:48 You can see, you have severe bone loss by measuring both long bones, von kossa staining, can look through the MicroCT or look through the vertebrae, but you can also look in the calvaria the drastically reduced bone mass, and BV/TV is clearly reduced. 00:48:06 You can now go on and mark as for early and late osteoblast lineage and you find that also the osteoblast lineage is affected. You can do this in vitro. I go over it very quickly. You see, hardly any bone nodule is being formed in the absence of Fra-2, but what about the osteoclasts? 00:48:22 This is an osteoclast session, let me show you that indeed, there is a giant osteoclast phenotype, meaning that you have huge osteoclasts, multi-nucleated osteoclasts, and also the number and the size of these clasts are drastically increased in the absence of Fra-2. 00:48:37 As you can see here by these parameters and also illustrated by some markers for the osteoclast lineage, carbonic anhydrase II or cathepsin K. Now, you would like to mimic that phenotype in vitro and this we failed completely. So, we cannot culture FOSL1 (ph). 00:48:56 FOSL2 extends to fos-like antigen 2, this is the geneticists insist, it's identical to Fra-2. You can see that you cannot form proper osteoclasts and the markers of osteoclasts are drastically down, but MCF RANKL is not sufficient, you could argue then, well, you have to do your coculture 00:49:15 experiments and then play around with the cytokines and maybe, you can rescue this deficiency or even go and make giant osteoclasts in vitro. This is what we have done, but we failed as you can see here. A coculture never can rescue this defect; however, we are pretty good with MCF Rank ligand and TNF, but we are best if we add LIF. 00:49:36 So the important thing is that the big osteoclasts are completely rescued if you add LIF. Now since I have to give you a good introduction, you may not know what LIF is. So LIF is called leukemia inhibitory factor. LIF is part of the LIF receptor gp 130 complex, and is activated Raf, Jak-STAT. 00:49:56 It has important functions in mouse development, ES cell differentiation, hematopoietic differentiation, etc. and in bone biology, as you will see in a minute. It is expressed in blasts and chondrocytes and it can effect the differentiation of chondrocytes, blasts and clasts as shown by Natalie Sims. 00:50:05 Interestingly, LIF receptor knockouts die after birth. They have a placental phenotype and they have most importantly, osteopenia with a giant osteoclast phenotype. They reminisce with the Fra-2 knockout mice as shown. So now the next, what you will do is, you go and see biochemically is indeed LIF...is this controlled by Fra-2? 00:50:36 For this, you do chromatin IP analysis and I am not going to explain to you exactly how this is done biochemically. For you, important to know is that in bone cells, Fra-2 and c-Jun, they can bind directly to LIF promoter element whereas in the absence of LIF...absence of Fra-2, they cannot bind and then Fra-1 is the core thing to decide. 00:51:01 So it's a direct transcriptional target, that's the bottomline. This is the important conclusion from this analysis. Now, then you go and assay directly in bones whether indeed LIF receptor, LIF and stock components of the stockpiles are down regulated, that you can see here very nicely, that 00:51:22 this pathway is completely down regulated, that the RNA and that the protein level, you can do immunohistochemistry for the receptor, and you can see indeed this drastic, reduced amounts of the receptor present in dissections. So then you ask, well, if indeed if LIF is the target, if you look in 00:51:39 LIF knockout pups, do they develop also bone phenotype? And the answer is yes. And they also develop, as you can see here, for the Fra knockout, a giant osteoclast phenotype. We are in the process of doing detailed histomorphometry to really assess the fact that indeed, you have giant osteoclast presence. 00:51:58 And by virtue of culturing all these cells in vitro, you can analyze which lineage is indeed affected and to our surprise, it was the osteoclast lineage. No osteoclast can be formed in the absence of LIF. This deficiency can be rescued by addition of LIF and if you look at the osteoblast lineage 00:52:14 here for nodule formation, there is no difference. So what do we conclude from this data? We can conclude that Fra-2 knockout pups exhibit a severe osteopenia, there are osteoblast defects, and most importantly, they come down with a severe osteopenia due to giant osteoclast phenotype. 00:52:31 And I've shown you the evidence how Fra-2 regulates the LIF pathway, that we have a direct binding of LIF and Jun to promote the element, and that the differentiation defect can be in vitro rescued by addition of this growth factors, and that the LIF knockout and the LIF receptor knockout have very similar in vivo phenotypes. 