Survey
* Your assessment is very important for improving the work of artificial intelligence, which forms the content of this project
Electrocardiography wikipedia , lookup
Heart failure wikipedia , lookup
Coronary artery disease wikipedia , lookup
Mitral insufficiency wikipedia , lookup
Lutembacher's syndrome wikipedia , lookup
Cardiac surgery wikipedia , lookup
Myocardial infarction wikipedia , lookup
Hypertrophic cardiomyopathy wikipedia , lookup
Antihypertensive drug wikipedia , lookup
Arrhythmogenic right ventricular dysplasia wikipedia , lookup
Quantium Medical Cardiac Output wikipedia , lookup
Atrial septal defect wikipedia , lookup
Dextro-Transposition of the great arteries wikipedia , lookup
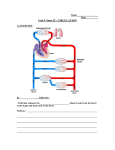
Circulation in the Three - Chambered Snake Heart By K J E L L JOHANSEN, CAND. READ. Our knowledge on the physiologic evolution of the cardiovascular system is fragmentary. The reptiles and the snakes in particular represent a stage where our knowledge is especially poor. The present study deals with the circulatory dynamics of the snake heart. It may bo suggested that the snake heart effectively prevents extensive mixing of arterial and venous blood. The durations of ventricular filling and contraction differ "•reatlv from that in the mammalian heart. 5 specimens of the viper, Vipera benis. The grass snakes weighed from 150 to 220 Gin. and the vipers approximately 100 Gra, The length of the snakes ranged from 60 to 130 cm. The snakes were placed in polyvinyl tubes, slightly longer than the animals. The inner diameter of the tubes was likewise slightly larger than that of the animals to allow for unrestricted respiratory movements. The snakes were fastened to the tubes by needles through the tube wall and the skin of the animals. The operations necessary to dissect free the heart and vessels were performed through windows in the poly-vinyl tube. Following this procedure the animals were always well confined. Electrocardiograms were recorded by means of needle electrodes placed subcutanously in the animals. One electrode was placed near the heart in cranial position while the other was placed near the cloacal opening. The pressures in the heart and vessels were obtained either through small cannulas (18 gage) or catheters (P.E. 50, 0.5S mm. I. D. 0.965 mm. O. D. 20 cm. long) and recorded by means of Statham pressure transducers on a Sanborn recorder. The pressure transducers were always placed level with the tip of the catheter and level served as the zero of the manometers. In several experiments an indirect, nontraumatizing method for recording of blood pressure was used.2 Relative variations in pressure were obtained by photoelectric recording of variations in the short axis of the vessel cross-section during slight compression of the vessel. The method has practically no instrumental phase lag regarding the beginning of pressure rise and is of great value for timing of dynamic circulatory events. In order to avoid changes in body temperature, the experiments were performed in a temperature stabilized room. T Downloaded from http://circres.ahajournals.org/ by guest on June 14, 2017 H E cardiovascular system in reptiles covers an important stage in the physiologic phylogeny of circulation. The three-chambered reptilian heart (crocodiles excepted) has a complete atrial septum while the single ventricle is subdivided by an incomplete septum. The conus arteriosus is absent and the arterial trunks have separate origins from the lumen of the ventricle. 1 The amphibian heart, on the other hand, has an atrial septum which in most, cases is perforated. The ventricle has no septum and the ventricular wall has developed several muscular trabeculae giving it a cavernous structure. The blood leaves the ventricle through the conus arteriosus which is followed by a bipartient truncus arteriosus. The ventral part of this truncus leads to the pulmonary artery, while the dorsal part gives rise to the aorta. Thus the reptilian heart is distinctly advanced over the amphibian heart, and shows an obvious approach toward a condition found in the higher amniotes. However, there is no complete separation of the pulmonary and systemic circuits. The present study seems to be the first one dealing with the dynamics of circulation in the snake heart which, among the reptiles, is specially well suited to studies of this kind. The elongated body form, with the visceral organs lying one after another, make special demands on the heart capacity. METHODS RESULTS The material used consists of 14 specimens of the common grass snake, Tripoclonotus natrix, and The blood pressure in the left and dorsal aorta showed relatively great individual variations. Thus the highest values of systemic blood pressure recorded during the experi- From the University Institute for Experimental Medical Research, Ullevaal Hospital, Oslo, Xorway. Received for publication April 9, 1959. 