Survey
* Your assessment is very important for improving the work of artificial intelligence, which forms the content of this project
Arrhythmogenic right ventricular dysplasia wikipedia , lookup
Cardiac surgery wikipedia , lookup
Drug-eluting stent wikipedia , lookup
History of invasive and interventional cardiology wikipedia , lookup
Quantium Medical Cardiac Output wikipedia , lookup
Dextro-Transposition of the great arteries wikipedia , lookup
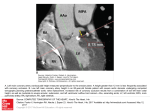
490 Dynamic Relation Between Myocardial Contractility and Energy Metabolism During and Following Brief Coronary Occlusion in the Pig Gregory G. Schwartz, Saul Schaefer, Dieter J. Meyerhoff, Joel Gober, Patricia Fochler, Barry Massie, and Michael W. Weiner Downloaded from http://circres.ahajournals.org/ by guest on June 16, 2017 Changes in high-energy phosphate metabolism may be important in the regulation of myocardial contractile function during ischemia. This study sought to determine the dynamic relation between myocardial contractile function and high-energy phosphate metabolism during and following brief (24-second) coronary occlusion, when large and rapid changes in both parameters occur. Eight anesthetized, open-chest pigs were instrumented with a Doppler flow probe and occluder on the anterior descending coronary artery, segment length crystals in the anterior left ventricular wall, and a surface coil for phosphorus-31 nuclear magnetic resonance spectroscopy. Phosphorus-31 spectra were reconstructed with a 4.8-second time resolution by summing corresponding short blocks of data from multiple occlusions. Metabolic and functional parameters were unchanged during the first 4.8 seconds of occlusion. During the remainder of occlusion, phosphocreatine progressively declined to 66±3% of control, inorganic phosphate rose to 170+±8% of control, and segment shortening fell to 25±9%o of control. A strong linear correlation was found between dynamic changes in segment shortening and phosphocreatine (r2=0.97), inorganic phosphate (r2=0.96), and the ratio of phosphocreatine to inorganic phosphate (r2=0.98) during occlusion. At any level of the ratio between phosphocreatine and inorganic phosphate, segment shortening was greater during reflow than during occlusion. The close, dynamic relation between segment shortening and phosphorus metabolites supports the regulation of contractility by changes in energy metabolism or its by-products during ischemia. During reactive hyperemia, the high coronary flow rate may be an independent factor modulating contractility. (Circulation Research 1990;67:490-500) yocardial ischemia perturbs high-energy phosphate metabolism in the cardiac myocyte and leads to a decline in contractile function."2 For over 20 years, these observations have led to hypotheses that cardiac contractility is regulated by levels of myocardial high-energy phosphates or impaired by metabolic by-products of ischemia, such as H' or inorganic phosphate (Pi).3 M From the Magnetic Resonance Unit and Cardiology Section, Veterans Administration Medical Center, and the Cardiovascular Research Institute and Departments of Medicine and Radiology, University of California, San Francisco, Calif. Supported in part by National Institutes of Health awards 1K11-HL-02155 (G.G.S.), K08-HL-02131 (S.S.), and R01AM-33923 (M.W.); the American Heart Association, California Affiliate (S.S.); Philips Medical Systems; and the Veterans Administration Medical Research Service (M.W.W., B.M.). Address for correspondence: Gregory G. Schwartz, MD, PhD, Cardiology Section (lllC), VA Medical Center, 4150 Clement St., San Francisco, CA 94121. Received October 20, 1989; accepted April 3, 1990. Because of the limited temporal resolution of metabolic measurements made by direct biochemical techniques or by nuclear magnetic resonance (NMR) spectroscopy, most of the data supporting these hypotheses in intact animals has stemmed from studies of prolonged myocardial ischemia or hypoxia.4-7 However, the most rapid and pronounced changes in myocardial contractile function and energy metabolism occur during the first 30 seconds of ischemia and during the initial period of recovery after a brief ischemic interval.8-13 Elucidating the dynamic relation between such large and rapid changes in cardiac contractile function and energy metabolism may help define the mechanisms by which cardiac contractility is regulated during and following ischemia. In addition to changes in energy metabolism, other factors may have contributory, if not primary, roles in the regulation of contractile function in the intact animal. For example, an increase in coronary flow or perfusion pressure may independently augment myo- Schwartz et al Dynamic Changes of Myocardial Function and Energy Metabolism cardial contractility. This observation has been termed the Gregg phenomenon.1415 With release of a brief coronary occlusion, coronary blood flow rises to several times its control level during reactive hyperemia; however, it is unknown whether myocardial contractile function is affected by this elevation of coronary blood flow. The purpose of this study was to determine the dynamic relation between changes in myocardial contractile function and high-energy phosphate metabolism during brief coronary occlusion and subsequent reflow in the in situ, blood-perfused heart. To augment the temporal resolution of the metabolic measurements, 31P NMR spectra were reconstructed from corresponding brief time intervals of repeated, identical interventions. Downloaded from http://circres.ahajournals.org/ by guest on June 16, 2017 Materials and Methods Animal Preparation Eight female Yorkshire-Landrace pigs weighing 30-40 kg were used. After premedication with ketamine HCl (20 mg/kg i.m.), anesthesia was induced in six pigs with halothane (3%) by face mask and sodium pentobarbital (10 mg/kg i.v.). In two pigs, anesthesia was induced with a-chloralose (150 mg/kg i.v.). Normal saline (500 ml i.v.) was administered rapidly with induction of anesthesia, followed by a continuous infusion of 75 ml/hr. After endotracheal intubation, the animals were mechanically ventilated with oxygen by a pressure-cycled respirator. Ventilatory rate and tidal volume were adjusted to maintain arterial pH between 7.30 and 7.45 with Po2 greater than 100 mm Hg. Anesthesia was maintained with halothane (0.5-1.0%) and sodium pentobarbital (1-2 mg/kg i.v. every 30 minutes) in six pigs and with a-chloralose (20 mg/kg i.v. every 30 minutes) in two pigs. An 8F fluid-filled introducer sheath was inserted in a carotid artery for monitoring of arterial pressure. A 7F pigtail catheter was advanced