Survey
* Your assessment is very important for improving the workof artificial intelligence, which forms the content of this project
1123
Variation of Perisylvian and
Calcarine Anatomic Landmarks
Within .S tereotaxic Proportional
Coordinates
Helmuth Steinmetz 1
Gunter FOrst2
Hans-Joachim Freund 1
This investigation describes the variability in location of functionally important perisylvian landmarks and of the calcarine sulcus within the Talairach stereotaxic grid, a
system frequently used for cortical localization in functional images. Twenty healthy
volunteers (40 hemispheres) had MR imaging under stereotaxic conditions. Outlines of
the following structures were directly identified on sagittal 5-mm MR sections and
marked on individual proportional grid overlays: inferior central sulcus, inferior precentral
sulcus, inferior postcentral sulcus, anterior ascending ramus and posterior rami of the
sylvian fissure, superior temporal sulcus, and calcarine sulcus. Maximal variation zones
for these landmarks were defined by superimposition of the standardized individual
data on a standard stereotaxic grid. The sulcal variation zones measured 1.5-2.0 em.
The findings indicate that macroanatomic individuality in the cerebral surface cannot
be accounted for adequately by proportional coordinates, and that this method does not
allow precise definition of anatomically based regions of interest for functional imaging.
Instead, MR mapping of the individual sulcus pattern should be used to generate brain
templates.
AJNR 11:1123-1130, November/December 1990
Received March 12, 1990; revision requested
June 5, 1990; revision received July 2, 1990; accepted July 4, 1990.
This work was supported by grants from the
Deutsche Forschungsgemeinschaft (SFB 200fZ2).
' Department of Neurology, Heinrich-Heine-University, Moorenstr. 5, D-4000 Dusseldorf, F.R.G .
Address reprint requests to H. Steinmetz.
The current revival of stereotaxic brain imaging reflects the need for a precise
topographic analysis of structural and functional image data. The Talairach stereotaxic proportional grid was originally developed for pneumoencephalography [1]
and has received renewed attention since the discovery that MR can provide
noninvasive application [2-5]. The anatomic atlas of Talairach et al. [1] is based
on this grid system and has recently been used by several investigators for
interpretation of CT [6] , MR [3 , 4], and positron emission tomography (PET) [716]. This interest prompted the publication of a new brain atlas by Talairach and
Tournoux [17], which , according to the preface, was "derived from one particular
[postmortem] brain, applicable to all other brains under examination ." Drawings of
contiguous frontal , coronal, and sagittal sections of this "brain of reference" were
displayed with the corresponding grid overlays. Unlike the original work of Talairach
et al. [1], interindividual variability of cortical structures was not assessed quantitatively in the new atlas. The problem of morphologic individuality, however, is
relevant for any atlas-based interpretation of image data. Variability of functionally
important perisylvian landmarks (Figs. 1 and 2) and of the calcarine sulcus within
the Talairach proportional grid is described in this article, which continues a previous
study on the variation of superior hemispheric sulci and of the sylvian fissure [5] .
On the basis of the results, the suitability of the proportional grid for image analysis
is discussed .
2
Department of Diagnostic Radiology, HeinrichHeine-University, D-4000 Dusseldorf, F.R.G.
Subjects and Methods
0195-6108/90/ 1106-11 23
© American Society of Neuroradiology
Forty cerebral hemispheres were evaluated in 20 healthy volunteers comprisi ng eight
women and 12 men 22-63 years old . Nineteen probands considered themselves right-
1124
STEINMETZ ET AL.
AJNR :11 , November/ December 1990
Key to Abbreviations Used in Figures
AAR
AGPG
Fig. 1.-Standard sulcal arrangement of lateral hemispheric aspect. See
key for abbreviations. (Rendering adapted from Brodmann (18].)
10
•
Fig. 2.-Base planes of proportional grid as applied to left (L) and right
(R) hemispheres of two different-sized adult brains show consistent location of sulcus centra lis within reference system. See key for abbreviations.
Locations of precentral sulcus (open arrows) and superior temporal sulcus
(solid arrows) show perisylvian variability, even within proportional frame.
