Survey
* Your assessment is very important for improving the workof artificial intelligence, which forms the content of this project
Electrocardiography wikipedia , lookup
Mitral insufficiency wikipedia , lookup
Hypertrophic cardiomyopathy wikipedia , lookup
Jatene procedure wikipedia , lookup
Atrial fibrillation wikipedia , lookup
Ventricular fibrillation wikipedia , lookup
Arrhythmogenic right ventricular dysplasia wikipedia , lookup
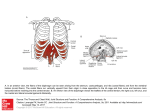
794 Fiber Types and Myosin Types in Human Atrial and Ventricular Myocardium An Anatomical Description Patrice Bouvagnet, Jocelyne Leger, Francoise Pons, Claude Dechesne, and Jean J. Leger From the Institut National de la Sante et de la Recherche Medicale, Laboratoire de Pathologie Moleculaire du Muscle, Institut de Biologie, Montpellier, France SUMMARY. Hybridomas were prepared from mice immunized with myosin from the enlarged left ventricle of a 53-year-old female with an obstructive cardiomyopathy. The specificity of 15 monoclonal antibodies to myosin heavy chains was assessed by the reactivity of muscle extracts and of chymotryptic myosin fragments of different sizes with these antibodies, as determined by the immune replicate technique; some of the monoclonal antibodies cross-reacted only with the ventricular V3-type myosin from hypothyroid rats, whereas the other antibodies cross-reacted both with the latter and with the ventricular VI-type myosins from normal young rats. Immunological heterogeneity of the fibers from human atrial muscles and from human ventricular muscles was detected by some of the antimyosin antibodies by means of indirect immunofluorescence. Histochemical fiber heterogeneity was also detected by adenosine triphosphatase staining of the same tissues. Because of the close correspondence observed between the immunological and histochemical responses of atrial fibers, it has been postulated that at least two distinct types of myosin exist in the human atrium, each myosin form being histochemically related to either a- or /3-like ventricular myosin heavy chains. In contrast, there was no direct correspondence between the two experimental approaches in human ventricles, and it is postulated that at least three distinct types of myosin exist within the human ventricles, one VI-type myosin, presumably corresponding to the very rare fibers with an alkaline-stable adenosine triphosphatase activity, and two other V3-type myosins corresponding to immunologically different fibers, each having an alkaline-labile adenosine triphosphatase activity. Monoclonal antibodies that can distinguish among the different myosin variants were further used to provide the basis for an anatomical description of fiber types and myosin types within the human atrial and ventricular myocardium in the whole hearts of two young boys who died sudden violent deaths. Small zones of myosin variation were seen to be scattered, but probably not randomly distributed, within large areas of myocardium in which the cellular distribution of myosin was constant; the large areas had one myosin distribution specific for each cardiac cavity. No clear-cut conclusions can yet be made concerning the physiological role of the regional variations observed in the distribution of the different molecular forms of myosin (Circ Res 55: 794-804, 1984) CARDIAC myosins exhibit molecular heterogeneity within the ventricles and atria of some mammals, according to evidence derived from several different experimental approaches (Hoh et al., 1978; Flink and Morkin, 1979; Klotz et al., 1981; Lompre et al., 1981; Sartore et al., 1981; Chizzonite et al., 1982; Gorza et al., 1982; Litten et al., 1982; Banerjee, 1983). These molecular variants of myosin heavy chains play a role in controlling myocardial function, both under normal conditions of development and during heart disease (Schwartz et al., 1981; Rupp, 1982; Swynghedauw and Delcayre, 1982). Regional variations in muscle fiber distribution, corresponding to different distributions of molecular variants of myosin heavy chains, have been detected in beef atrial and ventricular myocardium by immunofluorescence: it has been suggested that certain muscle fibers having a specific myosin content may be specialized for faster conduction (Sartore et al., 1981; Gorza et al., 1982). Several recent publications mention the possible existence of molecular heterogeneity in human cardiac myosins, but relatively less work has been done on myosins in human hearts. Human ventricular and atrial myosins have different enzymatic activities and different light chain contents (Klotz et al., 1983). Sequence microheterogeneities have been detected in the light chain 2 of human ventricular myosin (Klotz et al., 1982), and variations in light chain contents under pathophysiological conditions have been seen in human atria (Cummins, 1982) and in human ventricles (Tuschmid et al., 1983). Human ventricular and atrial myosins have different heavy chains, as shown by peptide mapping (Klotz et al., 1983), but migrate as a unique and identical band under nondissociating conditions (Lompre et Downloaded from http://circres.ahajournals.org/ by guest on May 12, 2016 Bouvagnet et a/./Isomyosins in Human Atria and Ventricles al., 1981; Clark et al., 1982; Klotz et al., 1983). Human ventricular myosin reacts four to six times more with monoclonal antibodies specific for the V3 myosin heavy chains of embryonic chicken ventricle than with monoclonal antibodies specific for the VI myosin heavy chains of adult rabbit ventricle (Clark et al., 1982). Schiaffino et al. (1983) and Mercadier et al. (1983) have shown that normal human heart contains 0-15% of total ventricular myosin which reacts with antibodies to the rat and beef VI myosin isoform; no similar information is available concerning human atrial myosin. A recent histochemical study of normal human heart has conclusively shown that heterogeneity with respect to fiber ATPase activity exists not only between atrial and ventricular myocardia but also within normal atrial and ventricular myocardium (Thornell and Forsgren, 1982). A rather new experimental method for studying molecular variations in any molecule is the use of monoclonal antibodies; these permit a detection of even slight antigenic differences between closely related molecules, and the hypothetically heterogeneous molecule under study can be used as an immunogen. This method has recently been used to detect previously known and currently unknown molecular heterogeneities of myosins in certain animal muscles (Bader et al., 1982; Chizzonite et al., 1982; Winckelmann et al., 1983; Miller et al., 1983; Wachsberger et al., 1983). Thus, we prepared hybridomas from mice immunized with myosin from the left ventricle of a hypertrophic human heart and observed that muscle fiber heterogeneity can be detected by indirect immunofluorescence within both human atrial and ventricular myocardium. This muscle fiber heterogeneity detected by antimyosin heavy chain antibodies is variably distributed in each cavity of the human heart. We compare it to that detected by histochemistry in beef, human, and rat hearts, by immunofluorescence in rat ventricles, and by other methods in human hearts. We discuss the functional significance of this variable cellular distribution of the different molecular forms of atrial or ventricular myosins in terms of their location in human heart. 