Survey
* Your assessment is very important for improving the workof artificial intelligence, which forms the content of this project
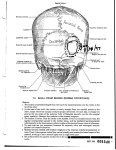
THE ANATOMICAL RECORD 262:344 –368 (2001) New Fossil Hominid Calvaria From Indonesia—Sambungmacan 3 SAMUEL MÁRQUEZ,1– 4 KENNETH MOWBRAY,5,6 G J SAWYER,5 TEUKU JACOB,7 AND ADAM SILVERS8 1 Departments of Cell Biology and Anatomy, Mount Sinai School of Medicine, New York 2 Department of Anthropology, City University Graduate Center, CUNY, New York 3 Department of Anthropology, Lehman College, City University of New York, New York 4 New York Consortium in Evolutionary Primatology, Department of Anthropology, The Graduate School, CUNY, New York 5 Division of Anthropology, American Museum of Natural History, New York 6 Department of Anthropology, Rutgers University, New Jersey 7 Laboratory of Bioanthropology and Paleoanthropology, Faculty of Medicine, Gadjah Mada University, Yogyakarta, Indonesia 8 Department of Radiology, Mount Sinai School of Medicine, New York ABSTRACT A morphologically distinct partial calvaria of Homo cf. erectus from Java, Indonesia is described. The fossil hominid Sambungmacan 3 (Sm 3) was first discovered in 1977 from the banks of the Solo River near the village of Poloyo, Sambungmacan district, in central Java. It was later recovered in a New York City natural history establishment in 1999 and quickly returned to the Indonesian authorities. Examination of Sm 3 shows that the calvaria is well preserved with only portions of the cranial base missing. The most striking characteristics of Sm 3 include: the presence of a vertically rising forehead, more open occipital/nuchal and frontal angles, a more globular vault, and a cranial capacity within the Homo erectus range. Most notably absent in Sm 3 are a number of the classic characters attributed to Homo erectus, such as a strongly expressed angular torus and a continuous supratoral sulcus. The absence of such characters would normally place the calvaria outside the range of Homo erectus (sensu stricto), however, overall quantitative and qualitative morphological assessments of Sm 3 place it within the Homo erectus spectrum. The combination of the morphological characters in Sm 3 may be interpreted in several ways: 1.) the known cranial variation of H. erectus from Indonesia and China is extended; 2.) this calvaria shows evidence of evolutionary change within H. erectus; or 3.) more than one species of Homo existed in the (presumed) Middle Pleistocene of Java.) Anat Rec 262:344 –368, 2001. © 2001 Wiley-Liss, Inc. Key words: Homo erectus; Java; Sm 3; vertically rising frontal; open occipital/nuchal plane angle; cranial capacity The “rediscovery” of a morphologically distinct partial calvaria of Homo cf. erectus from Indonesia was announced in New York on August 29, 1999. Its recent history is somewhat unclear (discussed below), but its unique features were quickly recognized in the summer of 1999 when it was brought to the American Museum of Natural History (AMNH). During its eight-week stay at the AMNH, we felt it was important to thoroughly examine the fossil in order to document its morphology with written descriptions, photographs, and computer tomography (CT) imaging, and subsequently report our findings as a preliminary chronicle of its recovery (Mowbray et al., 2001). Accompanying this description are two articles involving a paleoneurological assessment of the specimen (Broadfield et al., 2001), and a brief interpretive study using three-dimensional coordinate data and comparative morphological observations on selected specimens attrib© 2001 WILEY-LISS, INC. uted to African and Asian Homo erectus and extant Homo sapiens (Delson et al., 2001). According to several sources, Sm 3 was discovered in 1977 from the banks of the Solo River near the village of Poloyo, Sambungmacan district, Sragen country, in central Java, some 50 km north of the Sangiran Dome. The Grant sponsor: National Science Foundation; Grant number: SBR9634519; Grant sponsor: National Institutes of Health; Grant number: F31-DC00255-01. Correspondence to: Samuel Márquez, Department of Cell Biology and Anatomy, Mount Sinai School of Medicine, One Gustave L. Levy Place, Box 1007, New York, NY 10029. E-mail: [email protected] Received 12 June 2000; Accepted 20 December 2000 Published online 28 February 2001 SM 3 CALVARIA 345 Fig. 1. Map of Indonesia with the inset highlighting the site of Poloyo, Java where Sm 3 was first reported to have been found. Map courtesy of Angela M. H. Schuster, Archaeology Magazine. calvaria was reportedly discovered by miners excavating sand from the Solo River for use as construction material. Although the exact location and date of discovery cannot be confirmed (Jacob, 1999), it is thought that it may have been discovered near Ngadirejo in Sambungmacan between the villages of Chemeng and Poloyo (Fig. 1). The calvaria was purchased from the discoverer by an unknown agent and eventually ended up in an antiquities shop in Jakarta. In 1997, Dr. Boedhihartono, an Indonesian paleoanthropologist, was invited to examine the specimen in the antiquities shop in order to determine its authenticity; he published a brief description and provided some measurements and photographs (Boedhihartono, 1998). In the spring of 1999 the specimen “surfaced” in Maxilla & Mandible, Ltd., a New York City natural history establishment owned by Henry Galiano. Galiano, who is quite familiar with the techniques used for cleaning original fossils from his days in the Department of Vertebrate Paleontology at the AMNH, cleaned the external surface of the calvaria and removed the matrix from the internal surface for future geological study. In late June, Galiano brought the fossil to the AMNH for examination. While I. Tattersall, K. Mowbray, G.J. Sawyer, and E. Delson examined the calvaria to determine its authenticity, it was learned from Dr. Hisao Baba of the National Science Museum in Tokyo, Japan, and Professor Teuku Jacob of the Gadjah Mada University, Yogyakarta, Indonesia, that the calvaria might be the same fossil hominid that an Indonesian antiquities dealer had tried to sell the previous year in Jakarta. After receiving a partial copy of Boedhihartono’s 1998 report with photos of the calvaria, it became evident to us that the fossil hominid under examination and the calvaria from Jakarta were the same specimen. Accordingly, Mr. Galiano contacted the Indonesian government and arranged to return the fossil to them. On August 29, 1999, in a formal ceremony conducted at Maxilla & Mandible Ltd., Mr. Galiano presented the specimen to Professor Jacob and various representatives of the Indonesian government. In return, Mr. Galiano was honored with an award from the Indonesian government in recognition of his generosity and thoughtful commitment to science. While in New York, Professor Jacob temporarily assigned the designation of Poloyo 1 (Pl-1) (Mowbray et al., 2000) to the specimen but, once back in Java, he changed the accession code to Sambungmacan 3 (Sm 3) in recognition of its provenance in the same region as the calvaria Sm 1 (Jacob, 1975) and tibial fragment Sm 2 (Baba et al., 1990 and Matsu’ura 2000). The precise sites of discovery may be designated Sambungmacan Locality 1 and Locality 2. 