Both sulci are classical landmarks of functionally important regions (precentral sulcus is posterior border of Broca's area and superior temporal
sulcus is inferior border of Wernicke's area). Anterior ascending ramus is
the sulcus anterior to the precentral sulcus in both hemispheres. (Reprinted
with permission from Talairach and Tournoux [17).)
anterior ascending ramus of sylvian fissure
bicommissural plane that runs tangent to upper margin of anterior commissure and to
lower margin of posterior commissure
AHR
anterior horizontal ramus of sylvian fissure
same as AGPG
GaGp
GALGS calcarine sulcus
central sulcus
GS
inferior frontal sulcus
IFS
intraparietal sulcus
IPS
marginal sulcus (terminal ascending ramus
MS
of cingulate sulcus)
posterior ascending ramus of sylvian fissure
PAR
posterior descending ramus of sylvian fisPDR
sure
posterior horizontal ramus of sylvian fissure
PHR
postcentral sulcus
POGS
PRGS
precentral sulcus
sylvian fissure
SF
STS
superior temporal sulcus
VAG
plane perpendicular to bicommissural plane
and to midsagittal plane and tangent to
posterior margin of anterior commissure
same as VAG
Vca
Vcp
same as VPG
plane perpendicular to bicommissural plane
VPG
and to midsagittal plane and tangent to
anterior margin of posterior commissure
handed . Methodologic principles of the proportional grid system were
described in a previous study of 15 probands [5) who were also
included in this investigation . Following orthomorphic alignment of
the brain (Fig. 3A), sagittal T1-weighted spin-echo (SE) images (Figs.
3B-3F) were obtained from the whole brain with the use of a 0.35-T
superconductive magnet. The technical factors were 500/40/4 (TR/
TEfexcitations), 256 x 256 image matrix, and 5-mm-thick contiguous
slices. For each of the 40 hemispheres an individual proportional grid
was constructed on a transparent overlay on the sagittal images
(Figs. 3B-3F). Identical orientation of the overlay on images not in
the midplane was facilitated by the scanner's millimetric grid on the
images serving as a fixed ·:external" reference (the grid is not displayed in Figs . 3B-3F). Image distortions within the field of interest
were excluded , as also described previously [5). For the current study
the following sulci were identified directly on each hemisphere and
their course was marked on the overlay (see below for anatomic
criteria): inferior central sulcus , inferior precentral sulcus, inferior
postcentral sulcus , anterior ascending sylvian ramus , posterior horizontal sylvian ramus , posterior ascending sylvian ramus , and horizontal segment of the superior temporal sulcus (see Fig . 1 for
"standard " perisylvian topography). The calcarine sulcus was outlined
on the medial hemispheric surface. The callosal sulcus was determined on the midsagittal section. Except for the callosal sulcus all
sulci were evaluated separately for left and right hemispheres .
Inferior Central, Precentral, and Postcentral Sulci
The inferior central and pre- and postcentral sulci were identified
directly by using the following procedure [19) : The central sulcus was
AJNR :11, November/December 1990
STEREOTAXY OF CORTICAL LANDMARKS
identified at the superior hemispheric margin , where it is constantly
found as the next sulcus anterior to the marginal sulcus (5 , 19-21].
From there, the central sulcus takes an unbroken course (21 , 22]
toward its inferior end. By using a real-time cinematographic display
mode on an off-line image-processing workstation (Mipron, Kontron
Bildanalyse, Eching , F.R.G.), we traced the bottom of the central
sulcus progressing from medial to lateral MR slices. The course of
the inferior central sulcus on the external hemispheric surface as
determined from the two or three most lateral sagittal images of each
hemipshere was marked on the individual grid overlay. The next sulci
running anterior and posterior to the inferior central sulcus were
identified and marked as the inferior pre- and postcentral sulci,
respectively (Figs . 3C-3E).
Anterior Ascending, Posterior Horizontal, and Posterior Ascending
Sylvian Rami and Superior Temporal Sulcus
The sylvian fissure is classically divided into five segments
[21 , 23] . These are the main horizontal stem (posterior horizontal
ramus) and two anterior (anterior ascending and anterior horizontal
rami) and two posterior (posterior ascending and posterior descending rami) branches (see Fig . 1). The anterior and posterior ascending
rami constitute functionally important landmarks. The anterior ascending ramus divides the posterior inferior frontal gyrus into the pars
opercularis (between precentral sulcus and anterior ascending ramus)
and pars triangularis (between anterior horizontal ramus and anterior
ascending ramus). The anterior ascending ramus is thus centered in
the anterior speech area on the dominant hemisphere. The posterior
ascending ramus cuts into the supramarginal gyrus, which is part of
the inferior parietal lobule (and posterior speech region). The anterior
ascending ramus was directly identified with MR by using the following criteria (19]. The anterior ascending ramus originates in the depth
of the insular cistern , where it is continuous with the so-called circular
(periinsular) sulcus. From there, the anterior ascending ramus was
traced laterally on the 5-mm sagittal MR sections by using the
aforementioned fast display mode. The anterior ascending ramus
cuts through the whole depth of the frontal operculum and thus can
be discerned from other anterior perisylvian sulci. The posterior
ascending ramus was identified as the terminal , upswinging portion
of the posterior sylvian fissure (Fig . 1). The posterior horizontal ramus
was outlined between the points of origin of the anterior and posterior
ascending rami. The superior temporal sulcus was identified as the
next sulcus running inferior and parallel to the posterior horizontal
ramus (Figs. 3D-3F).