795 ventricle and the left atrium were frozen directly in liquid nitrogen and stored at —80°C, if not immediately used. Serial tissue sections (6-8 /xm) were cut with a cryostat at —20°C and either processed for immediate immunofluorescence and histoenzymological studies or stored at —80°C until use. Identical results were observed under both conditions. The anatomical description of fiber types and myosin types within the human heart was made on the whole heart of two 16-year-old boys who died sudden violent deaths. Seventeen tissue samples, each about 1 cm long, were excised from different regions of the hearts immediately after death (Fig. 1). Four samples came from the right atrium and three from the left atrium; six samples came from the right ventricle and four from the left ventricle. Two atrial samples (3 and 12) came from regions of postulated preferential conduction pathways (James and Sherf, 1971); samples 2 and 13 came from the right and left auricle, respectively, whereas the three other samples (1, 4, and 11) came from common atrial myocardium. Two ventricular samples (5 and 6) came from regions of postulated preferential conduction pathways, RA Methods Tissue Sources Adult human hearts were obtained either within 12 hours after death or from renal transplant donors (Nephrology and Urology Units, Hopital Saint-Charles, Montpellier). Prior to surgery, informed consent from the donor's family was obtained. Myosin for mouse immunization was prepared (Klotz et al., 1981) from the hypertrophic left ventricle of a 53year-old female who had an obstructive cardiomyopathy (heart weight: 690 g; degree of hypertrophy: about 200%). Immunofluorescence studies were routinely made on fragments from autopsied adult hearts weighing 350 g or less and apparently free of any cardiac disease. Tissue samples excised from the posterolateral wall of the left L V LA FIGURE 1. Location of excised tissue samples (Netter, 1974). Upper panel: right atrium (RA) and ventricle (RV) opened and viewed from the front. Lower panel: left ventricle (LV) flap opened in the posterolateral wall and sectioned left atrium (LA) from a posterolateral view. Specimens were excised from: RA, 1, anterior trabeculated wall; 2, auricle; 3, crista terminalis; 4, interatrial septum above the limbus of the fossa ovalis. RV, 5, interventricular septum below the supraventricular crest; 6, septa! band; 7, anterior papillary muscle; 8, anterior free wall; 9, conus arteriosus; 10, interventricular septum near the apex. LA, 11, posterior free wall; 12, superior free wall; 13, auricule. LV, 14, mid-portion of the muscular interventricular septum; 15, mid-portion of the posterolateral free wall; 16, anterior papillary muscle; 17, apex. Downloaded from http://circres.ahajournals.org/ by guest on May 12, 2016 Circulation Research/Vol. 55, No. 6, December 1984 796 and 15 came from free walls, and samples 10 and 17 came from the apex, whereas sample 9 was excised from the conus arteriosus located near the pulmonary artery (Fig. 1). All tissue samples were prepared for immunofluorescence and histoenzymological studies as already described. Normal Wistar rats (6 weeks old) and hypothyroid rats of the same strain (5 mg of propylthiouracil forcibly fed daily for 4 weeks) were killed by a sharp blow on the neck. The hearts were rapidly removed and processed like human hearts. Monoclonal Antibodies Mice (strain CB 20 or Biozzi HR) were immunized with myosin from a stock solution prepared from a hypertrophic left ventricle (see tissue sources). This myosin, used as an immunogen, was previously denatured with 2% sodium dodecyl sulfate (Schwartz et al., 1980) and then dialyzed in 50 mM sodium pyrophosphate, pH 7.5. The mice were immunized twice subcutaneously with 100 /xg of myosin emulsified in Freund's adjuvant at intervals of a few weeks. The mice were boosted intraperitoneally with 100 ng myosin 3 days before the fusion. Isolated splenocytes were fused with a P3-X63-Ag8-653 mouse myeloma in the presence of polyethylene glycol, as described by Di Pauli and Raschke (1978). The supernatants of 1024 hybridoma cell cultures from two fusions were screened for human ventricular myosin with a direct solid phase radioimmunoassay (RIA). Most of the positive hybridomas were not immediately cloned, but were kept in culture in RPMI 1640 with 15% fetal calf serum. A few cell lines which appeared to be of special interest for the detection of myosin heterogeneity (see Results) were additionally cloned using a fluorimetric cell sorter; the corresponding immunoglobulins were purified from the ascite fluid of mice injected by each hybridoma line by passage through protein A Sepharose columns. For use in immunofluorescence studies, culture supernatants or purified ascite fluids were pooled and their IgG content was precipitated by the addition of ammonium sulfate at a final concentration of 50%; the collected pellets were solubilized in 150 mM NaCl, 10 mM sodium phosphate, pH 7.4 (PBS), and stored at -20°C until use. The direct solid phase RIA used for screening cell line supernatants for immunoglobulin production and for subsequent antibody titrations was processed as follows: an excess of human ventricular myosin (50 ttl at 20 ttg/ml in 0.5 M NaCl, 50 mM sodium pyrophosphate, pH 7.4) was coated in each well of a 96-well polyvinyl microtiter plate. Fifty microliters of each hybridoma medium were then incubated for 3 hours at 25°C with the coated myosin. After washing, I25I-Iabeled goat antimouse IgG was added and incubation continued for 3 hours at 25°C. After washing, the radioactivity contained in each well was counted. A cell culture supernatant containing no antimyosin antibody was used as a control. The competitive RIA for testing the specificity of any antibody to different human myosins was