00:52:52 So to put it in a scheme, LIF is an important regulator. This is a model how Fra controls LIF expression. Fra is expressed in all bone cells and give LIF and LIF receptor expression in chondrocytes, osteoblasts, and their receptor in osteoclasts, and if you now remove Fra-2, you drastically reduce the amount of LIF, circulating LIF. 00:53:16 This, mechanistically we don't know, it can be either inhibition of cell of apoptosis where other mechanisms which leads to extended survivor and duplication and fusion of osteoclasts, which are responsible for the giant osteoclast phenotype. So I will end here, I think on time, with my last 00:53:37 slide, which shows you that signaling that from Fos proteins and other in osteoclastogenesis is always dependent from which angle you look at. You have this lineage, which is obviously shared with the microphages in the first phase of differentiation. 00:53:56 And then you have, as you heard also in the first talk, the M-CSF pathway. For signaling people, it goes via the Met kinase pathway down to AP1 and activate via the Fra-1 or Fra-2, these are important regulators of osteoclast differentiation. And then, in the next step, you have the osteoblasts, 00:54:14 which can provide the soluble or membrane-bound ligands, which again can function via JNK and NF Kappa B pathways to influence the decision to make a proper osteoclast. And I summarize with the last part, which you have heard already that, there is clearly the ITAM molecules and signaling through 00:54:34 PLC gamma, which again adds additional complexity in the interferon pathway to make a proper osteoclast and keep the osteoclast in shape and in proper function. So, the takehome message, hopefully, I've shown you how mouse models are being generated. 00:54:51 I've lined out why I think mouse models are important. I have given you three examples from the Fos field and I have...at the very last, I have shown you a new pathway, which I think is very important in osteoclastogenesis and generally, in bone cell differentiation. Thank you for your attention. 00:55:14 Steven Teitelbaum: Erwin, the morphology of those osteoclasts look very much like the ship knockout osteoclasts. 00:55:22 Erwin F. Wagner: Oh, I know that this is your favorite molecules, but... 00:55:25 Steven Teitelbaum: Well, I didn't want to say that Erwin, but you said it. But the question is, those cells are hypersensitive than M-CSF for example. Have you looked at that to see whether or not there is an acute sensitivity to osteoclastogenic cytokines? 00:55:42 Erwin F. Wagner: Well, the strict answer is no, we haven't looked because we've only recently established a LIF colony and doing the colonization (ph) of the isolate symptoms right now. Steven Teitelbaum: Patty? 00:55:53 F. Patrick Ross: Erwin, that was lovely. The size of the osteoclast suggests there may be more effusion, so if you looked at the levels of DC-STAMP for example... Erwin F. Wagner: Yes...yeah... 00:56:02 F. Patrick Ross: ...in the LIF knockouts or the LIF receptor...? Erwin F. Wagner: ...we've looked at all of these candidates. DC-STAMP is not altered at all, but MITF is drastically up regulated, so maybe this is another target and this could be explained in fusogenic ability. 00:56:16 I think that we have to analyze in greater detail, but DC-STAMP is not altered. Steven Teitelbaum: Eddie? 00:56:23 Eddie Lee: Excellent talk. Do you get induction of NFATC1 in the Fra knock into the Fas locus mice? Erwin F. Wagner: Yeah, I... Eddie Lee: So you don't need transactivation domain to induce NFATC1. 00:56:37 Erwin F. Wagner: Yes, I think the answer is yes. 00:56:40 Steven Teitelbaum: Thank you Erwin. Our next speaker is Laurie McCauley. Laurie is Professor and Chair at Department of Periodontics and Oral Medicine at the University of Michigan. As you all know, parathyroid hormone has become a major 00:57:06 player in both the anabolic and catabolic activities of skeleton, and nobody really has contributed more to our understanding of this at the cellular level than has Laurie and so, very delighted that you are going to talk to us about "Parathyroid Hormone and Osteoclasts, Changing and Bone Remodelling and Regeneration." 00:57:25 Laurie K. McCauley: Thank you Dr. Teitelbaum, Dr. Teti, and the organizers. I am truly honored to be here and share with you our work on the role of PTH and the putative role of an osteoclast in the actions of PTH in bone remodeling and regeneration. As an introduction to remind you, PTH and PTHrP are potent agents in bone. 00:57:58 They have overlapping biologic activities by binding to the same receptor on osteoblasts and their actions depend on the dose and their mode of administration. PTH has been well characterized as a catabolic agent in bone and it does this indirectly through its action on osteoblast to upregulate 00:58:19 RANK ligand and stimulate osteoclast differentiation. PTH is also well known for its anabolic actions in bone, although these are less clearly defined. There have been several theories that it increases osteoprogenitor cell proliferation, osteoblast survival, it upregulates various growth factors 00:58:39 and potential transcription mediators in both osteoblasts and osteoclasts. The likely, least well-characterized role of PTH is that of an antianabolic action, and this occurs at least through one mechanism of increasing matrix Gla protein. So, these three apparently divergent roles of PTH in bone 00:59:05 have set up a complex scenario to try and dissect out mechanisms of action. Our lab has focused on the anabolic actions of PTH and I am going to try and convince you that these do not act singularly through the cells in the osteoblast lineage. So, if we simply try and understand an increase in 00:59:27 bone mass or an anabolic action, we can look at cells of the osteoblast lineage that go through a differentiation and positively contribute to bone mass. So the osteoclast lineage undergoing their own well-characterized differentiation balance out that bone mass as negative mediators. 