1125
Results
Inferior Central, Precentral, and Postcentral Sulci
The variation zones of the right and left inferior central and
pre- and postcentral sulci measured between 1.5 and 2 em
in the anteroposterior axis (Figs. 4 and 5). As gyral widths
were of the same order, only minimal overlap occurred between the sulcal zones. The location and variation of the
inferior central sulci corresponded to that derived by Talairach
et al. [1] from their study of 20 postmortem hemispheres. No
clear left-right asymmetries were seen for the central or
precentral sulcus. The inferior postcentral sulcus was more
variable and tended to be located farther posteriorly on the
left than contralaterally (Fig. 5). The inferior pre- and postcentral sulci had not been investigated by Talairach et al. [1 ].
Sylvian Rami and Superior Temporal Sulcus
The vertical variation of the posterior horizontal ramus and
superior temporal sulcus and the anteroposterior variation of
the anterior and posterior ascending rami also ranged between 1.5 and 2 em (Figs. 4 and 6). The variation zone of the
posterior horizontal ramus extended more posteriorly in the
left than in the right hemispheres, thus producing a larger left
parietal operculum in the majority of cases. The latter finding
agrees with several anatomic reports [21, 23, 26-31 ]. Also,
distribution of the posterior end points of the posterior horizontal ramus was more scattered on the left than contralaterally, so that the location of the posterior ascending ramus
was more variable among left hemipsheres (Fig . 6). No leftright asymmetries were observed for the anterior ascending
ramus and superior temporal sulcus.
In six right hemispheres, a direct transition of the posterior
ascending ramus into the inferior postcentral sulcus was seen ,
which explains the considerable overlap between these variation zones (compare Figs. 5A and 6A). In five left hemispheres, a posterior ascending ramus could not be identified,
which explains the posterior extension of the posterior horizontal ramus zone beyond that of the posterior ascending
ramus in Figure 68.
Calcarine and Callosal Sulci
Calcarine Sulcus
The calcarine sulcus is centered in the striate cortex (18, 24] and
originates about 2 em posterior to the splenium of the corpus callosum from a common stem with the inferior parietooccipital sulcus
[20, 23]. From there, the calcarine sulcus takes a slightly curved
course toward the occipital pole. It has a sulcal depth of 14-20 mm
[20, 25]. Accordingly, the calcarine sulcus was identified and marked
on immediate paramedian MR sections (Fig. 3C). The callosal sulcus
was determined as the outer r:nargin of the corpus callosum on the
midsagittal image.
The variation zone of the calcarine sulcus measured almost
2 em in the vertical axis {Fig. 7). It corresponded exactly to
that indicated by Talairach et al. [1] in their pneumoencephalographic study of 30 hemispheres. There were no left-right
asymmetries.
Standardization Procedure
All sulcal markings on the individual overlays referred to midpoints
between opposed gyral lips at the external hemispheric surface. To
obtain statistical graphic summations of the individual data, all overlays were magnified proportionally and projected stepwise, rectangle
by rectangle , on a standard grid (Figs. 4-7).
Discussion
The distinction between direct and indirect techniques of
cerebral localization is fundamental to brain stereotaxy
[1 , 17]. Direct localization means recognition of characteristic
cerebral landmarks. Indirect localization , such as with the
proportional grid, relies on coordinate systems constructed in
reference to (directly identified) landmarks. Errors are inherent
in any indirect method owing to interindividual variability of
the spatial relation between targets and references . Direct
STEINMETZ ET AL.
1126
AJNR :11 , November/ December 1990
A
D
E
F
AJNA :11 , November/December 1990
STEREOTAXY OF CORTICAL LANDMARKS
identification is therefore preferable, but until now has remained restricted to only a few morphologically unique structures .
