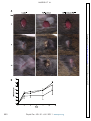
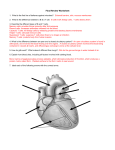
Survey
* Your assessment is very important for improving the work of artificial intelligence, which forms the content of this project
Physiol Rev 91: 931–972, 2011 doi:10.1152/physrev.00016.2010 FACTOR XIII: A COAGULATION FACTOR WITH MULTIPLE PLASMATIC AND CELLULAR FUNCTIONS László Muszbek, Zsuzsanna Bereczky, Zsuzsa Bagoly, István Komáromi, and Éva Katona Clinical Research Center and Thrombosis, Haemostasis and Vascular Biology Research Group of the Hungarian Academy of Sciences, University of Debrecen, Medical and Health Science Center, Debrecen, Hungary L I. II. III. IV. V. VI. VII. VIII. INTRODUCTION GENERAL OVERVIEW ON STRUCTURE,... ACTIVATION AND REGULATION OF... THE ACTION OF ACTIVATED FACTOR XIII... FACTOR XIII IN WOUND HEALING AND... OTHER EFFECTS OF FACTOR XIII FACTOR XIII AS AN INTRACELLULAR... SUMMARY, CONCLUSIONS, AND... 931 932 940 945 947 952 957 961 became apparent only after Duckert et al. (94) demonstrated that the severe bleeding diathesis of a patient was due to the deficiency of fibrin stabilizing factor. In 1963, soon after this clinical finding, the International Committee on Blood Clotting Factors acknowledged this protein as a clotting factor and termed it factor XIII. In 2007 the International Society of Thrombosis and Haemostasis, Scientific and Standardization Committee published a recommendation on the terms and abbreviations concerning FXIII (258), which will be followed throughout this review. I. INTRODUCTION The history of blood coagulation factor XIII (FXIII) started more than 70 years ago, when Barkan and Gaspar (33) observed that fibrin clots formed in the presence of Ca2⫹ become insoluble in weak bases. Some 20 years later, Robbins (314) performed the experiment with purified fibrinogen and concluded that in addition to Ca2⫹, a “serum factor” is also needed to render the clot insoluble in weak acids and bases. Laki and Lóránd (196) and Lóránd (221, 222) were the first who realized that the serum factor, which made the fibrin clot insoluble in concentrated urea solution, was thermo-labile and nondialyzable, and they called this protein fibrin stabilizing factor. Fibrin stabilizing factor was then purified from plasma, and the enzymatic nature of it was first revealed and characterized by Loewy and co-workers (215–219). The clinical significance of these findings For a relatively long time, FXIII did not raise much interest among scientists, and research on this field was practiced only by a few devotees, although their contributions were essential for later developments. It was revealed that FXIII is a zymogen, and in contrast to all other zymogen clotting factors, its active form (FXIIIa) is a transglutaminase (TG; protein-glutamine: amine ␥-glutamyltransferase, EC 2.3.2.13) that forms ⑀(␥-glutamyl)lysyl crosslinks between two polypeptide chains. The mechanism of the activation of zymogen FXIII and the basic enzymology of FXIIIa were clarified. FXIII was also found in platelets, and structural differences between plasma FXIII (pFXIII) and cellular factor (cFXIII) were revealed. Clinical data on the consequences of inherited FXIII deficiency provided important pieces of information on features of the disease and on the physiological role of FXIII in hemostasis. Neverthe- 931 Downloaded from http://physrev.physiology.org/ by 10.220.32.246 on June 14, 2017 Muszbek L, Bereczky Z, Bagoly Z, Komáromi I, Katona E. Factor XIII: A Coagulation Factor With Multiple Plasmatic and Cellular Functions. Physiol Rev 91: 931–972, 2011; doi:10.1152/physrev.00016.2010.—Factor XIII (FXIII) is unique among clotting factors for a number of reasons: 1) it is a protransglutaminase, which becomes activated in the last stage of coagulation; 2) it works on an insoluble substrate; 3) its potentially active subunit is also present in the cytoplasm of platelets, monocytes, monocyte-derived macrophages, dendritic cells, chondrocytes, osteoblasts, and osteocytes; and 4) in addition to its contribution to hemostasis, it has multiple extra- and intracellular functions. This review gives a general overview on the structure and activation of FXIII as well as on the biochemical function and downregulation of activated FXIII with emphasis on new developments in the last decade. New aspects of the traditional functions of FXIII, stabilization of fibrin clot, and protection of fibrin against fibrinolysis are summarized. The role of FXIII in maintaining pregnancy, its contribution to the wound healing process, and its proangiogenic function are reviewed in details. Special attention is given to new, less explored, but promising fields of FXIII research that include inhibition of vascular permeability, cardioprotection, and its role in cartilage and bone development. FXIII is also considered as an intracellular enzyme; a separate section is devoted to its intracellular activation, intracellular action, and involvement in platelet, monocyte/macrophage, and dendritic cell functions. MUSZBEK ET AL. less, FXIII used to be considered a stepchild among clotting factors for a number of reasons: 1) it falls outside the mainstream of the clotting cascade, and it only starts working after the visible end result, fibrin formation, had been completed; 2) as opposed to other clotting enzymes, it is making rather than breaking peptide bonds; and 3) it is working on an insoluble substrate matrix that is rather unattractive for enzymologists. This review, besides giving a short overview on earlier discoveries concerning structure and biochemical functions of FXIII, mainly deals with recent developments on the regulation of FXIII, with its role in fibrinolysis, and with its functions which partially or completely lie outside hemostatic mechanisms. The latter topic includes the cellular effects of FXIII and its involvement in angiogenesis and wound repair as well as in maintaining pregnancy. The intracellular activation and function of FXIII will also be discussed. II. GENERAL OVERVIEW ON STRUCTURE, GENE, POLYMORPHISMS, AND EXPRESSION OF FACTOR XIII The structure and synthesis of FXIII have been reviewed earlier in detail (124, 263); here only a general overview with emphasis on new discoveries will be provided. FXIII is a pro-TG that circulates in plasma in tetrameric form (FXIII-A2B2) (124, 259, 263). It consists of two potentially active, catalytic A subunits (FXIII-A) and two carrier/inhibitory B subunits (FXIII-B). FXIII-B is in excess; in the 932 A. Factor XIII A Subunit and Cellular Factor XIII FXIII-A, a protransglutaminase, belongs to the family of TGs. Eight members of the family (FXIII-A and TG1-TG7) possess potential enzymatic activity; while an additional protein, erythrocyte band 4.2, has a similar domain structure but, due to the replacement of active site amino acid, lacks enzymatic activity. (A more detailed review on TGs is provided in Reference 95.) All TGs are monomers; only FXIII-A forms a dimeric structure. The three-dimensional structure of three human TGs, FXIII-A2, TG-2, and TG-3, has been resolved (14, 213, 377, 383). It is interesting that despite the modest sequential agreement, their secondary structure shows striking similarity. A detailed comparison of the structure of the three TGs is given in Reference 187. FXIII-A consists of 732 amino acids including an initiator methionine (http://www.uniprot.org/uniprot/P00488); its molecular mass is 83 kDa. Serine is in the penultimate position, which favors the removal of initiator methionine and then N-acetylation. The NH2-terminal amino acid in the mature molecule is, indeed, an acetylated serine residue. In the following, amino acid numbering starting with the Ser residue will be used. No hydrophobic leader sequence could be identified in the primary structure of FXIII-A (144). There are nine cysteine residues in the protein, including the active site cysteine (Cys314), none of which forms disulfide bonds. Although the molecule has potential sites for N-glycosylation, no carbohydrate residue is detected. FXIII-A is expressed in all vertebrates investigated so far. A phylogenetic tree and sequence identity in different species are shown on FIGURE 1. Recombinant FXIII-A2 (rFXIII) has been expressed in Escherichia coli (45), in Saccharomyces cerevisiae (42, 157), in Schizosacharomyces pombe (52), and even in tobacco plant cells and whole tobacco plants (108). Features of rFXIII are identical to those of cFXIII. The first atomic resolution structure of FXIII-A2 was published (383) one and a half decades ago. Two crystal forms, orthorhombic (383) and monoclinic (377), were obtained for FXIII-A2, and X-ray crystallography of both crystal forms resulted in essentially the same structure. Studies on the X-ray structure of rFXIII-A2 (102, 377, 382, 383) revealed numerous structural details of FXIII-A2 and provided invaluable information on the structure-function relationships. The X-ray structure was also a great help in understanding the structural consequences of mutations causing FXIII deficiency (173). Physiol Rev • VOL 91 • JULY 2011 • www.prv.org Downloaded from http://physrev.physiology.org/ by 10.220.32.246 on June 14, 2017 Discoveries starting from the 1980s changed this trend, and since then FXIII has become an increasingly attractive field of research. The milestones of FXIII research were the following: 1) the realization that FXIII plays a highly important role in the regulation of fibrinolysis. 2) The discovery of cFXIII in monocytes and different types of macrophages, including tissue macrophages, revealed that this proenzyme is present in most organs and tissues of the body. 3) The primary protein structure and the genomic structure of FXIII subunits were published in the second half of the 1980s followed by the exact identification of molecular genetic defects in an increasing number of FXIII deficient patients. In 1994 the three-dimensional structure of cellular FXIII has also been published. 4) The availability of recombinant cFXIII (rFXIII) significantly promoted basic biochemical studies. 5) New, relatively simple reliable methods for the determination of FXIII activity and concentration in the plasma became available and made large-scale clinical studies possible. 6) The discovery of FXIII polymorphisms induced biochemical and clinical studies to explore their consequences. 7) It was realized that FXIII is a multifunctional protein that, beside hemostasis, plays an important role in a wide variety of physiological and pathological processes. plasma, ⬃50% of it exists in free noncomplexed form. FXIII-A dimer (FXIII-A2), but not FXIII-B, is also present in the cytoplasm of certain cells, particularly of platelets and monocytes/macrophages. The primary structure of both subunits has been determined by cDNA cloning and amino acid sequence analysis (127, 145, 146, 359). MULTIPLE FUNCTIONS OF FACTOR XIII FXIII-A consists of four main structural domains, -sandwich (amino acid 38 –184), catalytic core (185–515), -barrel 1 (516 – 628), and -barrel 2 (629 –731) domains, plus an NH2-terminal activation peptide (AP-FXIII) (FIGURE 2). The latter is cut off by thrombin during activation (see sect. IIIA for details). The sandwich and barrel domains almost exclusively consist of -sheets with only a few small helical structural elements in the sandwich and -barrel 1 domains. The core domain contains both -sheets and helices. On the basis of thermal denaturation studies (92, 193), the core domain can be further divided into NH2- (189 –332) and COOH-terminal (333–515) subdomains. Two nonproline cis peptide bonds were observed in the core domain (377) that also exists in human TG-2 (213) and TG-3 (14). The FXIII-A subunit monomers are arranged in a dimeric structure resembling a hexagon with C2 symmetry, in which the two central core domains are surrounded by the six -sheet domains (FIGURE 2). The X-ray studies also revealed that the arrangement of the amino acid residues playing a key role in the catalytic process (Cys314, His373, Asp396) resembles the catalytic triad in cysteine proteases (FIGURE 3A). Therefore, it was assumed that the catalytic reaction corresponds to a reverse proteolysis (294). However, due to the active site specificity of the enzyme, it catalyzes interchain peptide bond formation between the glutamine and lysine side chains instead of peptide chain elongation. In the reaction catalyzed by cys- teine proteases, the stabilization of the intermediates by the so-called oxyanion hole is of crucial importance. Based on the structural analogy with papain, it has been assumed that the NH group of the indole side chain of Trp279 and the NH group of Cys314 