828 Circulation Research, Volume VII, November 1959 THKEE-CHAMBERED SNAKE HEAET ECG T P 829 T P. mmHg ECG 45 40 mmHg 35 ECG I Downloaded from http://circres.ahajournals.org/ by guest on June 14, 2017 FIG. 1 Top. Blood pressure record from the left aorta of a grass-snake, Tripodonotus natrix, weight 170 Gni., length 130 cm. Time marTcs, 1 sec. FIG. 2 Middle. Blood pressure record from the pulmonary artery of a grass-snake, weight 170 Gm., length 130 cm. Time marks, 1 sec. ments were 80 mm. Hg systolic and 66 mm. Hg diastolic, with a pulse pressure of 14 mm. Hg. More often, the blood pressure ranged from 50 to 60 mm. Hg systolic and 40 to 50 diastolic. Figure 1 shows a representative tracing of blood pressure recorded in the left aorta. The notable features are the extraordinarily long Q-T duration and the short T-P interval in the electrocardiogram. The pulse contour is similar to that seen in mammals. A pressure record from the pulmonar}artery is shown in figure 2. The diastolie pressure is lower than the systemic diastolic, while the systolic pressures of both show only small differences. Thus the pulmonary pulse pressure is approximately 20 mm. Hg or about twice as large as that from the left aorta. The pulmonary artery pulse curve shows an earlier start than that recorded from the left aorta (fig. 3). The difference in time is about 0.04 to 0.05 sec. This finding was reinvestigated with the photoelectric method and the same difference in time was encountered. Likewise the Statham manometers were interchanged in order to exclude differences in the manometric systems. Thus this finding rests on a physiologic basis, rather than being an instrumental artifact. 1 second FIG. 3 Bottom. Blood pressures in the left aorta and pulmonary artery of a snake. Note the earlier start of pressure rise in the pulmonary artery. The efficiency of the pulmonary and aortic valves was tested by retrograde infusion of saline into the heart. These experiments revealed a high degree of valvular efficiency in the snake heart. The intracardiac pressures are shown in figure 4. The intraventricular pressure ranged between 40 to 70 mm. Hg systolic and approximately zero diastolic. A ventricular period of diastasis is absent and this finding correlates with the very long Q-T interval. The atrial pressures ranged from 3 to 5 mm. Hg systolic and slightly above zero diastolic. The systolic pressure in the left atrium was on several occasions higher than that of the right atrium. The systolic pressures in the left and right aorta were essentially similar. The aortic pressure records show a slow diastolic gradient which points toward a considerable amount of peripheral resistance. The period of isometric contraction (fig. 5) varied between 0.08 and 0.15 sec. and the isometric relaxation period showed a definitely longer duration. The total duration of the ventricular systole (60 to 70 heart beats/min.) varied between 0.35 and 0.48 sec. It was further observed on several occasions that whenever the snakes became restless a higher frequency occurred. The propagation of the arterial pulse throughout the dorsal aorta of a snake is shown in figure 6. The velocity of propagation JOHANSBN 830 ECG Left aortic pressure Right atrium Ventricular pressure Left atrium mmHg Downloaded from http://circres.ahajournals.org/ by guest on June 14, 2017 50 25 0 Dorsal aorta R cm behind heart Ventricle 1 Isec Dorsal aorta 32 cm behind heart ' TIG. 4. Intra cardiac pressure relations in the snake heart. was around 1.3 M./sec. From other experiments the value has been found to vary between 1.2 and 1.5 M./sec. DISCUSSION The values reported regarding pressures in the heart and vessels of snakes are consistently higher than those reported for other reptiles. Steggerda and Essex3 found ventricular systolic pressures around 32 mm. tig in the mud turtle (Chelhydta serpentina). The systolic pressure in the left aorta was around 30 mm. Hg with a pulse pressure around 10 mm. Hg. It is conceivable that the snake heart has established a greater pumping capacity in accordance with the relatively longer arterial tree that necessarily results from the elongated body form. It may also be that the snakes studied, living 60° N have developed a more efficient circulation by adaptation to the colder climate. The pulse contour from the aortic pressure in snakes shows a diastolie gradient pointing toward a high peripheral resistance allowing a gradual efflux from the arterial reservoirs during diastole. Comparing the left aortic pressure drop with that of the ventricle it is completely shut off from the aorta. Thus the 1 second FIG. 5 Top. Diagram