346 MÁRQUEZ ET AL. TABLE 1. Metrical descriptions and definitions (taken from Wood, 1991) No. Description 1. 2. 3. Length (glabella—opisthocranion) (C) Minimum frontal breadth Maximum bi-parietal breadth 4. Maximum bi-temporal breadth 5. 6. Biporionic breadth (C) Supramastoid crest breadth (C) 7. Maximum mastoid breadth at base (C) 8. 9. 10. 11. 12. 13. 14. 15. 16. Glabella—bregma (C) Glabella—bregma (A) Bregma—left pterion (C) Bregma—left pterion (A) Bregma—right pterion (C) Bregma—right pterion (A) Parietal sagittal length (C) Parietal sagittal length (A) Parietal right temporal length (C) 17. 18. Parietal right temporal (A) Parietal left temporal length (C) 19. 20. Parietal left temporal (A) Parietal coronal breadth (C) 21. Parietal coronal breadth (A) 22. Parietal right lambdoidal length (C) 23. Parietal right lambdoidal length (A) 24. Parietal left lambdoidal length (C) 25. Parietal left lambdoidal length (A) 26. 27. 28. 29. 30. 31. 32. Bregma right asterion (C) Bregma right asterion (A) Bregma Left asterion (C) Bregma Left asterion (A) Lambda—inion (C) Lambda—inion (A) Inion—opisthion 33. 34. 35. 36. 37. Occipital Sagittal length (C) Occipital Sagittal length (A) Biasterionic breadth (C) Biasterionic breadth (A) Width temporal gutter 38. 39. Width temporal gutter Vertical distance of supraorbital torus 40. Vertical distance of supraorbital torus 41. Anteroposterior thickness of right supraorbital torus (C) 42. Anteroposterior thickness of right supraorbital torus (A) 43. Anteroposterior thickness of left supraorbital torus (C) Definition Chord distance Minimum chord distance between frontotemporale (ft-ft) Breadth across homologous points on the parietal bone at the site of maximum breadth. Breadth across homologous points on the temporal squama at the site of maximum breadth. Chord distance Chord distance across the maximum projection of the supramastoid crests Chord distance across the points of maximum projection on the mastoid process, wherever that occurred, but excluding the supramastoid crests. Chord distance Arc length Chord distance Arc length Chord distance Arc length Chord distance Arc length Chord distance along the temporal border of the intact parietal (sphenion—asterion) Arc length along the temporal border of the intact parietal Chord distance along the temporal border of the intact parietal (sphenion—asterion) Arc length along the temporal border of the intact parietal Chord distance along the coronal border of the intact parietal (bregma—sphenion) Arc length along the coronal border of the intact parietal (bregma—sphenion) Chord distance along the lambdoid border of the intact parietal (lambda—asterion) Arc length along the lambdoid border of the intact parietal (lambda—asterion) Chord distance along the lambdoid border of the intact parietal (lambda—asterion) Arc length along the lambdoid border of the intact parietal (lambda—asterion) Chord distance Arc length Chord distance Arc length Chord distance Arc length Opisthion not clearly demarcated but measure approximates the posterior border of foramen magnum Chord distance Arc length Chord distance Arc length Maximum distance from the outer wall of the temporal squama at the anterior end of the root of the zygomatic process to the outer lip of the process at that location. Left damaged Chord distance taken at the highest point of the left supraorbital margin. Chord distance taken at the highest point of the right supraorbital margin. Chord distance, taken from above, between the most anterior projection of the torus at the highest point on the superior orbital margin and the posterior limit of the torus, or the shallowest point in the post-toral sulcus. Arc length, taken from above, between the most anterior projection of the torus at the highest point on the superior orbital margin and the posterior limit of the torus, or the shallowest point in the post-toral sulcus. Same as in 41. 347 SM 3 CALVARIA TABLE 1. Metrical descriptions and definitions (taken from Wood, 1991) (continued) No. 43. Description 45. Anteroposterior thickness of left supraorbital torus (C) Anteroposterior thickness of left supraorbital torus (A) Frontal torus breadth (C) 46. Frontal torus breadth (A) 47. Mandibular fossa length (C) 48. Mandibular fossa length (A) 49. Mandibular fossa breadth (C) 50. Mandibular fossa breadth (A) 51. Mandibular fossa depth 44. Definition Same as in 41. Same as in 42. Maximum chord distance across the frontal toral region. The termini for this measure usually lie lateral to frontotemporal orbitale. Arc length across the superior aspect of the torus as defined in 45. Only left is sufficiently preserved. Minimum chord distance between the tympanic plate and the most inferior projection of the articular eminence; taken midway along the breadth measurement Arc distance between the termini described above. Only left fossa was measured. Minimum chord distance in the coronal plane between the tip of the entoglenoid process and the most lateral extent of the articular margin on the articular eminence. Arc distance between the termini described above. Only left fossa was measured. Maximum perpendicular distance between an imaginary line connecting the most inferior projection of the articular eminence and the tip of the postglenoid process, and the roof of the mandibular fossa. MATERIALS AND METHODS This article focuses upon the description of the original fossil of Sm 3 and reports a series of standard morphometric measurements (see Table 1). Quantitative measures that take into account clearly identifiable anatomic landmarks and craniometric points are reported in Table 2. The description and discussion of measurements found in Results are grouped by regions following a protocol developed by Schwartz and Tattersall (in preparation) that strives to encompass all aspects of available skull anatomy, while ensuring the utilization of a uniform