00:59:44 Much of the work of PTH has focused on the cells with receptors, the mature osteoblasts, and looking at genes that may positively mediate bone mass, although this singularly has not been able to account for the anabolic actions of PTH. What I am going to share with you today is that in fact, on a 01:00:06 mature osteoblast, PTH has a fairly negative effect at evoking an action on bone mass and the effects of PTH on mature osteoclasts are very little. So, this sets up a scenario that the mature cells in the bone microenvironment are likely not the key players in anabolic actions of PTH. 01:00:30 However, we have found that PTH is a very positive regulator of less differentiated or preosteoblastic cells. And is also a positive regulator of hematopoietic stem cell or early cells in the osteoclast lineage. We know as well that there is important crosstalk between these cell types as 01:00:53 PTH upregulates RANK ligand to support osteoclast differentiation through the osteoblast. And I am also going to show you data that suggests that cells in hematopoietic and osteoclast lineage may also crosstalk back and positively support osteoblast activity. 01:01:10 So, I am going to focus first at the little bit of data on the cells in the osteoblast lineage and then spend a little bit more time on the osteoclast hematopoietic and then the crosstalk between the two. So, Nabanita Datta in our group has shown that PTH has very divergent actions on cells in 01:01:32 the osteoblast lineage, that on the preosteoblast, under defined conditions, we see that it actually upregulates cyclin D1 to promote cells to go through the cell cycle and increase in cell numbers. However, in cells that are mature osteoblasts, PTH has actually an opposite effect, also through 01:01:56 cyclin D1, inhibiting cyclin D1 and preventing cell cycle progression. In addition, PTH prominently inhibits mineralization in vitro and if we just look at a classical mineralized nodule formation assay, PTH dramatically inhibits mineralized nodule formation in this. 01:02:20 And this is consistent with studies in the literature that have shown that PTH down-regulates various osteoblast phenotypic markers that would be associated with bone formation and upregulates matrix Gla protein. So, if the direct action of PTH on osteoblasts is to inhibit mineralization, but we 01:02:41 still get an anabolic action of PTH, this suggests to us that there is an intermediate cell or an indirect effect of PTH on the anabolic actions. So what I have just shown you is that PTH inhibits the cells that are actually producing the matrix, but it actually has an effect on the earlier cells to increase 01:03:03 their proliferation and hence then promote downstream bone formation. One of the questions that we have been seeking to ask is, is this a direct or indirect effect in vivo? Several years ago, we looked at a candidate...is the cells in the osteoclast lineage as candidates for this? 01:03:25 And we've have looked at several different stages of osteoclast differentiation in studies that we performed with Russell Taichman at Michigan. We looked at blocking stromal-derived factor 1, which we showed was upregulated in the bone microenvironment with PTH. 01:03:41 It's an early promoter of cells in the hematopoietic lineage and if we blocked SDF1, we are able to dampen an anabolic action of PTH. In addition, if we administered OPG, we were also capable at dampening an anabolic action. And then, moving to genetic models that you just heard about in the 01:04:03 previous talk, we looked at anabolic actions of PTH in c-Fos knockout mice, which have no osteoclasts, and we totally abolished the anabolic action of PTH, and I will show you some of this data in the next two slides. But if we looked further in osteoclast differentiation at the cSrc knockout mice 01:04:23 that have osteoclasts but do not function, we were still able to mount an anabolic response to PTH, suggesting that there is a cell in the osteoclast lineage, but perhaps not an active or a functional osteoclast. Just a couple of slides to represent this data and this is a model system where we 01:04:46 administer PTH daily during the time of very active bone growth and I would just note at this time that this model and the other models that I am going to show you are really models of bone growth or regeneration and not necessarily models of osteoporosis. 01:05:03 So what we found is when we administered PTH to the wild-type mice, it had a dramatic increase in vertebral bone area, similar findings with the heterozygote mice, but the knockout mice, they are being osteopetrotic at a higher bone area to begin with, but they responded in an opposite manner such 01:05:24 that it was clear they responded to PTH, not only did they not have an anabolic response, but there was an apparent inhibition. So one of the goals was to try and rescue the osteoclast phenotype in these mice to see if the osteoclast was playing an important role in this action. 