Survey
* Your assessment is very important for improving the workof artificial intelligence, which forms the content of this project
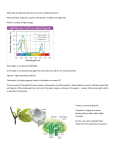
Energy Pathways (Metabolism) in Cell A. Energy (def)-capacity to do work or cause change 1. Cells transform energy from one type to another. Energy can exist in 2 states: a. potential—stored (energy that matter possesses because of its location or structure) Chemical energy is PE available for release in a chemical reaction b. kinetic—associated with the relative motion of objects; expended in the process of doing work example: photons of light can be captured & their energy harnessed to power photosynthesis 2. Thermodynamics: study of energy transformations definitions within biology: system—matter under study surroundings—everything outside the system a. closed system: matter isolated from its surroundings (like coffee in a thermos) b. open system: energy & matter can be transformed between the system & its surroundings. c. organisms: open systems. They absorb energy (light or chemical) in the form of organic molecules, and release heat and metabolic wastes like urea & carbon dioxide to their surroundings. 3. Laws of Thermodynamics a. law of energy conservation: energy can neither be created or destroyed; only converted from one form to another. The electric company doesn’t make energy-- neither do plants— but during every energy transfer/transformation, some energy becomes unusable or unavailable, leading to: b. every energy transfer increases the entropy of the universe events in the universe proceed in a downhill direction (from higher to lower energy levels) Entropy: a quantity used as a measure of disorder and randomness --means anytime energy is exchanged, some usable energy is lost (entropy) examples: abandoned house going into disrepair cars converting only about 25% of energy in gas into motion c. for a process to occur on its own, without outside help in the form of energy input, it must increase the entropy of the universe 1) spontaneous processes occur without energy input—need not occur quickly. Example: car rusting 2) free energy (G) change of a reaction tells whether reaction is spontaneous or requires energy input 3) free energy is used as a measure of the stability of a system (the greater the G, the more unstable the system). Example: compressed spring d. for living things to maintain low entropy, there must be steady energy inputs 4. Energy Transfer Molecule a. ATP—adenosine triphosphate (Fig 8.8) components: one ribose (pentose) sugar one adenine (nitrogenous base) three phosphate groups—bonds between them can be broken by hydrolysis b. “energy currency” used by cells when energy is needed (cells can release energy stores, or store energy to use later) c. humans have only enough ATP to last for 20 seconds; must be continually replenished (ATP cycle) ATP is formed from the burning, or oxidizing of food molecules, in process called cellular respiration. Bonds between the last 2 phosphates release energy to be used in cells d. exergonic vs endergonic reactions (Fig 8.6) ATP → energy + ADP + P (exergonic reaction: free energy is released; ∆G is negative) catabolic ADP + P + energy → ATP (endergonic reaction: absorbs free energy & stores it in molecule). Called phosphorylation e. approximately 50% of energy is lost as heat & utilized by warm-blooded animals (heat can only do work when temp goes down from warmer to cooler locale; if temp is uniform, like in a cell, it can only warm the organism) f. some life forms use ATP & release light; example: fireflies (bioluminescence) 5. Metabolic pathways --metabolism: all an organisms chemical reactions. Specific enzymes catalyze each step a. catabolic-degrade complex compounds; “breakdown pathways” (exergonic-means ‘energy out’) release energy by breaking down complex molecules to simpler compounds ex. cellular respiration: glucose is broken down in the presence of oxygen to carbon dioxide & water b. anabolic-“biosynthesis pathways” (endergonic- means ‘energy in’) consume energy to build complicated molecules ex. synthesis of proteins from amino acids c. energy coupling (ATP is responsible for managing most energy coupling in cells) Energy released from the downhill reactions of catabolism can be stored to drive the uphill reactions of the anabolic pathways. *note: the release of energy from hydrolysis of ATP comes from chemical change to state of lower free energy, not from the phosphate bonds themselves. 