descriptive nomenclature that purposely lacks systematic implications. The subsections include: Sex Determination, Supraorbital Region, Cranial Roof, Cranial Walls, Cranial Rear, Cranial Base, Cranial Sutures and Thickness, Anterior Endocranial Compartment, Middle Endocranial Compartment, Posterior Endocranial Compartment, Sinus Drainage System, and Middle Meningeal Vessels. Two subsections (sex determination, cranial sutures, and cranial thickness) not included in the Schwartz and Tattersall protocol are incorporated into the description and require brief explanations. Sex determination in the human fossil record is based on the sexual dimorphism of modern humans (and other primates), that may be attributed to hormonal development as represented by muscle attachment to bone. Marked nuchal rugosities, prominent surpraorbital tori, and strong temporal lines are generally representative of males, while minimal expression of these characters is indicative of females. Problems regarding the accuracy of sexing skeletal material arise when the range of variation for a particular population is unknown, when only one skeletal element is being assessed, with qualitative vs. quantitative approaches, and with “experience” vs. statistical “standardization” (Krogman and Iscan, 1986). These factors may explain why certain characters might not all score as the same sex. Following in the tradition of a number of workers (e.g., Santa Luca, 1980; Weidenreich, 1943; Wolpoff, 1999), the precarious nature of applying these techniques to the fossil record was taken into account when assessing sex. The description of cranial sutures in humans is well documented (Todd and Lyon, 1925), and students of human variation and evolution have used the degree of sutural closure as an age indicator of when death occurred. However, the application of these techniques using adult specimens, fossil or recent, is not without controversy and debate (Brooks, 1955; Buikstra and Ubelaker, 1994; Meindl and Lovejoy, 1985; Perizonius, 1984; Singer, 1953). For example, Weidenreich (1951) evaluated the Ngandong cranial assemblage and estimated that nearly half of the specimens recovered were juveniles based primarily on incomplete ectocranial synostosis. Antón and Franzen (1997) argue convincingly that other cranial features, such as relative cranial development and cranial thickness, should be included during fossil crania examination, rather than relying solely on the lack of overall sutural closure to estimate age. With this caveat in mind, a description and a scoring of the degree of sutural closure are reported (Table 3) along with measurements of cranial thickness (Table 4). To facilitate our examination of endocranial, ectocranial and pneumatization morphologies, we utilized CT imaging of the fossil specimen, and a first generation hemisected cast. Quantitative descriptions are based on standard measurements taken with polycarbonate vernier calipers, spreading calipers, and a flexible cloth tape. Frontal angle and vertex of the calvaria were derived with the specimen in the Frankfurt Horizontal (FH) based on Weidenreich (1951) and Santa Luca (1980) estimated orbital height measure of 38 mm in order to approximate infraorbitale since the calvaria lacked its orbital floors. Endocranial volume determination. Endocranial volume was measured by two different methods: seed fill- 348 MÁRQUEZ ET AL. TABLE 2. Calvaria dimensions of Sm 3 (in millimeters) 1. Length (glabella—opisthocranion) 2. Minimum frontal breadth 3. Maximum bi-parietal breadth 4. Maximum bi-temporal breadth 5. Biporionic breadth 6. Supramastoid crest breadth 7. Maximum mastoid breadth at base 8. Glabella—bregma (C) 9. Glabella—bregma (A) 10. Bregma—left pterion (C) 11. Bregma—left pterion (A) 12. Bregma—right pterion (C) 13. Bregma—right pterion (A) 14. Parietal sagittal length (C) 15. Parietal sagittal length (A) 16. Parietal right temporal length (C) 17. Parietal right temporal (A) 18. Parietal left temporal length (C) 19. Parietal left temporal (A) 20. Parietal coronal breadth (C) 21. Parietal coronal breadth (A) 22. Parietal right lambdoidal length (C) 23. Parietal right lambdoidal length (A) 24. Parietal left lambdoidal length (C) 25. Parietal left lambdoidal length (A) 26. Bregma right asterion (C) 27. Bregma right asterion (A) 28. Bregma left asterion (C) 29. Bregma left asterion (A) 30. Lambda—inion (C) 31. Lambda—inion (A) 32. Inion—opisthion 33. Occipital Sagittal length (C) 34. Occipital Sagittal length (A) 35. Biasterionic breadth (C) 36. Biasterionic breadth (A) 37. Temporal gutter width (right) 38. Temporal gutter width (left) 39. Vertical distance of supraorbital torus (left) 40. Vertical distance of supraorbital torus (right) 41. Anteroposterior thickness of right supraorbital torus (C) 42. Anteroposterior thickness of right supraorbital torus (A 43. Anteroposterior thickness of left supraorbital torus (C) 44. Anteroposterior thickness of left supraorbital torus (A) 45. Frontal torus breadth (C) 46. Frontal torus breadth (A) 47. Mandibular fossa length (C) (only left side is sufficiently preserved) 48. Mandibular fossa length (A) (only left side is sufficiently preserved) 49. Mandibular fossa breadth (C) 50. Mandibular fossa breadth (A) 51. Mandibular fossa depth ing with oil rapeseeds and water displacement. Using the seed-filling method, an initial endocranial volume was obtained directly from the calvaria without any attempt to reconstruct its missing cranial base. A second endocranial volume was derived by reconstructing its cranial base with the use of inert modeling clay. Continual gentle tapping of the calvaria during the filling process ensured seeds were fully packed and settled. Before and after each seed-filling, the calvaria was weighed with an electronic scale (to 0.1 gm). The procedure was repeated five times for both unreconstructed and reconstructed fossil. Weight data were then transformed to volumetric measurements (in milliliters) by linear regression. A reference regression line (r2 ⫽ 0.99 P ⬍ 0.001) was created by using the weights 178.5 101 126.5 127.5 131.5 145.5 126.5 103 106 87 98 87.5 100 98 105 59.7 59.9 62.4 63 72.5 73 70.5 71 71.5 75 125.5 140 123.5 140 50.5 54 40.1 80.5 96 118 126 6.4 — 12.5 13.5 5.9 32 25.2 30 111.2 138 20 21 18.5 20 14.2 for nine known volumes of seeds (5 ml to 45 ml at 5 ml intervals), whereby each was measured twice using two differently shaped graduated cylinders. Rapeseeds were also introduced into the calvaria and measured in a graduated cylinder. No statistical significant differences were recorded for the two seed-filling