essentially the same as described by Winckelman et al. (1983). To test the specificity of the antibodies to the myosin heavy chains, total muscle extracts in SDS buffer (Chizzonite et al., 1982), myosin samples, and chymotryptic treated myofibril samples (Klotz et al., 1981) were first separated by electrophoresis in SDS PAGE using the buffer system of Laemmli (1970). The protein bands were then electrophoretically transferred to nitrocellulose paper according to the method of Towbin et al. (1979) and, after incubation with each cell line supernatant, the immune bands were revealed, using rabbit antimouse IgG coupled with peroxidase (Matus et al., 1980). Immunofluorescence Study of Heart Cells Indirect immunofluorescence was performed on transverse sections of frozen cryostat tissue samples from different regions of the human heart with appropriate dilutions of the antibody solutions, according to the method of Gorza et al. (1982). Atrial and ventricular sections were processed with fluorescein-labeled rabbit antimouse IgG specially prepared to avoid nonspecific cross-reaction (Nordic Immunologic Laboratories). The slides were mounted in a mixture consisting of an equal volume of PBS and glycerol. Sections were examined with a Leitz Orthoplan microscope equipped with epifluorescence optics. Controls for immunofluorescence included sections incubated with a cell culture supernatant either free of monoclonal antibody or containing a monoclonal antibody to renin (a generous gift from B. Pau). Enzyme Histochemistry of Heart Cells Serial sections adjacent to those used for immunofluorescence studies were stained for myofibrillar ATPase (myosin adenosine triphosphatase, calcium activated, EC 3.6.1.3) according to the method of Brooke and Kaiser (1970). The initial preincubation was made at pH 10.2, 10.4, 10.6, and 10.8 according to the method of Thornell and Forsgren (1982). The sections were examined under a Leitz Orthoplan photomicroscope. Results Antibody Characterization and Selection Fifteen stable hybridomas specific for human ventricular myosin were selected by direct RIA from two separate cell fusions; two hybridomas were generated from a CB 20 mouse, and the other 13 hybridomas were from a Biozzi HR mouse. Both immunizations used SDS-denatured myosin (Schwartz et al., 1980) prepared from the hypertrophic left ventricle of a 5 3-year-old female who had an obstructive cardiomyopathy. The 15 cell lines were cloned and their supernatants reacted only with the myosin heavy chains and not with the myosin light chains or other trace contaminants present in the whole muscle extract or in the myosin preparation as determined by the immune replicate technique (Fig. 2, a,a' and b,b') (Towbin et al., 1979). The specificity of the antibodies to myosin heavy chains was further tested by the reactivity of the two main chymotryptic fragments of the myosin heavy chains (e.g., rod and subfragment 1) (Fig. 2, c,c',c") and in some cases to smaller chymotryptic fragments of purified myosin rod (Fig. 2, d,d',d") or subfragment 1 (Klotz et al., 1981) to these antibodies. These different blotting experiments enabled us to verify that the antibodies were directed to the myosin heavy chain molecule, even though the myosin used as an immunogen contained trace contaminants. Most of the antibodies reacted differently with four different human myosins from the ventri- Downloaded from http://circres.ahajournals.org/ by guest on May 12, 2016 Bouvagnet et a/./Isomyosins in Human Atria and Ventricles MyHC- 797 MyHC - ROD —, SF1 -«» « Ach rv< Achn- — VLC1. VLC2a a' b b c d cle, atrium, skeletal muscle, and uterus as determined by competitive radioimmunoassay (data not shown). These competition assays and the assays of specificity to different chymotryptic myosin fragments (Fig. 2, d,d',d") suggest that most of the antibodies were presumably specific for different epitopes along the myosin heavy chain molecule. In a preliminary study, the 15 monoclonal antibodies were applied at different dilutions on serial cryostat sections of human atrial and ventricular tissue sections, to test their histoimmunological cross-reactivity with the cardiac fibers detected by fluorescein-labeled rabbit antimouse IgG. All 15 antibodies had a high response in terms of fluorescence level, and the response varied with the antibody dilution. Fiber typing was carried out on more than c' d' cd" FIGURE 2. a, b, c, d: SDS polyacrylamide gel electrophoreses (SDS PAGE) of different preparations of myosin or myosin fragments detected by Coomassie blue, and 2 a', b', c', c", d', d": corresponding immunoreplicates reacting with different hybridoma supematants and revealed using rabbit antimouse igC coupled with peroxidase. a: total muscle extract on 11% SDS PACE: MyHC, VLC1, VLC2 are the myosin heavy and light chains, respectively; a': immunoreplicate of a with a monoclonal antibody called 4F4. b: myosin on 11% SDS PACE; b': immunoreplicate of b with the 4F4 monoclonal antibody; c: myosin rod and subfragment 1 obtained after limited chymotryptic treatment of myofibrils on 14% SDS PAGE; c', c": immunoreplicate of c with 4F4 and another monoclonal antibody called 2C9, respectively; d: chymotryptic fragments of purified myosin rods on 14% SDS PAGE; d', d": immunoreplicates of d with 4F4 and 2C9 monoclonal antibodies, respectively. 50 tissue blocks coming from 10 different human atria and ventricles. The controls carried out with culture medium and labeled with antimouse IgG displayed a total absence of staining in all atrial and ventricular fibers (data not shown). Fiber Typing in Human Atria To determine whether any of the 15 available monoclonal antibodies would react differently with the hypothetically multiple molecular forms of myosin in human heart, tissue samples were initially excised from the free anterior wall of the right ventricle and of the anterior wall of the right atrium of different adult human hearts. A few of these muscle sections adjacent to those used for im- A A «* ^ »*®«M& rf «2> FIGURE 3. Three serial tissue sections from the anterior trabeculated wall of the right atrial human myocardium (sample 1 in Fig. 1) processed for indirect immunofluorescence with antiventricular human myosin antibodies (a) and (b) and for the histochemical staining of myosin ATPase activity after preincubation at pH 10.6 (c). Hybridomas which uniformly stained all atrial fibers (a) constituted the 1A group. The others, which preferentially stained certain fibers (b), constituted the 1A group. Arrowhead: an atrial fiber highly reactive with the 2A hybridoma group (b) and exhibiting weak