through the sheath into the left ventricle for pressure measurement. The heart was exposed via a median sternotomy, and two proximal segments of the left anterior descending coronary artery were dissected free for placement of a hydraulic cuff occluder (In Vivo Metric, Healdsburg, Calif.) and a perivascular Doppler flow probe (Crystal Biotech Inc., Holliston, Mass.). The occluder was placed as proximally as possible on the left anterior descending coronary artery to obtain a large ischemic region. In no case did a major diagonal branch originate proximal to the occluder. Bipolar pacing wires were inserted in the left atrial appendage. The ischemic region was defined by inflating the occluder until an area of gross cyanosis and akinesis was clearly demarcated on the anterior left ventricular surface. A two-turn, 2.5-cm diameter surface coil tuned to the 31P resonance frequency was glued with cyanoacrylate in the central portion of this region. A pair of ultrasonic dimension gauges (Triton Technology, San Diego) was implanted 1 cm apart 491 along the axis of fiber orientation16 in the subendocardium 1-2 mm adjacent to the surface coil. In all pigs, the visibly ischemic region was much larger than the area encompassed by the coil and dimension gauges. Segment shortening and coronary flow velocity were measured with a Triton sonomicrometer and flowmeter, respectively. Aortic and left ventricular pressures, left ventricular dP/dt, coronary flow velocity, subendocardial segment length, and the intramyocardial electrogram (from the dimension gauges) were recorded on a multichannel recorder (Gould, Cupertino, Calif.). Experimental Protocol The pig was placed in a 1-m bore Philips Gyroscan NMR spectrometer (Shelton, Conn.), operating at 2.0 T. Heart rate was maintained at 100 beats/min by atrial pacing. The pacing stimulus was used to gate the spectrometer acquisitions. To avoid radiofrequency noise introduced by wires leading into the spectrometer bore, the stimulus was transmitted to the spectrometer console by a telemetry system (Hewlett-Packard, Palo Alto, Calif.). After the phosphorus surface coil had been tuned to proton frequency (85.9 MHz) with a GordonTimms arrangement,17 the magnet was shimmed on cardiac water protons to an average linewidth of 25 Hz. The surface coil was then tuned to phosphorus frequency (34.8 MHz). A small glass vial containing hexamethylphosphorous triamide, placed in the center of the coil, served as a standard for determining the phosphorus 900 pulse length at the center of the coil. Spectroscopy was then performed with 1800 pulses gated 50 msec after the atrial pacing stimulus (at end diastole) of every second cardiac cycle, resulting in a repetition time of 1.2 seconds. A 1800 pulse length at the center of the coil was selected for data acquisition on the basis of computer simulations18 to provide maximal weighting of the NMR signal at a depth of approximately 0.7 cm from the coil, in the region where segment shortening was measured. However, this pulse length also allowed some spectral contribution from 2,3-diphosphoglycerate (DPG) in left ventricular cavity blood. The experimental intervention was a 24-second occlusion of the left anterior descending coronary artery, followed by complete release of the occlusion. With each intervention, spectroscopy was performed over a 2-minute interval spanning a 34-second preocclusion control period, the 24-second occlusion, and 62-second recovery following occlusion release. Groups of four successive acquisitions, each group spanning 4.8 seconds, were stored separately. Left anterior descending coronary artery occlusion (24 seconds) was repeated a total of 20 times at 5-minute intervals. Measurement of coronary flow and segment shortening simultaneous with NMR acquisitions introduced excessive radiofrequency noise in the NMR receiver. Therefore, these data were obtained during different occlusions. Coronary 492 Circulation Research Vol 67, No 2, August 1990 flow and segment shortening were measured during occlusions 1-3, 9-12, and 18-20. Spectroscopy was performed during occlusions 4-8 and 13-17. Before spectrosocpy, the cables from the flow probe and dimension gauges were disconnected from the flowmeter and sonomicrometer and coiled inside the magnet bore; the tuning and matching of the surface coil were then readjusted, if necessary. Control (nonischemic) hemodynamic measurements and a control spectrum of 40 acquisitions were obtained before the first coronary occlusion and following every 3-5 occlusions thereafter. At the conclusion of an experiment, the pig was killed with a lethal overdose of sodium pentobarbital. The position of the segment length crystals in the subendocardium was confirmed in all pigs by postmortem inspection. Downloaded from http://circres.ahajournals.org/ by guest on June 16, 2017 Data Analysis Systolic segment shortening was computed according to the method of Theroux et al.8 End-diastolic segment length was measured at the peak of the intramyocardial ventricular electrogram, and endsystolic segment length was measured 20 msec before peak negative left ventricular dP/dt. The timing intervals were established with a short length of fluidfilled tubing before the pig was placed in the spectrometer. Percent segment shortening was calculated as 100 x (end-diastolic- end-systolic length)/enddiastolic length. Coronary blood flow velocity and segment shortening were averaged during the same 4.8-second intervals as the NMR measurements. At 2-T field strength, sufficient NMR data cannot be acquired in 4.8 seconds to yield a useful 31P spectrum. To improve temporal resolution, 4.8second blocks of data, each consisting of four acquisitions, were summed from the corresponding time intervals of 10 interventions. Thus, 40 acquisitions were used to reconstruct a phosphorus spectrum from each 4.8-second interval preceding, during, and following coronary occlusion. The summed free induction decays were processed by exponential multiplication with line broadening of 20 Hz, convolution difference with a line broadening of 200 Hz and a convolution factor of 0.9, fast Fourier transformation, and phasing with zero- and firstorder corrections. Peak position and height were determined for Pi including superimposed 2,3-DPG, phosphocreatine (PCr), and the 3-phosphate of ATP. To allow adequate time for the phosphorus metabolites to reach a steady state of partial saturation, the first eight acquisitions during the control period of each intervention were not analyzed. During the remaining 24 seconds of the preocclusion control period, the average peak height of each metabolite was computed