techniques. Rhodorsil威 silicon rubber endocasts were made from the original Sm 3 specimen and endocast reconstructions were displaced in water to calculate endocranial volume. Results from seed filling and water displacement techniques are shown in Table 5. Imaging modalities. CT imaging was performed using a HiSpeed CT/i scanner (GE Medical Systems, Mil- 349 SM 3 CALVARIA TABLE 3. Assessment of ectocranial suture closure Midline Sagittal 1: pars bregmatica 2: pars verticis 3: pars obelica 4: pars lambdica Coronal 1: pars bregmatica 2: pars complicata 3: pars stephanica 4: pars pterica Lambdoid 1: pars lambdica 2: pars intermedia 3: pars asterica Parieto-mastoid Occipito-mastoid Squamous Spheno-frontal Spheno-temporal Spheno-parietal Left Right 0 0 0 0 0 0 eroded 0 0 1 0 eroded 0 0 0 0 eroded 3 1 0/1 0 0 eroded 0 eroded eroded 1 n/a 0 TABLE 4. Calvaria thickness of Sm 3 in millimeters Glabella Glabella—Bregma midpoint Bregma Bregma-Lambda midpoint Bregma to lambda midpoint Lambda External occipital protuberance Pterion (left) Pterion (right) Pterion Asterion (left) Asterion (right) Stephanion (left) Stephanion (right) Parietal eminence (left) Parietal eminence (right) 21.0 8.5 11 10 11 (adjacent to left parietal) 11 (adjacent to right parietal) 11.5 16 12.5 12 7.5 15 15 7 7.5 12 12 waukee, WI) at the Mount Sinai Hospital/NYU Medical Center. Helical scanning was used with images reconstructed at 1 mm intervals. The CT machine specifications were a 25 cm field of view (DOV), 140 kV, and 170 mA with a 1:1 pitch helical scan. The images were processed on an Advantage Windows Workstation (ADW) (GE Medical Systems) using ADW 3.1 software for surface and volume rendering and to reconstruct other localized three-dimensional structures employing sagittal and coronal reformations. RESULTS Preservation and Condition of Sambungmacan 3 The calvaria shows excellent preservation in both ectocranial and endocranial anatomy with no distinctly noticeable taphonomic deformities. The blackish-gray calvaria is heavily mineralized, extremely well preserved, and shows no evidence of either complete fracture or any bone perforation. There are scratches throughout the ectocranial portion of the calvaria which require further investigation but do not appear to be the result of speci- TABLE 5. Endocranial volume data from Sm 3 Seed filling without reconstructing basicranium Seed filling with reconstructing basicranium 870 cc 890 cc Average water displacement of reconstructed endocast 917 cc men cleaning. The neurocranium is almost complete, while the splanchnocranium and major portions of the cranial base are missing (see Figs. 2–7). Both temporal lines are discernible and the amount of sagittal keeling can be seen anteriorly at bregma. The orbital roofs are relatively complete while an area of postmortem damage inferior to glabella serves as a portal into a possible frontal sinus region (Fig. 8). The mandibular fossae, temporal planes, and mastoids are well preserved, but only the left external acoustic meatus is present. Both mastoid tips are broken, revealing varying degrees of pneumatization. The majority of the sphenoid, basilar, and condylar portions of the occipital and petrous portions of the temporal bones are missing. The nuchal and occipital planes are sufficiently preserved on the external and internal surfaces to determine nuchal rugosity, the angle between the two planes, and to provide information about the posterior cranial fossa. Sutures remain open (see Table 3) on the ectocranial surface of the vault, however, the endocranial surface reveals sutural closure but not complete obliteration. The degree of cranial sutural closure and overall vault thickness suggests that this individual was most likely a young adult when death occurred (see Antón, 1999). Sex Determination The calvaria exhibits a suite of characters that makes sex determination difficult to ascertain. In this regard, male characters include well-developed and elongated suprameatal crests, a well-developed occipital protuberance, and pronounced lateral arcus superciliaris. Characters attributed to females include an overall small calvaria, lack of strongly developed superior and inferior temporal lines, a bulbous frontal region, and a depressed glabellar region. Although its mosaic of characters may be too ambiguous to allow us to properly determine the sex of Sm 3, the overall smaller size and somewhat less rugose expression of features in Sm 3, as compared to most presumed male Javanese fossils (Santa Luca, 1980; Weidenreich, 1951), suggest a female identification (see also Delson et al., 2001). Overall Appearance of Sm 3 The calvaria is relatively short and broad with a steeply rising frontal and a fairly open angle between the occipital and nuchal planes resulting in a relatively globular appearance in side view. In lateral profile, the vault shows little supraorbital distension while exhibiting a globular shaped morphology (see Figs. 2 and 3). Based on CT scans and hemisected casts, the forehead rises steeply before arching posteriorly. Maximum length of the calvaria (glabella to opisthocranion) measures 178.5 mm. Maximum breadth of 145.0 mm across the squamous temporals, and minimum frontal breadth of 101.0 mm. A cranial index (CI) of 81 further indicates a broad-headed individual. 350 MÁRQUEZ ET AL. Fig. 2. Right lateral view of Sm 3 calvaria with original above and cast seen below. SM 3 CALVARIA Fig. 3. Left lateral view of Sm 3 calvaria with original above and cast seen below. 351 352 MÁRQUEZ ET AL. Fig. 4. Superior view of Sm 3 calvaria with original above and cast seen below. SM 3 CALVARIA Fig. 5. Inferior view of Sm 3 calvaria with original above and cast seen below. 353 Fig. 6. Anterior view of Sm 3 calvaria with original above and cast seen below. SM 3 CALVARIA Fig. 7. Posterior view of Sm 3 calvaria with original above and cast seen below. 