showing the period of isometric contraction (a-6) and isometric relaxation (c-d) in the snake heart. FIG. 6 Bottom. Propagation of the arterial pulse in the dorsal aorta of a snake. apparent that during this period the heart is aortic semilunar valves seem effectively to prevent regurgitation during diastole. There are important differences between aortic and pulmonary pressures. The pulse pressure in the pulmonary artery is twice as large as that recorded in the left aorta. This finding agrees with what Steggerda and Essex found in the turtle. The greater pulse pressure probably results from a lesser resistance in the lung vessels than in the systemic circuit. The mean pressure in the pulmonary vein may be considered low and the high pressure gradient between the pulmonary artery and vein leads to a high velocity of the blood in the pulmonary circuit. This situation resembles that seen in mammals, but the relatively higher pulmonary arterial pressure in snakes further accentuates this phenomenon. The low diastolie pressure in the pulmonary artery has obvious consequences. During ventricular systole the ejection of blood must necessarily first start through the pulmonary THREE-CHAMBERED SNAKE HEART Downloaded from http://circres.ahajournals.org/ by guest on June 14, 2017 trunk because of the least resistance there. This artery leaves the heart close to the entrance of the great systemic veins whereby the venous blood preferentially fills the pulmonary artery during ventricular contraction. Although the pressure rise in the pulmonary artery appears only around 0.5 sec. before the pressure rise in the aorta, this may be a factor preventing extensive shunting of venous blood to the systemic apertures. It seems likely that inside the ventricle a directional flow from left to right is favored, evidently similar to the situation encountered in the mammalian heart with congenital ventricular septal defects.3 Steggerda and Essex followed the course of blood through the heart and great vessels of the turtle by means of dye dilution technic. They reported that the dye injected into the right atrium appears very rapidly in the pulmonary artery and only a small fraction appears in the aortas. The recorded pressures revealed a difference between the systolic pressures in the left and right atrium. Steggerda and Essex reported from their experiments on turtles that the systolic pressure in the left atrium was more variable than that of the right atrium. The authors felt that this might be related to the capacity and pressure of the circulation through the lung. On the basis of the present study it may be suggested that the relatively great pressure gradient through the lung with its probably high flow result in a permanently increased pressure in the left atrium. The consequence, however, may be that already during ventricular filling, i.e., atrial contraction, there will be slight pressure gradients and left-to-right shunts inside the ventricle. The dynamics of heart beat in the snake thus seem to prevent a high degree of venous blood admixture to the arterial side of the ventricle. Tt may be mentioned that Steggerda and Essex found only slight changes in oxygen saturation between the right and left aorta and that these vessels carried blood with far greater oxygen content than did the pulmonary artery. This finding is contradictory to older reports 4-5 based on anatomic features 831 rather than exact physiologic measurements. In agreement with the findings of Steggerda and Essex, the present study revealed no difference in systolic pressures between the right and left side of the ventricle. A comparison of the pressures in the ventricle and left aorta reveal a distinct period of isometric contraction (fig. 5 a, b). The duration of isometric contraction is in good accord Math that reported by Shannon and Wiggers0 experimenting on the frog and turtle hearts. Likewise the period of isometric relaxation is definitely longer than isometric contraction, also agreeing with the report of Shannon and Wiggers. Another point of interest has to do with the correlation of electrical and mechanical changes throughout the heart cycle. The snakes exhibit an extraordinarily brief isopotential period between the T-wave and P-wave, in relation to the whole cardiac cycle. The T-P interval is only about one seventh the P-P duration. This peculiar situation is likewise reported for other species of reptiles. Davies et al.7 studied the electrocardiograms of the Nile crocodile and reported a T-P interval that is one fourth the entire cycle. Among several other investigators, Wiggers8 states that the end of ventricular systole cannot be definitely related to any phase of the T-wave. If, however, the completion of the T-wave indicates that repolarization has been