for each pig. During each ensuing occlusion and postocclusion time interval, the peak height of each metabolite was normalized to its average preocclusion control value. We have demonstrated (unpublished data) that the linewidths of PCr and 3-ATP do not change appreciably during or following brief coronary occlusion; therefore, nor- TABLE 1. Effect of Repeated Brief Coronary Occlusion on Metabolic and Functional Parameters SS (%) PCr/Pi PCr/,3-ATP End End Beginning End Beginning Beginning 19±3 2.0±0.2 19±3 1.9±0.1 2.5±0.3 2.3±0.2 p=0.49 p=0.25 p=0.19 Beginning and end refer to the total period of data collection. Metabolite values are expressed as ratios of peak heights. All values are mean±SEM. PCr, phosphocreatine; Pi, inorganic phosphate; SS, segment shortening. p values computed with paired t tests (n=8). malized changes in metabolite peak height should closely approximate normalized changes in peak area. During and immediately following ischemia, intracellular myocardial pH was estimated from the chemical shift of the Pi peak'9; however, spectral overlap of Pi and 2,3 -DPG precluded accurate pH determination under nonischemic conditions. Segment shortening and metabolite peak heights for each pig were normalized to the control levels for that animal. The normalized data from all eight pigs were then pooled. In the pooled data, the significance of changes from control in each occlusion and postocclusion time interval were assessed by Student's t test with a Bonferroni correction20 for 18 comparisons (five occlusion and 13 recovery intervals). A value of p<0.05 was considered statistically significant. Data are reported as mean±SEM. Results Under control conditions, systolic, diastolic, and mean aortic pressures were 74±3, 54±4, and 64±3 mm Hg, and systolic segment shortening was 19±3%. The PCr/,f-ATP peak height ratio was 2.0±0.2, with half-height linewidths of PCr and ,B-ATP of 1.2±0.1 and 1.4±0.1 ppm, respectively. The signal-to-noise ratios for PCr and ,B-ATP were 8.3 ±0.4 and 4.8 ±0.3. With occlusions spaced 5 minutes apart, there were no changes in systemic hemodynamics, coronary blood flow, or segment shortening, either under control conditions or in response to occlusion, during the course of the experiment. There were also no cumulative changes in control phosphorus spectra. Table 1 compares metabolic and functional data obtained under control (nonischemic) conditions at the beginning and the end of data acquisition. The absence of discernible persistent effects of repeated brief coronary occlusion validates the technique of spectral reconstruction using acquisitions from successive interventions. Figure 1 shows typical recordings of segment length and arterial pressure with corresponding phosphorus NMR spectra reconstructed from control, ischemic, and recovery time intervals. Normalized phosphorus metabolite peak heights during each 4.8-second interval are listed in Table 2, along with calculated intracellular pH values. Table 3 lists normalized end-diastolic and end-systolic segment Schwartz et al Dynamic Changes of Myocardial Function and Energy Metabolism CL C B A 493 E E Q! 7S so4 <0 1 "c - Downloaded from http://circres.ahajournals.org/ by guest on June 16, 2017 PCr 10.0 -10.0 0.0 [p p m] -20.0 0.0 10.0 [ p p m] -10.0 -20.0 10.0 o.0 -10.0 -20.0 [ p p m] FIGURE 1. Top panel: Segment length in the subendocardium of the anterior left ventricle and aortic blood pressure recorded before, during, and following 24-second coronary occlusion. Heart rate is 100 beats/min. Bottom panel: Representative 31P nuclear magnetic resonance spectra reconstructed from corresponding 4.8-second time intervals. Panel A: Preocclusion control interval. Panel B: Occlusion (19.2-24 seconds). Panel C: Reflow (19.2-24 seconds). Pi inorganic phosphate; 2,3-DPG, 2,3diphosphoglycerate; PCr, phosphocreatine. length, segment shortening, and coronary blood flow velocity during each of these intervals. Metabolite peak height and segment shortening data are plotted in Figure 2. During the first 4.8 seconds of occlusion, there were no detectable changes in any phosphorus metabolite peaks, nor in segment shortening (Tables 2 and 3). Beginning in the second time interval of occlusion (4.8-9.6 seconds) and continuing through the remainder of occlusion, PCr peak intensity declined and Pi rose, with little change in ,3-ATP. Concomitantly, end-diastolic segment length increased slightly and end-systolic segment length increased markedly, resulting in a progressive decline of systolic segment shortening. By the last time interval of occlusion (19-24 seconds), PCr (normalized to its control value) fell to 0.66±0.03, and normalized Pi rose to 1.70±0.08. A slight decrease in the 13-ATP peak was noted at the end of occlusion and in the early recovery period, but this trend did not achieve statistical significance. Segment length (normalized to control) rose to 1.06+0.08 at end-diastole and 1.25 ±0.11 at end-systole, resulting in a decline of segment shortening to 5±+2% (0.25+±0.11 normalized to control). Segment shortening, PCr, and Pi were all still changing when the occlusion was released at 24 seconds (Figure 2); therefore, it is reasonable to expect that longer occlusions would have caused even more severe metabolic and functional derangements (e.g., net systolic segment expansion). With coronary occlusion, both systolic and diastolic aortic pressure fell by 3-7 mm Hg, with occlusion release blood pressure returned to control within 10-15 seconds. Left ventricular end-diastolic pressure increased by 1-2 mm Hg during occlusion from its control value of 5 + 1 mm Hg. Reactive hyperemia accompanied occlusion release, with coronary blood flow velocity rising to 2.95 ±0.22 times control at the peak of the response, 10-15 seconds after release. Segment shortening began to recover in the first postocclusion time interval (0-4.8 seconds after release), while phosphorus metabolites began to recover in the second postocclusion time interval (4.8-9.6 seconds after release). By the interval 19-24 seconds after occlusion release, all phosphorus metabolites and segment shortening had returned to within +5% of their respective control values (Figure 2, Tables 2 and 3). Beginning approximately 30 seconds after occlusion release, an undershoot in Pi peak height to 494 Circulation Research Vol 67, No 2, August 1990 TABLE 2. Metabolic Changes During and Following 24 Seconds of Coronary Occlusion -3-ATP PCr/Pi PCr Time (sec) Pi Downloaded from http://circres.ahajournals.org/ by guest on June 16, 2017 Occlusion 0-4.8 4.8-9.6 9.6-14.4 14.4-19.2 19.2-24 Recovery 0-4.8 4.8-9.6 9.6-14.4 14.4-19.2 19.2-24 24-28.8 28.8-33.6 33.6-38.4 38.4-43.2 43.2-48 48-52.8 52.8-57.6 57.6-62.4 1.00+0.03 0.88±0.02 0.80±0.05 0.75 ±0.03* 0.66±0.03* 0.99±0.09 1.07±0.05 1.31±0.09 1.04±0.10 0.81±0.04 0.62±0.07* 1.05±0.03 0.95±0.05 0.95±0.06 1.53±0.14 1.70±0.08* 0.51±0.06* 0.38±0.03* 0.89±0.05 0.95+0.07 0.38±0.05* 0.47±0.05* 0.68±0.04* 0.90±0.06 1.07±0.13 1.06±0.08 1.19±0.14 1.38±0.16 1.12±0.08 1.26±0.07 1.29±0.12 1.30±0.13 1.15±0.08 Metabolite levels are normalized to their respective control values. pH values are Pi, inorganic phosphate. 