Survey
* Your assessment is very important for improving the workof artificial intelligence, which forms the content of this project
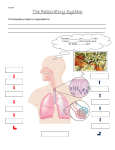
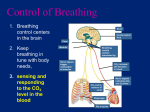
Course: BIOL1040 Topic: Circulation Produced by cyrionTM. © 2014. All rights reserved. Module IV – Circulation and Gas Exchange GAS EXCHANGE Respiration occurs in two different ways: o Cellular respiration – The process of producing energy from glucose. o Gas exchange – The process of taking up oxygen and discharging carbon dioxide. The method by which oxygen is inhaled and carbon dioxide is exhaled is simply passive diffusion – simple diffusion that allows O2 and CO2 to move along its concentration gradient. Passive diffusion always implies movement from regions of high to low partial pressure. The concentration of gases can be measured using partial pressure – this is the pressure exerted by a particular gas in a mixture of gases. partial pressure = % of gas × pressure of gas EXAMPLE: Air usually contains 21% oxygen, and at sea level, the atmospheric pressure of oxygen is 760 mmHg. Hence the partial pressure of oxygen is p𝑂2 = 0.21 × 760 = 160 mmHg. Partial pressure applies to liquids as well – the solubility of a gas in liquids is directly proportional to the partial pressure of that gas in equilibrium with the liquid. The respiratory medium is the source of the oxygen – this can either be air or water. Gas exchange occurs at the respiratory surface of the respiratory organ – this is where oxygen moves in and carbon dioxide moves out via passive diffusion. As a result, respiratory surfaces tend to be moist, large and thin. There are many types of respiratory organs: o Gills in fish are outfoldings of the body that are suspended in water. They have a very large surface area to provide more respiratory surface for gas exchange. A process called ventilation maintains the partial pressure gradients of O2 and CO2 across the gill. It is a closed system. o Tracheal systems in insects are made up of air tubes that branch throughout the body. The largest tubes (tracheae) open to the outside of the insect’s body surface. The air sacs are formed from enlarged portions of the tracheae, and are found near organs that require a large supply of oxygen. The surfaces of the air sacs are moist to transfer oxygen. It is an open system. o Lungs in mammals are localised respiratory organs. Air is inhaled from the nasal cavity and pharynx, passing through the larynx, trachea and bronchi to the bronchioles, which end in microscopic alveoli lined with thin, moist epithelium. The highly branched structure of the lungs provides huge surface areas (80 – 100m2) for gas exchange. Branches of pulmonary arteries transport oxygen-poor blood to the alveoli, while branches of pulmonary veins transport oxygen-rich blood back to the heart. 1 The process of ventilation maintains high O2 and low CO2 concentrations at the respiratory surface. The process that ventilates the lungs is called breathing. There are two main types of breathing: o An amphibian employs positive pressure breathing, inflating lungs with forced air flow. During inhalation, muscles lower the floor of the oral cavity to draw air in, and subsequently the floor rises to push air down the trachea. During exhalation, air is forced back out via compression of the muscular body wall. o A mammal employs negative pressure breathing – this is the process of pulling rather than pushing air into the lungs. During inhalation, the diaphragm contracts and moves down, expanding the thoracic/chest cavity, lowering the air pressure in the lungs below that of the air outside their body. As gas moves from high to low partial pressures, air rushes through the nostrils and mouth down to the alveoli. During exhalation, the diaphragm relaxes and moves up, reducing the volume of the chest cavity, forcing air up and out of the body. Within the chest cavity, a pleural sac forms a double membrane surrounding the lungs, similar to a fluidfilled balloon surrounding an air filled balloon. The inner membrane adheres to the outside of the lungs, while the outer membrane adheres to the wall of the chest cavity. The pleural sac contains a thin space filled with pleural fluid – this sticks the two membranes of the sac together, allowing them to slide smoothly across each other, but not allowing them to be pulled apart easily. Negative pressure must be maintained – normally lungs are inflated such that they fit snugly within the ribcage. This is because there is an elastic recoil that pulls the chest wall outward. If the lungs are stabbed, they collapse, and the surface area reduces. The tidal volume refers to the volume of air inhaled and exhaled with each breath (~ 500 mL). o The vital capacity refers to tidal volume during maximal inhalation and exhalation (3.4 – 4.8 L). o The residual volume is air that remains after a maximal exhalation (~ 1.2 L). With age, residual volume increases at the expense of vital capacity. 