completed, the period between repolarization and the next depolarization is extremely short and obviously different from that in the mammalian heart cycle. The absence of a definite period of diastasis in snakes seems to have a bearing on the extraordinarily long duration of ventricular activity in these animals. Another consequence must be that ventricular filling chiefly takes place during atrial contraction and thereby differs entirely from ventricular filling in mammals which mainly takes place prior to and during ventricular diastasis. SUMMARY The dynamics of heart beat have been studied in snakes. The pressures in the left aorta 832 J0J1AXSEN Downloaded from http://circres.ahajournals.org/ by guest on June 14, 2017 showed values of 50 to 60 mm. Hg systolic and 40 to 50 mm. Hg diastolic. The systolic pressure in the pulmonary artery was equal to that in the systemic arteries, while the diastolic pressure was distinct]}' lower. The pressure in the left atrium showed values more variable and higher than those fiom the right atrium. The circulatory dynamics of the snake heart seems to prevent a high degree of admixture of venous blood to the arterial side of the ventricle. The ventricular pressure record lacks a definite period of diastasis. The extraordinarily long duration of ventricular activity is also revealed from the electrocardiograms. The ventricular filling in snakes must chiefly take place during atrial contraction and thereby differ from the situation seen in mammals. SUMMARIO IN INTERUNGUA Le dynarnica del pulso cardiac esseva studiate in serpentes. Le tension in le aorta sinistre habeva valores de 50 a 60 mm Hg systolic e de 40 a 50 mm de Hg diastolic. Le tension systolic in le arteria pulmonar esseva equal al tension systolic in le arterias systemic durante quc le tension diastolic in le arteria pulmonar esseva distinctemente plus basse. Le tension in le atrio sinistre inonstrava valores plus variabile e plus alte que le tension in le atrio dextere. II pare que le dynamica cireulatori del corde de serpentes preveni un alte grado de adinixtion de sanguine venose al latere arterial del ventriculo. Le curvas del tension ventricular non exhibi un definite periodo de diastase. Le extraordinarimente longe duration del activitate ventricular es etiam revelate per le electrocardiogramma. On debe supponer que le plenation ventricular in serpentes occurre primarimentc durante le contraction atrial. Isto differe ab le situation in mammiferos. REFERENCES 1. MATHUR, P. X.: The anatomy of the reptilian heart. Proe. Indian Acad. Sc. 20: 1, 1944; 23: 129, 1946. 2. WEHN, P. S.: Pulsatory Activity of Peripheral Arteries. Oslo, Oslo University Press, 1957. :). STEGGERDA, F. R., AND ESSEX, H. E.: Circula- tion and blood pressure in the great vessels and heart of the turtle (Chelydra serpeniinu). Am. J. Physiol. 190: 310, 1957. ; l. GOODRICH, E. S.: Note on the reptilian heart. J. Anat. 53: 298, 1919. n . EWER, R. F.: Haemodynnmic factors in the evolution of the double circulation in the vertebrates. Am. Naturalist 84: 215, 1950. (i. SHANNON, E. W., AND WIGGERS, C. J.: The dynamics of the frog and turtle hearts. The non-refractory phase of systole. Am. J. Physiol. 128: 709, 1939. 7. DAVIES, F., FRANCIS, E. T. B., AND KING, T. S.: Electrocardiogram of the crocodilian heart. Nature 167: 146, 1951. .8. WIGGERS, C. J.: The electrocardiogram: Its relation to cardiodynamic events. Arch. Int. Med. 20: 93. 1917. Circulation in the Three-Chambered Snake Heart KJELL JOHANSEN, CAND. READ. Downloaded from http://circres.ahajournals.org/ by guest on June 14, 2017 Circ Res. 1959;7:828-832 doi: 10.1161/01.RES.7.6.828 Circulation Research is published by the American Heart Association, 7272 Greenville Avenue, Dallas, TX 75231 Copyright © 1959 American Heart Association, Inc. All rights reserved. Print ISSN: 0009-7330. Online ISSN: 1524-4571 The online version of this article, along with updated information and services, is located on the World Wide Web at: http://circres.ahajournals.org/content/7/6/828 Permissions: Requests for permissions to reproduce figures, tables, or portions of articles originally published in Circulation Research can be obtained via RightsLink, a service of the Copyright Clearance Center, not the Editorial Office. Once the online version of the published article for which permission is being requested is located, click Request Permissions in the middle column of the Web page under Services. Further information about this process is available in the Permissions and Rights Question and Answer document. Reprints: Information about reprints can be found online at: http://www.lww.com/reprints Subscriptions: Information about subscribing to Circulation Research is online at: http://circres.ahajournals.org//subscriptions/