0.66±0.04* 0.71±0.04* 0.90±0.04 0.97+0.03 1.03±0.07 1.08±0.04 1.06±0.04 1.04±0.04 0.96±0.06 0.96±0.03 1.07±0.05 1.06±0.03 1.05±0.03 1.77±0.15* 1.52±0.10* 1.28±0.04* 1.07±0.06 0.96±0.05 1.01±0.08 0.91±0.07 0.80±0.09 0.86±0.08 0.75±0.03* 0.83±0.07 0.85±0.08 0.89±0.04 pH 7.15±0.06 7.19±0.05 7.18±0.03 7.15±0.04 0.81±0.05 7.23±0.05 0.89±0.04 7.18±0.02 1.02±0.07 0.98±0.06 1.01±0.04 1.05±0.07 0.98±0.05 0.86±0.04 1.00±0.03 0.97±0.05 1.05+0.11 1.07±0.04 1.03±0.06 absolute. PCr, phosphocreatine; *p<0.05 compared with control, using Student's t test with Bonferroni's correction for multiple observations. approximately 10-20% below control was noted; concomitantly, segment shortening demonstrated an overshoot to approximately 5-10% above control (Figure 2, Table 3). This pattern was observed in seven of the eight pigs studied. End-diastolic and end-systolic segment lengths were slightly shorter than control during this period. The overshoot in segment shortening and the undershoot of Pi per- TABLE 3. Segment Shortening and Coronary Blood Flow During and Following 24 Seconds of Coronary Occlusion Cor Q EDL ESL SS Time (sec) Occlusion 0-4.8 4.8-9.6 9.6-14.4 14.4-19.2 19.2-24 1.00±0.07 1.02±0.08 1.04±0.08* 1.05±0.08* 1.06±0.08* 1.00+0.09 1.06±0.10* 1.15±0.11* 1.20±0.11* 1.25±0.11* 0.99+0.01 0.84±0.05 0.55±0.09* 0.41±0.09* 0.25±0.09* 0 0 0 0 0 Recovery 2.18±0.20* 0.38±0.09* 2.84±0.21* 0.65±0.08* 2.95±0.22* 0.90±0.06* 2.72±0.23* 1.04±0.04 2.48±0.24* 1.06±0.03 2.25±0.24* 1.07±0.04 2.00+0.21* 0.96+0.07 1.06+0.04 0.95+0.08* 1.80±0.19* 1.06±0.03 33.6-38.4 0.95±0.08* 0.96±0.07 1.68±0.18 1.06±0.04 38.4-43.2 0.95±0.08* 0.96±0.07 1.56±0.20 1.06±0.03 0.94±0.08* 43.2-48 0.96±0.07 1.43±0.17 1.05+0.03 48-52.8 0.95±0.08* 0.96±0.07 1.38±0.15 1.09+0.03 52.8-57.6 0.95±0.08* 0.96±0.07* 1.36±0.14 57.6-62.4 1.08±0.03 0.96±0.07 0.95±0.08* All values are normalized to their respective controls. EDL, end-diastolic length (mean control, 10.8 mm); ESL, endsystolic length (mean control, 8.7 mm); SS, segment shortening (mean control, 19.3%); Cor Q, coronary blood flow 0-4.8 4.8-9.6 9.6-14.4 14.4-19.2 19.2-24 24-28.8 28.8-33.6 1.06+0.08* 1.04±0.08* 1.01±0.08 0.99±0.07 0.98±0.07 0.97±0.07 1.22±0.11* 1.13±0.10* 1.03±0.09 0.98±0.08 0.96±0.08 0.95±0.08* velocity. *p<0.05 compared with control, using Student's t test with Bonferroni's correction for multiple observations. 495 Schwartz et al Dynamic Changes of Myocardial Function and Energy Metabolism 3 - - * 2 0 0 + 0 0 O.. PCr Pi ATP -.. SS A ....Cor Flow 0 n~~~~~~~~~ . 0 20 40 60 Time (seconds) FIGURE 2. Changes in phosphorus metabolite peak heights and systolic segment shortening during and following 24 seconds of coronary occlusion. Combined data from eight pigs. Values are normalized to their respective preocclusion control levels. Standard errors for the data are listed in Tables 1 and 2. Note lack of change of systolic shortening (SS) or phosphorus metabolites during the first 4.8 seconds of occlusion, rapid recovery of SS and metabolites following occlusion release, and late undershoot of inorganic phosphate (Pi). PCr, phosphocreatine; Cor Flow, coronary Downloaded from http://circres.ahajournals.org/ by guest on June 16, 2017 flow. sisted at the end of the data collection period (62 seconds after occlusion release), but resolved fully during the 5-minute interval between occlusions. During occlusion, the dynamic changes in normalized segment shortening exhibited a strong, linear correlation with those of normalized PCr (slope=2.3, r2=0.97, SEE=0.06), Pi (slope=-1.1, r2=0.96, SEE= 0.06), 1/Pi (slope= 1.7, r2=0.98, SEE=0.03), and PCr/ Pi (slope=1.2, r2=0.98, SEE=0.05). There was a poor correlation between changes in segment shortening and ATP (slope=3.8, r2=0.51, SEE=0.25). These relations are shown in Figure 3. When the data from individual pigs were analyzed separately, similar linear relations were found, with average r2 values of 0.84 (segment shortening versus PCr), 0.74 (versus 1/Pi), 0.78 (versus PCr/Pi), and 0.31 (versus ATP). During the recovery from ischemia, the relations between segment shortening and PCr and Pi became curvilinear. At a given level of phosphorus metabolites, segment shortening was greater during the recovery from, as compared with the onset of, ischemia. Figure 4 and Tables 2 and 3 illustrate the relation between segment shortening and PCr/Pi. For example, during the interval 14.4-19.2 seconds of occlusion, normalized PCr/Pi was 0.51 and normalized segment shortening was 0.41. During the interval 4.8-9.6 seconds of recovery, PCr/Pi was similar at 0.47, but segment shortening was considerably greater at 0.65. Analysis of data from individual pigs revealed a similar "hysteresis loop" in six of the eight animals. There were no significant differences in calculated intracellular pH between points on the descending and ascending limbs of the hysteresis loop. Discussion Dynamic Relation Between Contractile Function and Energy Metabolism This study demonstrates that pronounced changes in myocardial contractile function and high-energy phosphate metabolism occur synchronously during brief coronary occlusion. With a time resolution of 4.8 seconds, dynamic changes in regional segment shortening bore a strong, linear relation to concomitant changes in PCr, Pi, and the PCr/Pi ratio. As in all studies of myocardial function and energy metabolism during ischemia, this investigation cannot prove a causal relation between variables that change in parallel. Such proof awaits an experimental intervention that alters the intracellular level of one phosphorus metabolite without producing the other metabolic effects of ischemia. However, the extremely close temporal correlation between direct measures of mechanical function and energy metabolism demonstrated in this study is consistent with a causal relation. Several mechanisms have been proposed to explain the impairment of cardiac contractility by perturbed myocardial energy metabolism during (I) to * * 0 0 E a Pcr/Pi PCr 1/Pi ATP z 0.0 0.2 0.4 0.6 0.8 1.0 1.2 Normalized peak height FIGURE 3. Relation between systolic segment shortening (SS) and phosphorus metabolites during 24 seconds of coronary occlusion. Combined data from eight pigs. Time intervals after release of occlusion are indicated to the left of data. Note that changes in SS are strongly correlated with those of phosphocreatine (PCr) (r2=0.97), l/inorganic phosphate (Pi) (r2=0.99), and PCr/Pi (r2=0.98), but notATP (r2=0.51). 