01:05:42 And to do this, we've used vertebral transplant model where we dissect out vertebral bodies from mice at 4 days of age, and implant them subcutaneously into immunocompromised mice, administer PTH or vehicle, and then analyze the bone growth in the implants. 01:06:02 This is a histologic view at day 4 prior to implantation. And this is after 21 days. We call these implants now vossicles. They are ossicles from the vertebrae, and at this point, we know that they consist of mesenchyme that is from the donor mice, and the marrow is mostly derived from the host, but 01:06:24 there is some carryover of elements of the hematopoietic cells from the donor as well. This shows one of the implants from a c-Fos knockout mice and compared to the wild-type, which at 4 days, you already see a marrow cavity and trabecular bone developing, in the c-Fos knockout mice, you have a much more rudimentary bone organ. 01:06:47 However, after 21 days of implantation, we can see in a c-Fos knockout mouse, with TRAP staining, you see numerous TRAP-positive osteoclasts and a rescue of the c-Fos osteoclast defect from the host. And now, when we look at the anabolic actions in this model looking at the vossicle implants, we 01:07:11 see that vossicles from wild-type, heterozygote, and knockout mice respond in a similar fashion and all display an anabolic action to PTH. So this suggests that the rescue of the hematopoietic lineage cells have restored the ability of PTH to mount an anabolic response. 01:07:31 And then, looking at the c-Src knockout mice, we saw in knockout mice that are, again, they are osteopetrotic as well, but we were able to still see an anabolic response with PTH, suggesting that the necessity for functional resorbing osteoclast was not necessary. 01:07:52 So, what I've just showed you here...the working model at this point is that PTH acts on cells in the osteoblast lineage to evoke an effect in the cells in the hematopoietic lineage. We know they differentiate into c-Fos positive cells and then ultimately c-Fos and c-Src positive cells, but we 01:08:15 think these cells may be signaling to the proliferative osteoblast to stimulate matrix production. Now, as I mentioned to you before, the thought that this anabolic action may be mediated by the release of growth factors would be suggested against with the c-Src model. 01:08:37 Another evidence for cells in the hematopoietic lineage to play an important role is we routinely do BrdU labeling in these model systems, and we see a lot of BrdU positivity lining the tubercular surfaces in vehicle mice, but in mice treated with PTH, we see a more widespread and greater BrdU 01:09:00 labeling suggesting that there are cells in the marrow that are targets of PTH action. So, from this then, we decided to try and dissect out more PTH actions on cells in the osteoclast lineage, and as I indicated before to you, we know that PTH crosstalks to stimulate osteoclast differentiation 01:09:24 through RANK ligand, and there has been several other mediators that have been shown to be affected by PTH action, M-CSF, MCP-1 and VCAM-1 that are expressed in osteoblasts in response to PTH or PTHrP. But the one lingering question was could PTH or PTHrP act directly on cells in the osteoclast lineage? 01:09:49 And so, we set out to answer this question by looking at PTH effects in osteoclast differentiation from a stage of early hematopoietic cells through to differentiated osteoclasts. In this system, we isolate total bone marrow from mice and we amplify the bone marrow for 8 days with the addition of 01:10:09 Flt3 ligand, which increases the myeloid population in the marrow. And so, the first time point is after 8 days is just an enumeration of cells, and we know that Flt3 ligand amplifies cells in this, and this has been shown in the literature. If we look at PTH alone, it has no effect on this 01:10:32 amplification; however, if we looked at PTH in combination with Flt3 ligand, it acted as an accessory to further the amplification of cells in the hematopoietic lineage. The next data point then is to replate the cells at equal numbers and stimulate them to differentiate with the addition of M-CSF 01:10:52 and RANK ligand and enumerate TRAP positive cells. And now when we look at this, in this second phase of the assay system, which at this point now, there are no longer stromal cells present. We see that the flt3 ligand was capable at increasing osteoclastic cells, but PTH alone had no effect, and 01:11:12 at this point, did not significantly alter the flt3 ligand effect. This sets up a scenario where PTH is evoking an action at early progenitor cells, but not specifically at the differentiation of osteoclasts in the absence of stromal cells. We have also done a similar study in vivo where we 01:11:34 isolate bone marrow from mice that have been treated with PTH or vehicle and do flow cytometric analysis of the lineage negative Sca-1+, c-kit+ population and see that PTH increases that population in vivo as well. Later in the differentiation, and activation scheme of osteoclasts, you heard about 01:11:56 the cytoskeleton in the first talk of the session, osteoclasts undergo a very well characterized patterning when they are plated on glass. And we evaluated the effects of PTH on the formation of podosome clusters, rings, and belts on glass, and then finally, the formation of a ceiling zone on a mineralized substrate. 