6. Oxidation & Reduction (all about electrons) a. oxidation state-measure of energy content most energy is released from highly reduced molecules b. oxidized-loses one or more electrons c. reduced-gains one or more electrons d. called redox reactions e. example: salt f. happens with covalent bonds too—example using C: C & H have about same affinity for valence electrons, but when methane reacts with oxygen to form carbon dioxide, electrons end up farther away from the C and closer to the O, which are very electronegative methane (most reduced state) electrons pulled in by C methanol formaldehyde formic acid carbon dioxide (most oxidized state) electrons pulled away by O B. Processing Energy: Photosynthesis—def: process by which molecules are produced by cells using light energy 1. Importance --energy for life --transferring of light energy into chemical energy --most important set of reactions 2. autotrophs & heterotrophs-both need to obtain organic compounds a. autotrophs (producers) -use CO2 as their source of carbon. REQUIRES ENERGY - most use light energy (photosynthesis) - some use inorganic substances like sulfur & ammonia (chemosynthesis) b. heterotrophs (consumers) - must consume organic molecules - more than 80% of living species 3. Overview a. overall chemical equation b. two basic phases or parts: 1. light dependent reactions -light energy is absorbed, stored in chemical energy in ATP & NADPH -also involves the splitting of water; O2 released as a byproduct 2. light independent reactions (used to be called the dark reactions) -carbohydrates are synthesized from CO2 using energy from ATP and NADPH 4. Light-dependent reactions (occur in the thylakoid membranes) “photo” part of photosynthesis; convert solar energy to chemical energy a. light: visible light is the radiation that drives photosynthesis (1) Light travels in waves, in packets called photons (2) energy content depends on wavelength; shorter wavelengths = higher energy (3) when light meets matter, it may be reflected, transmitted, or absorbed (4) spectrophotometer-measures the ability of a pigment to absorb various wavelengths of light b. pigments: (1) absorb wavelength whose energy matches the energy needed to boost an electron to a higher orbital; it excites them (2) absorb light in the visible spectrum: 380nm to 750 nm (an absorption spectrum plots a pigments light absorption vs. its wavelength) (3) major groups that trap light energy (a) chlorophylls: green pigments in chloroplasts of plants; what makes leaves green. --Chlorophyll a --Chlorophyll b: “accessory pigment” funnels the light energy it absorbs to chlorophyll a Chloroplasts are mostly found in mesophyll cells that form the tissue in the interior of the leaf. All green parts of a plant have chloroplasts, but the leaves are where most photosynthesis occurs. Chlorophylls reflect green and absorb red & blue wavelengths. They also contain Mg (b) carotenoids: accessory pigments that funnel energy from other wavelengths to chlorophyll a; also protects against excessive light. Reflect red & yellow; absorb blue & green wavelengths. c. chloroplast organization: (see Figure 10.4 p186) Thylakoid: site of light dependent reactions (1) thylakoids contain the chlorophyll (2) grana- stacks of thylakoid membranes (3) stroma- background material between the grana d. organization of photosynthetic pigments embedded in thylakoid membranes are clusters called photosystems there are several thousand per chloroplast Photosystems consist of: (1) center called reaction center—particular chlorophyll a molecule and a primary electron acceptor. Either: Photosystem I : P700 means reaction center chlorophyll a has an absorption peak of 700nm or Photosystem II: P680 with absorption peak at 680nm (2) light harvesting complex “antenna pigments” that surround each reaction center (250-350 per center) receive wavelengths (energized) and pass the energy to the reaction center (3) reaction center passes photoexcited electrons to an electron acceptor (4) P700+ and P680+ are left behind (lost an electron, or oxidized) e. Noncyclic Photophosphorylation: predominant route. Involves both Photosystems I & II --produces both ATP (for chemical energy) & NADPH (for reducing power) (1) electrons leave both P680 & P700 and are received by separate electron acceptors, forming P680+ and P700+ (2) photolysis occurs (water molecules are split) when the attraction of P680+ pulls the lower energy electrons from water H2O → ½ O2 + 2e-- + 2H+ (a) O2 is released into atmosphere (as a waste or byproduct) not from CO2 (b) the electrons restore P680+ to P680 (c) H+ is released into the thylakoid lumen (as a waste or byproduct) (3) electrons which left P680 and went to its primary electron acceptor are relayed through an electron transport system to Photosystem I; as they “fall” to a lower energy level, their energy is harnessed to produce ATP (4) electrons from Photosystem I (P700) are received by another electron acceptor; the electron hole left in the P700 is filled by an electron that reaches the bottom of the electron transport chain from PS2. (a) they react with NADP+ and H+ to form NADPH (used later) (5) ATP formed from chemiosmosis (fig 10.18 p 197) H+ gradient—more concentrated in the thylakoid lumen, so ATP is produced as they return to stroma (6) Summary: Noncyclic electron flow pushes electrons from water, where they’re at a low state of potential energy, to NADPH, where they’re stored at a high state of potential energy. ATP is also generated. Therefore, thylakoids convert light energy to chemical energy stored in NADPH and ATP (oxygen is a byproduct) f. Cyclic Photophosphorylation (1) only Photosystem I (no NADPH production or photolysis, so no O2 either) (2) P700 releases energized electrons, which are received by an electron acceptor & passed through an electron transport system back to P700+ (3) transfer moves H+ into the thylakoid lumen to form H+ gradient—ATP is produced by chemiosmosis 5. Light-independent reactions (because they don’t need light directly) occur in stroma; “synthesis” part