355 356 MÁRQUEZ ET AL. Fig. 8. Photograph of inferior view showing frontal bone region. Fig. 9. A three-quarter view of a three-dimensional reconstruction of Sm 3. Note the steeply rising frontal squama with no supratoral extension at the midline. Supraorbital Region The lateral margins of the supraorbital region of Sm 3 markedly project anteriorly, whereas the medial portions protrude minimally in the same direction. The marginal surface above each orbit is continuous and relatively uniform in vertical thickness. The two orbital margins, however, are not completely confluent across glabella. As such, glabella is slightly depressed when viewed superiorly, and lacks a distinct angle or bony bulge that clearly delineates it superiorly. The lateral surfaces of the orbital roofs roll smoothly anteriorly, superiorly, and subsequently posteriorly where they form flat plateaus prior to gradually sloping into the lateral walls of the frontal squama. The medial surface of the left orbital roof (the right side is damaged on its inferior surface), however, rolls smoothly anteriorly and superiorly to become somewhat flat and slightly angled as it forms the supraorbital ridge. The medial surfaces of the supraorbital region, unlike the lat- eral surfaces, quickly meet the steeply rising slope of the frontal squama (Fig. 9). A distinct supraorbital notch occurs on the superior margin of the left orbital region and causes a superoinferior thinning in this area. The right side could not be assessed. The middle supraorbital region shows only a slight supratoral sulcus, but both lateral margins have moderate depressions that may be interpreted as sulci (Fig. 10). Post-orbital (PO) constriction is minimal, as reflected by a PO index of 88 (see Fig. 7 in Delson et al., 2001), which is calculated by the (minimum frontal breadth measured at FT-FT/maximum breadth of the supraorbital ridge ⫻ 100). Postmortem damage to the inferior portion of the frontal bone exposes the outer and inner tables of the region with the intervening space filled with matrix. Whether the present infill reflects the replacement of trabecular bone or of empty space could not be deter- SM 3 CALVARIA 357 30 mm superior to glabella and continues through bregma. Sagittal keeling progresses along most of the sagittal suture, tapering off approximately 34 mm anterior to lambda (Fig. 13). Cranial Walls Fig. 10. A superior view of a three-dimensional reconstruction showing the lateral wing extension of the supratorus. mined. Macroscopic imaging of the frontal did not reveal any clear demarcation between matrix infill and mineralized bone. CT imaging of the region suggests that sinus cavities might exist as seen in the very subtle differences of opacity (see Fig. 11A,B,C). These voxel intensity differences, however, could just as likely be from the porous nature of the trabeculae meshwork that may have existed previously. Subsequent to its return to Java, the infill has been removed, revealing an apparently well-developed frontal sinus morphology (Jacob, personal communication). Just above the right supraorbital ridge on the ectocranial surface of the frontal squama are a series of raised interconnecting bony nodules. These osteogenic calluses form a C-shaped narrow ridge of raised bone that is contained within a 20 mm2 area. A few less prominent unconnected nodules also rise up superior to glabella. Examination of the calvaria in its entirety reveals that the raised calluses are concentrated on the periosteum of the frontal bone alone. It is unclear whether they are the result of a nonlethal trauma or of a localized infection that had healed before death. Cranial Roof Beginning at glabella and ending at bregma, the frontal squama in sagittal profile presents a moderate arc. At bregma, the vault begins a symmetrical, curved, backward descent until it reaches opisthocranion. Maximum cranial height of the sagittal profile was estimated to lie posterior to bregma. The metopian– glabella vector formed a 75.3° angle with the estimated FH (Fig. 12). Maximum cranial breadth occurs on the laterally protrusive supramastoid crests. A moderate frontal and a prominent sagittal keel are clearly defined. Frontal keeling begins approximately In coronal profile, at the mastoid region, the walls of the calvaria are mainly parabolic in shape. From the base of the mastoids, the walls initially slope laterally only to quickly rise vertically until they cross the temporal lines, after which the walls curve inward towards the midline (Fig. 14). The temporal lines are weakly expressed on both sides of the calvaria. They originate from the lateral wing of the superior orbital margins and course posteriorly in low arcs, ending near asterion. The posterior root of the zygomatic process of the right temporal bone originates anterior to the external auditory meatus while the left zygomatic process is missing. Both sides have confluent suprameatal and supramastoid crests. On the right, the root of the zygomatic process is continuous with the suprameatal crest. Both suprameatal crests roof over the external auditory meatus. The supramastoid crests are prominent and sweep markedly upward. The left parietal shows a sharp oval depression measuring 1.1 mm in maximum length with three groups of parallel scratch marks. Two of these groups radiate diagonally from the oval “puncture” mark in a posterior direction, while the third group is situated closer to the sagittal–lambdoidal suture. The mastoid processes are of moderate to small size and have a slightly narrow base. The tips of both mastoid processes are broken off, exposing matrix-filled air cells. The left mastoid process is moderately excavated while matrix fills the right process to its outer rim. In inferior view, the processes are triangular with their internal peaks angled obliquely anterior. Their lateral surfaces (mastoid crests) are slightly convex. The left external acoustic meatus is round in outline with its tympanic portion contributing significantly to the deep posterior wall of the mandibular fossa. Only the superiormost portions of the greater wings of the sphenoid are present with the left side clearly intruding between the temporal and frontal bones. This configuration helps form an acute sphenoidal angle of the parietal bone. The squamous portion of temporal bone is moderately long (approximately 1/3 of the overall cranial length). Its maximum height occurs anteriorly, well before its midpoint. This causes its superior border to slope posteriorly until reaching a deeply incised parietal notch. Cranial Rear The sagittal profile of the posterior region is moderately curved with a relatively open angle where the occipital and nuchal planes intersect. The posterior end of the sagittal keel superior to lambda is relatively flat. At the lambdoidal suture, incomplete ectocranial synostosis between the parietals and the occipital produces moderate relief. Continuing inferiorly from lambda, the occipital squama is very slightly curved, approximating an almost flat surface. At the midpoint between the superior and highest nuchal lines the occipital squama presents a moderate buildup of bone. Here, the nuchal plane undercuts the occipital plane forming a 117.7° angle at opisthocranion (Fig. 15). From a posterior view, the superior