Our evaluation of the (indirect) proportional grid system
using techniques of direct sulcus mapping revealed variation
zones for all major sulci of 1 .5-2 em measured on the external
brain surface (Figs. 4-7). Similar spatial variability must be
assumed for deeply located parts of the sulci . The inaccuracy
of the proportional grid system was the same in the anteroposterior and vertical axes and increased with the distance
between the target and the inner reference points of the
anterior and posterior commissures. Variation of the callosal
sulcus (Fig. 7), for example, was about half that of the central
sulcus, posterior horizontal ramus, or calcarine sulcus, which
corresponds to about double the distance of the latter sulci
to the anterior or posterior commissure. Significant overlapping between variation zones was observed only for the
posterior suprasylvian landmarks. The sulcal zones may
therefore provide a guide for the identification of individual
sulci visible on high-resolution images. Structural regions of
interest based on proportional coordinates, however, would
be unreliable for an anatomically based data analysis. The
individual sulcus pattern was too unpredictable to allow focal
distinctions between, for instance, precentral or postcentral,
supra- or infrasylvian, and parietal opercular or angular locations by means of Talairach atlas coordinates (see Figs. 47). In fact, variability of the central sulcus or posterior horizontal ramus within the proportional grid was not less than
within the standard system for electroencephalographic scalp
electrode placement, which is based solely on the external
cranial landmarks of the nasion, inion, and preauricular points
[32).
These conclusions clearly depend on the ability to confidently identify the individual sulci with our MR technique . It is
no problem to recognize the rami of the sylvian fissure or the
superior temporal , calcarine, or callosal sulci on 5-mm sagittal
sections. It is more difficult to outline the inferior extension of
the central sulcus. The central sulcus was identified at the
superior hemispheric margin, where it is always found immediately in front of the so-called marginal sulcus; that is, the
terminal ascending ramus of the cingulate sulcus [5 , 19, 21,
1127
33). From there, the central sulcus was traced inferiorly by
using a cinematographic image display mode [19]. Since the
course of the central sulcus is unbroken through its entire
length [21 , 22] , the procedure left no doubt among the authors
that the sulcus named was actually the inferior central sulcus
in each individual. Therefore, identification of the inferior preand postcentral sulci was self-evident. It should be noted that
almost no overlap occurred between the variation zones of
these three sulci (Figs. 4 and 5). Thus , if any false identifications had occurred , the present investigation would have
actually underestimated anatomic variability. Strong evidence
against any misinterpretation of the suprasylvian sulcus pattern is provided by the original Talairach atlas (plate 154 of
Talairach et al. [1 ]), which shows the same location and
variation of the inferior central sulcus derived from 20 postmortem hemispheres.
Several studies used the Talairach atlas [1) to correlate
PET with anatomic structures [7 -1 6]. Fox et al. reported
highly focal cerebral blood flow (CBF) responses of the primary sensory cortices to cutaneous tactile vibration [11) or
retinal field stimulation [1 0, 12). The following anteroposterior
and vertical mean coordinates and standard deviations were
found for the primary somatosensory projection areas in eight
healthy subjects [11) (positive values were superior to the
estimated bicommissural line (ACPC) or anterior to the estimated midcommissuralline (MC) [11 ); MC would be equidistant to lines VAG and VPC in Figures 4-7 [11 ]):
1. Lip area, right hemisphere: MC , +0.2 ± 0.5 em (range,
+1.2 to -0.4 em); ACPC , +2.7 ± 0.6 em.
2. Finger area, right hemisphere (digits 1-4): MC, -0.8 ±
0.6 em (range, +0.1 to -1.9 em); ACPC, +4.8 ± 0.3 em .
3. Toe area, midsagittal plane (digits 1 and 2, bilateral
stimulation): MC, -1 .1 ± 1.1 em (range, +0.6 to -3.0 em);
ACPC, +5.1 ± 0.4 em .
Location and width of the sensory lip area are compatible
with the MR localization of the inferior postcentral gyrus (Figs.
4 and 5; it must be taken into account that the inferior central
sulcus has a slightly oblique, anterolateral to posteromedial ,
depth orientation , which influences the location of the sensory
CBF activation). Midsagittal CBF responses to toe vibration ,
however, were dislocated anteriorly by Fox et al. [11] com-
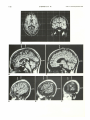
Fig. 3.-Stereotaxic MR evaluation of perisylvian and calcarine sulci. See key for abbreviations.
A, Axial and coronal T1-weighted images (SE 500/30/1, 1-cm slice thickness) show orthomorphic position within field of view.
8-F, Sagittal T1-weighted images (SE 500/40/4, 5-mm slice thickness) with individually constructed proportional grid overlays at 0 (8), 5 (C), 45 (0),
50 (E), and 55 (F) mm from midsagittal plane.
C, Direct identification of central (white curved arrow) and calcarine (black curved arrow) sulci. Marginal sulcus (straight arrow).
D and E, Direct identification of anterior (white open arrows) and posterior (solid arrows) ascending rami. Bottom of central sulcus (black open arrows)
has been traced in 5-mm lateral steps from C. Sulci anterior and posterior to central sulcus are pre- and postcentral sulci.