01:12:18 PTH, at 15 minutes, in osteoclastic cells had basically a similar effect as in control and that of a typical podosome formation. At 2 hours, we saw similar belt formation in control in PTH samples, and on a mineralized matrix, the formation of a similar ceiling zone in control in PTH-treated 01:12:44 cultures suggesting that PTH is not impacting this late stage activity of osteoclasts. So from here, we decided then to go back into the in vivo system where we could try to separate out the mesenchymal and hematopoietic compartments, and for this, we use an ossicle model system where bone marrow stromal cells are expanded in vitro. 01:13:09 We place it into a Gelfoam sponge, implanting on the back of athymic mice, PTH or vehicle is administered, and what we get are the small ossicles, ectopic ossicles of bone that are well vascularized. This model system is actually very responsive to PTH. 01:13:29 We get a more dramatic anabolic response in this model system than we do in the endogenous bone of the mice bearing the implants. We see a dramatic increase in trabecular and cortical bone and overall increase in bone volume. Histologically, we see increase in trabecular bone in the model system 01:13:50 and I think you can also appreciate the fact that we see a much more hematopoietic marrow with PTH administration versus a more fatty marrow in vehicle implants. We have characterized this model system temporally in its response to PTH in a very detailed manner and I am just going to show you a couple of key points of the model system. 01:14:16 We typically find the best response, anabolic response of PTH if these cells are implanted, then one week later, PTH is administered for three weeks and that's shown here. Now, sometimes questions arise as to a potential mesenchymal contribution from the host carrying the implants and we've 01:14:39 looked at this in various different modes, and to our knowledge, there is no or very little contribution from the host to the mesenchymal compartment. One piece of evidence for that is, if we do the implants and then wait 12 weeks without treatment, and then we treat for three...the same three-week 01:14:59 period with PTH, we don't see an anabolic response. And in fact, these ossicles appear as dense cortical shells with no or very little trabecular bone suggesting that the host was not able to replace mesenchymal cells to the implant to respond to PTH. 01:15:16 The other point that's worth mentioning is, we have found that during the first week, the first week after implantation, and we start our injections after one week, during this next week is the period of time that these ossicles are very responsive to PTH action. 01:15:36 And at that time, we have characterized the gene expression in this system and we know that the cells are osteocalcin-negative at this point, but after one week, they are highly osteocalcin-positive at similar levels to endogenous bone organs suggesting that PTH is targeting this window of cells as they 01:15:58 are transiting from being preosteoblasts to osteoblasts. Using this model system, we've also done a modification of it where we can track cells in vivo, in real-time, using a luciferase tracking. So we've isolated bone marrow stromal cells from mice that constitutively express luciferase in all their tissues. 01:16:22 Now, these ossicles of bone contain bone marrow stromal cells that are luciferase positive, and we can image these using a CCD imaging to track the growth of the bone ossicles in the same animal over time. In this data panel I am showing you now, we followed a similar regime where the cells were 01:16:46 implanted and then after seven days, injections were begun. And what we noticed is in the controls, we see an increase in luciferase activity and that this is associated with the normal growth of the ossicles. The PTH has a dramatic upregulation and the luciferase activity in these ossicles that 01:17:06 peaks out after a few weeks, but the most dramatic upregulation is during this first week. This model system, we think is...this is the first time such a model system has been shown to show that in vivo, in real-time, there is an increase in the cell numbers in response to PTH, and they are the cells 01:17:30 that we've implanted and not cells from the host. We validated this system by looking at BrdU positive labeling where we see an increase with PTH and we've also done luciferase immunostains to show that the luciferase positive cells are cells that are morphologically consistent with cells of the osteoblast and osteocyte lineage. 