nuchal lines exhibit well-defined bilateral arches that converge at the inferi- Fig. 11. A: A close-up of the frontal bone from figure 11B showing either some vacuities of the remnant cancellous bone or frontal pneumatization, which has been somewhat obfuscated by mineral infill. B: An arial view of Sm 3 with arrows pointing to the pneumatized regions of the frontal and occipital bones. C: A close-up of the occipital bone region from Figure 11B illustrating cancellous bone. Fig. 12. Lateral view of Sm 3 showing the metopian-glabella and FH vectors forming a 75.3° frontal angle. Fig. 13. Three quarter view of a three-dimensional reconstruction showing midsagittal keeling. Fig. 14. Anterior view of a three-dimensional CT reconstruction coronal slice through the region of the mastoids of Sm 3 illustrating its cranial wall shape. Fig. 15. A sagittal three-dimensional CT reconstruction of Sm 3 showing how the nuchal plane undercuts the occipital plane forming a 117.7° angle at opisthocranion. Fig. 16. A posterior three-dimensional CT reconstruction view of Sm 3 revealing the detailed bilateral arches of the superior nuchal lines that converge at the inferiorly projecting external occipital protuberance. Fig. 17. A close-up posterior view of Sm 3 showing a wormian bone at lambda. 362 MÁRQUEZ ET AL. Fig. 18. bone. A right posteroinferior three-quarter view showing Waldeyer’s crests lying entirely on the occipital orly projecting external occipital protuberance (Fig. 16). The surface surrounding the external occipital protuberance exhibits a markedly rugose texture. The occipital plane is shorter in length compared to the nuchal plane. Likewise, the overall occipital breadth is wide, with the lambdoidal suture exhibiting a low and broad pattern. The lambdoidal suture runs between the asterionic borders with a wormian bone at lambda interrupting it (Fig. 17). Cranial Base There is a well-developed and continuous median nuchal crest from inion to the remaining edge at opisthion. On both sides of the crest, at its superior border, are distinct scalloped regions that correspond to the attachment sites for mm. semispinalis capitis. Similarly, just below the inferior nuchal line, two smaller scalloped regions indicate where mm. rectus capitis posterior minor once attached. Lateral to those depressions, the surface attachment areas of other nuchal muscles are less distinct (i.e., mm. superior oblique). Two large distinct Waldeyer’s crests of varying prominence lie entirely on the occipital, paralleling the occipitomastoid sutures (Fig. 18). Medial to the mastoid processes are distinct occipitomastoid crests; the left is well developed and better preserved than the right. Only the left posterior lateral border of the foramen magnum near opisthion is present. This border exhibits a distinct swelling of bone, which morphologically corresponds to a postcondylar tuberosity (see Weidenreich, 1951). From the remnant of the border, one can discern a narrowing at the posterior region of the foramen magnum just anterior to opisthion, which we refer to as the opisthionic recess. The left mastoid process has better preserved outer walls and presents a hollowed out cavity. The damaged outer walls of the right mastoid process are flush with the lateral aspect of the cranial vault. Both mastoid notches are narrow and relatively short, with the left side more deeply incised than the right. The notches open posteriorly into shallow and narrow digastric fossae. Anteriorly, the left mastoid notch courses into the stylomastoid foramen whereas the right leads into a large excavation that may be related to the jugular foramen. Both styloid processes are lacking and overall damage to the region has obliterated that part of the tympanic forming the vaginal process. An incomplete but discernable C-shaped foramen spinosum is present on the remnant of the left sphenoid, but due to erosion the right is poorly expressed. The damaged left carotid foramen faces posteriorly, but the posi- SM 3 CALVARIA Fig. 19. axis. 363 An inferior view showing the squamotympanic fissure vector forming a 63.5° angle to the sagittal tion of the right carotid is unclear due to poor preservation of the region. The anterior portion of the left mastoid is distinctly separated from the tympanic plate by a prominent tympanomastoid fissure (sometimes referred to as the mastoid fissure). At its widest breadth, the left tympanomastoid fissure measures at least 6.1 mm, while the right side is indiscernible due to the missing tympanic plate. The anterior and posterior walls of the left mandibular fossa parallel each other while the fossa is greater in mediolateral width than in anteroposterior length. Of particular note is the squamotympanic fissure, sometimes incorrectly referred to as the “Glaserian” fissure, which runs the entire length of the floor in the deepest portion of the fossa. The fissure forms an acute angle of 63.5° (i.e., mandibular fossa angle) to the sagittal axis and thus its medial alignment is obliquely orientated (Fig. 19). The well-preserved left fossa is relatively deep, measuring approximately 14 mm, and is constricted somewhat anteroposteriorly. The fossa is bounded anteriorly by a steep pre-articular plane that opens onto a weakly developed articular eminence. A relatively shallow gutter precedes the eminence. The lateral edges of the mandibular fossae do not extend beyond the cranial wall; instead, they are positioned beneath and medial to the protruding suprameatal and supramastoid crests. Cranial Sutures and Thickness Having discussed the precautionary aspects of this subsection earlier, the ectocranial sutures of Sm 3 are uniform, weakly interdigitated, and mostly show lapsed union. We follow here the classificatory groupings of sutures with their subdivisions from Cobb (1952) using the schematic descriptions from Stewart (1954, see appendix 1). The ectocranial sutural interdigitation pattern of Sm 3 is illustrated in Figure 20. Of particular note is the superior border of the left squamosal suture which exhibits a more horizontal rather than arc-like pattern (but see Holloway and Shapiro, 1992). The right squamosal suture is not sufficiently preserved to determine its pattern. At their posterior ends, the squamosal sutures form clear mastoid angles of the parietal bone. The parietomastoid suture is moderate in length and sweeps obliquely in a posterosuperior direction. It measures 3.1 mm in length on the left, and approximately 3.4 mm on the right. Endocranially, both the right and left coronal sutures are completely fused; the right is almost