496 Circulation Research Vol 67, No 2, August 1990 1.2 - recovery 1.0 - I_ , control (I) U') 0.8 - 10 0.6 - ._! 0 E 0 z ..' .," i; ,t: T/ Occlusion * - - -0 -- 0.4 0.2 - 19.2-24 sec occlusion . n 0.v n -0_ 0.0 0.2 0.4 0.6 0.8 1.0 1.2 Reflow FIGURE 4. Relation between systolic segment shortening (SS) and ratio of phosphocreatine to inorganic phosphate (PCr/ Pi) during and following coronary occlusion. Combined data from eight pigs. Data are nornalized to preocclusion control values. Arrows indicate temporal sense of relation. Note that at a given level of PCr/Pi, SS is greater during the recovery than the onset of ischemia. 1.4 Normalized PCr/Pi Downloaded from http://circres.ahajournals.org/ by guest on June 16, 2017 ischemia. These include an inhibitory effect of Pi on myofilament calcium sensitivity,21,22 a depressant effect of intracellular acidosis23 or lactate,24 or a reduction in the cytosolic phosphorylation potential ([ATP]/[ADP][P,])25 or free energy of ATP hydrolysis.26 For any of these factors to be considered a likely regulator of contractile function during ischemia, changes in that factor should occur in advance of, or simultaneously with, changes in contractility. In addition, changes in contractile function should be strongly correlated with changes in the proposed regulating factor. Clarke et a19 examined the dynamic relation between energy metabolism and contractile function (assessed by rate-pressure product) in isolated, perfused rat hearts during and following 2 minutes of global ischemia. High-energy phosphate metabolites and pH were measured by 31P NMR with a 10-second time resolution. These investigators concluded that the cytosolic phosphorylation potential controls myocardial contractile function, because this was the only metabolic parameter to decline at a rate faster than the rate-pressure product during ischemia. However, the rate-pressure product is an indirect index of contractile state, and data from isolated, crystalloidperfused hearts may not fully apply to the regulation of contractility in the in situ, blood-perfused heart. In the current experiment, segment shortening was strongly and synchronously related to changes in PCr, Pi, and the PCr/Pi ratio (which has been used as an approximation to the phosphorylation potential27). During the first 4.8 seconds of occlusion, no changes were detected either in segment shortening or in any of the phosphorus metabolites. This brief period of metabolic and functional stability may reflect the utilization of residual oxygen in capillary blood or oxygen stored in myocardial myoglobin.28 The decline in segment shortening that occurred during the remainder of the occlusion period bore a strong linear relation to the simultaneous changes in PCr (r2=0.97), Pi (r2=0.96), and PCr/Pi (r2=0.98). The precise temporal correspondence of these functional and metabolic changes suggests that alterations in high-energy phosphate metabolism rapidly and directly regulate cardiac contractility at the onset of myocardial ischemia; however, it must be reemphasized that temporal correspondence is not proof of a causal relation. The hypothesis is further supported by a study of contractile function during steady-state, graded ischemia in the pig, performed in this laboratory.29 Under these disparate experimental conditions, the relation between segment shortening and PCr/Pi was nearly identical to that determined in the present study, suggesting that the same regulatory mechanism was operative. However, because ischemia causes concomitant changes in several parameters of energy metabolism, it is difficult to assign the primary regulatory role to the changes in Pi, the phosphorylation potential, or the free energy of ATP hydrolysis. In the current study, the marked decline in contractile function during ischemia was accompanied by small, poorly correlated changes in the f3-ATP resonance. This result corroborates previous findings that ATP depletion is prevented during brief periods of ischemia by utilization of PCr via the creatine kinase reaction.1,7,913 It is also unlikely that intracellular acidosis mediated the pronounced decline in segment shortening that occurred during 24 seconds of ischemia. The change in pH was most likely quite small, because the pH values calculated from the chemical shift of Pi during ischemia and early recovery (7.15-7.23, Table 2) are similar to those determined under nonischemic conditions in other experimental models.6"19'30'31 In addition, the consumption of protons accompanying the breakdown of 34% of myocardial PCr by the creatine kinase reaction can be estimated to have raised intramyocardial pH by 0.04 units.3 Other investigators32 have found that intracellular pH actually rises during an initial period of hypoxia via this mechanism. Why Is the Relation Between Contractility and Phosphorus Metabolites Altered During Reactive Hyperemia? An important finding of this study is that the relation between segment shortening and phosphorus metabolites is different during the recovery from ischemia compared with the onset of ischemia. At a given level of PCr, Pi, or PCr/Pi ratio, segment Schwartz et al Dynamic Changes of Myocardial Function and Energy Metabolism Downloaded from http://circres.ahajournals.org/ by guest on June 16, 2017 shortening was greater during the recovery from ischemia than during the onset of ischemia. The relation between segment shortening and phosphorus metabolites is therefore not unique and most likely reflects a multifactorial regulation of contractility, with changes in another, independent factor during the recovery from brief ischemia. This additional independent factor may be the increased coronary blood flow rate during reactive hyperemia. In 1963, Gregg'4 described an augmentation of systolic performance and oxygen consumption resulting from an increase of coronary flow and/or perfusion pressure to supranormal levels. The Gregg phenomenon has been demonstrated in experimental models in which coronary flow is increased by pharmacological vasodilation, without an increase in perfusion pressure.33 The most widely proposed mechanism for the phenomenon is that mechanical distension of myofibrils by adjacent distended intramyocardial vessels34,35 leads to increased contractility, perhaps by a localized Frank-Starling mechanism. An