01:17:54 So finally, using this model system, we asked the question, if we inhibited osteoclast activity with the addition of a potent bisphosphonate, zoledronic acid, could we alter the anabolic response of PTH? So, with the same regime where we implant and one week later, then administer our treatments, 01:18:17 and then go for a three-week period of time and sacrifice. These are representative images and this graph now shows just the control samples where you see a small increase, but just not a very impressive increase with luciferase activity. The thing that I think is important to note is that 01:18:39 zoledronic acid alone is basically paralleling and exactly the control samples suggesting that zoledronic acid itself is not altering the cell numbers, the bone marrow stromal cell numbers. PTH has a dramatic increase in the luciferase activity reflecting our increase in bone marrow stromal cell 01:18:59 numbers and the combination of the bisphosphonate with the PTH was capable at blunting this action. Now, I also just want to point out to you that if you look at the combination, early on, you see that it's appearing to increase similar to PTH, but then blunts off with time. 01:19:19 And this may be relevant to the interpretation of the next data slides that I am going to show you. But this shows you the temporal course over a period of 24 days. I am going to show you some of the endpoint data points at day 28 now. So looking at the cellular profile in the implants, we see 01:19:39 that the numbers of osteoclast per millimeter of bone, as we would expect, are increased with PTH. Zoledronic acid was capable at reversing that. The BrdU positive cells that we note in the bone marrow were increased with PTH and also reversed with the zoledronic acid. 01:19:56 Our serum biochemistry follows a similar trend. Our TRAP 5b levels are elevated with PTH, reversed with a combination of zoledronic acid, and our serum osteocalcin levels elevated with PTH and reversed with the addition of zoledronic acid. So, at this point, all the data points suggest that 01:20:17 zoledronic acid is really compromising the anabolic action of PTH. So we were a little bit surprised when we saw the bone areas and bone volume fractions of these by MicroCT. What we found was that PTH has an increase in percent bone volume. The zoledronic acid had a similar, if not higher, increase 01:20:42 in bone area by traditional histomorphometry or by MicroCT analyses. However, if we look at this data in a little bit different way, if we look at the overall size of the implants, remembering that these are implants that start from no-bone and then progress into a bone organ, the overall size of 01:21:05 the implant was largest in PTH as shown diagrammatically below. With addition of Zoledronic acid, the overall size was similar to the vehicle or zoledronic acid. The total bone volume, and that's the total amount of bone, was higher in PTH, and zoledronic acid was capable of blunting that, although I note not all 01:21:27 the way back to vehicle or zoledronic acid alone levels. But the percent bone volume that I showed you in the previous slide, was similar in the two, suggesting and our calculated marrow volumes being that the combination of zoledronic acid was capable at restricting the marrow cavity. 01:21:46 So this suggests that PTH increased not only the present bone volume, but also the overall bone volume and the zoledronic acid was capable at blunting the increase in the overall volume of the implants. So I would just also like to mention...emphasize that in the bone marrow microenvironment, we have more than just osteoblasts and osteoclasts. 01:22:12 There is a wide variety of other cell types that could be important in some of the messages between PTH and its targets. In a poster that we described yesterday and in other work that we found in the lab, I can just briefly say to you that we see no alteration in anabolic actions of PTH in the 01:22:31 absence of T-cells when we have reduced numbers of B-cells, when we've increased dendritic cells, when we reduced the hematopoietic rapidly proliferating cells through irradiation. And in our hands, what we see the most dramatic downregulation or inhibition of anabolic actions of PTH has been when 01:22:53 there is a significant compromise in the osteoclast population. So, in summary, what we have found is that PTH increases a proliferative...has a very positive proliferative phenotype on cells in the osteoblast lineage and we believe that this is mediated through an intermediary cell in the osteoclast lineage. 01:23:14 Just to acknowledge some of the people who are responsible for this work. Nabanita Datta in our lab, Amy Koh, and Glenda Pettway were the major contributors to this work. And collaborators, Burak Demiralp, at Hacetteppe did some of the initial c-Fos work, and my collaborators in Lyon, Pierre Jurdic Adrienne Anginot and Anne Chabadel in 01:23:36 his lab helped with the osteoclast and cytoskeletal assays. Steve Goldstein and Jeff Meganck in his lab, who have been long-term collaborators for our microCT work, and Russell Taichman for his contributions to the bone marrow osteoblast environment. Thank you and I'll now entertain questions. 01:24:05 Steven L. Teitelbaum: Really big message. Roberto? 01:24:09 Roberto Pacifici: Laurie, it was really a wonderful talk. I was wondering if you could illuminate us on the nature of the signal that allows the cell, the intermediate cell as you call in the osteoclastic lineage to potentiate or make possible an anabolic response to PTH. Is it a soluble factor? Is it a cell to cell communication? 