backbone form the oxyanion hole (294) (FIGURE 3A). Trp279 is highly conserved in vertebrate TGs, and its direct involvement in the TG reaction has been shown by kinetic study with TG-2 (147). Although the geometrical arrangement of the catalytic triad and the oxyanion hole in the catalytic domain corresponds to the arrangements in cysteine proteases (288), X-ray structure also revealed that the catalytic cysteine residue is completely buried by AP-FXIII (FIGURE 3A). A strong saltbridge between the side chain of Arg11= in AP-FXIII extending over from the opposite subunit and the side chain of Asp343 of the core domain enforces AP-FXIII into a position that makes Cys314 inaccessible for the substrate and even for the solvent molecules. Therefore, the removal or disposition of AP-FXIII is a prerequisite of activation. However, this is not sufficient since the accessibility of the active site to the substrate is also prevented by the strong hydrogen bond that exists between the O atom of Tyr560 and S␥ atom of Cys314 residues (FIGURE 3A). Tyr560 is located on a loop of the -barrel 1 domain, which means that the transposition of this loop from the immediate vicinity of the active site is also required for the interaction with the substrate. Considering the extreme stability of barrel domains (193) and the size of the natural Physiol Rev • VOL 91 • JULY 2011 • www.prv.org 933 Downloaded from http://physrev.physiology.org/ by 10.220.32.246 on June 14, 2017 FIGURE 1 Phylogenetic tree and sequence identity of A subunit of factor XIII (FXIII-A) in different species. A: phylogenetic tree of FXIII-A was constructed by the BIONJ version (109) of neighbor-joining algorithm using the SplitsTree4 software (141) downloaded from the www.splitstree.org website. FXIII-A protein sequences were aligned by ClustalX software (198). Distance scale represents the number of differences between sequences in species since their divergence; 0.1 means 10% difference between two sequences. B: identity percent represents the degree of homology in different organisms compared with human FXIII-A protein and coding DNA. Data were obtained from the HomoloGene database (www.ncbi.nlm.nih.gov/sites/homologene). The amino acid sequence of salmon FXIII-A was downloaded from the Universal Protein Resource (UniProt) web server (http://www.uniprot.org), and its identity percent value was calculated by BLAST software (www.blast.ncbi.nlm.nih.gov) using the alignment obtained by ClustalX software (198). MUSZBEK ET AL. FIGURE 2 Ribbon and tube representation of the zymogen rFXIII-A2 homodimer based on its X-ray structure (377) and visualized by the Chimera software (299). The relative positions of the constituting domains are marked for only one of its monomeric FXIII-A subunit. Helical and -sheet structural elements are colored orange and purple, respectively. The disordered (random coil like) structural elements and loops are shown in cyan, and the activation peptides are depicted in bright green. substrates, the removal of the whole -barrel 1 from its original position is an even more likely option. Ca2⫹ binding sites on the surface of FXIII-A2 are essential for the activation of FXIII. FXIII-A, but not FXIII-B, tightly binds The gene coding for human FXIII-A (F13A1) spans over 160 kb and has been localized to chromosome 6p24 –25 (46). It is transcribed into a 3.9-kb mRNA, with an 84-bp 5=-untranslated region, a 2.2-kb open reading frame, and a 1.6-kb 3=untranslated region. F13A1 contains 15 exons and 14 introns (144). Exon I consists of the 5= noncoding region, and exon II encodes AP-FXIII (FIGURE 4). The -sandwich domain, the catalytic core domain, and the two -barrel domains are encoded by exons II-IV, exons IV-XII, exons XII-XIII, and exons XIII-XV, respectively. shows the polymorphisms and their location in the coding region of FXIII-A gene. Data provided by the International HapMap project (www.hapmap.org) on the frequency of the polymorphisms in the Caucasian, African, FIGURE 4 FIGURE 3 Structures around the catalytic triad (A) and the main Ca2⫹ binding site (B) of FXIII-A. Structural elements are shown by “ribbon and tube” representation; helical and -sheet structural elements are colored orange and purple, while disordered (random coil like) structural elements and loops are shown in light green. Side chains of selected amino acid residues are represented by balls and sticks. The N, C, O, and S atoms are colored blue, gray, red, and yellow, respectively. A: the amino acids of the catalytic triade (Cys314-His373Asp396) and Trp279 participating in the formation of oxyanion hole are shown together with interacting amino acid residues that fix the structure in inactive conformation and bury the active site cysteine. The X-ray structure published by Weiss et al. (377) was used for the preparation of the picture (377). Arg11= represents the Arg residue in AP-FXIII of the opposite subunit. B: amino acid residues implicated in the binding of Ca2⫹ (green ball) at the main Ca2⫹ binding site. The structure based on the X-ray coordinates published by Fox et al. (102) was visualized by the Chimera software (299). 934 Physiol Rev • VOL 91 • JULY 2011 • www.prv.org Downloaded from http://physrev.physiology.org/ by 10.220.32.246 on June 14, 2017 one Ca atom with a Kd of 0.1 mM as determined by equilibrium dialysis and fluorescence studies (209), while a Kd of 0.51 mM was determined by 43Ca NMR experiments (19). These reports also suggest the existence of additional low-affinity Ca2⫹ binding sites. From X-ray structural studies of FXIII-A2 cocrystallized with Ca2⫹, Str2⫹, and Yb3⫹, a single main cation binding site per subunit was identified (102). The carboxylate group of Asp438, Glu485, and Glu490 side chains, the carbonyl O atom of Asn436, as well as the backbone carbonyl O atom of the Ala457 residue participate in cation binding (either directly or through a water bridge) to a different extent depending on the nature of the cation (FIGURE 3B). The backbone carbonyl O atom of Ala457 and the carboxylate groups of Glu485 and Glu490 side chains have been reported to be directly involved in Ca2⫹ binding (102, 195). MULTIPLE FUNCTIONS OF FACTOR XIII Downloaded from http://physrev.physiology.org/ by 10.220.32.246 on June 14, 2017 FIGURE 4 Distribution of SNPs published so far within the coding region of F13A1 gene. The “A” of the translation initiation codon “ATG” is considered as nucleotide one, and nucleotide positions within cDNA are given accordingly. The amino acid numbering is given according to Ichinose et al. (144). Exons encoding a particular domain are color-coded. The mutations resulting in amino acid replacements are depicted in purple, while silent mutations are shown in black. Physiol Rev • VOL 91 • JULY 2011 • www.prv.org 935 MUSZBEK ET AL. tive effect against venous thromboembolism (378) and myocardial infarction (370). The complex interrelationship of FXIII polymorphisms with the risk of thrombotic disease is beyond the scope of this review; interested readers should consult a most recent review (259). The biochemistry of other common FXIII-A polymorphisms has not been investigated in such details as that of Val34Leu polymorphism. The Phe204 allele of FXIII-A Tyr204Phe polymorphism was reported to be associated with decreased pFXIII level and activity, whereas the Leu564 allele of the Pro564Leu variant resulted in lower FXIII plasma level with increased FXIII activity (22, 107). However, these results still need to be confirmed. FXIII-A is expressed primarily in cells of bone marrow origin. It is present in platelets in huge quantity (55, 228). The average FXIII-A content of a single platelet was estimated to be 60 ⫾ 10 fg, which corresponds to 3% of total platelet proteins (176). The concentration of FXIII-A is ⬃100- to 150-fold higher in platelet cytoplasm than in plasma. FXIII-A is also present in megakaryocytes (10, 179, 231, 256) and in their precursor cells (10). Megakaryocytes synthesize FXIII-A and package it, together with its encoding Table 1 Racial variations of the polymorphisms in factor XIII-A and factor XIII-B subunit genes Nucleotide Variation F13A1 c.103G⬎T c.614A⬎T c.996A⬎C c.1652C⬎T c.1694C⬎T c.1704A⬎G c.1696T⬎A c.1951G⬎A c.1954G⬎C F13B c.344G⬎A c.456G⬎A c.765C⬎T c.1049A⬎G c.1707T⬎G c.1806T⬎C c.1952 ⫹ 144 C⬎G Allele Frequency Amino Acid Variation CEU YRI JPT HCB p.V34L p.Y204F p.P331P p.T550I p.P564L p.E567E p.L588Q p.V650I p.E651Q 0.767/0.233 0.983/0.017 0.853/0.147 1 0.758/0.242 0.890/0.110* 0.975/0.025 0.950/0.050 0.775/0.225 0.883/0.117 1 0.933/0.067 0.992/0.008 0.864/0.136 0.991/0.009 1 0.942/0.058 0.742/0.258 1 1 1 1 0.705/0.295 1 1 0.909/0.091 0.909/0.091 1 1 0.989/0.011 1 0.733/0.267 1 1 0.911/0.089 0.911/0.089 p.R95H p.T132T p.C235C p.H330R p.D549E p.N582N - 0.075/0.925 0.925/0.075 1 1 1 0.492/0.508 0.85/0.142 0.725/0.275 0.307/0.693 0.983/0.017 0.933/0.067 0.908/0.092 0.842/0.158 0.98/0.017 0.034/0.966 1 1 0.989/0.011 0.966/0.034 0.244/0.756 0.42/0.578 0.044/0.956 1 1 0.989/0.011 0.956/0.044 0.189/0.811 0.35/0.644 In the case of F13A1 and F13B polymorphisms, amino acid positions are numbered according to Ichinose et al. (144) and according to Bottenus et al. (50), respectively. Allele frequency data of the International HapMap project (www.hapmap.org) were used; the first numbers represent the frequency of ancestral allele. CEU, Utah residents with ancestry from Northern and Western Europe; YRI, African population in Yoruba in Ibadan, Nigera; JPT, Japanese in Tokyo, Japan; HCB, Han Chinese in Beijing, China. *As in the case of c.1704A⬎G mutation, no HapMap data for CEU were available; data were taken from the Centre d’Etude du Polymorphisme Human (CEPH) project (www.cephb.fr). 936 Physiol Rev • VOL 91 • JULY 2011 • www.prv.org Downloaded from http://physrev.physiology.org/ by 10.220.32.246 on June 14, 2017 and Asian populations demonstrate considerable racial variation (TABLE 1). Detailed studies on the biochemical and clinical consequences of the polymorphisms were carried out only in the case of Val34Leu polymorphism, a primarily Caucasian mutation first described by Mikkola et al. (244). This mutation increases the rate of FXIII activation (23, 31, 374) (see sect. IIIA for details). The structure of fibrin clots is also influenced by FXIII-A Val34Leu polymorphism (23), and this effect is modulated by fibrinogen concentration (211). At high fibrinogen levels, plasma samples from homozygotes for the Leu34 allele form clots having looser structure, thicker fibers, and increased permeability, while at low fibrinogen concentrations fibrin meshwork had thinner, more tightly packed fibers, and lower permeability. Practically no fibrinogen concentration-dependent changes were observed in the plasma samples of wild-type individuals. The effect of the Val34Leu polymorphism on the specific activity of FXIIIa was controversial for a while; it is now clear that the specific activity of fully activated pFXIII (23, 31), cFXIII (374), and rFXIII-A2 (201) of different FXIII-A Val34Leu genotypes are identical. Although there are contradictory results on the clinical implications of this polymorphism, meta-analyses demonstrated its protec- MULTIPLE FUNCTIONS OF FACTOR XIII mRNA, into newly formed platelets (5, 6, 231). It is now clear that FXIII-A in platelets is essentially of cytoplasmic localization (51, 220, 278, 346). Platelets could take up a tiny amount of FXIII-A2B2 from the plasma which appears in ␣-granules (233); this amount is negligible compared with the amount of FXIII-A in the cytoplasm. The presence of FXIII-A mRNA and the synthesis of FXIII-A have also been verified in the above cell types (4, 5, 190, 271). The transcriptional regulation of cell type-specific expression of FXIII-A was investigated in monocytoid (U937) and megakaryocytoid (MEG-01) cell lines containing FXIII-A mRNA (178). Reporter gene assays revealed that a 5=-fragment was sufficient to support the basal expression in the above cells, but not in other cell types. Promoter elements for a myeloid-enriched transcription factor (MZF-1) and for the ubiquitous transcription factors, SP-1 and NF-1, seem to be important for the basal FXIII-A expression. Another region had enhancer activity in MEG-01 cells but silencer activity in B. Factor XIII B Subunit FXIII-B is a glycoprotein consisting of 641 amino acids and containing 8.5% carbohydrate; its molecular mass is ⬃80 kDa. It is a typical mosaic protein consisting of 10 short tandem repeats, called sushi domains or GP-I structures, each containing ⬃60 amino acids and held together by a pair of internal disulfide bonds (FIGURE 5). The same structures have also been found in more than 20 other