alternative explanation is that a surfeit of myocardial oxygen delivery leads to increased oxygen utilization,36 improved energetics with an increased phosphorylation potential, and augmented ATP utilization for contractile activity. In the present study, contractile function during early reactive hyperemia exceeded that during ischemia, under comparable metabolic conditions (Figure 4). Therefore, the wide, initial portion of the hysteresis loop cannot be explained by a change in energetic state, but may reflect a direct augmentation of contractility by the elevated coronary blood flow of reactive hyperemia via the Gregg phenomenon. In addition, the final recovery points in Figure 4 cluster at a normalized segment shortening value of approximately 1.1 and a normalized PCr/Pi value of approximately 1.2. This overshoot could reflect an augmentation of the phosphorylation potential by excess myocardial oxygen delivery during reactive hyperemia, with a resultant increase in contractility. Pagani et a137 also observed an overshoot in contractile function following 1 minute of ischemia in dogs, which could be abolished by preventing reactive hyperemic coronary flow from exceeding the control flow rate. Stahl et a138 found that dipyridamole- or papaverine-induced hyperemia augmented segment shortening and shifted the end-systolic pressurelength relation to the left in stunned canine myocardium. While energy metabolism was not assessed in these studies, the results also support enhancement of postischemic contractile function by an elevated coronary blood flow rate. In the isolated ferret heart, Kihara et a139 recently found that abnormal intracellular calcium transients and left ventricular dysfunction produced by ischemia could be partially reversed by hypoxic perfusion. These data suggest that coronary blood flow, even in the absence of oxygen delivery, may modulate contractility after ischemia via an effect on calcium transients. 497 Several alternative explanations for the hysteresis loop appearance of Figure 4 must be considered. A macroscopic Frank-Starling mechanism does not explain the results, because end-diastolic segment length was not different during ischemia versus recovery at a given level of PCr/Pi (Table 3). Similarly, calculated intracellular pH was not significantly different between the two limbs of Figure 4. Lower levels of lactate (because of washout) during recovery as compared with the onset of ischemia are unlikely for two reasons. First, no pH differences are apparent at comparable PCr/Pi levels. Second, the decline in myocardial lactate during reactive hyperemia has been shown to be slower than its increase during brief ischemia in dogs.13 It is possible that autonomic reflexes, such as activation of baroreceptors by the small decline in arterial pressure during occlusion, increased the activity of cardiac sympathetic nerves during reactive hyperemia, with a consequent increase in contractility. However, it is unlikely that a 4-7 mm Hg fall in arterial pressure could provoke a 25-45% increase in segment shortening (at a given PCr/Pi) by sympathetic activation. In the study by Pagani et al,37 the postischemic overshoot in contractility was not affected by P-blockade with propranolol or depletion of catecholamine stores with reserpine. Differences in intramyocardial blood volume between occlusion and reflow must also be considered in interpreting the data of Figure 4. Enddiastolic intramyocardial blood volume has been estimated to constitute 13% of myocardial mass in anesthetized dogs, and this percentage may increase with vasodilation.40 Conversely, when arterial inflow is occluded, intramyocardial blood volume falls because of systolic compression of capacitance vessels.41 Increased intramyocardial blood volume during reflow, compared with occlusion, could cause an apparent increase in Pi (and an apparent decrease in PCr/Pi) because of a larger superimposed signal from intramyocardial 2,3-DPG. Thus, some of the hysteresis in Figure 4 could reflect a leftward shift of the ascending limb because of increased intramyocardial blood volume. It is doubtful, however, that this is the sole explanation for the hysteresis. A plot of normalized segment shortening versus normalized PCr (Figure 5) also exhibits hysteresis, though somewhat less prominently than Figure 4. Because blood contains no PCr, the hysteresis evident in Figure 5 is not readily explained by changes in intramyocardial blood volume. Finally, changes in left ventricular wall thickness beneath the surface coil must be considered in explaining the hysteresis relation of Figures 4 and 5, since left ventricular wall thickness may vary with changes in coronary blood flow.42 NMR acquisitions were gated to end-diastole to minimize the effects of changing left ventricular wall thickness during and following ischemia. A change in wall thickness would reciprocally alter the amount of myocardium and intracavitary blood contained within the sensitive volume of the coil, possibly affecting the determination of 498 Circulation Research Vol 67, No 2, August 1990 1.2 - recovery 2 1.0- _i control C') UI) 0.8- / "'i. 10 N -*- 0.6- --- *-- E 0 0.4 - z 19.2-24 sec occlusion 0.2 Occlusion Reflow FIGURE 5. Relation between systolic segshortening (SS) and phosphocreatine (PCr) during and following coronary occlusion. Combined data from eight pigs. See Figure 4 for explanation. ment 0.0 0.4 0.6 0.8 1.0 1.2 Normalized PCr Downloaded from http://circres.ahajournals.org/ by guest on June 16, 2017 metabolite levels. To determine the extent of changes in end-diastolic wall thickness in this experiment, two pigs were instrumented with a pair of wall thickness crystals adjacent to the surface coil. In these pigs, end-diastolic wall thickness remained between 100% and 104% of control during all occlusion and recovery time intervals. Such small changes in end-diastolic wall thickness are unlikely to have significantly affected the relations shown in Figures 4 and 5. Limitations Metabolic measurements made by 31P NMR. While 31P NMR is a powerful tool for the repetitive, nondestructive assessment of myocardial energy metabolism, it has several important limitations. Because the sensitive volume of the surface coil includes some intracavitary ventricular blood, spectral overlap between 2,3-DPG and Pi is unavoidable. This overlap reduces the accuracy of Pi peak height and chemical shift determinations, especially under nonischemic conditions, when Pi is low. Although the Pi peak was clearly discernible during ischemia and early recovery, conclusions regarding the role of Pi and pH in the regulation of contractility must be tempered by this limitation. 