01:24:33 Laurie K. McCauley: I would love to be able to illuminate you on that. We, at this point we really don't know. We've done a series of microarray analyses to look at potential candidates, but what I can tell you is that...my gut 01:24:51 impression would be that it was a cell to cell contact, but I really have no solid data to say that. We have some putative mediators, but we are way too early to even speculate. 01:25:00 Roberto: Thank you. Steven L. Teitelbaum: Go. 01:25:03 Joseph Lorenzo: Laurie, again very nice presentation. I am trying to understand what you are implying. Are you saying that you think that there is an early hematopoietic cell that might directly respond to PTH? 01:25:16 Laurie K. McCauley: Actually, I don't think it's the early...okay, let me back up. We definitely see that PTH increases the early hematopoietic cells, but I don't think that it's an early hematopoietic cell that actually signals to the osteoblasts. Those are probably two separate events, but you know, they could be integrally related. 01:25:37 But I think it's a cell later in that lineage that's actually providing the signal. 01:25:43 Joe: But do you think that cell has a PTH receptor? That's what I am getting at. 01:25:48 Laurie K. McCauley: I don't think the cell has a PTH receptor. 01:25:50 Joe: Do you think it has to go through a stromal response? 01:25:53 Laurie K. McCauley: I think so. Joe: Okay, thanks. Steven L. Teitelbaum: Nancy... 01:25:59 Nancy Lane: Hi Laurie, great presentation. I just have a question. I want to put this in the perspective of an old person. What would happen if you didn't take 4-day old neonatal vertebrae and put it in? If you took a mature animal and did this experiment, do you have any idea what you might see or have you done that? 01:26:21 Laurie K. McCauley: Yeah...no, we haven't done that experiment. It's a good question. We, you know, today, we have limited ourselves to models that would be perhaps more appropriate to applications in regenerative... 01:26:32 Nancy: Yeah, I see... Laurie K. McCauley: ...but, and so, but, you know, it's true it could definitely have applications in an older individual or model as well. I mean, when we have done implants of older bone, you would just don't see such a robust bone formation. 01:26:47 So, what we've been really pleased with this model is that it provides us kind of a window on time that we get a very dramatic response and that we can really start to sort out mechanisms that perhaps then we could take to another model like you are saying. 01:27:03 You know, after we get an idea in a model that is really robust then do we see a similar phenomenon in another model. I mean, the other complication with an older model or an OVX model is you have cells at all stages of the lineage that are responding in you know... 01:27:19 Nancy: Sure...sure...sure. Laurie K. McCauley: ...in different actions... Nancy: Sure. Laurie K. McCauley: ...of PTH... Nancy: Sure. 01:27:22 Laurie K. McCauley: ...was in this model we have... Nancy: Much work... Laurie K. McCauley: ...a really bulked for the cells that are giving us this response. 01:27:28 Nancy: Well, I hope you get a chance to look at older models because we see a lot of activity in the marrow, kind of even when they are older. Thank you. Laurie K. McCauley: Thank you. 01:27:36 David Roodman: Roodman, Pittsburgh. Really wonderful presentation. Laurie K. McCauley: Thank you. 01:27:41 David: Flt3 ligand expands both stem cells as well as progenerators and those can be sorted by a variety of markers nowadays. Have you had the opportunity to look at subpopulations within the expanded population? One would predict that if you really think it's cell to cell interaction, you could 01:28:00 take stromal cells and take different subpopulations and then do time-course experiments to sort out which population is doing it. 01:28:07 Laurie K. McCauley: Yeah. We are actually doing those experiments right now, the sorting that you talked about. Thank you. 01:28:13 Sayyid: Sayyid from Rochester, Minnesota. A great talk and I had a question on the fact that you said that the irradiation in stem cells did not result in alteration in the anabolic effect of PTH, but earlier on, I was wanting to understand that PTH is anabolic in your ossicle model on early, but 01:28:34 not late cells, so what I am trying to understand is, how does this translate in terms of the origin of osteoblastic cells? 01:28:43 Laurie K. McCauley: Yeah, I guess I am not exactly sure what you are asking, but the experiments that we did with the irradiation were we irradiated and then treated with 01:28:54 PTH for a period of 20 to 30 days and then, the endpoints what we saw was no compromise and in fact, if anything we may have seen, an increase. One of the, perhaps confounding things in that model is that the irradiation will affect the rapidly proliferating cells, but then you have downstream 01:29:16 ramifications of that and so we haven't totally sorted out all of those aspects. Sayyid: Thank you. 01:29:24 Morten Karsdal : Morten Karsdal of Copenhagen, Denmark. So you are suggesting that one of the things is that osteoclasts are secreting something affecting osteoblasts and so we are 01:29:33 investigating the question of osteoclast anabolic, is there something coming from osteoclasts that are stimulating osteoblasts? And we've investigated that in some detail and Dr. Hendrickson from our group is today presenting that. Indeed, there are secreted signals from mature osteoclasts that are stimulating bone formation. 