ATPase activity after alkaline preincubation id). Arrow: an atrial fiber weakly reactive with the 24 hybridoma group (b) and exhibiting high ATPase activity after alkaline preincubation (d). Initial magnification, lOOx. Bar: 20 fim. Downloaded from http://circres.ahajournals.org/ by guest on May 12, 2016 Circulation Research/Vol. 55, No. 6, December 1984 798 TABLE 1 Correspondence of Fiber Types as Deduced from Indirect Immunofluorescence and Histochemical ATPase Staining of Serial Sections of Human Atrial Myocardium Atrial fiber type Immunofluorescence Hybridoma group: 1A 2A ATPase staining pH incubation: 10.6 A+ At Homogeneous Intense Weak Pale Dark munofluorescence studies were stained for histochemical determination of myofibrillar ATPase. Nine of the 15 hybridoma supernatants examined on atrial tissue sections reacted with the same affinity for all atrial fibers, and the immunofluorescence level observed varied according to the antibody dilution. This group of hybridomas, whose supernatants reacted homogeneously with all atrial fibers, was designated as 1A (Fig. 3a). The supernatants of the six other hybridomas,-at specific antibody dilutions, displayed a heterogeneous pattern of fluorescence reactivity (Fig. 3b): some fibers designated as A+ fibers were bright; others fibers designated as A— fibers were relatively dark, and one group of fibers showed various intermediate stainings. This group of hybridomas, whose supernatants reacted heterogeneously with the atrial fibers, was designated as 2A. By using serial sections, we were able to verify that all the 2A hybridoma supernatants consistently reacted with the same atrial fibers (A+ fibers), and conversely, not with the A— fibers. In order to compare the heterogeneity of human atrial fibers detected here by antimyosin antibodies to that previously reported in histochemical deter- minations of ATPase activities (Thornell and Forsgren, 1982), serial sections of the same atrial samples were incubated at pH 10.6 and then stained for ATPase activity. The atrial fibers which were poorly labeled immunologically with the 2A-hybridoma group (A— fibers), were always much more heavily stained for histochemical ATPase activity than the fibers which were highly labeled with the 2A hybridoma group (A+ fibers) (see arrow in Fig. 3, b and c, and arrowhead in Fig. 3, b and c, respectively). Fibers with intermediate stainings both in fluorescence and ATPase activity could be observed. Therefore, there is apparently a close correspondence between the immunological and histological methods for detecting heterogeneity in human atrium (see Table 1). Fiber Typing in Human Ventricles Nine of the 15 hybridoma supernatants studied stained all ventricular fibers homogeneously, but at variable immunofluorescence levels according to the antibody dilution used (Fig. 4a); this group of cell lines was designated as IV. The supernatants of the six other available hybridomas stained certain sparse fibers intensely (designated as V+ fibers) and reacted weakly and uniformly with the other ventricular fibers (designated as V— fibers) (Fig. 4b); some very sparse fibers were intermediately stained. This group of hybridomas was designated as 2V. The experimental controls in the absence of any hybridoma supernatants displayed a total absence of staining in all ventricular fibers (data not shown). Serial cryostat sections of ventricular samples adjacent to those used for immunofluorescence studies were incubated at different alkaline pH and stained for ATPase activity (Thornell and Forsgren, 1982). The few ventricular fibers which reacted intensely with the supernatants of the 2V-hybridoma group FIGURE 4. Two serial tissue sections from the free anterior wall of the right ventricle (sample 8 in Fig. 1) processed for indirect immunofluorescence with antiventricular human myosin antibodies: (a) and (b). Hybridomas which uniformly stained all ventricular fibers (a) constituted the IV group. The others, which preferentially stained certain sparse fibers (b), constituted the 2V group. Initial magnification, 40x. Bar: 50 pm. Downloaded from http://circres.ahajournals.org/ by guest on May 12, 2016 Bouvagnet et a/./Isomyosins in Human Atria and Ventricles 799 FIGURE 5. Three serial tissue sections from the anterior papillary muscle of the left ventricle (sample 16 in Fig. 1) processed for indirect immunofluorescence with a 2V hybridoma (a) and for ATPase activity after preincubation at pH 10.4 (b) and at pH 10.6 (c). Arrow: a group of three ventricular fibers whose ATPase activity was inhibited at both pH and which corresponds for immunofluorescence to V+ fiber type. Arrowhead: a fiber which exhibited a slightly higher ATPase activity at pH 10.6 but which reacted like other V- fibers for ATPase activity at pH 10.4 or for immunofluorescence. Initial magnification, 1OOX. (V+ fibers) showed no ATPase activity after alkaline preincubation of the tissue in the pH range between 10.2 and 10.8 (see arrow in Fig. 5, a-c). The other ventricular fibers which immunologically reacted weakly with the supernatants of the 2V hybridoma group (the V— fibers), exhibited homogeneous ATPase activity after preincubation of the tissue at pH 10.4 or below (Fig. 5b), but curiously exhibited heterogeneous ATPase activity after preincubation of the tissue at higher pH (Fig. 5c). Under the latter conditions, a few V— fibers were uniformly and highly stained for ATPase activity (arrowhead in Fig. 5c), whereas most of the V— fibers exhibited an intermediate stain for ATPase activity. These data are summarized in Table 2. Therefore, both the immunological and the histochemical method detect heterogeneity in human ventricle, but the close correspondence between the two experimental approaches does not exist as in the studies on human atrium. Comparative Fiber Typing in Human Atrium and Ventricle We then made a comparative study of the immunological reactivities of the supernatants of the 15 hybridomas on serial sections of ventricular fibers and atrial fibers, with both muscle sections applied to the same slide and with the supernatant at the same dilution. Two of the six hybridoma supernatants of the 2V group, which heavily stained a few ventricular fibers (V+ fibers in Fig. 4b), displayed a heterogeneous pattern of reactivity wih atrial fibers (Fig. 3b); this indicates that the labeled atrial fibers previously designated as A+ fibers differ not only from the unlabeled atrial A— fibers, but also from the unlabeled ventricular V— fibers. This also indi- cates that unlabeled atrial A - fibers differ from the labeled ventricular V+ fibers (Table 