proteins, including 2-glycoprotein I, many of which are encoded by genes clustered in chromosome 1 band q32 (143). By electron microscopy, the B subunit appears as a thin, flexible, and kinked strand (58). FXIII-B has not been crystallized, and no reliable information on its three-dimensional structure is available. While FXIII-A in the absence of FXIII-B forms homodimer, contradictory publications concerning the state of noncomplexed FXIII-B were obtained. Gel filtration of purified FXIII-B suggested the presence of FXIII-B dimers (335), while sedimentation analysis revealed monomeric form (58). In a most recent report, gel filtration of recombinant FXIII-B supported the former finding (350). There is no good explanation for the discrepancy, and in plasma conditions, the state of free FXIII-B remains to be explored. The gene of FXIII-B subunit (F13B) is located at position 1q31–32.1; it is ⬃28 kb in length and composed of 12 exons producing a 2.2 kb mRNA (146). Exons are interrupted by 11 introns. Exon I encodes a 20-amino acid leader sequence characteristic for proteins that are secreted via the classical secretory pathway (50, 169, 375). Exon II-XI code for the sushi domains, each of which is encoded by a single exon. The sushi domains show a high degree of homology, suggesting gene duplication and exon shuffling during evolution (292, 293). The last exon codes for a COOH-terminal region of FXIII-B, for the 3=-untranslated region, and for the polyA tail. F13B is directly regulated by transcription factor HNF1␣ and HNF4␣ (143, 214). Accordingly, FXIII-B is expressed in the liver and secreted by hepatocytes (146, 149, 266). The polymorphic nature of FXIII-B was revealed a long time ago (43, 81, 207). On the basis of isoelectric focusing experiments, three major population-associated phenotypes were described: FXIII-B*1, FXIII-B*2, and FXIII-B*3, characteristic of European, African, and Asian populations, respectively. Since the first report of a F13B gene polymorphism by Board (43), several new polymorphisms have been described. FXIII-B polymorphisms and their frequency in different races are summarized on FIGURE 6 and in TABLE 1. By screening all the exons of F13B, Komanasin et al. (186) found three poly- Physiol Rev • VOL 91 • JULY 2011 • www.prv.org 937 Downloaded from http://physrev.physiology.org/ by 10.220.32.246 on June 14, 2017 FXIII-A was also detected in monocytes (257) and in their bone marrow precursor cells (10). Like platelets, monocytes also lack FXIII-B. A most recent study revealed that FXIII-A may be differently expressed in monocyte subsets; higher FXIII-A mRNA expression was demonstrated in Ly-6Chigh monocytes than in their Ly-6Clow counterparts (358). FXIII-A in monocytes is also of cytoplasmic localization, although during differentiation into macrophages it also appears in the nucleus (7). The results of the first discovery were soon confirmed, and FXIII-A was also detected in a number of monocyte-derived macrophages including macrophages of different serous cavities, alveolar macrophages, tumor-associated macrophages, histiocytic and dendritic reticulum cells of lymph nodes, connective tissue histiocytes, perivascular dendritic macrophages, dermal dendrocytes etc. (reviewed in Refs. 6, 263). FXIII-A has also been demonstrated in chondrocytes, osteoblasts, and osteocytes (279) (see sect. VID). In early studies, FXIII-A was also identified in placenta and in uterus (48, 61). With the use of monocyte and macrophage differentiation markers, it has been clearly demonstrated that FXIII-A is confined to monocyte-derived tissue macrophages in the placenta (9, 171) and in the uterus (11) as well. The expression of FXIII-A in monocytes is retained after malignant transformation; in fact, its expression in the respective cells is upregulated in acute myelomonocytic, monocytic leukemias and in chronic myelomonocytic leukemias (172). It turned out that the detection of FXIII-A by flow cytometry in leukemic cells is a useful intracellular marker in the classification of acute myeloid leukemias. It is interesting that FXIII-A becomes expressed in part of the lymphoblasts in acute B cell lymphoid leukemia, while it is absent in lymphocytes, normal lymphoid precursors, and mature lymphocytes from B cell chronic lymphoid leukemia (183). U937 cells, and the GATA-1 element was found to be responsible for the enhancer activity. MUSZBEK ET AL. morphisms in healthy Caucasian subjects. An A to G transversion within exon III (rs6003) leads to His to Arg amino acid exchange at codon 95 in the second sushi domain of the mature protein. The frequency of FXIII-B Arg95 carriers among healthy Caucasians is ⬃15%, it is more frequent among Africans (also named as FXIII-B*2 at the phenotypic level), and it is missing from the Asian population (155, 186). The polymorphism did not influence FXIII-A, FXIII-B, or pFXIII antigen levels. Increased subunit dissociation was found in plasma from subjects possessing the Arg allele. However, when the variants were purified to homogeneity and binding was analyzed by steady-state kinetics, no difference was observed (186). Most recently, a C-to-G change at position 29756 in intron K (IVS11⫹144) leading to a novel splice acceptor site was described (155, 321). This polymorphism results in allele-specific splicing products, and a protein 15 amino acids longer at the COOH terminus than its wildtype counterpart is synthesized. The variant sequence includes two additional lysine and one glutamic acid res- 938 idues. These charged amino acids change the pI of the protein. The polymorphism characteristically occurs in Asians; it corresponds to FXIII-B*3 at the phenotype level. Its frequency is less in Caucasian populations, and it does not seem to be present among Africans. Although such a profound structural change would be expected to alter some of the biochemical features of the molecule, this possibility has not been explored, yet. C. Factor XIII Complex in Plasma The origin of FXIII-A in pFXIII complex has been a debate for a long period of time. Studies characterizing phenotypes of plasma FXIII subunit A after bone marrow and liver transplantation clearly indicated that the major source of plasma FXIII-A is a cell population or cell populations of bone marrow origin (303, 379). In one of the studies, a minor source of recipient FXIII-A, independent of donor hematopoesis, was also detected (303); in theory, such source(s) could be long-persisting tissue macrophages or Physiol Rev • VOL 91 • JULY 2011 • www.prv.org Downloaded from http://physrev.physiology.org/ by 10.220.32.246 on June 14, 2017 FIGURE 5 Schematic structure of FXIII-B consisting of 10 sushi domains each held together by two pairs of disulfide bonds. Amino acids in the leader sequence are depicted in purple, and those in the mature protein are shown in cyan. The solid stars point to the site of N-glycosylation, and the empty star points to a potential glycosylation site to which no carbohydrate is attached. MULTIPLE FUNCTIONS OF FACTOR XIII liver cells. However, FXIII-A and its encoding mRNA could be detected in hepatocytes only in trace amounts using highly sensitive techniques (5, 6), and there is no evidence on the contribution of tissue macrophages to FXIII-A in the plasma. Bone marrow ablation in patients undergoing autologous peripheral blood stem cell transplantation decreased platelet count to a very low level (⬎90% decrease), while the reduction of plasma FXIII level was only ⬃25% (152). It has also been shown that in long-standing severe thrombocytopenia, the reduction of plasma FXIII was less than expected (179). These findings suggest that in the case of highly impaired production of FXIII-A by bone marrow cells, synthesis of FXIII-A could be taken over by other cell types. It is to be noted that in early embryonic life, well Physiol Rev • VOL 91 • JULY 2011 • www.prv.org 939 Downloaded from http://physrev.physiology.org/ by 10.220.32.246 on June 14, 2017 FIGURE 6 Distribution of SNPs published so far within the coding region of F13B gene. The “A” of the translation initiation codon “ATG” is considered as nucleotide one, and nucleotide positions within cDNA are given accordingly. The amino acid numbering is given according to Bottenus et al. (50). The mutations resulting in amino acid replacements are depicted in purple, and silent mutations are in black. The intronic mutation resulting in an alternative splice site is shown in green. MUSZBEK ET AL. Only the liver has been demonstrated as the source of FXIII-B in plasma (see above). This conclusion was also supported by phenotyping plasma FXIII-B following liver transplantation. In this case, the phenotype of recipient’s FXIII-B changed to the donor’s phenotype, while the phenotype of FXIII-A remained unchanged (379). FXIII subunits derived from separate cellular sources form tetrameric complex in the circulating blood. A reference interval of 14 –28 mg/l has been established for FXIII-A2B2 in human plasma (175). It is surprising that there is still uncertainty concerning the affinity of the two types of FXIII subunits. Apparent binding constants of 4 ⫻ 10⫺7 M (305) and 8.06 ⫻ 10⫺8 M (186) were reported. However, when calculating with these binding constants, a significant part of FXIII-A should be in free form, which is evidently not the case. In our laboratory one magnitude lower binding constant (2.73 ⫻ 10⫺9 M) was measured; this value fits better with the fact that FXIII-A is present in the plasma in practically fully complexed form. In normal conditions, FXIII-B is in excess in the plasma, and ⬃50% of it circulates in free, noncomplexed form (384). In patients with FXIII-A deficiency, the total amount of FXIII-B in the plasma is reduced while the concentration of free B subunits remains constant. The FXIII-A domain(s) that are responsible for the association with FXIII-B have not been identified. Most recently, binding analysis with various truncated forms of rFXIII-B suggested that the first sushi domain was responsible for the binding of FXIII-B to FXIII-A (350). In the plasma, practically all FXIII molecules are bound to fibrinogen (Kd ⬃10⫺8 M) and the association is independent of the presence of calcium ions (123, 125, 215). The ␥=-chain in ␥A␥= fibrinogen (fibrinogen 2) provides the major binding site for FXIII (91, 250). The ␥=-chain is present in ⬃15% of plasma fibrinogen. Compared with the ␥A chain, the variant chain contains a 20-amino acid exten- 940 sion, which is the binding site for FXIII through its B subunit and also for thrombin exosite 2. III. ACTIVATION AND REGULATION OF FACTOR XIII: THE BIOCHEMICAL ACTION OF ITS ACTIVE FORM A. Activation of Factor XIII in the Plasma and Within Cells The activation of pFXIII occurs in the final phase of the clotting cascade by the concerted action of thrombin and Ca2⫹ (FIGURE 7). In the initial step of the reaction, thrombin cleaves off AP-FXIII from the NH2 terminus of FXIII-A by hydrolyzing the Arg37-Gly38 peptide bond. Then, in the presence of Ca2⫹, the inhibitory B subunits dissociate, which is a prerequisite for the truncated FXIII-A dimer (FXIII-A2=) to assume an enzymatically active conformation (FXIII-A2*). The conformational change of FXIII-A2= resulting in an active TG also requires Ca2⫹. The proteolytic cleavage of FXIII-A by thrombin considerably weakens the interaction between A and B subunits (224, 305). The binding of Ca ions to the high-affinity Ca2⫹ binding sites on the A subunits is sufficient to dissociate the subunits and to activate the released A= dimer (138, 210). Interestingly, in the presence of Ca2⫹, complete activation of the FXIII-A dimer occurs if only one AP-FXIII is released from one of the two A subunits: in other words, the dimer of a cleaved and an uncleaved A subunit (FXIII-A*A°) possesses full enzymatic activity (138). It is to be noted that Siebenlist et al. (342) reported a low innate activity of the zymogen FXIII-A2B2 that can slowly cross-link fibrin(ogen) in the presence of Ca2⫹, but it is ineffective on low-molecular-weight substrates (342). It is not clear if such an activity is due to the presence of a small fraction of free FXIII-A2 that remains uncomplexed or dissociates from the complex and undergoes nonproteolytic activation (see below) or it is the intrinsic property of the fibrin(ogen)-bound tetrameric zymogen. Thrombin is not the only serine protease that could cleave and activate FXIII in the presence of Ca2⫹. Several other proteases, including batroxobin marajoensis (371), thrombocytin (273), trypsin (188, 334), and activated factor X (FXa) (236) have been reported to be able to activate FXIII. Although the cleavage site of these enzymes on