F, Image showing most lateral portion of posterior horizontal ramus of sylvian fissure (white open arrow) and superior temporal sulcus (black open
arrow). In this subject, perisylvia[l sulci were mapped in the following fashion: Portions of the central and pre- and postcentral sulci visible on E were
drawn on the individual overlay. The upper ends of these outlines were connected with those points on D where the central and pre- and postcentral sulci
open onto the external brain surface. Outlines of the anterior and posterior ascending rami were drawn as visible on E. Lateral-most portions of posterior
horizontal ramus and superior temporal sulcus were drawn as visible on F and connected with their anterior and posterior continuations on E. This
procedure gives a very close estimate of superficial course of sulci.
STEINMETZ ET AL.
1128
----~----~---
AJNR :11 , November/December 1990
R
L
cs
cs
/
/
~t/.1
I
/
1"\
1"\
/
/
I
--::7
H
1/
. :=
·. .: ::::::::: l::;.d;r/
:: :::;.;.;;
I
i\
I
""
u
v
\ !'---
\
ACPC
ACPC
1\
\
w-v
0..
>
B
A
Fig. 4.-A and 8, Perisylvian variation of right (R) and left (L) central and superior temporal sulci as obtained from sagittal MR imaging in 20 probands.
See key for abbreviations. Shaded areas represent va riation zones of posterior horizontal ramus of sylvian fissu re measured between points of origin of
anterior and posterior ascending rami. Zones of variation of central sulcus, posterior horizontal ramus, and superior temporal sulcus measure 1.5-2 em.
Note farther backward extension of zone of posterior horizontal ramus on left compared with right. Sulcal outlines at superior hemispheric convexity
represent superomedial central sulcus as identified on immediate paramedian sections, serving as starting points from which course of central sulcus
was traced laterally.
-
1-- ,..-
~
/
rJ fY I) Pocs
1i~ . .-::
/
I
Vj
I
~
PRCS
~
~~
~ I\ }iL 1
7
:::::
~Ill( < :)
\
/\}
\
~
nr
<
""' "' ,"\
~
/
\
\
I
~
PRCS
~ !ft~
ACPC
u
0..
>
A
fp o cs
~:: ::=<
V)/
~
.·.·
;:)
~ ::: ;.
ll\ tY.t n:
:··
:::-·
u
!'---
~ Ill\
\
<
>
\
~
I
~
JF
'XS
/
I
"" ......__v
--
p
R
· :· .
~
::.
L
""' "'1"\
\
\
· r::::==
.
ACPC
·' [/iF'
\
I
""
......__ v
~
B
u
~
'
u
1'---..
0..
>
Fig. 5.-A and 8, Perisylvian variation of right (R) and left (L) pre- and postcentral sulci as obtained by sagittal MR imaging in 20 probands. See key for
abbreviations. Shaded areas represent variation zones of posterior horizontal ramus of sylvian fissure. Variation zones of pre- and postcentral sulci
measure 1.5-2 em. Outlines of postcentral sulcus on left are more scattered throughout their band of variation and tend to lie more posterior than those
in right hemispheres.
pared with the MR localization of the bottom of the central
sulcus at the medial hemispheric aspect, which in our study
was found at an MC coordinate of -2 .1 ± 0.6 em (range,
-1 .2 to -3.4 em) (Fig . 4). The difference is statistically significant (p < .05; t test). The sensory toe representations
indicated by Fox et al. [11] extended up to 1.8 em anteriorly.
The increased range of variation may be due to their technique
of estimating the individual ACPC lines from the glabella-inion
line on lateral skull radiographs [8 , 9]. Variable tilts of the
estimated ACPC line thus may have added to the inaccuracy
of the proportional grid system . In the studies on the primary
visual projection , Fox et al. [1 0, 12] applied checkerboard
stimuli to restricted parts of the retina. The findings were
summed up from six normal subjects and confirmed retino-
tropy on the medial occipital lobe. However, the reported
coordinates of the CBF peak responses [1 0, 12] differ from
the MR location (Fig. 7) and from the Talairach atlas coordinates of the calcarine sulcus [1 , 17], which harbors most of
the striate cortex [18, 24]: Stimulation of the macula and
perimacula, for instance, produced maximal midsagittal activation below the estimated ACPC line: for macula, ACPC =
-0.50 em and MC = -6.86 em ; for perimacula, ACPC =
-0.12 em and MC = -6.29 em; for retinal periphery, ACPC
= +0.64 em and MC = -5.96 em. Mean values are from
Table 2 of Fox et al. [12]. (Compare with Fig. 7 or with atlas
plates [1 , 17].) Taken together, the findings of Fox et al. [1 012] suggest displacement of the CBF responses with respect
to the primary sensory cortices in two directions: anterior shift
STEREOT AXY OF CORTICAL LANDMARKS
AJNR :11 , November/December 1990
/
I
~
--- --
~
1/
![}? ~ /
?R ~~
A ARt
\ J,~
\~~\ .::::::
~Nfjj)
~\
\
I/ YJ
;::::
r>1:>
<
:-
~
"""1'\
1'--
/
\
\
I
/
I( . ~ ~;(;J h::::
1\
u
A
I"--
. f'\=''.