3). However, this hybridoma group, 2V(2A), does not support the evidence for a difference between atrial and ventricular labeled fibers (A+ vs. V+ fibers) and between atrial and ventricular unlabeled fibers (A— vs. V— fibers). The four other supernatants of the 2V hybridoma group uniformly and weakly stained all atrial fibers: this indicates that the atrial fibers previously reactive with the 2V(2A) hybridoma group (A+ fibers) were different from the more reactive group of ventricular fibers (V+ fibers). This hybridoma group was called 2V(1A). In contrast, four of the nine cell line supernatants which uniformly and intensely stained all ventricular fibers (Fig. 4a, IV hybridoma group) displayed a heterogeneous pattern of fluorescence reactivity in atrium (Fig. 3b). This indicates that the unreactive atrial A— fibers were different from all ventricular fibers which were reactive with this 1V(2A) hybridoma group (V— or V+ fibers). The supernatants of the last five hybridomas of the set uniformly and intensely stained TABLE 2 Correspondence of Fiber Types as Deduced from Indirect Immunofluorescence and Histochemical ATPase Staining of Serial Sections of Human Ventricular Myocardium Ventricular fiber type Immunofluorescence Hybridoma group: IV 2V ATPase staining pH incubation: 10.6 Ve V- Homogeneous Intense Weak Pale Interm/Dark Some fibers displayed an intermediate (interm.) reactivity. Downloaded from http://circres.ahajournals.org/ by guest on May 12, 2016 Circulation Research/Vo/. 55, No. 6, December 1984 800 TABLE 3 Classification of the Set of the Hybridomas Relative to Their Reactivity for Ventricular and Atrial Fibers Hybridoma groups Immunofluorescence fiber response ntric Ventricles Name No. V+ V- A+ A- 2V (2A) 2V (1A) IV (2A) IV (1A) 2 4 4 5 Intense Intense Intense Intense Weak Weak Intense Intense Intense Weak Intense Intense Weak Weak Weak Intense the atrial and ventricular fibers 1V(1A) and possiblycorresponded to epitopes common to all human cardiac myoxins. To summarize, human atrium, as well as human ventricle, is composed of at least two different antigenic types of muscle fiber, both specific for each cardiac cavity. Anatomical Distribution of Human Heart Fibers Immunofluorescence microscopy was further performed on 17 different regions of the hearts of two 16-year-old boys, to determine the fiber distribution and, consequently, the cellular distribution of the multiple molecular forms of atrial and ventricular myosin recognized by the antibodies of the 2A and 2V hybridoma groups, respectively. About 100 serial 7-/im-thick frozen sections per tissue block were examined. The most frequent pattern of reactivity observed in all atrial samples treated with the supematants of the 2A hybridoma group is shown on Fig. 6a: Many atrial fibers were either intensely (i.e., the A+ fibers) or weakly (i.e., the A— fibers) stained, whereas many others were intermediately stained. All the tissue sections of the atrial samples which were excised from one region of postulated conduction pathways (12) or from common atrial myocardium (4 and 11) contained only a half-and-half mixture of the A+ and A— fibers. In a few longitudinally oriented bundles, A+ and A— fibers (reactive or unreactive, respectively, with antibodies from the 2A hybridoma group) were observed near each other (Fig. 6b): this indicates the apparently discontinuous arrangement of the different types of atrial fibers. The four other atrial samples examined contained half FIGURE 6. Tissue sections from four different regions of human atrial myocardium processed for indirect immunofluorescence with the supematants of the 1A hybridoma group, a: transverse section of the anterior wall of the RA (no. 1 in Fig. 1), b: longitudinal section of the interatrial septum (no. 4 in Fig. 1), c: transverse section of the subendocardial area of the left auricle (no. 13 in Fig. 1), d: transverse section of the subendocardial area of the crista terminalis (no. 3 in Fig. 1). Note that the fiber patterns in (a) and (b) are observed in all atrial tissue samples, whereas the fiber patterns in (c) and (d) are observed in only a few sections of specifically located atrial samples. Initial magnification: a, c, d, 40x. Bar: 50urn;b, 100X. Bar: 20 pm. Downloaded from http://circres.ahajournals.org/ by guest on May 12, 2016 Bouvagnet et a/./Isomyosins in Human Atria and Ventricles of the tissue sections with the equilibrated mixture of heterogeneously reactive fibers described above; the other half of the tissue sections corresponded to two opposite situations in terms of the regional distribution of fiber types. One situation, which corresponds to the presence of a majority of weakly reactive A— fibers with a few highly reactive A+ fibers (Fig. 6c), was observed in both auricles (samples 2 and 13). The other situation, which corresponds to the presence of a majority of highly reactive A+ fibers, was observed only in the subendocardial area of the right atrium (samples 1 and 3, Fig. 6d). However, samples 3 and 12, which were both excised from regions of a postulated conduction pathway displayed a different fiber pattern. The same regional distribution of the different atrial fibers was observed in both hearts. From this study of about two thousand sections from seven different atrial regions in two normal human hearts, it can be concluded that human atrium contains two different fiber types which are invariably located and exist in roughly the same proportion. The most frequent pattern of reactivity observed in all ventricular samples treated with the supernatants of the 2V hybridoma group is shown in Figure 7a. Most of the ventricular fibers reacted weakly but uniformly with the antimyosin antibodies, and only a few fibers were highly reactive. The most frequent pattern of ventricular fibers therefore consists of a majority of V— fibers with a small amount of the V+ fibers, ranging from approximately 2 to 5% of all fibers. This regional distribution of fiber types was observed in all ventricular sections from samples 6, 8, 14, 15, and 16, and on some sections of samples 5 and 7. The number of rare, highly reactive V+ fibers increased significantly in some sections from sample 5, which contained a maximum proportion of about 15% V+ fibers. However, this 801 increase in V+ fibers was not observed in sample 6 which, like sample 5, was excised from a region of postulated conduction pathways. On the few sections from sample 5, where the V+ were numerous, they were arranged in bundles (Fig. 7b). Conversely, these highly reactive V+ fibers were totally absent in most sections from samples