FXIII-A has not been identified, based on their substrate specificity and on the Mr of the truncated FXIII-A, it is assumed that they cleave the same Arg37-Gly38 peptide bond as thrombin, and the active form of FXIII-A produced by these proteases, like the thrombin activated form, also has an NH2-terminal glycine. In contrast to thrombin, the proteolytic cleavage of FXIII-A by FXa requires Ca2⫹. No evidence has been provided on the physiological implication of FXa-induced FXIII activation. The mannan-binding lectin associated serine protease 1 (MASP1) that is involved in the complement Physiol Rev • VOL 91 • JULY 2011 • www.prv.org Downloaded from http://physrev.physiology.org/ by 10.220.32.246 on June 14, 2017 before the development of bone marrow, FXIII-A is present in mesenchymal histiocytes, and by week 20, FXIII-A also appears in liver cells (170). In summary, in normal conditions, FXIII-A present in pFXIII is predominantly synthesized by cells of bone marrow origin; however, in the case of impaired bone marrow function, other not yet identified cell types might take over the production of FXIII-A. Another unresolved question is how FXIII-A is released from cells that synthesize it. In the absence of leader sequence, the classical secretory pathway does not operate. In a most recent paper, cFXIII in macrophages was found in association with podosomes and other structures adjacent to the plasma membrane, and FXIII-A was present in intracellular vesicles positive for Golgi matrix protein 130, which has been implicated in the delivery of nonclassically secreted proteins to the plasma membrane (75). Although this finding raises the possibility of a nonclassical secretory pathway, clear-cut evidence supporting this hypothesis is still to be provided. MULTIPLE FUNCTIONS OF FACTOR XIII Downloaded from http://physrev.physiology.org/ by 10.220.32.246 on June 14, 2017 FIGURE 7 Different mechanisms of FXIII activation. FXIII-A=, A subunit of FXIII from which the activation peptide was cleaved off; FXIII-A*, FXIII-A= transformed into an active transglutaminase in the presence of Ca2⫹; FXIII-A°, noncleaved FXIII-A transformed into an active transglutaminase by the nonproteolytic mechanism in the presence of Ca2⫹. Green and orange cylinders represent -barrel and -sandwich domains of FXIII-A, respectively. The central core domains in FXIII-A are depicted as horseshoes in magenta. The activation peptides are shown as red loops. The elongated bended structure consisting of 10 pearls surrounding FXIII-A2 corresponds to FXIII-B. Physiol Rev • VOL 91 • JULY 2011 • www.prv.org 941 MUSZBEK ET AL. system and possesses thrombin-like activity also cleaves FXIII-A at Arg37-Gly38, although at a much slower rate than thrombin (191). It is not likely that FXIII activation by MASP1 has any physiopathological importance. Recently, it has been shown that limited cleavage of FXIII by human neutrophil elastase resulted in the activation of both pFXIII and cFXIII with TG activities of ⬃50% of thrombin-activated FXIII (28). MALDI-TOF analysis of the cleaved fragments and NH2-terminal Edman degradation of the truncated protein identified Val39-Asn40 as the primary cleavage site, and the NH2-terminal amino acid of this novel active form of FXIII is Asn. The presence of polymerizing fibrin enhances the activation of pFXIII by ⬃100-fold (122, 123, 159, 210, 272). The binding to fibrin makes the orientation of both pFXIII and thrombin favorable for the proteolysis of FXIII-A at Arg37Gly38. Both fibrin I, which is devoid of fibrinopeptide A, and fibrin II, which is devoid of fibrinopeptides A and B, are effective accelerators. This effect is partly attributed to the FXIII-B-driven optimal orientation of the tetramer molecule. It facilitates the interaction with thrombin and accelerates the thrombin-induced cleavage of FXIII-A in the presence of fibrin (122, 137). In the absence of FXIII-B, i.e., in the case of cFXIII, no fibrin-induced promotion of the removal of AP-FXIII could be observed (121, 137). Experiments with thrombin mutants demonstrated that His66, Tyr71, and Asn74, surface residues of thrombin, are involved in high-specificity fibrin-enhanced factor XIII activation. These residues, representing a distinct interaction site on thrombin (within exosite 1), are also employed by thrombomodulin in the cofactor-enhanced activation of protein C (300). Enhancement of factor XIII activation by fibrin was competitively inhibited by thrombomodulin. When the degree of fibrin ␥-chain cross-linking exceeds 942 Results concerning the effect of FXIII binding to fibrinogen ␥=-chain on FXIII activation are contradictory. Moaddel et al. (247) reported that ␥A␥A fibrin accelerated the activation of pFXIII less efficiently than ␥A␥= fibrin. In contrast, Siebenlist et al. (342) demonstrated more rapid FXIII activation in the presence of ␥A␥A fibrin(ogen) than with ␥A␥= fibrin(ogen). Atroxin, a thrombin-like snake venom enzyme that does not bind to the ␥= sequence, activated FXIII in the presence of ␥A␥A and ␥A␥= fibrin(ogen) to the same extent. This finding indicates that rate differences observed with thrombin are related to the binding of thrombin rather than to the binding of FXIII to the ␥= sequence. In the plasma FXIII becomes effectively activated only on the surface of newly formed fibrin, and the truncated active dimer remains associated with its substrate; no thrombincleaved FXIII could be detected in the serum (341), while following dissociation, FXIII-B becomes detached from fibrin. Fluorescence studies showed that cFXIII also binds to fibrin, but with less affinity than pFXIII. The affinity, however, is increased upon cFXIII activation (137). Fibrin(ogen) also regulates the Ca2⫹ requirement for the activation of truncated FXIII-A2= (78, 79). It brings down the optimal Ca2⫹ concentration from ⬎10 mM to the physiological interval present in the plasma. Considering that the location of FXIII-A Val34Leu polymorphism is just 3 amino acids upstream from the thrombin cleavage site, one would expect its influence on thrombininduced FXIII activation. Indeed, it was demonstrated with both cFXIII (374) and pFXIII (23, 31) that the thrombininduced release of AP-FXIII from the Leu34 FXIII-A variant, as well as the consequent activation of FXIII, proceed at a 2.5-fold higher rate than in the case of the Val34 variant. Faster activation of FXIII results in accelerated fibrin cross-linking and in a higher rate of ␣2-PI incorporation into fibrin (23, 31, 330, 374). The introduction of Val34Leu and Val35Thr double mutations into FXIII-A increased its rate of activation even further (by 7.6-fold) (20). Investigation of the effect of FXIII-A Val34Leu genotype on thrombin-induced FXIII activation in the more complex environment of human plasma led to a similar conclusion (341). In plasma, the onset of FXIII activation was initiated by fibrin polymerization independently of the polymorphism; however, after initiation, the release of Leu34 AP-FXIII occurred significantly faster than that of Val34 AP-FXIII. A few pieces of biochemical evidence suggest major conformational changes of FXIII during the process of activation; however, the active form of FXIII has not been crystallized Physiol Rev • VOL 91 • JULY 2011 • www.prv.org Downloaded from http://physrev.physiology.org/ by 10.220.32.246 on June 14, 2017 The question whether AP-FXIII is released into plasma after thrombin cleavage or it remains noncovalently associated with the truncated FXIII molecule has been addressed by X-ray crystallography and by biochemical techniques. In the crystal prepared from FXIII-A2 treated with thrombin and Ca2⫹, the AP-FXIII appeared in the same position as it was in the zymogen (382). It is also known that in certain conditions the proteolytic removal of AP-FXIII is not a prerequisite of the active configuration (79, 223, 302). However, as discussed earlier, the data obtained by X-ray crystallography do not fit well with the biochemical data obtained with FXIIIa in solution. Recent findings obtained with antibodies specific for free AP-FXIII indicate that in plasma AP-FXIII dissociates from the truncated parent molecule and appears in the serum (287, 332). While noncleaved AP-FXIII is strongly hydrogen bonded to the adjacent -sandwich and to the -barrel 1 domain of opposite FXIII-A monomer, according to NMR and circular dichroism studies its released form seems unstructured (287). ⬃40%, the promoter effect of fibrin is lost (210), i.e., the formation of cross-linked product downregulates the activation of cross-linking enzyme. MULTIPLE FUNCTIONS OF FACTOR XIII While X-ray crystallographic study could not reveal significant difference in the structure of nonactivated and thrombin and Ca2⫹-activated FXIII-A2 (382), the above findings suggest a considerable conformational change during the transformation of FXIII-A2 into FXIII-A2*. It is interesting that Pinkas et al. (301) trapped TG-2 in complex with an inhibitor peptide, Ac-P(DON)LPF, that mimics a natural Gln substrate and solved the X-ray crystal structure (301). They demonstrated that the substrate analog stabilizes active TG-2 in an extended conformation, dramatically different from earlier TG structures. No such crystal structure of FXIIIa has been produced. Based on the homology with TG-2, a molecular model of FXIII-A2* with extended conformation has been developed, and this model showed good correlation with the biochemical data (187). Although in physiological conditions pFXIII is generally activated by the concerted action of a protease (thrombin) and Ca2⫹, proteolytic cleavage is not an absolute requirement for the activation of the tetramer molecule. In nonphysiological conditions, i.e., at extremely high Ca2⫹ concentrations (ⱖ100 mM), the A2B2 complex dissociates and the nontruncated FXIII-A2 becomes transformed into an active TG (FXIII-A2°) (79, 223). The occupancy of lowaffinity Ca2⫹ binding sites might play a role in this process. In sharp contrast to the nonproteolytic activation of pFXIII, low Ca2⫹ concentration is sufficient to induce a slow progressive activation of cFXIII at physiological ionic strength (302). The rate of this nonproteolytic activation of cFXIII is greatly enhanced by increasing the ionic strength, and complex formation with FXIII-B fully abrogates the activation process (302). This finding suggests that in the case of pFXIII, high Ca2⫹ concentration is only required for the dissociation of FXIII-B from FXIII-A2. In extracellular conditions, cFXIII could be activated by thrombin and Ca2⫹ the same way as pFXIII, excluding the dissociation of FXIII-B. It was suggested that cFXIII on platelets is activated by the cysteine protease calpain (21). However, in the intracellular environment, cFXIII does not need proteolytic cleavage for activation, and during platelet activation, no proteolytic truncation of FXIII-A occurs (261, 262). The elevation of intracellular Ca2⫹ concentration, as a consequence of cell activation, seems sufficient to bring about the enzymatically active configuration (FXIIIA2°). The activation of cFXIII by the nonproteolytic mechanism (i.e., by the elevation of intracellular Ca2⫹ concentration) represents the physiological way of cFXIII activation in platelets (261, 262) and most likely in monocytes (1) as well. In platelets, elevation of intracellular Ca2⫹ concentration, as a result of activation induced by thrombin and Ca2⫹ ionophore, initiated the intracellular activation of cFXIII without the involvement of proteolysis (261, 262). Similarly, elevation of intracellular Ca2⫹ and stimulation of monocytes by angiotensin II resulted in nonproteolytic activation of FXIII, which catalyzed the dimerization of angiotensin receptor 1 (AT1) (1) (see more details in sect. VIIB). B. Enzymatic Reaction Catalyzed by Activated Factor XIII: Substrate Specificity TGs, including FXIIIa, catalyze an acyl transfer reaction that consists of two major steps (124, 148, 263). First, the peptide-bound glutamine substrate forms a binary complex with the enzyme (thioester linkage between the carboxamide group of the glutamine and the active site cysteine). The reaction proceeds through an oxyanion intermediate, and ammonia is released as the acyl-enzyme intermediate is formed. In the second step, if a substrate primary amine group is present, the acyl group is transferred to the acyl acceptor amine through a second oxyanion intermediate. The amine becomes attached to the ␥-glutamyl residue via peptide (“isopeptide”) bond, and the active-site cysteine becomes deacylated. In the absence of a substrate amine, the end result of the reaction is the deamidation of the substrate glutamine residue. If