obliterated while the left can be faintly detected. The sagittal and lambdoidal sutures are completely fused but not obliterated whereas the squamosal sutures are completely obliterated. Likewise, the left and right sphenofrontal, sphenoparietal, and sphenosquamosal sutures are obliterated. Fig. 20. The sutural interdigitation pattern of Sm 3. Polysiloxane high-resolution dental impression material was used to mold the sutures and all post-depositional cracks along the sutural borders. As seen in the schematic drawing (see inset), the vinyl mold covered the area of the sutures as they naturally exist in three-dimensional space. After remov- ing the mold from the calvaria, it was sliced at bregma to allow the mold to be laid out on a flat surface. This enabled the imprints of the sutures and perisutural cracks to be accurately traced onto acetate paper, where they are now depicted on a one-to-one scale in two dimensions. Fig. 21. Midsagittal three-dimensional reconstruction shows the relatively high frontal and open occipital angle. Note the relative thickness throughout the cranial vault. The middle meningeal vascular branching pattern can also be appreciated. SM 3 CALVARIA 365 Fig. 22. A three-dimensional CT reconstruction from an oblique posterior view showing the internal morphology of the frontal crest. The parietomastoid and occipitomastoid sutures are fused and lie within slightly concave troughs that follow along their sutural borders. Wormian bones can be seen at lambda and at the junction of the right lambdoidal and occipitomastoid sutures, the latter being unobservable ectocranially. Overall, the calvaria exhibits a fairly thick roof and walls (Fig. 21). The superior portions of the greater wing of the sphenoid are thick and measure about 9 mm at their superiormost border. sella turcica, foramen ovale, foramen lacerum, and the basiocciput is missing. Preserved portions of the sphenoid reveal the lateral wall of the foramen spinosum. Most of the superior portion of the right petrosal revealing the internal acoustic meatus and approximately one half of the left petrosal is preserved. Its damaged anterior portion exposes the superior carotid canal. The superior surfaces of both petrosal portions of the temporal bone exhibit a weak arcuate eminence on the right but absent on the left. Anterior Endocranial Compartment Posterior Endocranial Compartment The anterior endocranial compartment lies largely above the orbits, more so medially than laterally. Clear and distinct anterior rami of the middle meningeal vessels are present on both sides of the concave endocranial surfaces of the frontal squama. There is a sharp and welldeveloped frontal crest that is moderate in length and tapers superoposteriorly (Fig. 22). The foramen cecum is not present. The right petrosal is well preserved, however, the posterior aspect of the left petrosal is somewhat eroded, obscuring morphological detail. The right petrous pyramid acuminates apically throughout the length of its superior surface. Its posterior surface reveals a narrow diagonal channel that runs from the arcuate eminence towards the jugular foramen below the internal acoustic meatus. It is unclear whether this narrow groove is the remnant of a sinus or simply an artifact resulting from taphonomical processes. The aqueduct of the vestibule is present and slightly roofed over. Although there is no subarcuate fossa, the area of the petrous bone corresponding to the Middle Endocranial Compartment Most of the sphenoid bone forming the optic canals, anterior and posterior clinoid processes, pituitary fossa, 366 MÁRQUEZ ET AL. Fig. 23. A: Lateral view showing the left surface area of the Sm 3 endocast. Note temporal pole has been reconstructed. B: Basal view of reconstructed endocast. 367 SM 3 CALVARIA superior vestibular canal is marked by a slight depression. The internal rims of both jugular fossae are missing or damaged. The cruciform eminence prominently demarcates the moderately impressed cerebral and cerebellar fossae. Sinus Drainage System The superior sagittal sinus is distinctly impressed only from bregma to lambda. It divides into two slightly incised transverse sinuses. The right transverse sinus flows into a narrow and short sigmoid sinus, while the left flows into a much wider but equally short sigmoid. Examination of the endocast reveals a slightly impressed occipital sinus accompanied by a strongly expressed marginal sinus on the right side that communicates with the right sigmoid before leading into the jugular foramen (see Endocast of Sambungmacan 3 (Sm 3): A New Homo erectus From Indonesia by Broadfield et al., this volume). Middle Meningeal Vessels The impressions of the middle meningeal vessels are well developed on both sides of the calvaria. On each side of the endocast, the main branch bifurcates as it courses over the anterolateral wall of the temporal lobe. On the calvaria, the anterior division of the main branch courses upwards toward bregma with smaller bifurcations supplying the frontal region and the anterior portion of the parietals. The posterior division of the main branch courses posterolaterally where it soon bifurcates into smaller side branches that supply the posterior region of the parietals and the occipital region. Endocast An endocast was generated directly from the fossil. As Figure 23 shows, the missing portions were reconstructed in order to obtain an endocranial volume through water displacement procedures (see Table 5 for results). DISCUSSION AND EVOLUTIONARY IMPLICATIONS This preliminary study provides a basic description of the newly recovered fossil hominid calvaria (Sm 3) from Java, Indonesia. In general, the calvaria appears morphologically most similar to Homo erectus, depending of course on one’s definition of that particular species. As usual in human systematics, there are differences of opinion as to which characters are diagnostic of Homo erectus (see Rightmire 1993; Stringer, 1984; Wood, 1984; 1991; see also Hublin, 1986; Kennedy, 1991; Wolpoff, 1998), and some authors would restrict this species to eastern Asia (Tattersall, 1995). Additional studies have also detected diversity within the Homo erectus group (e.g., Schwartz and Tattersall, in press). In the context of the traits that distinguish Indonesian Homo erectus from the Chinese and African forms (Andrews, 1984; Rightmire, 1993; Wood, 1991), the calvaria falls comformably within the Indonesian spectrum. There are, however, a number of traits that place the calvaria outside the classic definition of H. erectus (sensu stricto). These include: an absent