excised from the apex (10 and 17) and from the conus arteriosus (9). In terms of the myosin distribution, no differences were observed between endocardial and epicardial areas. The same regional variation in the distribution of the V+ and V— fibers was observed in both hearts. On the basis of this study of about 3500 sections from 10 different ventricular regions in two human hearts, it can be concluded that human ventricle contains two different fiber types but that the amount of the V+ type fibers varies roughly between zero to 15% of the total fibers, depending on the location of the ventricular tissue. Discussion The present study shows that human atrium and human ventricle each contain muscle fibers that differ in their antigenic reactivity with a set of myosin heavy chain specific antibodies prepared from an enlarged human ventricle. On the basis of these observations, we can assume that at least two antigenic types of myosin are present in the ventricle, as well as in the atrium. Since different hybridoma supernatants are able to detect these antigenic differences, and since the epitopes of the immunoglobulins used are located at different positions along the myosin chains, it is likely that the two (or more) atrial myosins and the two (or more) ventricular myosins detected each have different primary structures. In addition, since the set of antimyosin hybridomas can be grouped into four subclasses according to the unique or dual specific reactivity of their FIGURE 7. Tissue sections from two different regions of the human ventricular myocardium processed for indirect immunofluorescence with the supernatants of the 2V hybridoma group, a: transverse section of the subendocardial region of the mid-portion of the posterior left ventricular wall (no. 25 in Fig. 1), b: oblique section of the region below the supraventricular crest of the RV (no. 5 in Fig. 1). Note that the fiber pattern in (a) is observed in almost all tissue samples, whereas the fiber pattern in (b) is observed in only a few sections of specifically located ventricular samples. Initial magnification, 40x. Bar: 50 pm. Downloaded from http://circres.ahajournals.org/ by guest on May 12, 2016 Circulation Research/Vo/. 55, No. 6, December 1984 802 FIGURE 8. Tissue sections from the left ventricle of a normal young rat (a and b) and of a hypothyroid rat (c and d) processed for indirect immunofluorescence with 2V antimyosin antibodies. Some 2V antibodies reacted only with the V3-type of rat ventricular myosin (a) and (c); other 2V antibodies reacted in a single way with the VI- and V3-types of rat ventricular myosin (b). supernatants with the atrial or ventricular fibers (Table 3), it can be assumed that the two human ventricular myosins (corresponding to the V+ and V— fibers) are molecularly different from the two human atrial myosins (corresponding to the A+ and A— fibers) (Table 3). In general, our immunological results support the view that human atrium and human ventricle each contain at least two molecular variants of myosin heavy chains, as do rat, rabbit, and beef atria and the ventricles of many animals (Hoh et al., 1978; Flink and Morkin, 1979; Klotz et al., 1981; Lompre et al. 1981; Chizzonite et al., 1981; Sartore et al., 1982; Gorza et al., 1982; Litten et al., 1982; Banerjee, 1983). Heterogeneity of cardiac fibers with respect to myosin composition consequently appears to be a general property of mammalian hearts, regardless of the size of the animals. The heterogeneous reaction of the fibers to ATPase after alkaline preincubation within human atrium was also demonstrated in beef atrium by Gorza et al. (1982), but, curiously, these results differ from those of Thornell et al. (1982), who detected a heterogeneous ATPase reaction within human atrium only after acid preincubation. Reservations are therefore required in any interpretation of the ATPase data. By comparing the histoenzymatic response to the immunological typing of the same bovine atrial fibers, Gorza et al. (1982) have observed that fibers which were ATPase resistant at alkaline pH, cross-reacted with anti-beef atrium (or anti-Vl or anti-myosin heavy chain a) serum, and that the other minor atrial fibers which lost their ATPase activity at alkaline pH, cross-reacted with anti-beef ventricle (or anti-V3 or anti-myosin heavy chain /3) serum. Because of their similar histochemical behavior, it is tempting to suggest that human atrial A— fibers with an alkaline stable ATPase activity are structurally related to a-like myosin heavy chains and that human A+ fibers with an alkaline labile ATPase activity are related to /3-like myosin heavy chain; the set of antibodies used in the present work would then react either with V3type myosin heavy chains (2A-hybridoma group) or with VI- and V3-type myosin heavy chains (1Ahybridoma group), and none of the antibodies would react with VI-like myosin heavy chains alone. The presence of two different molecular variants of human atrial myosin, each having a structural relationship with one of the two main types of ventricular myosin, but all of which are different (see Results and Table 3), is in accordance with the results of Clark et al. (1982) which showed the simultaneous presence of VI-type and V3-type epitopes in atrial myosins from chicken, pig, and beef. The patterns of alkaline pH sensitivity of the histochemical ATPases of all human ventricular V+ fibers and of most of the V— fibers are rather similar to those observed within the ventricles of beef and hypothyroid rats, which lose their ATPase activity above pH 10.4 and contain mainly the V3-type of ventricular myosin (Sartore et al., 1981; Weisberg et al., 1983). The other very rare human V— fibers which still have an ATPase activity at pH 10.6 (Fig. 5c), behave histochemically like the ventricular fibers of euthyroid rats which contain mainly the VItype of ventricular myosin. The finding that none of the 15 monoclonal antibodies used here would react with any human fiber containing only VI-type myosins, was further tested on ventricular fibers prepared from a 1-month-old euthyroid rat and from a hypothyroid rat. We thus observed that some of our 2V hybridoma stained intensely certain fibers of the left ventricle of the normal rat (Fig. 8a) and all fibers of the left ventricle of the hypothyroid rat Downloaded from http://circres.ahajournals.org/ by guest on May 12, 2016 Bouvagnet et a/./Isomyosins in Human Atria and Ventricles (Fig. 8c), assessing that these monoclonal antibodies reacted with epitopes present only in the V3-type of rat ventricular myosin. We also observed that the other 