of photosynthesis (anabolic) a. overall function: to use the energy from ATP and energy rich electrons from NADPH to form a product (often glucose). ATP & NADPH contain an abundance of chemical energy, but are not stable enough to store that energy for more than a few minutes. Goal is to convert them to high energy compounds that can be used for a long time b. three variations identified in plants: C3 , C4, and CAM c. C3 pathway is the most common by far (Calvin-Benson cycle) CO2 enters the cycle and leaves as sugar --occurs mostly during the day anyway, because that’s when the light reactions can provide the ATP & NADPH the Calvin cycle needs. (1) carbon dioxide fixation: first step (a) CO2 is fixed to a 5 carbon molecule, RuBP (ribulose biphosphate) this is catalyzed by the enzyme RuBP carboxylase, commonly known as RUBISCO, probably the most abundant protein on earth. (b) forms an unstable 6 carbon molecule which immediately forms two 3 carbon molecules of PGA (phosphoglyceric acid; or 3-phosphoglycerate in Campbell textbook) This is a low energy molecule (2) reduction of PGA to PGAL (phosphoglyceraldehyde, or G3P in text) occurs next (a) ATP supplies the energy (b) NADPH supplies the electrons (3) regeneration of RuBP: PGAL (G3P) is higher in energy. Some is converted to various products, like glucose, amino acids, fatty acids, etc. The rest is converted to 5 carbon molecules of RuBP to feed the cycle. d. C4 and CAM synthesis (see Problem with Calvin Cycle page & Figure 10.21 p 202 --different method of fixing CO2 -- modification for better survival in hot, dry habitats (1) C4 photosynthesis—spatial separation of steps C4 plants first fix CO2 in a 4 carbon compound examples: sugarcane, corn (a) plants have 2 types of photosynthetic cells; bundle sheath cells and mesophyll cells (b) Calvin cycle: occurs in bundle sheath cells, but CO2 is incorporated into organic molecules in the mesophyll first (c) instead of being fixed by rubisco into PGA, the CO2 combines with PEP (phosphoenolpyruvate) to form OAA (oxoloacetate) in the mesophyll cells. OAA has 4 carbons, that’s where the name C4 comes from. The key enzyme involved in this is PEP carboxylase. (d) PEP carboxylase has a much higher affinity for CO2 than rubisco, and no affinity for O2. (e) the mesophyll cells pump the 4 carbon compounds into the bundle sheath cells (in the form of malate) where it’s converted to pyruvate and CO2 . Now rubisco can use CO2 to start the Calvin cycle, and not use the O2 instead. Pyruvate is sent back to the mesophyll cells, where it’s converted back to PEP so the process can repeat (2) CAM photosynthesis—temporal separation of steps Crassulacean Acid Metabolism (CAM) --similar to C4 photosynthesis (a) PEP still fixes CO2 to OAA, but instead of maleate, OAA is converted to malic acid (this is a minor difference, malic acid is just an ionized form of maleate) (b) Malic acid is shuttled to the vacuole of the cell (c) Stomata open at night. PEP carboxylase is active, malic acid accumulates in the cell’s vacuole instead of going to the bundle sheath. (d) Stomata close during the day (reverse of other plants) This is when malic acid is sent out of the vacuole and converted back to OAA, releasing CO2 that can enter the Calvin cycle (e) advantage: CAM can proceed during the day with the stomata closed, so plant loses less water. This adaptation is seen in desert plants, including cacti. The name came from the family of plants CAM was discovered in. Problem with Calvin Cycle: One of the major problems facing terrestrial plants is dehydration. The stomata are not only the major route for gas exchange (CO2 in and O2 out), but also for the evaporative loss of water. On hot, dry days, plants close their stomata to conserve water. This causes problems for photosynthesis. In most plants (C3 plants like rice, wheat, and soybeans), initial fixation of CO2 occurs via rubisco, forming a three-carbon compound, 3-phosphoglycerate. When their stomata partially close on a hot, dry day, CO2 levels drop as CO2 is consumed in the Calvin cycle. At the same time, O2 levels rise as the light reaction converts light to chemical energy. While rubisco normally accepts CO2, when the O2:CO2 ratio increases (on a hot, dry day with closed stomata), rubisco can add O2 to RuBP. When rubisco adds O2 to RuBP, RuBP splits into a three-carbon piece and a two-carbon piece in a process called photorespiration. The two-carbon fragment is exported from the chloroplast and degraded to CO2 by mitochondria and peroxisomes. Unlike normal respiration, this process produces no ATP, and furthermore, consumes it. Unlike photosynthesis, photorespiration does not produce organic molecules; it decreases photosynthetic output by siphoning organic material from the Calvin cycle. A hypothesis for the existence of photorespiration is that it is evolutionary baggage. When rubisco first evolved, the atmosphere had far less O2 and more CO2 than it does today, so the inability of the active site of rubisco to exclude O2 would have made little difference. Today it does make a difference. Photorespiration can drain away as much as 50% of the carbon fixed by the Calvin cycle on a hot, dry day. Certain plant species have evolved alternate modes of carbon fixation to minimize photorespiration.