31P NMR cannot provide a direct measurement of free ADP, one of the determinants of the phosphorylation potential and the free energy of ATP hydrolysis. In addition, the interaction between changes in high-energy phosphate metabolism and intracellular calcium levels cannot be assessed with this technique. Regional heterogeneity of metabolic and functional Both the metabolic and functional of the myocardium to ischemia may vary transmurally. Therefore, it is desirable to obtain these measurements from the same transmural layer of the left ventricle. NMR techniques capable of precise spatial localization usually require long data acquisition times. In the present experiment, such techniques would have required averaging data from an inordinately large number of interventions to achieve the desired temporal resolution with an acceptable signal-to-noise ratio. However, the subendocardial NMR signal was weighted more heavily than the subepicardial signal by choosing a pulse responses. responses length that provided maximal sensitivity in the subendocardium. Concomitantly, segment shortening was measured in the subendocardial layer. On the left ventricular surface, the ischemic region was large compared with the area encompassed by the coil and dimension gauges, making substantial lateral contamination of the NMR signal from nonischemic myocardium unlikely. Therefore, metabolic and functional correlations should be possible in this experimental model, despite the constraints of spatial heterogeneity. Effects of anesthesia. Arterial pressure in this group of pigs was somewhat depressed, especially in those receiving halothane and sodium pentobarbital. However, the low left ventricular end-diastolic pressures and normal segment shortening8 suggest peripheral vasodilation, rather than significant myocardial depression. In addition, control phosphorus spectra showed no evidence of metabolic impairment, and the 300% increase in peak reactive hyperemic flow implies that coronary vasodilator reserve was far from exhausted. The two pigs anesthetized with chloralose demonstrated similar responses during ischemia to those of the pigs anesthetized with halothane and pentobarbital (nadir segment shortening, 31% of control; nadir PCr, 66% of control; peak Pi, 184% of control; segment shortening versus PCr, r=0.81; segment shortening versus Pi, r= -0.81). In addition, both chloralose-anesthetized pigs demonstrated "hysteresis" in these relations during recovery. These data suggest that the results of the current study do not depend strongly on the type of anesthetic used. Summary Numerous previous studies have supported the regulation of myocardial contractile function during ischemia by changes in high-energy phosphate metabolism, based on measurements made under steady-state or slowly changing conditions. However, the most pronounced changes in both contractile function and energy metabolism occur early in ischemia. In this study, a precise temporal correspondence was demonstrated between changes in contractile function and PCr, Pi, and PCr/Pi in the in situ Schwartz et al Dynamic Changes of Myocardial Function and Energy Metabolism pig heart subjected to brief coronary occlusion. These data strongly support a direct regulatory relation between high-energy phosphate metabolism and contractile function during ischemia. During reactive hyperemia, increased coronary blood flow may independently augment contractility. This study also demonstrates that high temporal resolution NMR data can be obtained in a large, intact animal preparation at moderate field strength, with spectral reconstruction from multiple, reproducible interventions. Acknowledgments We sincerely thank Gerald Matson, PhD; Donald Twieg, PhD; Mr. James Buchanan; and Alexander Smekal, MD; for technical advice and assistance. References Downloaded from http://circres.ahajournals.org/ by guest on June 16, 2017 1. Covell JW, Pool PE, Braunwald E: Effects of acutely induced ischemic heart failure on myocardial high energy phosphate stores. Proc Soc Exp Biol Med 1967;124:126-131 2. Braasch W, Gudbjarnason S, Puri PS, Ravens KG, Bing RJ: Early changes in energy metabolism in the myocardium following acute coronary artery occlusion in anesthetized dogs. Circ Res 1968;23:429-438 3. Allen DG, Orchard CH: Myocardial contractile function during ischemia and hypoxia. Circ Res 1987;60:153-168 4. Liedtke AJ, Hughes HC, Neely JR: Metabolic responses to varying restrictions of coronary blood flow in swine. Am J Physiol 1975;228:655-662 5. Bittl JA, Balschi JA, Ingwall JS: Contractile failure and high-energy phosphate turnover during hypoxia: 3`P-NMR surface coil studies in living rat. Circ Res 1987;60:871-878 6. Guth BD, Martin JF, Heusch G, Ross J Jr: Regional myocardial blood flow, function and metabolism using phosphorus-31 nuclear magnetic resonance spectroscopy during ischemia and reperfusion in dogs. JAm Coll Cardiol 1987;10:673-681 7. Camacho SA, Lanzer P, Toy BJ, Gober J, Valenza M, Botvinick EH, Weiner MW: In vivo alterations of high-energy phosphates and intracellular pH during reversible ischemia in pigs: A 31P magnetic resonance spectroscopy study. Am Heart J 1988;116:701-708 8. Theroux P, Franklin D, Ross J Jr, Kemper WS: Regional myocardial function during acute coronary occlusion and its modification by pharmacologic agents in the dog. Circ Res 1974;35:896-908 9. Clarke K, O'Connor AJ, Willis RJ: Temporal relation between energy metabolism and myocardial function during ischemia and reperfusion. Am J Physiol 1987;253:H412-H421 10. Farmer D, Lipton MJ, Higgins CB, Ringertz H, Dean PB, Sievers R, Boyd DP: In vivo assessment of left ventricular wall and chamber dynamics during transient myocardial ischemia using cine computed tomography. Am J Cardiol 1985; 55:560-565 11. Wohlgelernter D, Cleman M, Highman HA, Fetterman RC, Duncan JS, Zaret BL, Jaffe CC: Regional myocardial dysfunction during coronary angioplasty: Evaluation by twodimensional echocardiography and 12-lead electrocardiography. JAm Coll Cardiol 1986;7:1245-1254 12. Carlson EB, Hinohara T, Morris KG: Recovery of systolic and diastolic left ventricular function after a 60-second coronary arterial occlusion during percutaneous transluminal coronary angioplasty for angina pectoris. Am J Cardiol 1987;60:460-466 13. Dunn RB, McDonough KM, Griggs DM Jr: High energy phosphate stores and lactate levels in different layers of the canine left ventricle during reactive hyperemia. Circ Res 1979;44:788-795 14. Gregg DE: Effect of coronary perfusion pressure or coronary flow on oxygen usage of the myocardium. Circ Res 1963; 13:497-500 