an ⑀-amino group of a peptide-bound lysine residue is the acyl acceptor primary amine for the reaction, ⑀(␥-glutamyl)lysyl is formed and the two peptide bonds become cross-linked. A more detailed description of the reactions catalyzed by TG is given in References 147 and 148. Physiol Rev • VOL 91 • JULY 2011 • www.prv.org 943 Downloaded from http://physrev.physiology.org/ by 10.220.32.246 on June 14, 2017 and X-ray structural studies could not be carried out. The active-site cysteine, originally buried within the catalytic core domain, becomes unmasked during activation and available for reaction with its substrates and also with alkylating agents (301). Hydrogen-deuterium exchange and chemical modification of amino acid side-chains (cysteine alkylation and lysine acetylation) provided further insight into structural changes occurring during the activation process. Hydrogen-deuterium exchange evaluated by MALDITOF mass spectrometry showed that FXIII activation exposes the potential substrate recognition region in the catalytic core domain (residues 220 –230), while decreased deuteration occurred in regions 98 –104 of the -sandwich and 526 –546 of -barrel 1 (364). Results from chemical modifications demonstrated that Lys156 and Lys221, the active sites Cys314 and Cys409 (both cysteines in the catalytic core domain), as well as Cys695 in the -barrel 2 become exposed upon activation by thrombin and Ca2⫹ (365). In a further study, the exposure of Cys238, Cys327, and Lys68 was also revealed (322). Experiments using an inhibitory analog of FXIIIa substrate K9 DON (a -caseinderived nonapeptide in which the substrate Gln residue was replaced by the Gln isostere, 6-diazo-5-oxo-norleucine; DON) provided further insight into the changes of FXIII-A2 structure upon activation and upon the binding of the substrate analog. K9 DON inhibited FXIIIa and provided even greater protection of the -sandwich and the -barrel 1 from deuteration. At the same time, the K9 DON was able to block the alkylation of Cys409 in the catalytic core near the dimer interface (322). MUSZBEK ET AL. The substrate specificity of FXIIIa is more restricted than that of other TGs. The primary physiological substrates of FXIIIa are fibrin and ␣2-PI (225, 324). Twenty-three additional FXIIIa substrate proteins are enumerated in the TRANSDAB database (http://genomics.dote.hu/wiki/). The substrate proteins can be divided into the following categories: coagulation factors, components of the fibrinolytic system, adhesive and extracellular matrix proteins, intracellular cytoskeletal proteins, and others (TABLE 2). The interaction of FXIIIa only with part of these proteins has physiopathological implications; these interactions will be discussed in the respective sections of this review. Most recently, an interesting study demonstrated that, in addition to the TG activity of FXIIIa, FXIII, just like TG-2, also possesses protein disulfide isomerase (PDI) activity 944 (194). Although both activities are located on FXIII-A, they are distinct. FXIII without being transformed into an active TG as well as active-site alkylated FXIIIa exert PDI activity. The physiological importance of PDI activity of FXIII still needs to be explored. C. Inactivation of Activated Factor XIII Within the Clot Extensive cross-linking of fibrin increases the resistance of fibrin clot to fibrinolysis (103, 104). This finding suggests that noncontrolled cross-linking by FXIIIa results in overcross-linked fibrin, and extensive cross-linking of other plasma proteins to fibrin, that could lead to undesired prolonged persistence of thrombi. A gradual decrease of FXIIIa activity, which has been demonstrated within the fibrin clot and in an experimental pulmonary emboli model (catalytic half-life: ⬃20 min) (315), suggests the existence of inactivation mechanism(s) that operate in whole plasma clot and in thrombi. The inactivation of FXIIIa is much less investigated than its activation. Activated clotting factors are inactivated by two mechanisms (70). Proteolytically active factors are inhibited by specific serine protease inhibitors, serpins (like antithrombin III or tissue factor pathway inhibitor), or by less specific protease inhibitors, like ␣2-macroglobulin. The other way of inactivation of active factors is performed by proteolytic enzymes. The latter can be carried out by highly specific proteases, like the cleavage of activated factor V (FVa) and factor VIII (FVIIIa), by activated protein C, or by a proteases with much broader substrate specificity, like plasmin. Plasmin degrades fibrin, fibrinogen, FVa, and FVIIIa. As no plasma protein inhibitor of FXIIIa has been discovered, one has to consider the proteolytic inactivation mechanism. The fibrinolytic enzyme plasmin could be a possible candidate for such a role. However, both plasma and cellular FXIII and their activated forms are highly resistant to plasmin (310). In addition, the powerful inhibitory effect of fibrin-linked ␣2-PI also makes it unlikely that plasmin could effectively be involved in the inactivation of FXIIIa in the fibrin clot. In contrast, polymorphonuclear granulocytes (PMNs) became activated and released proteases [for instance, elastase, cathepsin G, and matrix metalloproteinase (MMP-9)] in the fibrin clot. The released PMN proteases proteolytically degraded both FXIII subunits and inactivated FXIIIa within the fibrin clot (29). Such a mechanism also operated in clots made from whole plasma, and the main physiological inhibitor of PMN proteases, ␣1-antitrypsin, provided only limited protection. The time course of FXIIIa degradation by PMNs suggests that the proteolytic degradation of FXIIIa by the released proteases does not interfere with the initial crosslinking events; however, it could prevent the formation of over-cross-linked plasma-clot and facilitate the elimination of fibrin when it is no longer needed. It would be interesting to investigate if this mechanism also plays a role in other processes, in which FXIII is implicated and PMN leukocytes Physiol Rev • VOL 91 • JULY 2011 • www.prv.org Downloaded from http://physrev.physiology.org/ by 10.220.32.246 on June 14, 2017 As glutamine-containing peptides with different primary structure around the reactive Gln residue could serve as acyl donor substrate for FXIIIa-catalyzed TG reaction (80, 296, 356) (http://genomics.dote.hu/wiki/), the substrate-enzyme interaction could only be described for individual peptides. Oligopeptides corresponding to the NH2-terminal amino acid sequence of Asn1-␣2-plasmin inhibitor (Asn1-␣2-PI), an isoform of ␣2-PI, the major plasmin inhibitor and an excellent glutamine substrate of FXIIIa (see sect. IV for details), are the most well-studied acyl donor peptide substrates in this respect. Enzyme kinetic and NMR experiments with such 15-mer and 12-mer oligopeptides and with their chemical mutants shed some light on the role of amino acid side chains around the Gln2 substrate site in the substrate-enzyme interaction (63, 64, 232, 296). Kinetic experiments with peptides, in which Asn1 residue was either truncated or replaced by Ala, and proton NMR analysis of FXIII-A2*-N1-␣2-PI(1–12) complex demonstrated that Asn1 residue is essential for effective enzyme-substrate interaction (296). Further studies demonstrated the supportive role of Glu3, Gln4, and Lys12 side chains in the reaction (64, 296). It was shown by solution NMR methods that the reactive peptide glutamines are encountering a distinctive environment within the FXIIIa active site and the glutamines and residues located COOH-terminally come in direct contact with the enzyme and adopt an extended conformation (232, 296). It is interesting that the replacement of Gln4 by Ser or Leu doubles the catalytic efficiency of the reaction, indicating that the FXIIIa subsite for this residue must be relatively broad (63). Experiments with COOHterminally truncated peptides proved that amino acid residues 7–12 are essential for the interaction of N1-␣2-PI(1– 12) with the enzyme and suggested the existence of a secondary binding site on FXIII-A2*. Hydrophobic residues, particularly Leu10, and the COOH-terminal Lys12 seemed especially important in this respect, and direct interaction between hydrophobic COOH-terminal residues and FXIIIA2* has been demonstrated by STD NMR (296). MULTIPLE FUNCTIONS OF FACTOR XIII Table 2 Protein Coagulation factors Substrate proteins of activated factor XIII Substrate Sequence Around Reactive Gln Fibrin(ogen) ␣ chain 77, 234, 312, 348 2, 60 105, 140, 372 181, 204 49, 212 37, 38 367 229 328, 347 99, 238 168, 349 27 65 69 1 298 39 249 ND, not determined. accumulated. For instance, wound healing could be a target of such an investigation. IV. THE ACTION OF ACTIVATED FACTOR XIII ON FIBRIN: ITS ROLE IN THE REGULATION OF FIBRINOLYTIC PROCESS FXIIIa cross-links fibrin ␥- and ␣-chains into ␥-chain dimers and ␣-chain polymers, respectively. (For detailed information on fibrinogen and fibrin clot structure, see References 93, 250, and 376.) The extremely rapid ␥-dimer formation is the result of reciprocal intermolecular bond formation between ␥406 lysine of one ␥-chain and ␥398/399 glutamine residue of another aligning ␥-chain (60). Cross-linking of ␣-chains, a much slower process, occurs among multiple glutamine and lysine residues, resulting in ␣-oligomers and high Mr ␣-polymers. A small amount of ␣-␥-chain heterodimers and ␥-chain trimers or tetramers have also been identified in fibrin (251, 343), and their presence was related to increased resistance to fibrinolysis (344). Although ␣-chain cross-linking confers the final stability to the fibrin clot allowing strength, rigidity, and resistance to fibrinolysis (103, 106), ␥-chain dimerization also contributes to clot stiffness (117, 353). It has been reported that ␥A␥= fibrin becomes more extensively cross-linked by activated FXIII than fibrin containing only ␥A homodimers (97, 247, 248). In sharp contrast to these findings, Siebenlist et al. (345) found that the overall rate of ␣-polymer formation was slower for ␥A␥= fibrin than for ␥A␥A fibrin and suggested that the presence of ␥=-chain suppresses FXIII-induced fibrin cross-linking. An interesting question is whether cFXIII, present in platelets incorporated into thrombi, contributes to the crosslinking of the fibrin clot. Elevated platelet count and pFXIII concentration both increase the formation of highly crosslinked ␣-polymers (104). In the presence of normal platelets, the fibrin clot formed from FXIII-free fibrinogen becomes cross-linked (308); however, the contribution of platelet FXIII, compared with the contribution of plasma FXIII, was only negligible (132). Platelet FXIII is neither secreted nor exposed to the platelet surface during platelet activation (167, 265). The tiny amount of pFXIII, taken up by platelets and packed into the ␣-granules, could be released during clot formation (233), but its involvement in Physiol Rev • VOL 91 • JULY 2011 • www.prv.org 945 Downloaded from http://physrev.physiology.org/ by 10.220.32.246 on June 14, 2017 -S-Q221-L-Q-K-V-P-P-Q237-M-R-M-E-L-N-Q328-N-P-G-S-P-G-Q366-W-H-S-E-S-G-Q398-Q399-H-H-L-G-GFibrin(ogen) ␥ chain Factor V N.D. ␣ -Plasmin inhibitor ( ␣ -antiplasmin) -N-Q -E-Q-V-S-PFibrinolytic proteins 2 2 2 Lipoprotein A ND Plasminogen -S-Q322-V-R-W-E-Y Procarboxypeptidase B/U -F-Q2-S-G-Q5-V-L-A-A-L-S-Q292-H-I-V-F-P(thrombin-activable fibrinolysis inhibitor; TAFI) Adhesive/matrix proteins Thrombospondin ND Vitronectin -E-Q93-T-P-V-L-KFibronectin -A-Q3-Q-I-V-Q-POsteopontin -S-Q34-K-Q36-N-L-L-A-PCytoskeletal proteins Vinculin ND Actin -H-Q41-G-V-M-V-GMyosin ND -L-Q315-L-L-K-Y-IOthers AT1 receptor Semenogelins I and II ND Protein synthesis initiation factor ND 5A ␣2-Macroglobulin -Q-Q670-Y-E-M-H-G- Reference Nos. MUSZBEK ET AL. the cross-linking reaction is insignificant. Platelets, however, have another effect on the cross-linking reaction that is independent of their FXIII content. When fibrin cross-linking was induced by highly purified plasma FXIII, the formation of ␣-chain polymers and the incorporation of ␣2-PIfibrin ␣-chain heterodimer into these polymers was significantly accelerated by intact FXIII-free platelets obtained from patients with severe FXIII-A deficiency (132). The results indicate that, in physiological conditions, platelets promote the cross-linking reaction by providing a catalytic surface, on which the cross-linking of fibrin polymers is enhanced. In addition, activated platelets bind pFXIII through fibrinogen (265) and could deliver it to the thrombus. Cross-linking of fibrin chains by FXIIIa, in the absence of ␣2-PI, decreased the rate of lysis induced by exogenous plasmin (103). FXIII, in