v
:::::
\
\
::::- .
ACPC
u
<(
>
"~'---- v
~
0-
>
'\
I
""
<(
/
~.,
"" """
.. ,: { ' /' {:/
ll'\1!, it..:<
ACPC
:::>
L
.:\ iJWJ 1:>
AAR
I ~\J
>
'\~
PAR)
1/
u
/
~
f.--
~
I
""'~-
--
R
PAR
/.
/
1129
u
0-
>
B
Fig. 6.-A and B, Perisylvian variation of right (R) and left (L) anterior and posterior ascending rami of sylvian fissure as obtained by sagittal MR imaging
in 20 probands. See key for abbreviations. Shaded areas represent variation zones of posterior horizontal ramus of sylvian fissure. Variation zones of
anterior and posterior ascending rami measure 1.5-2 em. Zone of left posterior ascending ramus is broader and extends farther posteriorly than
contralaterally, so that left parietal operculum is longer than its mate in most cases.
of the toe response and downward shift of the visual response, each on the order of 1 em. With our current MR
material , we tested their method to estimate the ACPC line
from the glabella-inion line [8). The estimated midcommissural
point was found at 4.8 mm superior to 4.5 mm inferior (mean,
0.83 mm superior; SD = 2.49 mm) and at 1.5-7 .3 mm
posterior (mean, 4.06 mm posterior; SD = 1.36 mm) to the
true midcommissural point (perpend icular measurements
from the true ACPC and MC on midsagittal MR slices). The
inclination of the estimated ACPC with respect to the true
ACPC ranged from -4.0° to +11 .8° (-0.06 ± 3.64° ; positive
values correspond to anterior elevation of the estimated
ACPC). Nevertheless, this only partly explains the discrepancies between functional localization in the studies of Fox et
al. [1 0-12] and direct anatomic localization.
Several groups proposed other methods to match PET with
structural standard templates [34-42]. Of these, Bajcsy et al.
[34] , Bohm et al. [35, 37 , 42] , and Dann et al. [41] described
the computerized superimposition of digitized, transformable
postmortem brain atlases on individual tomographic data.
Brain surface and ventricles served as main reference structures . These systems allow global transformations of scale,
rotation , and translation in x, y, and z axes as well as elastic
deformation of the atlas structures. They are superior to the
proportional grid method [42]. However, they do not solve
the principal problem of indirect localization ; that is, the comparison of individual brains under study with one standard
reference organ . Furthermore, changes of surface profile or
ventricular contours cannot b~ excluded in postmortem atlas
brains . Vice versa, structural abnormalities may be present in
patients. These issues were also addressed by Herholz et al.
[38] and Evans et al. [40] . In a comprehensive approach,
Evans et al. [ 40] developed a standard region-of-interest (ROI)
atlas derived from an in vivo MR data set. To account for
morphologic variability , they used an interactive procedure to
correct and redefine single standard ROis according to individual brain contours seen on MR , thus combining indirect
with direct localization .
/
~
----
f.-
-
~
~
""" '\
/
I
I
~
ACPC
1,_
\
~ ~ \I
~
~JI
~
~~\
~
""
I'-- ___./
~
-
\
\~
I
~
'\
'--
~
Fig. 7.-Variations of calcarine and callosal sulci as obtained by MR
imaging in 20 probands (pooled data from 40 hemispheres). See key for
abbreviations. Variation zone of callosal sulcus measures less than 1 em.
With increasing distance from center of reference system, sulcal variation
zones become broader (compare Figs. 4-6 and calcarine with callosal
sulcus).
Further improvement of direct localization will come from
gradient-echo MR volume measurements yielding isotropic
data matrices [43, 44). Direct MR identification and mapping
of cortical landmarks will thus allow the generation of individual brain templates and improved accuracy of structural ROI
placement. Although computationally expensive and userintensive, the intraindividual three-dimensional matching of
such MR data with PET appears to provide the most reliable
way for studies of structural-functional relationships in normal
and abnormal conditions [45]. It also renders identical repositioning unnecessary. Our group is currently validating an
intermodal surface-fitting method similar to that described by
Pelizzari et al. [46] .