2V hybridomas and all IV hybridomas reacted more or less intensely with the ventricular fibers of both rats, but in a single way, suggesting that these monoclonal antibodies reacted with epitopes common to VI- and V3-types of ventricular myosin (Fig. 8, b and d). We also confirmed the histochemical observations made by Weisberg et al. (1983) and the relationship between immunological and histochemical fiber typings. Rare Vl-type myosins already have been detected in human ventricles by Thornell et al. (1982), using histochemistry, and by Schiaffino et al. (1983) and Mercadier et al. (1983), using monoclonal and polyclonal antibodies specific to the rat and beef Vl-type myosins. The probable absence of anti-Vl monoclonal antibodies in our hybridoma set could be an unexpected consequence of the initial choice of an antigen which did not in fact contain this Vl-type myosin (Mercadier et al., 1983). On the basis of the histochemical and immunological information currently available, we tentatively conclude that human ventricle contains very few Vl-type myosins (presumably, the V— fibers with an alkaline-stable ATPase activity) and two molecular variants of V3-type myosins, one being relatively rare (the V+ fibers with an alkalinelabile ATPase activity), and the other representing 90-95% of the total myosin amount (the V— fibers with an alkaline-labile ATPase activity). A presence of two forms of V3-type myosin in rat ventricles has already been suggested by Watras (1981). More recently, Mahdavi et al. (1983) have reported a similar heterogeneity involving two different primary amino acid sequences within the Vl-type myosins in the rat ventricle. The most interesting result concerning the anatomical distribution of human heart fibers reported here is the observation of small zones of myosin variation scattered but not randomly distributed within a large area of myocardium whose cellular distribution of myosin is constant. The subendocardial region of the crista terminalis and the anterior trabeculated wall of the right atrium are rich in highly immunoreactive myosins, whereas both auricles are rich in nonimmunoreactive myosins. These regional variations in myosin distribution could perhaps be related to the tension of the different cardiac walls as the result of a process of adaptation to the same internal pressure (Sandier and Dogge, 1963). Our results in human atria were not the same as those obtained by Gorza et al. (1982), who have suggested that certain typical patterns of muscle fiber in beef atrial myocardium may be specialized for faster conduction. The origin of the discrepancy is still unknown, but it is probably related to the postulated existence of specialized conduction pathways in atrial human myocardium (Anderson et al., 1981). In the ventricle, a distribution gradient of the small zones of variation in myosin distribution ap- 803 pears to exist within normal myocardium: highly immunoreactive myosins are more abundant near the auriculoventricular valves than in the midportion of the ventricular wall or septum, and are nearly absent in the apex and conus arteriosus. No significant differences in the regional distribution of different myosin types between the left and right ventricle have been detected, with our approach, as have been found in beef ventricles (Sartore et al., 1981). However, it should be emphasized that our anatomical study was limited to whole normal hearts of two young boys, and that the antibodies used probably react only with V3-type myosins. The regional distribution of V— fibers with an alkalinestable ATPase activity is thus not described by the present results. Our anatomical study is currently being extended to a variety of other whole human hearts in different physiological or pathophysiological states, in order to detect any possible variation in their myosin distributions (Schier and Adelstein, 1982; Mercadier et al, 1983; Leger et al., 1983). We wish to thank Henri Haton for his excellent technical assistance, Dr. G. Biozzi for the kind gift ofH. R. mice, and Dr. ]. Demaille for valuable discussion. We also thank Drs. L Gorza, S. Sartore, and S. Schiaffino for their assistance in establishing optimal conditions for indirect immunofluorescence. We are grateful to Dr. B. Pau for his assistance in introducing us to hybridoma technology, and for the support given by the "Service d'lmmunodiagnostic" of the Clin Midy firm. This work was supported in part by the "Ministere de I'lndustrie et de la Recherche" (DGRST81J0260), by TInstitut National de la Sante et de la Recherche Medicate" (CRL826030), and by la Fondation de la Recherche Medkale. Address for reprints: /. /. Leger, INSERM U249, Institut de Biologie, Boulevard Henri IV, 34000 Montpellier, France. Received September 9, 1983; accepted for publication August 22, 1984. References Anderson RH, Becker AE, Tranum-Jensen J, Janse MJ (1981) Anatomico-electrophysiological correlations in the conduction system: A review. Br Heart J 45: 67-82 Bader D, Masaki T, Fischman DA (1982) Immunochemical analysis of myosin heavy chain during avian myogenesis in vivo and in vitro. J Cell Biol 95: 763-770 Banerjee SK (1983) Comparative studies of atrial and ventricular myosin from normal, thyrotoxic, and thyroidectomized rabbits. Circ Res 52: 131-136 Brooke MH, Kaiser KK (1970) Muscle fiber types: how many and what kind? Arch Neurol 23: 369-379 Chizzonite RA, Everett AW, Clark WA, Jakovcic S, Rabinowitz M, Zak R (1982) Isolation and characterization of two molecular variants of myosin heavy chain from rabbit ventricle. J Biol Chem 257: 2056-2065 Clark WA, Chizzonite RA, Everett AW, Rabinowitz M, Zak R (1982) Species correlations between cardiac isomyosins. A comparison of electrophoretic and immunological properties. J Biol Chem 257: 5449-5454 Cummins P (1982) Transitions in human atrial and ventricular myosin light chain isoenzymes in response to cardiac pressure overload induced hypertrophy. Biochem J 205: 195-204 Di Pauli R, Raschke WC (1978) A hybridoma secreting IgM anti Iglb allotype. In Current Topics in Microbiology and Immunology 81, edited by F Melchers, M Potter, NL Warner. Berlin, Springer-Verlag, pp 37-39 Downloaded from http://circres.ahajournals.org/ by guest on May 12, 2016 804 Circulation Research/Vol. 55, No. 6, December 1984 Flink IL, Morkin E (1979) Thyroid hormone stimulates synthesis of a cardiac myosin isozyme. J Biol Chem 254: 3105-3110 Gorza L, Sartore S, Schiaffino S (1982) Myosin types and fiber types in cardiac muscle. II: Atrial myocardium. J Cell Biol 95: 838-845 Hoh JFY, McGrath PA, Hale PT (1978) Electrophoretic analysis