excess to its normal plasma concentration, increased the extent of fibrin cross-linking and inhibited plasmin-induced degradation of fibrin (103, 104). However, the lysis rate for cross-linked fibrin prepared from plasma was much slower than the lysis rate of crosslinked fibrin prepared from purified fibrinogen, and 10-fold higher plasmin concentration was needed to bring the lysis rate of plasma-derived fibrin to that of fibrin formed from purified fibrinogen. This finding emphasizes that plasma component(s) incorporated into the clot are required for the effective protection of fibrin from fibrinolysis. The impaired binding of plasminogen to cross-linked fibrin (237, 326) might also play a role in the FXIII-induced resistance to fibrinolysis. ␣2-PI, the main physiological inhibitor of plasmin, is an excellent substrate of FXIIIa, and it becomes cross-linked to fibrin ␣-chains during the initial phase of cross-linking reaction (324). It is synthesized in the liver and becomes secreted as a protein of 491 amino acids starting with a methionine (Met1-␣2-PI). In the plasma, a protease, antiplasmin cleaving enzyme (APCE), cleaves off the NH2-terminal propeptide of 12 amino acid residues (202) and transforms Met1-␣2-PI into Asn1-␣2-PI (32, 357). The transformation is only partial; in plasma, the ratio of Met1-␣2-PI to Asn1␣2-PI is 3:7. Only the Asn1-␣2-PI isoform is a good substrate for FXIIIa (357). It provides a Gln residue penultimate to Asn1 (Gln2; Gln14 in Met1-␣2-PI) to the crosslinking reaction. The same Gln residue is sheltered by the NH2-terminal 12-residue peptide of Met1-␣2-PI and makes it inaccessible for FXIIIa (203). In fact, the addition of only three COOH-terminal amino acids of the propeptide makes Asn-␣2-PI a poor substrate for the TG reaction (136). With the use of recombinant Gln2Ala ␣2-PI mutant, three further 946 During the formation of the fibrin clot, the Gln2 site becomes cross-linked to Lys303 residue of ␣-chain of fibrin (181). It is interesting that this Lys residue is not involved in the formation of ␣-chain polymers. The formation of ␣2-PIfibrin ␣-chain heterodimers is a rapid reaction; it only slightly lags behind fibrin ␥-chain dimerization and precedes fibrin ␣-chain cross-linking. It can also be cross-linked to fibrinogen, although at a significantly slower rate. The existence of ␣2-PI cross-linked to plasma fibrinogen has also been reported (252). There is no acyl acceptor lysyl residue in the ␣2-PI molecule; therefore, no ␣2-PI dimer is formed (360). Only Asn1-␣2-PI is cross-linked to fibrin, which explains that part of ␣2-PI remains in the serum. Cross-linked ␣2-PI retains its full inhibitory activity and makes fibrin strikingly resistant to digestion by plasmin (202, 325). The concerted action of FXIII and ␣2-PI plays a key role in the protection of fibrin clots against prompt elimination by fibrinolysis. The dodecapeptide with the NH2-terminal sequence of Asn1-␣2-PI was shown to inhibit the cross-linking of ␣2-PI to fibrin, resulting in accelerated fibrinolysis (182). ␣2-PI deficiency is a very rare, but severe bleeding diathesis with symptoms similar to those of FXIII-A deficiency (56). Phenylglyoxal inactivated form of ␣2-PI (205) or the active site Arg364Ala mutant (206) enhanced urokinase-induced plasma and whole blood clot lysis significantly by competing with the native ␣2-PI for FXIIIa catalyzed incorporation into fibrin. Jansen et al. (158) reported that the tissue plasminogen activator (tPA)-induced lysis of whole blood clot from a patient with ␣2-PI deficiency was not or only slightly increased by an antibody that inhibited FXIIIa. This finding suggests a primary role of ␣2-PI to fibrin cross-linking in the reduction of clot lysis by FXIII. However, Sakata et al. (326) demonstrated in ␣2-PI depleted plasma, and in purified system without ␣2-PI, that fibrin cross-linking itself and the consequent decrease in the binding of plasminogen to fibrin also contribute to the reduction of fibrinolysis by FXIII. The above findings clearly prove that FXIII has a major role in protecting fibrin from degradation by the fibrinolytic system. Most recently, the protective effect of FXIII against fibrinolysis was also demonstrated in clots formed under flow (264). Fibrin cross-linking and the binding of ␣2-PI to fibrin by FXIIIa might work together or consecutively. Using a unique antibody, which inhibited FXIIIa-fibrin and FXIIIa-fibronectin interaction but did not influence the cross-linking of ␣2-PI to fibrin, McDonagh and Fukue (235) demonstrated that both ␣2-PI-fibrin cross-linking and Physiol Rev • VOL 91 • JULY 2011 • www.prv.org Downloaded from http://physrev.physiology.org/ by 10.220.32.246 on June 14, 2017 FXIII may influence fibrinolysis by two mechanisms: 1) the cross-linking of fibrin ␣-chains into high-Mr ␣-polymers may render the clot more resistant to fibrinolysis, and 2) the binding of ␣2-PI, and perhaps also other plasma components, to fibrin may protect the fibrin clot and prevent its prompt elimination by the powerful fibrinolytic system. glutamine donor sites were found in the molecule at positions 21, 419, and 447. However, even in the mutant protein, their kinetic efficiency was much lower than that of the Gln2 residue (204). It has been shown that the Arg6Trp polymorphism in Met1-␣2-PI affects its cleavage rate by APCE in favor of the Arg6 variant, but this polymorphism does not affect the poor cross-linking of Met1-␣2-PI to fibrin (203). MULTIPLE FUNCTIONS OF FACTOR XIII ␣-chain polymerization were necessary to regulate the rate of fibrinolysis. In experimental pulmonary embolism, both FXIIIa-mediated fibrin cross-linking and ␣2-PI-fibrin crosslinking increased the resistance to endogenous and tPAinduced fibrinolysis (307). In summary, the above studies indicate that the cross-linking of ␣2-PI to fibrin occurs at an early stage of thrombus formation and provides protection of the newly formed fibrin clot against the prompt elimination by the fibrinolytic system, while the increased resistance to thrombolysis of matured thrombi could be the consequence of the much slower, extensive cross-linking of ␣-chains, including ␣-chains to which ␣2-PI had been attached. Thrombin activatable fibrinolysis inhibitor (TAFI; also known as procarboxypeptidase U, procarboxypeptidase B, or procarboxypeptidase R) plays an important role in the regulation of fibrinolysis (71, 253, 311). TAFI is activated by thrombin, a process that is greatly enhanced in the presence of thrombomodulin. Activated TAFI inhibits fibrinolysis by abrogating the following plasmin-mediated positive-feedback mechanisms of fibrinolysis. Once formed, plasmin starts to digest the clot by catalyzing cleavages after selected arginine and lysine residues. These cleavages provide extra binding sites to Glu plasminogen and tPA by exposing COOH-terminal lysine residues. This way the cofactor activity of fibrin in Glu plasminogen activation increases. In addition, the partially cleaved fibrin serves as a cofactor for the conversion of Glu plasminogen into Lys plasminogen by plasmin. The activation of Lys plasminogen by tPA is ⬃20-fold more efficient than the activation of Glu plasminogen. TAFI interferes with these positivefeedback mechanisms by removing the newly exposed COOH-terminal lysine residues of degraded fibrin, and thereby diminishing the catalytic efficiency of plasmin formation. TAFI has been shown to be a substrate for FXIIIa (367), which catalyzed the polymerization of TAFI and its cross-linking to fibrin. TAFI contains both acyl donor Gln and acyl acceptor Lys residues; Gln2, Gln5, and Gln294 are the preferred acyl donor sites. It has been suggested that the cross-linking may facilitate the activation of TAFI, stabilize the enzymatic activity, FXIIIa catalyzed the incorporation of labeled amines and a Gln-containing substrate peptide into plasminogen, demonstrating that it contains both glutamine and lysine donor sites (37). FXIIIa can polymerize plasminogen into high Mr multimers and form fibronectin-plasminogen heteropolymers. The physiological significance of these findings, if any, remains to be elucidated. The protein part of lipoprotein (a) [Lp(a)], an atherogenic risk factor, strikingly similar to plasminogen, also contains acyl donor glutamine residues, for the FXIIIa catalyzed TG reaction (49). In native Lp(a), part of the substrate glutamyl sites are buried and inaccessible for FXIIIa. Lp(a) competes with plasminogen and tPA for binding sites on fibrin(ogen) and inhibits plasmin generation (227). FXIIIa cross-links Lp(a) to fibrinogen (317), but the concentration of FXIII in this experiment was well above normal plasma concentration, and the incubation times were too long to support the physiological relevance of this interaction. In summary, FXIII is an essential component of the clotting system; its deficiency causes severe bleeding diathesis. The main function of FXIII in hemostasis is the stabilization of newly formed fibrin. By cross-linking fibrin ␥-chains into dimers and fibrin ␣-chains into high-molecular-weight polymers, FXIII increases the rigidity and strength of the fibrin clot and protects it against shear stress in the circulation. FXIII also protects fibrin from the prompt elimination by the fibrinolytic system. This mechanism involves the covalent cross-linking of ␣2-PI, the major plasmin inhibitor, to fibrin strands, the diminished binding of plasminogen to cross-linked fibrin, and the reduced lysis of cross-linked fibrin by plasmin. V. FACTOR XIII IN WOUND HEALING AND ANGIOGENESIS A. Clinical Data and Animal Experiments Supporting the Involvement of Factor XIII in Wound Healing The first FXIII-deficient patient, recognized by Duckert et al. in 1960, demonstrated impaired wound healing and abnormal scar formation, in addition to the severe bleeding complications (94, 331). The frequency of poor wound healing in severe FXIII deficiency was reported between 14 –36% in various reviews (22, 44, 154, 336). The essential role of FXIII in the wound healing process was clearly proven in transgenic mice, in which the targeted deletion of FXIII-A gene caused complete FXIII-A deficiency (199). In Physiol Rev • VOL 91 • JULY 2011 • www.prv.org 947 Downloaded from http://physrev.physiology.org/ by 10.220.32.246 on June 14, 2017 There are also other interactions between FXIII and components of the fibrinolytic system. It has been shown that plasminogen activator inhibitor 2 (PAI-2), an inhibitor of urokinase-type plasminogen activator (uPA), is cross-linked to fibrinogen or fibrin by FXIIIa (163, 313). Cross-linked PAI-2 retained its activity and effectively inhibited uPAmediated fibrin clot lysis. PAI-2 is detectable in the plasma only during pregnancy; although it is mainly of cellular localization, peripheral blood monocytes incorporated into the fibrin clot are capable of secreting it. The cross-linking of PAI-2 to fibrin may serve compartmentalization of the inhibitor. Monocytes incorporated into thrombus could therefore play a role in the downregulation of plasminogen activation (313). and protect the active enzyme from further degradation. However, no experimental support has been provided to this hypothesis, and the long incubation time required for the cross-linking questions the physiological implications of this mechanism. MUSZBEK ET AL. the FXIII-A-deficient mice, the healing of excisional wound was considerably delayed (FIGURE 8); incomplete reepithelization, persisting necrotized fissure, and abnormal scar formation were observed at day 11 after excision, when wild-type mice demonstrated complete closure of the wound (150). Substitution with human pFXIII restored the normal healing process (FIGURE 8). The influence of FXIII polymorphisms on the prevalence and presentation of CVLU has been intensively studied by Gemmati and co-workers (111-113, 361) FXIII-A Val34Leu, Tyr204Phe, Pro564Leu, and FXIII-B Arg95His did not influence the prevalence of CVLU (361). However, the presence of Leu34 and Leu564 alleles was associated with smaller ulcer surface, and the size of ulcer inversely related to the number of Leu34 alleles (111, 113, 361). Healing time was increased in patients with the Val34Val genotype compared with Leu34 carriers (111). Furthermore, FXIII-A Leu34 beneficially influenced the healing time after superficial venous surgery (112). The above results suggest that FXIII is involved in the pathomechanism of the healing process of venous leg ulcer, its topical application is beneficial, and the accelerated rate of FXIII activation and altered fibrin structure in FXIII-A Val34Leu polymorphism decrease the severity of the disease. Further details on this