STEINMETZ ET AL.
1130
REFERENCES
1. Talairach J, Szikla G, Tournoux P, et al. Atlas d 'anatomie stereotaxique du
telencephale. Paris: Masson, 1967
2. Olivier A, Peters TM , Clark JA, et al. Integration de l'angiographie numerique, de Ia resonance magnetique, de Ia tomodensitometrie et de Ia
tomographie par emission de positrons en stereotaxie. Rev Electroencephalogr Neurophysiol Clin 1987;17: 25-43
3. Vanier M, Ethier R, Clark J, Peters TM , Olivier A, Melanson D. Anatomical
interpretation of MR scans of the brain . Magn Reson Med 1987;4:
185- 188
4. Rumeau C. Gouaze A, Salamon G, et al. Identification of cortical sulci and
gyri using magnetic resonance imaging: a preliminary study. In: Gouaze A,
Salamon G, eds. Brain anatomy and magnetic resonance imaging . Berlin:
Springer, 1988:11-31
5. Steinmetz H, FOrst G, Freund HJ. Cerebral cortical localization: application
and validation of the proportional grid system in MR imaging. J Comput
Assist Tomogr 1989;13 :10-19
6. Vanier M, Roch Lecours A, Ethier R, et al. Proportional localization system
for anatomical interpretation of cerebral computed tomograms. J Comput
Assist Tomogr 1985;9 :715-724
7. Reiman EM , Raichle ME, Butler FK, Herscovitch P, Robins E. A focal
abnormality in panic disorder, a severe form of anxiety. Nature 1984;31 0:
683- 685
8. Fox PT, Perlmutter JS, Raichle ME. A stereotactic method of anatomical
localization for positron emission tomography. J Comput Assist Tomogr
1985;9 : 141-153
9. Fox PT, Fox JM , Raichle ME , Burde RM . The role of cerebral cortex in the
generation of voluntary saccades: a positron emission tomographic study.
J Neurophysiol1985;54 :348-369
10. Fox PT, Mintun MA, Raichle ME, Miezin FM , Allman JM, Van Essen DC.
Mapping human visual cortex with positron emission tomography . Nature
1986;323 : 806-809
11 . Fox PT, Burton H, Raichle ME. Mapping human somatosensory cortex
with positron emission tomography . J Neurosurg 1987;67:34-43
12. Fox PT, Miezin FM, Allman JM, Van Essen DC, Raichle ME. Retinotopic
organization of human visual cortex mapped with positron-emission tomography. J Neurosci 1987;7 :913-922
13. Roland PE , Eriksson L, Stone-Eiander S, Widen L. Does mental activity
change the oxidative metabolism of the brain? J Neurosci 1987;7 :23732389
14. Petersen SE, Fox PT, Posner MI. Mintun M, Raichle ME. Positron emission
tomographic studies of the cortical anatomy of single-word processing.
Nature 1988;331 :585-589
15. Posner Ml , Petersen SE, Fox PT, Raichle ME. Localization of cognitive
operations in the human brain. Science 1988;240: 1627-1631
16. Roland PE, Eriksson L, Widen L, Stone-Eiander S. Changes in cerebral
oxidative metabolism induced by tactile learning and recognition in man.