of multiple forms of rat cardiac myosin: effects of hypophysectomy and thyroxine replacement. J Mol Cell Cardiol 10: 10531076 James TN, Sherf L (1971) Specialized tissues and preferential conduction in the atria of the heart. Am J Cardiol 28: 414-426 Klotz C Swynghedauw B, Mendes H, Marotte F, Leger JJ (1981) Evidence for new forms of cardiac myosin heavy chains in mechanical heart overloading and in ageing. Eur J Biochem 115: 415-421 Klotz C, Leger JJ, Elzinga M (1982) Comparative sequence of myosin light chains from normal and hypertrophied human hearts. Circ Res 50: 201-209 Klotz C, Dechesne C, Cardinaud P, Leger JJ (1983) Structural differences between atrial and ventricular myosins from normal human hearts. Biochimie 65: 569-578 Laemmli UK (1970) Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature (Lond) 227: 680-685 Leger Joe, Klotz C, Leger JJ (1983) Cardiac myosin heavy chains and rropomyosin in mechanical heart overloading and ageing. In Perspectives in Cardiovascular Research: Myocardial Hypertrophy and Failure, vol 7, edited by NR Alpert. New York, Raven Press, pp 385-392 Litten RZ III, Martin BJ, Low RB, Alpert NR (1982) Altered myosin isozyme patterns from pressure-overloaded and thyrotoxic hypertrophied rabbits hearts. Circ Res 50: 856-864 Lompre AM, Mercadier JJ, Wisnewski C, Bouveret P, Pantaloni C, D'Albis A, Schwartz K (1981) Species- and age-dependent changes in the relative amounts of cardiac myosin isoenzymes in mammals. Dev Biol 84: 286-290 Mahdavi V, Periasamy M, Nadal-Ginard B (1982) Molecular characterization of two myosin heavy chain genes expressed in the adult heart. Nature 297: 659-665 Matus A, Pehling G, Ackermann M, Maeder J (1980) Brain postsynaptic densities: Their relationship to glial and neuronal filaments. J Cell Biol 87: 346-351 Mercadier JJ, Bouveret P, Gorza L, Schiaffino S, Clark WA, Zak R, Swynghedauw B, Schwartz K (1983) Myosin isozymes in normal and hypertrophied human ventricular myocardium. Circ Res 53: 52-62 Miller DM, Ortiz I, Berliner GC, Epstein HF (1983) Differential localization of two myosins within nematode thick filaments. Cell 34: 477-490 Netter FH (1974) A compilation of paintings on the normal and pathologic anatomy and physiology, embryology, and diseases of the heart. In The Ciba Collection of Medical Illustrations, vol 5, edited by FF Yonkman. New Jersey, CIBA, pp 9-13 Rupp H (1982) Polymorphic myosin as the common determinant of myofibrillar ATPase in different haemodynamic and thyroid states. Basic Res Cardiol 77: 34-46 Sandier H, Dogge HT (1963) Left ventricular tension and stress in man. Circ Res 13: 91-96 Sartore S, Gorza L, Pierobon Bormioli S, Dalla Libera L, Schiaffino S (1981) Myosin types and fiber types in cardiac muscle. I: Ventricular myocardium. J Cell Biol 88: 226-233 Schiaffino S, Gorza L, Sartore S (1983) Distribution of myosin types in normal and hypertrophic hearts: an immunocytochemical approach. In Perspectives in Cardiovascular Research: Myocardial Hypertrophy and Failure, vol 7, edited by NR Alpert. New York, Raven Press, pp 149-166 Schier JJ, Adelstein RS (1982) Structural and enzymatic comparison of human cardiac muscle myosins isolated from infants, adults, and patients with hypertrophic cardiomyopathy. J Clin Invest 69: 816-825 Schwartz K, Lompre AM, Bouveret P, Wisnewsky C, Swynghedauw B (1980) Use of antibodies against dodecyl sulfate denatured heavy meromyosins to probe structural differences between muscular myosin isoenzymes. Eur J Biochem 104:341 346 Schwartz K, Lecarpentier Y, Martin JL, Lompre AM, Mercadier JJ, Swynghedauw B (1981) Myosin isoenzymic distribution correlates with speed of myocardial contraction. J Mol Cell Cardiol 13: 1071-1075 Swynghedauw B, Delcayre C (1982) Biology of cardiac overload. Pathobiol Annual 12: 137-183 Thornell LE, Forsgren S (19820 Myocardial cell heterogeneity in the human heart with respect to myosin ATPase activity. Histochem J 14: 479-490 Towbin H, Staehelin T, Gordon J (1979) Electrophoretic transfer of proteins from polyacrylamide gels to nitrocellulose sheets: Procedure and some applications. Proc Natl Acad Sci USA 76: 4350-4354 Tuchschmid CR, Srihari T, Hirzel HO, Schaub MC (1983) Structural variants of heavy and light chains of atrial and ventricular myosins in hypertrophied human hearts. In Cardiac Adaptation to Hemodynamic Overload, Training and Stress. Erwin Riesch Symposium, Tubingen, Darmstadt, edited by D Steinkopff Verlag, pp 123-128 Wachsberger P, Lampson L, Pepe FA (1983) Non-uniform staining of myofibril A bands by a monoclonal antibody to skeletal muscle myosin SI heavy chain. Tissue Cell 15: 341-349 Watras J (1981) Changes in rat cardiac myosin during development and in culture. J Mol Cell Cardiol 13: 1011-1021 Weisberg A, Winegrad S, Tucker M, McClellan G (1982) Histochemical detection of specific isozymes of myosin in rat ventricular cells. Circ Res 51: 802-809 Winckelmann DA, Lowey S, Press JL (1983) Monoclonal antibodies localize changes on myosin heavy chain isozymes during avian myogenesis. Cell 34: 295-306 INDEX TERMS: Anatomy • Human • Hybridoma • Immunofluorescence • Myosin • Myocardium Downloaded from http://circres.ahajournals.org/ by guest on May 12, 2016 Fiber types and myosin types in human atrial and ventricular myocardium. An anatomical description. P Bouvagnet, J Leger, F Pons, C Dechesne and J J Leger Circ Res. 1984;55:794-804 doi: 10.1161/01.RES.55.6.794 Circulation Research is published by the American Heart Association, 7272 Greenville Avenue, Dallas, TX 75231 Copyright © 1984 American Heart Association, Inc. All rights reserved. Print ISSN: 0009-7330. Online ISSN: 1524-4571 The online version of this article, along with updated information and services, is located on the World Wide Web at: http://circres.ahajournals.org/content/55/6/794 Permissions: Requests for permissions to reproduce figures, tables, or portions of articles originally published in Circulation Research can be obtained via RightsLink, a service of the Copyright Clearance Center, not the Editorial Office. Once the online version of the published article for which permission is being requested is located, click Request Permissions in the middle column of the Web page under Services. Further information about this process is available in the Permissions and Rights Question and Answer document. Reprints: Information about reprints can be found online at: http://www.lww.com/reprints Subscriptions: Information about subscribing to Circulation Research is online at: http://circres.ahajournals.org//subscriptions/ Downloaded from http://circres.ahajournals.org/ by guest on May 12, 2016