topic can be found in an excellent recent review (386). Early sporadic reports demonstrated the beneficial effect of FXIII supplementation on the healing of surgical wounds in postoperative situations (114, 115, 246). In inflammatory bowel disease, ulcerative colitis, and Crohn’s disease, the compensation of consumption and loss of FXIII into inflamed tissue by substitution seems to improve the healing tendency (133, 226, 337). Treatment 948 When acquired FXIII deficiency was induced in rats by CCl4 treatment, the healing of microvascular anastomosis and burn injury were impaired, but the healing process could be normalized by the administration of FXIII (153, 284). The healing of osteotomies in sheep could also be stimulated by FXIII, and the bone formed in this condition had higher tensile strength (62). B. Effect of Factor XIII on Fibroblasts, Macrophages, and Extracellular Matrix Fibrin gel formation, macrophage invasion, fibroblast migration/proliferation, production of extracellular matrix, and angiogenesis are the key events in wound healing. Earlier and recent studies enlightened certain details of the involvement of FXIII in these mechanisms. FXIII exerts its effect by cross-linking extracellular matrix proteins, by affecting fibroblasts and macrophages, and by promoting the complex mechanism of angiogenesis. The first publication in this field came directly after discovering the first FXIII-deficient patient. Beck et al. (36) demonstrated that growth of fibroblasts in the plasma of this patient was impaired, the contact between cells was lost, their normal elongated cell shape became irregular, and the cells failed to produce collagen fibers. Supplementation with normal plasma or with partially purified FXIII corrected the defects; however, a higher amount of FXIII was required for the correction of growth impairment than for restoring normal fibrin cross-linking. The enhancement of fibroblast proliferation by cross-linking fibrin matrix (366) and by direct proliferation-inducing effect of FXIII (54) were both reported in early preliminary studies. Another study indicated that FXIII binds to the extracellular matrix and matrix assembly sites, where it remains active (34). Its binding to a putative cell surface receptor, which mediates internalization and degradation, was also suggested. The migration of fibroblasts into fibrin gel was greatly enhanced by extensive cross-linking of fibrin ␣-chains, while the dimerization of ␥-chains was not sufficient to support fibroblast migration (53). Fibronectin, an important component of extracellular matrix, is cross-linked by FXIIIa at cellular matrix assembly sites (35), which complements disulfidebonded multimer formation in the stabilization of assembling fibronectin molecules. Fibronectin covalently incorporated into the fibrin clot by FXIIIa supports fibroblast adherence and promotes the migration of cells into the gel (126). Cell attachment and spreading on composite fibrin/fibronectin clot significantly decreased when recombinant fibronectin with mutations in the FXIIIa substrate glutamine sites was used, i.e., fibronectin must be crosslinked to fibrin by FXIIIa to ensure maximal cell attachment (74). In addition to fibrin and fibronectin, several Physiol Rev • VOL 91 • JULY 2011 • www.prv.org Downloaded from http://physrev.physiology.org/ by 10.220.32.246 on June 14, 2017 Chronic venous leg ulcer (CVLU) represents a peculiar form of impaired wound healing with multicausative background. The possible involvement of FXIII in the pathomechanism of this disease is of particular interest. An earlier study demonstrated decreased FXIII level in a small group of patients with CVLU (369), while in a more recent report FXIII-A antigen level did not differ significantly between CVLU patients and matched controls (n ⫽ 91 in both groups) (113). However, in the latter study, 11% of the cases had moderate FXIII deficiency (FXIII-A antigen ⱕ0.65 U/mL). Although only a relatively small number of patients were recruited for the four studies evaluating the effect of topical treatment of CVLU with FXIII concentrate, the studies unanimously reported a beneficial effect (131, 134, 297, 380). Accelerated wound surface reduction, shorter healing time, improved availability of granulation tissue, and decreased secretion and bleeding tendency were observed following daily local administration of the concentrate in CVLU, but not in arterial-venous mixed disease (380). with recombinant FXIII also improved established experimental colitis in rats (83). MULTIPLE FUNCTIONS OF FACTOR XIII other adhesive proteins, vitronectin, osteopontin, thrombospondin, and von Willebrand factor, are substrates of FXIIIa (TABLE 2). Their cross-linking to other connective tissue matrix components might have further impact on the attachment of cells to the matrix and on the healing of injuries. Macrophages have a well-known important role in the wound repair process (3, 118, 119). FXIII of tissue macrophages might play a role in maintaining and modulating the connective tissue matrix in normal conditions. This hypothesis, however, still lacks experimental support. In theory, FXIII of the extracellular compartment might act on monocytes/macrophages by influencing their activation, differentiation, and migration, while cFXIII present in these cells might play a role in the mechanism of macrophage migration and phagocytosis. Like in the case of fibroblasts, FXIIIa enhanced the proliferation of peripheral blood monocytes, accelerated their migration through the filter of a Boyden chamber by twofold, and significantly inhibited monocyte apoptosis (85). These changes were related to the downregulation of thrombospondin-1 and to the upregulation of c-Jun and Egr-1 in monocytes. Active-site blocked FXIIIa was ineffective, suggesting that the TG activity of FXIIIa is essential for its effects on monocytes (85). The above results seem to be in contradiction to the finding that anti-FXIII-A antibodies did not influence the binding of THP-1 cells (a human monocytic leukemia cell line) to fibronectin and their migration on fibronectin surface (16). However, the leukemic cells might behave differently from normal monocytes, and the inhibitory effect of the antibody was not proven in this publication. It was also shown that in fibrin gel containing cross-linked fibronectin the migration of monocytes slowed down (197). Although the relation of migration in fibrin gel to the migration required for wound healing is questionable, the adhesion to cross-linked fibronectin might represent an anchoring mechanism that keeps macrophages where they are needed for the wound healing process. The involvement of monocyte/macrophage cFXIII in the wound healing process has not been explored to a full extent. FXIII-A2 cannot be secreted by the classical pathway, and there is no proof for its secretion by other secretory mechanisms. Nevertheless, leakage of cFXIII from damaged cells might contribute to the local cross-linking of proteins. The activation of monocytes/macrophages by the alternative pathway, for instance, by interleukin-4, highly increased FXIII-A expression (362). Alternatively activated or M2 macrophages express scavenger receptors, bind apoptotic cells, and are important in mediating wound healing following tissue damage (96). cFXIII is involved in receptor-mediated phagocytosis by monocytes (6, 329). Removal of cell debris and apoptotic cells is an important step in the wound healing process, and the decreased phagocytosis of monocytes/ macrophages in FXIII-A deficient patients might contribute to the impaired tissue repair. Another intracellular mechanism involving cFXIII in wound healing could operate through the angiotensin II-induced release of monocytes from their splenic reservoir and migration to the site of injury (358). Binding of angiotensin II to its receptor (AT1) could lead to cFXIII activation and consequently to the covalent dimerization of AT1 by FXIIIa, which induces the activation and mobilization of monocytes (1). (The expression and activation of cFXIII in activated monocytes and the involvement of cFXIII in the function of these cells are detailed in section VIIB.) C. Proangiogenic Effect of Activated Factor XIII Angiogenesis is a complex process that includes remodeling and sprouting of new capillaries from existing Physiol Rev • VOL 91 • JULY 2011 • www.prv.org 949 Downloaded from http://physrev.physiology.org/ by 10.220.32.246 on June 14, 2017 A recent study confirmed and extended sporadic preliminary results on the direct effect of FXIII on fibroblasts using more elaborate techniques; this study also shed light on the mechanism by which FXIII exerts its effect (85). FXIIIa, but not nonactivated FXIII or inactivated FXIIIa, bound to skin fibroblast, enhanced their migration in a wound-migration assay, increased the incorporation of [3H]thymidine into the cells, and decreased fibroblast apoptosis. The finding that an antibody against ␣v3-integrin, the vitronectin receptor, blocked the binding of FXIIIa to fibroblasts implicated this receptor in the effect exerted by FXIIIa (85). In addition, FXIII counteracted the toxic effect of metalloproteinases on dermal fibroblasts, although it is not clear from this study if FXIII added to the cell culture became activated or not (385). Another interesting aspect of the relationship between macrophages and FXIII is the FXIII-dependent generation of a complement C5-derived monocyte chemotactic factor during clotting (286). The major monocyte chemotactic factor identified in the serum had a Mr (75 kDa) by gel filtration that is much higher than that of the active complement fragment C5a or C5a des-Arg (8 kDa). This finding suggests that C5a was cross-linked to another plasma protein, although no direct evidence has been provided. Most recently, a plasma protein with the features of ribosomal protein S19 (RP S19) was identified, which was dimerized by FXIIIa on the surface of activated platelets during coagulation and converted into a monocyte-selective chemoattractive factor (339). Interestingly, RP S19 showed immunological cross-reactivity with C5a. However, the Mr of the active dimeric form was only 32 kDa. It remains to be seen what is the exact relationship between the cross-linked C5a fragment and the active ribosomal protein S19-like factor. Such monocyte specific chemotactic factor(s) formed during clotting could be involved in recruiting monocytes/inflammatory macrophages to the site of injury. MUSZBEK ET AL. Downloaded from http://physrev.physiology.org/ by 10.220.32.246 on June 14, 2017 Physiol Rev • VOL 91 • JULY 2011 • www.prv.org 950 MULTIPLE FUNCTIONS OF FACTOR XIII blood vessels. It is involved in a wide range of physiopathological processes from wound healing to tumor growth. The results published in the last decade strongly suggest that the role of FXIII in tissue repair and wound healing is at least in part mediated by its proangiogenic effect. The interaction of FXIII with immortalized human endothelial cells was first reported a decade ago by Dallabrida et al. (84) and by Dardik et al. (89). The binding itself did not require TG activity, nonactivated FXIII, FXIIIa, and inactivated FXIIIa all bound to the cells via ␣v3 integrin (vitronectin receptor) and activation of the receptor by MnCl2 highly increased the binding (84, 89). In addition, FXIII seems to be capable of mediating endothelial cell-platelet interaction (89). Dardik et al. (90) also demonstrated in the Matrigel tube formation model that FXIIIa caused a dose-dependent enhancement array formation of HUVECs. No effect was observed on the addition of FXIII or iodoacemide inhibited FXIIIa, or when the TG activity of FXIIIa was blocked by a polyclonal antibody (90). Interestingly, an opposite effect was observed by Dallabrida et al. (84) using a simian virus 40 (SV40) transformed human dermal microvascular endothelial cell line (HMEC-1) and fibrin gel. Differences in the features of the two cell types, e.g., in their TSP-1 content The role of FXIII in angiogenesis has also been verified in several in vivo animal models. Injection of FXIIIa into the cornea subepithelially induced the formation of a rich network of capillaries within 36 h. At the same time, the positive staining for TSP-1 observed in the fibers of the stroma and in keratocytes of nontreated cornea disappeared (90). Topical application of FXIIIa produced a significantly higher number of new vessels in neonatal heterotopic mouse heart allograft model and increased the contractile performance of the allograft compared with untreated normal mice (86). Here again, FXIIIa decreased TSP-1 mRNA and protein