2 Breathing is regulated by involuntary mechanisms – these mechanisms ensure that gas exchange is coordinated with blood circulation and metabolic demand. o The stimuli for regulation of respiration comes not from levels of O2, but rather levels of CO2. o Sensors in the carotid arteries and aorta detect the pH of the blood, which send signals to the medulla. The medulla itself also detects the pH of the cerebrospinal fluid. This is because blood CO2 concentration determines the pH of the cerebrospinal fluid. o CO2 diffuses from the blood to the cerebrospinal fluid, where it reacts with water to form carbonic acid (H2CO3). o H2CO3 then dissociates into a bicarbonate ion (HCO3-) and a hydrogen ion (H+), reducing pH. 𝐶𝑂2 + 𝐻2 𝑂 ⇌ 𝐻2 𝐶𝑂3 ⇌ 𝐻 + + 𝐻𝐶𝑂3 − (decreased pH) EXAMPLE: During exercise, increased metabolic activity increases the concentration of CO2 in blood as it is not removed fast enough. This lowers the pH of the blood via the above mechanism. Sensors in blood vessels (such as the carotid arteries and aorta) detect the decrease in the blood pH, which then signal the medulla. The medulla itself also detects the decrease in the pH of cerebrospinal fluid. Subsequently, the medulla signals the rib muscles and diaphragm to increase the rate and depth of ventilation. This decreases CO2 levels, restoring the pH back to homeostatic levels. 3 There are two mechanisms for the transport of large quantities of oxygen and carbon dioxide: o PARTIAL PRESSURE – In the lungs, gradients of partial pressure favour the diffusion of O2 into the blood and CO2 out of the blood. The gradients exist due to cellular respiration, which removes O2 from and adds CO2 to the surrounding interstitial fluid. This is also known as Fick’s law of Inhaled air diffusion, which 𝑝𝑂2 = 160 mmHg states that the 𝑝𝐶𝑂2 = 0.2 mmHg diffusion rate of a gas across a fluid Exhaled air membrane is 𝑝𝑂2 = 120 mmHg proportional to the 𝑝𝐶𝑂2 = 27 mmHg difference in partial pressure, area and thickness of the membrane. Partial pressures of O2 and CO2 can vary in different parts of the circulatory system, as well as in inhaled air compared with exhaled air. The diagrams adjacent illustrate the partial pressures of O2 and CO2 at varying stages of respiration. o RESPIRATORY PIGMENTS – In the rest of the body, O2 is transported via proteins called respiratory pigments, found in the blood or haemolymph. Respiratory pigments overcome the low solubility of gases in blood by greatly increasing the amount of O2 that the blood can carry (from 4.5 mL dissolved O2 per litre of blood to 200 mL carried by respiratory pigments per litre of blood). Pigments have a distinctive colour, and usually consist of a protein bound to a metal. The blue pigment haemocyanin has copper as its oxygen-binding component, and is found in arthropods and many molluscs. The red pigment haemoglobin has iron as its oxygen-binding component, and is found in almost all vertebrates and many invertebrates. Haemoglobin consists of four subunits (polypeptide chains) each with a cofactor called a heme group with an iron atom at its centre. Each iron atom is able to bind one molecule of O2, and so one haemoglobin molecule can carry four molecules of O2. 4 o o When one O2 binds to a subunit, the other subunits change their shapes slightly, making it easier for more O2 to bind. This phenomenon is called positive cooperativity. The opposite is also true – when one subunit unloads O2, the other three subunits unload O2 more readily. As such, even a small change in 𝑝𝑂2 causes haemoglobin to load or unload a large amount of O2. The production of CO2 lowers the pH of surroundings, which in turn decreases the affinity of haemoglobin for O2, causing O2 to be unloaded more readily. Haemoglobin not only transports O2, but CO2 as well – this assists in buffering the blood. 5