angular torus, a more open occipital/nuchal angle, a divided supratoral sulcus, a more globular cranial vault, a vertically rising rather than a sharply receding frontal, and the lack of a continuous bar-like occipital torus. The presence/ absence of these characters may be interpreted in several ways: 1.) the combination of this group of characters in Sm 3 extends the known range of cranial variation of H. erectus from Indonesia and China; 2.) this calvaria shows evidence of evolutionary change within H. erectus; or 3.) more than one species of Homo existed in the (presumed) Middle Pleistocene of the Indonesian islands. More exhaustive and comprehensive studies of this fossil calvaria and of variation within living populations will undoubtedly allow us to better understand its taxonomy and evolutionary significance. This is important; for, after all, if one has not properly identified the actors, one cannot possibly hope to understand the play. ACKNOWLEDGMENTS We thank Mr. H. Galiano for allowing us to examine the specimen while it was in his care. We thank Drs. S.C. Antón, T. Bromage, P.J. Gannon, R.L Holloway, J.T. Laitman, O. Pearson, E. Sarmiento, I Tattersall, and B. Wood for their comments and suggestions. We also thank Dr. C. Swisher III for his assistance with the geological provenience of the fossil. KM would like to thank Drs. S. Cachel and M.E. Morbeck for their continued support. SM and KM thank Ian Tattersall for his inspiration and tireless assistance with this project. LITERATURE CITED Andrews P. 1984. On the characters that define Homo erectus. Courier Forschungsinstitut Senckenberg 69:167–175. Antón S, Franzen JL. 1997. The occipital torus and developmental age of Sangiran – 3. J Hum Evol 33:599 – 610. Baba H, Aziz F, Watanabe N. 1990. Morphology of the fossil hominid tibia from Sambungmacan, Java. Bull Nat Sci Mus Tokyo, ser D (Anth) 16:9 –18. Boedhihartono. 1998. A new Homo erectus finding. Antropologi 54: 121–125. Broadfield D, Holloway RL, Mowbray K, Silvers A, Márquez S. 2001. The endocast of Sm 3: a new Homo erectus from Java. Anat Rec 262: Brooks ST. 1955. Skeletal age at death: the reliability of cranial and pubic age indicators. Am J Phys Anthropol 13:567–589. Buikstra JE, Ubelaker DH. 1994. Standards: for data collection from human skeletal remains. Fayetteville, Arkansas: Arkansas Archeological Survey Research Series No. 44. Cobb WM. 1952. Skeleton. In: Lansing AI, editor. Cowdry’s problems of aging. Baltimore: Williams and Wilkins. Delson E, Harvati K, Reddy D, et al. 2001. The Sambungmacan 3 Homo erectus calvaria: a comparative morphometric and morphological analysis. Anat Rec 262:380 –397. Holloway RL, Shapiro JS. 1992. Relationship of squamosal suture to asterion in pongids (Pan): relevance to early hominid brain evolution. Am J Phys Anthropol 89:275–282. Hublin J-J. 1986. Some comments on the diagnostic features of Homo erectus. Fossil man, new facts—new ideas. Anthropus (Brno), 23: 175–187. Jacob T. 1975. Morphology and paleoecology of early man in Java. In: Tuttle R, editor. Paleoanthropology: morphology and paleoecology. The Hague: Mouton. p. 311–325. Jacob T. 1999. The two Sambungmachan skull caps (Sm 1 and Sm 3). Zhoukoudian Symposia, October 12, 1999. Kennedy G. 1991. On the autapomorphic traits of Homo erectus. J Hum Evol 20:375– 412. Krogman WM, Iscan MY. 1986. The human skeleton in forensic medicine. 2nd ed. Springfield: Charles C. Thomas Publisher. Matsu’ura S, Kondo M, Aziz F, Sudijono, Narasaki S, Watanabe N. 2000. First known tibia of an early Javanese hominid. Curr Anth 41:297–300. Meindl RS, Lovejoy CO. 1985. Ectocranial suture closure: a revised method for the determination of skeletal age at death based on the lateral-anterior sutures. Am J Phys Anthropol 68:57– 66. 368 MÁRQUEZ ET AL. Mowbray KM, Márquez S, Antón SC, et al. 2000. The newly recovered Poloyo hominin (PL-1) from Java. Am J Phys Anthropol Suppl. Vol. 30:233. (Abstract). Perizonius WRK. 1984. Closing and non-closing sutures in 256 crania of known age and sex from Amsterdam (A.D. 1883–1909). J Hum Evol 13:201–216. Rightmire GP. 1993. The evolution of Homo erectus, comparative anatomical studies of an extinct human species. New York: Cambridge University Press. Santa Luca AP. 1980. The Ngandong fossil hominids: a comparative study of a far eastern Homo erectus group. Yale University Pubs Anthro 78:1–75. Schwartz JH, Tattersall I. The human fossil record. Volume 1: craniodental morphology of Genus Homo. New York: Peter Nevraumont Publications (in preparation) Schwartz JH, Tattersall I. What constitutes Homo erectus? Proceedings of Zhoukoudian IVPP conference. Morphology of Genus Homo (in press). Singer R. 1953. Estimation of age from cranial suture closure. J Forensic Med 1:52–59. Stewart TD. 1954. Evaluation of evidence from the skeleton. In: Gradwohl RBH, editor. Legal medicine. St. Louis: Mosby. p 407– 450. Stringer CB. 1984. The definition of Homo erectus and the existence of the species in Africa and Europe. Courier Forschungsinstitut Senckenberg 69:131–143. Tattersall I. 1995. The fossil trail. New York: Oxford University Press. Todd TW, Lyon DW Jr. 1924. Endocranial suture closure, its progress Fig. A-1. and age relationship. Part I. Adult males of White stock. Am J Phys Anthropol 7:325–384. Todd TW, Lyon DW Jr. 1925. Cranial suture closure, its progress and age relationship. Part II. Ectocranial closure in adult males of White stock. Am J Phys Anthropol 8:23– 43. Weidenreich F. 1943. The skull of Sinanthropus pekinensis: a comparative study on a primitive hominid skull. Palaeontol. Sin Ser D 10:1–298. Weidenreich F. 1951. Morphology of Solo man. Am Mus Nat Hist, Anthropolog Pap 43:207–290. Wolpoff M. 1998. Human evolution. New York: McGraw Hill. Wolpoff M. 1999. Paleoanthropology 2nd ed. Boston: McGraw-Hill. Wood B. 1984. The origins of Homo erectus. Courier Forschungsinstitut Senckenberg 69:99 –111. Wood B. 1991. Koobi fora research project volume 4, hominid cranial remains. Oxford: Clarendon Press. APPENDIX 1 Using the Todd and Lyon (1924) rating scale permits the creation of a “closure formula,” thus allowing the scoring to act as a somewhat descriptive tool. Particular cranial landmarks further demarcate the three main sutures of the vault (i.e., coronal, sagittal, and lambdoidal), which enables a more detailed description of sutural classification. The descriptive categories of each suture are listed below along with a schematic illustration showing suture subdivisions and the stages of suture closure (Fig. A-1). Stages of suture closure and sutural identification in the human skull.