Eur J Neurosci 1989; 1 : 3-18
17. Talairach J. Tournoux P. Co-planar stereotaxic atlas of the human brain.
3-dimensional proportional system: an approach to cerebral imaging . Stu\1gart: Thieme, 1988
18. Brodmann K. Vergleichende Lokalisationslehre der Grosshirnrinde in ihren
Prinzipien dargestellt auf Grund des Zellenbaues. Leipzig: Barth, 1909
19. Ebeling U, Steinmetz H, Huang Y, Kahn T. Topography and identification
of the inferior precentral sulcus in MR imaging. AJNR 1989;10:937-942,
AJR 1989;153: 1051-1056
20. Jensen J. Die Furchen und Windungen der menschlichen GrosshirnHemispharen. Allg Psychiatr 1871;27 :473-516
21 . Eberstaller 0 . Das Stirnhirn. Vienna: Urban & Schwarzenberg, 1890
22 . Retzius G. Das Menschenhirn. Stockholm: Norstedt & Saner, 1896
23. Cunningham OJ . Contribution to the surface anatomy of the cerebral
hemispheres. Dublin: Royal Irish Academy, 1892
24. Stensaas SS, Eddington OK, Dobelle WH . The topography and variability
of the primary visual cortex in man. J Neurosurg 1974;40:747-755
z
AJNR:11 , November/December 1990
25 . Lang J, Gatzenberger H. Ober Ausbildung und Lage der Gyri et Sulci an
der Facies medialis hemispherii. Verh Anat Ges 1980;74 :687-688
26. von Economo C, Horn L. Ober Windungsrelief, Masse und Rindenarchitektonik der Supratemporalflache, ihre individuellen und ihre Seitenunterschiede. Z Neural Psychiatr 1930;130:678-757
27. Shellshear JL. The brain of the aboriginal Australian. A study in cerebral
morphology. Philos Trans R Soc Lond {Bioi] 1937;227 :293-409
28. Connolly CJ. External morphology of the primate brain. Springfield, IL:
Thomas, 1950
29. Rubens AB , Mahowald MW, Hutton JT. Asymmetry of the lateral (sylvian)
fissures in man. Neurology 1976;26:620-624
30. Yeni-Komshian GH, Benson DA. Anatomical study of cerebral asymmetry
in the temporal lobe of humans, chimpanzees, and rhesus monkeys.
Science 1976; 192: 387-389
31 . Habib M, Renucci RL, Vanier M, Corbaz JM , Salamon G. CT assessment
of right-left asymmetries in the human cerebral cortex. J Comput Assist
Tomogr 1984;8:922-927
32. Steinmetz H, FOrst G, Meyer BU. Craniocerebral topography within the
International 10-20 System. Electroencephalogr Clin Neurophysiol
1989; 72:499-506
33. Berger MS, Cohen W, Ojemann GA. Correlation of motor cortex brain
mapping data with magnetic resonance imaging. J Neurosurg 1990;
72:383-387
34. Bajcsy R, Lieberson R, Reivich M. A computerized system for the elastic
matching of deformed radiographic images to idealized atlas images. J
Comput Assist Tomogr 1983;7 :618-625
35. Bohm C, Greitz T, Kingsley D, Berggren BM, Olsson L. Adjustable computerized stereotaxic brain atlas for transmission and emission tomography. AJNR 1983;4:731-733
36. Mazziotta J, Phelps M, Plummer D, Schwab R, Halgren E. Optimization
and standardization of anatomical data in neurobehavioural investigations
using positron computed tomography. J Cereb Blood Flow Metab
1983;3[Supp11]:S266-267
37. Bohm C, Greitz T, Kingsley D, Berggren BM, Olsson L. A computerized
individually variable stereotaxic brain atlas. In: Greitz T, lngvar DH, Widen
L, eds. The metabolism of the human brain studied with positron emission
tomography. New York: Raven, 1985:85~91
38. Herholz K, Pawlik G, Wienhard K, Heiss W-0 . Computer assisted mapping
in quantitative analysis of cerebral positron emission tomograms. J Comput
Assist Tomogr 1985;9 :1 54-161
39. Samson Y, Baron J-C, Feline A, Bories J, Crouzel C. Local cerebral glucose
utilization in chronic alcoholics: a positron tomographic study. J Neurol
Neurosurg Psychiatry 1986;49 : 1165-1170
40. Evans AC , Beil C, Marrett S, Thompson CJ , Hakim A. Anatomical-functional correlation using an adjustable MAl-based region of interest atlas
with positron emission tomography. J Cereb Blood Flow Metab
1988;8: 513-530
41 . Dann R, Hoford J, Kovacic S, Reivich M, Bajcsy R. Evaluation of elastic
matching system for anatomic (CT, MR) and functional (PET) cerebral
images. J Comput Assist Tomogr 1989;13:603-611
42. Seitz RJ , Bohm C, Greitz T, et al. Accuracy and precision of the computerized brain atlas programme (CBA) for localization and quantification in
positron emission tomography. J Cereb Blood Flow Metab 1990;10:443457
43. Steinmetz H, Rademacher J, Huang Y, et al. Cerebral asymmetry: MR
planimetry of the human planum temporale. J Comput Assist Tomogr
1989;13:996-1 005
44. Steinmetz H, Rademacher J, Jancke L, Huang Y, Thron A, Zilles K. Total
surface of temporoparietal intrasylvian cortex: diverging left-right asymmetries. Brain Lang (in press)
45. Levin ON , Hu X, Tan KK, et al. The brain: integrated three-dimensional
display of MR and PET images. Radiology 1989;172:783-789
46. Pelizzari CA, Chen GTY, Spelbring DR, Weichselbaum RR , Chen CT.
Accurate three-dimensional registration of CT, PET, andfor MR images of
the brain. J Comput Assist Tomogr 1989;13:20-26