499 15. Feigl EO: Coronary physiology. Physiol Rev 1983;63:1-205 16. Streeter DD Jr, Spotnitz HM, Patel DJ, Ross J Jr, Sonnenblick EH: Fiber orientation in the canine left ventricle during diastole and systole. Circ Res 1969;24:339-347 17. Gordon RE, Timms WE: An improved tune and match circuit for BO shimming in intact biological systems. J Magn Reson 1982;54:323-324 18. Lawry TJ, Karczmar GS, Weiner MW, Matson GB: Computer simulation of MRS localization experiments: An analysis of ISIS. J Magn Reson 1989;9:299-314 19. Kusuoka H, Inoue M, Yutaka T, Watari H, Hori M, Abe H: Augmented energy consumption during early systole as a mechanism of cyclical changes in high-energy phosphates in myocardium assessed by phosphorus nuclear magnetic resonance. Jpn Circ J 1985;49:1099-1107 20. Miller RG: Simultaneous Statistical Inference. New York, McGraw-Hill Book Co, 1966, pp 116-126 21. Kentish JC: The effects of inorganic phosphate and creatine phosphate on force production in skinned muscles from rat ventricle. J Physiol (Lond) 1986;370:585-604 22. Kusuoka H, Weisfeldt ML, Zweier JL, Jacobus WE, Marban E: Mechanism of early contractile failure during hypoxia in intact ferret heart: Evidence for modulation of maximal Ca+2activated force by inorganic phosphate. Circ Res 1986; 59:270-282 23. Steenbergen C, Deleeuw G, Rich T, Williamson JR: Effects of acidosis and ischemia on contractility and intracellular pH of rat heart. Circ Res 1977;41:849-858 24. Yatani A, Fujino T, Kinoshita K, Goto M: Excess lactate modulates ionic currents and tension components in frog atrial muscle. J Mol Cell Cardiol 1981;13:147-161 25. Gibbs C: The cytoplasmic phosphorylation potential: Its possible role in the control of myocardial respiration and cardiac contractility. J Mol Cell Cardiol 1985;17:727-731 26. Kammermeier H: High energy phosphate of the myocardium: Concentration versus free energy change. Basic Res Cardiol 1987;82:31-36 27. Chance B, Eleff S, Leigh JS, Sokolow D, Sapega A: Mitochondrial regulation of phosphocreatine/inorganic phosphate ratios in exercising human muscles: A gated 31P NMR study. Proc NatlAcad Sci USA 1981;78:6714-6718 28. Olsson RA: Kinetics of myocardial reactive hyperemia blood flow in the unanesthetized dog. Circ Res 1964;14(suppl I): 1-81-1-85 29. Schaefer S, Schwartz GG, Gober JR, Wong AK, Camacho SA, Massie B, Weiner MW: Relationship between myocardial metabolites and contractile abnormalities during graded regional ischemia: 31P NMR studies of porcine myocardium in vivo. J Clin Invest 1990;85:706-713 30. Brindle KM, Rajagopalan B, Williams DS, Detre JA, Simplaceanu E, Ho C, Radda GC: 31P NMR measurements of myocardial pH in vivo. Biochem Biophys Res Commun 1988; 151:70-77 31. Katz LA, Swain JA, Portman MA, Balaban RS: Intracellular pH and inorganic phosphate content of heart in vivo: A 31P NMR study. Am J Physiol 1988;255:H189-H196 32. Allen DG, Morris PG, Orchard CH, Pirolo JS: A nuclear magnetic resonance study of metabolism in the ferret heart during hypoxia and inhibition of glycolysis. J Physiol (Lond) 1985:361:185-204 33. Abel RM, Reis RL: Effects of coronary blood flow and perfusion pressure on left ventricular contractility in dogs. Circ Res 1970;27:961-971 34. Arnold G, Kosche F, Meissner E, Nietzart A, Lochner W: The importance of the perfusion pressure in the coronary arteries for the contractility and the oxygen consumption of the heart. Pflugers Arch 1968;299:339-356 35. Poche R, Arnold G, Gahlen D: The influence of coronary perfusion pressure on metabolism and ultrastructure of the arrested, aerobically perfused isolated guinea pig heart. Virchows Arch [B] 1971;8:252-266 36. Bachaner MB, Lioy F, Visscher MB: Coronary blood flow, oxygen delivery rate and cardiac performance. JPhysiol (Lond) 1971;216:111-127 500 Circulation Research Vol 67, No 2, August 1990 37. Pagani M, Vatner SF, Baig H, Braunwald E: Initial myocardial adjustments to brief periods of ischemia and reperfusion in the conscious dog. Circ Res 1978;43:83-92 38. Stahl LD, Aversano TR, Becker LC: Selective enhancement of function of stunned myocardium by increased flow. Circulation 1986;74:843-851 39. Kihara Y, Grossman W, Morgan JP: Direct measurement of changes in intracellular calcium transients during hypoxia, ischemia, and reperfusion of the intact mammalian heart. Circ Res 1989;65:1029-1044 40. Hoffman EA, Ritman EL: Intramyocardial blood volume: Implication for analysis of myocardial mechanical characteristics via in vivo imaging of the heart, in Sideman S, Beyar R (eds): Activation, Metabolism and Perfusion of the Heart. Dordrecht, Netherlands, Martinus Nijhoff, 1987, pp 421-432 41. Kajiya F, Tsujioka K, Goto M, Wada Y, Chen XL, Nakai M, Tadaoka S, Hiramatsu 0, Ogasawara Y, Mito K, Tomonaga G: Functional characteristics of intramyocardial capacitance in the dog. Circ Res 1986;58:476-485 42. Gaasch WH, Bernard SA: The effect of acute changes in coronary blood flow on left ventricular end-diastolic wall thickness. Circulation 1977;56:593-598 KEY WORDS * energy metabolism * contractility * reactive hyperemia * magnetic resonance spectroscopy Downloaded from http://circres.ahajournals.org/ by guest on June 16, 2017 Dynamic relation between myocardial contractility and energy metabolism during and following brief coronary occlusion in the pig. G G Schwartz, S Schaefer, D J Meyerhoff, J Gober, P Fochler, B Massie and M W Weiner Downloaded from http://circres.ahajournals.org/ by guest on June 16, 2017 Circ Res. 1990;67:490-500 doi: 10.1161/01.RES.67.2.490 Circulation Research is published by the American Heart Association, 7272 Greenville Avenue, Dallas, TX 75231 Copyright © 1990 American Heart Association, Inc. All rights reserved. Print ISSN: 0009-7330. Online ISSN: 1524-4571 The online version of this article, along with updated information and services, is located on the World Wide Web at: http://circres.ahajournals.org/content/67/2/490 Permissions: Requests for permissions to reproduce figures, tables, or portions of articles originally published in Circulation Research can be obtained via RightsLink, a service of the Copyright Clearance Center, not the Editorial Office. Once the online version of the published article for which permission is being requested is located, click Request Permissions in the middle column of the Web page under Services. Further information about this process is available in the Permissions and Rights Question and Answer document. Reprints: Information about reprints can be found online at: http://www.lww.com/reprints Subscriptions: Information about subscribing to Circulation Research is online at: http://circres.ahajournals.org//subscriptions/