01:29:55 And secondly to that, that also means that if osteoclasts, some osteoclasts, and not all the osteoclasts are anabolic, you should be able to manipulate osteoclast function so you could inhibit osteoclast resorption and at the same time have more osteoclasts and stimulate formation. 01:30:13 And that can actually be done and we have seen it in the osteoimmunology session that if you inhibit acidification of the osteoclastic resorption, it could...I think those data go along line with the anabolic signal from osteoclasts... 01:30:26 Laurie K. McCauley: Yes, thank you. Morten: Just that... Laurie K. McCauley: I think what you are suggesting definitely is consistent to what we've seen. David? 01:30:32 David Rowe: Yeah Laura, just a followup on our poster conversation yesterday. As I mentioned to you that the first stage in these nodules that form is that the endothelial cells proliferate and expand and they are coming from the donor in those marrow stromal cells. 01:30:52 Do you know whether PTH has an effect on that aspect of the biology? Is that early proliferative effect that you are seeing with your luciferase? Could that be on an endothelial population and not on osteoprogenitive population? 01:31:08 Laurie K. McCauley: I think it's an intriguing hypothesis. We've not looked at that specifically, David, but I mean, I think it is certainly something that's worth evaluating. 01:31:16 David Rowe: Does anybody know whether PTH affects endothelial cell? 01:31:20 Laurie K. McCauley: Well, there has been a series of studies of PTH on angiogenesis and endothelial cells. As I know, most of them have been done in vitro that are specifically targeting the endothelial cells. Some of the angiogenesis studies have actually been a bit conflicting both pro and anti-angiogenic. 01:31:42 But in fact, there very likely is a contribution of kind of a pro-angiogenic contribution. 01:31:50 Steven L. Teitelbaum: Last question please? 01:31:53 Lynn Chin (ph): Lynn Chin from New Jersey. I actually have two questions. One is that in your ossicle model, I noticed that you have extra bone marrow stromal stem cells to implants, but later on, you noticed there are osteoclasts in this model. Do you know where these osteoclasts come from? 01:32:12 Laurie K. McCauley: Osteoclasts in our stromal cells? Lynn Chi: In your ossicle model. Laurie K. McCauley: Oh the ossicles. Okay, so the osteoclasts that we see later? 01:32:20 Lynn Chin: Yeah. Is that from a host or from the origin of... Laurie K. McCauley: Those are from the host. 01:32:24 Lynn Chin: Okay, so there is this invasion of the host cells... Laurie K. McCauley: Yes. Lynn Chin: ...into this model. Laurie K. McCauley: Yes. 01:32:29 Lynn Chin: And the second question is, I noticed the...that's very interesting, the PTH increases cyclin D1 expression and I noticed your poster, that's...it's about you did a test in the MC3T3 cells, and have you tried other cells and have you tried in the in vivo bones? 01:32:48 Laurie K. McCauley: We've looked at in vivo as well and in fact, we have looked at in this ossicle model, and interestingly, early in the ossicle development, the effect of PTH is to upregulate cyclin D1, but if we look later in the model, we no longer see an increase in cyclin D1 and it's 01:33:05 consistent with what we see as PTH having a proliferative effect early and not later. So, our in vivo studies are consistent with that. 01:33:16 Lynn Chin: Thank you. Laurie K. McCauley: Thank you. 01:33:17 Steven L. Teitelbaum: Absolutely the last question. 01:33:19 Richard Norles (ph): Okay thanks. Richard Norles from Wyeth. Very nice talk. The data with the bisphosphonate and zoledronate, have you thought of using any other antiresorptive agent such as raloxifene? If so, what was the data on that? 01:33:37 Laurie K. McCauley: We've really not looked...really not had an intention to compare bisphosphonates. In fact, you know when we set out to do the experiment, we really just wanted to go for...you know, using it as a tool, not at all to look at kind of therapeutic implications, but just to...how could we really wipe out osteoclasts? 01:33:58 And so, we went...with zoledronic acid, we used high dose and we used frequent administration. So, I guess to answer your question, that's not something that has been on our list of things to do. 01:34:10 Richard Norles (ph): Yeah, I am just wondering if it's a bisphosphonate's specific effect or if it's...? 01:34:12 Laurie K. McCauley: Yeah. I mean once that model system has been shown, it certainly could be used for something like that to compare potencies and/or for potential combinatorial applications. Richard Norles (ph): Thank you. 01:34:25 Steven L. Teitelbaum: Well, thanks to the speakers for this fabulous session. Thank you very much.