expression. These findings also demonstrate that FXIIIa exerts its proangiogenic effect, at least in part, through the downregulation of TSP-1. In FXIII-A knockout mice, the formation of new vessels in subcutaneously injected Matrigel plug dramatically decreased compared with normal mice (86). If FXIIIa was added to the Matrigel, the number of vessels almost reached the normal level. When a critical-sized bone defect in the tibia of rat was filled with hydroxyapatite, the addition of FXIII stimulated the in-growth of microvessels into the implant (180). Although FXIII added to hydroxyapatite was not preactivated, it was transformed somehow into its active form locally as demonstrated by staining with an antibody specific for the active form. The mechanism of the proangiogenic effect of FXIIIa was partly enlightened in Inbal’s laboratory. FXIIIa bound to ␣v3 receptor enhances its noncovalent interaction with VEGFR-2, and even resulted in partial cross-linking between the 3 subunit of vitronectin receptor and VEGFR-2 (88). FXIIIa then induced tyrosine phosphorylation and activation of VEGFR-2 that led to the phosphorylation and activation of Akt and the extracellular signal-regulated protein kinases (ERKs) and to the upregulation of the transcription factors c-Jun and Egr-1. Akt promotes growth factor-mediated cell survival and has been implicated in angiogenesis. The activation of ERKs together with the zinc-finger transcription factor, Egr-1-mediated pathway promotes cell proliferation. As mentioned above, the downregulation of TSP-1 is a key event in the proangiogenic effect of FXIIIa. Overexpres- FIGURE 8 Impaired wound healing in FXIII-A knock out mice. A: representative views of the gross appearance of excisional wounds of control, FXIII-A-deficient, and FXIII-A-deficient/pFXIII-treated mice on days 0, 8, and 11. B: follow up of wound scores at different time points of control (squares), FXIII-A-deficient mice (open triangles), and FXIII-A-deficient/pFXIII-treated mice (black triangles). Each group included 10 animals. Data are presented as means ⫾ SD. Mean ⫾ SD area under the curve (AUC) was calculated from the wound scores. AUC was significantly higher in either the control or FXIII-treated groups compared with the FXIII-A-deficient animals (P ⬍ 0.001 in both cases). [From Inbal et al. (150), with permission.] Physiol Rev • VOL 91 • JULY 2011 • www.prv.org 951 Downloaded from http://physrev.physiology.org/ by 10.220.32.246 on June 14, 2017 Although the binding of the nonactivated form might be important for targeting FXIII to endothelial cells, its active form is required to exert proangiogenetic effects. FXIIIa, but not the zymogen FXIII or active-site blocked FXIIIa, enhanced human umbilical endothelial cell (HUVEC) migration in two different migration assays (90). FXIIIa also increased [3H]thymidine incorporation into HUVECs and significantly suppressed the apoptosis of these cells as demonstrated in TUNEL assay (90). FXIIIa had no effect on the secretion of vascular endothelial growth factor (VEGF) into the medium, and neither VEGF protein level nor steady-state VEGF receptor-2 (VEGFR2) mRNA levels were affected by FXIIIa. In contrast, FXIIIa induced an almost complete disappearance of thrombospondin-1 (TSP-1) mRNA from the cells and markedly reduced the amount of TSP-1 in the conditioned medium. TSP-1, a homotrimeric multifunctional glycoprotein produced and secreted by many cells, is involved in the inhibition of angiogenesis by inhibiting endothelial cell migration and proliferation and by inducing apoptosis (200, 245, 276). The interference of TSP-1 with wound healing-associated angiogenesis was also demonstrated in transgenic mice with targeted overexpression of TSP-1 in the skin (355). (90), opposing response in VEGFR-2 and VEGF expression to ␣v integrin suppression (87), might be partly responsible for the discrepancy. Another significant difference was the composition of gel matrix; Matrigel, a gelatinous protein mixture secreted by Engelbreth-Holm-Swarm (EHS) mouse sarcoma cells, resembles the complex extracellular environment found in many tissues as opposed to fibrin gel consisting of a single component. MUSZBEK ET AL. sion of c-Jun in HUVECs and in cardiac allografts has been linked to the downregulation of TSP-1 (86, 88), and this effect is mediated by Wilm’s tumor-1 (WT-1), another transcription factor (88). As the level of WT-1 is not influenced by FXIIIa, its effect seems to be exerted through the modulation of WT-1 to TSP-1 association. VI. OTHER EFFECTS OF FACTOR XIII A. Role of Factor XIII in Maintaining Pregnancy The levels of fibrinogen, factors VII, VIII, IX, and X progressively increase during pregnancy (354). A similar pattern is followed by FXIII-B but not FXIII-A. FXIII-A concentration and consequently also FXIII-A2B2 concentration and FXIII activity start to decrease between the fifth to eighth gestational week (wG) and following a gradual reduction reaches ⬃50% of normal average (41, 73, 129, 277, 368). FXIII then rises with the onset of labor and falls FIGURE 9 A hypothetic scheme for the involvement of FXIII in wound healing. pFXIII activated in the coagulation pathway plays a pleiotropic role that involves antifibrinolytic effect, modulation of extracellular matrix, promotion of monocyte/fibroblast migration, proliferation, survival, and a complex effect on endothelial cells leading to angiogenesis. cFXIII in monocytes/macrophages recruited to the site of injury becomes upregulated and activated. Activated cFXIII of monocytes/macrophages promotes phagocytosis. This way it is involved in the removal of cell debris and apoptotic cells. The evidence for individual pathways shown is discussed and evaluated in the text. 952 Physiol Rev • VOL 91 • JULY 2011 • www.prv.org Downloaded from http://physrev.physiology.org/ by 10.220.32.246 on June 14, 2017 The above findings leave no doubt that FXIIIa is a proangiogenic factor, and this function of FXIIIa is important for the promotion of wound healing. The biochemical mechanism of FXIIIa-induced angiogenesis and its participation in wound healing have been partly clarified (FIGURE 9), but there are still a number of missing links that need to be explored. Further basic and clinical research in this area may lead to the exploitation of this proangiogenic effect in clinical practice. Topical application of FXIII concentrate might be used to promote the angiogenesis in poorly healing wounds, and the inhibition of FXIIIa-induced angiogenesis might have a potential in antitumor therapy. MULTIPLE FUNCTIONS OF FACTOR XIII again postpartum. Whether the reduction of plasma FXIII during pregnancy represents decreased synthesis of FXIII-A, increased utilization/consumption, or simple dilution by expanded plasma volume is not clear. The concentration of pFXIII required for maintaining pregnancy has not been established. The observation that FXIIIB-deficient women, who have FXIII level in the range of 5–10%, do not experience miscarriage (116, 323) suggests that pFXIII concentration significantly lower than normal is sufficient to maintain pregnancy without substitution therapy. A patient with pFXIII activity, as low as 0.3%, was able to carry two pregnancies until wG 31 and 37, and only then did ablation of placenta with bleeding complications occur that needed replacement therapy and caesarian sections (243). Present recommendations for FXIII substitution of FXIII-deficient patients suggest that 10% presubstitution FXIII level is ideal to prevent miscarriages (26, 139, 173). Ogasawara et al. (283) investigated FXIII level in 424 patients with a history of first trimester miscarriages. No difference in the subsequent miscarriage rates was found between patients with normal and moderately decreased level of FXIII. The above findings clearly demonstrate that pFXIII is essential to carry out normal pregnancy, but the level required to prevent miscarriages is significantly below the reference interval of pFXIII. FIGURE 10 Distribution of FXIII-A in the definitive placenta. The major components, related to the role of definitive placenta, are indicated. FXIII-A-containing cellular and structural elements are depicted in maroon. These include macrophages in the placental villi (Hofbauer cells) and in the decidua and the Nitabuch’s fibrinoid layer. This layer is located between the zona spongiosa and zona compacta where the release of placenta will take place. The fibrinoid deposits contain fibrin and fibronectin cross-linked by FXIIIa. Physiol Rev • VOL 91 • JULY 2011 • www.prv.org 953 Downloaded from http://physrev.physiology.org/ by 10.220.32.246 on June 14, 2017 FXIII-A is not required for conception, but it is essential for maintaining pregnancy; patients with severe FXIII-A deficiency and without replacement therapy experience recurrent pregnancy losses in the first trimester (see reviews in Refs. 26, 142, 151, 173). Homozygous female FXIII-A knock out mice were also capable of becoming pregnant; however, most of them died of excessive vaginal bleeding (189). Massive placental hemorrhage with subsequent necrosis developed in the uteri by day 10. As embryonic development of the FXIII-A-deficient fetus is normal, the defect should be at the maternal side. FXIII-A has been identified in histiocytes of the uterus (11), in macrophages of the implantation tissue (25), and in the placenta (9). In the placenta small mononuclear round-shaped FXIII-A-containing cells appear at the fifth wG and then they rapidly proliferate and differentiate into large stellate cells reaching ⬃30% of total cell number (171). From the eighth wG they tend to accumulate in the peripheral part of chorionic villi. The location of FXIII-A-containing cells in the definitive placenta is demonstrated in FIGURE 10. Although the presence of FXIII-A-containing cells both in the placenta and the implantation tissue suggests the involvement of these FXIII-A-containing macrophages in the development of normal pregnancy, the simple fact that replacement by pFXIII concentrate is sufficient to carry FXIII-A-deficient patients successfully through pregnancy (47, 100, 185, 316) proves the primary importance of pFXIII. The function of cFXIII present in uterine and placental macrophages remains unclear; it might be involved in the crosslinking of extracellular matrix proteins in these tissues. MUSZBEK ET AL. It is very likely that activated FXIII cross-links and stabilizes fibrin (and fibronectin) of the Nitabuch’s layer, and protects it from fibrinolysis. The urokinase-type plasminogen activator (uPA) is abundant in placental cells (98, 101) and, together with its cellular receptor and inhibitors (PAI-1 and PAI-2), FXIII may act as a regulator of uPA-induced fibrinolysis. The fibrinoid of the Nitabuch’s layer seems to play a complex role. In addition to its sealing effects, it also acts as a protective immunologic “barrier” between feto-maternal tissues and participates in anchoring the placenta. The absence of its cross-linking by FXIIIa could compromise all these functions. The angiogenic effect of FXIIIa may contribute to the angiogenesis during placentation. Insufficient formation of cytotrophoblastic shell was also reported in woman with FXIII deficiency (24). Although the essential role of pFXIII in maintaining pregnancy has been firmly established, we are only beginning to understand its precise role and the mechanisms through which it ensures normal placental development as well as prevents peri-placental bleeding, detachment of the placenta from the uterus, and subsequent spontaneous fetal loss. B. Effect of Factor XIII on Vascular Permeability The first report concerning the influence of FXIII on vascular permeability came from Hirihara et al. (135). In their experiments the vascular permeability enhancement induced by intradermal injection of anti-endothelial cell antiserum was demonstrated by the leakage of Evans blue from vascular compartment after 15 min and by measuring the swelling after 1, 2, or 3 days, respectively. Local administration of pFXIII suppressed both the acute and the subacute increase of permeability significantly. It was not explored if FXIII became activated following administration or the inactive form was sufficient to exert the inhibitory effect. A more detailed in vitro study used porcine endothelial cell culture and ischemic-reperfused rat heart in a Lan- 954 gendorf perfusion system to address the effect of FXIII on endothelial barrier function (275). Physiological concentration of activated pFXIII or cFXIII decreased the permeability of endothelial cell monolayer for Trypan blue-labeled albumin in a two-compartment system separated by a porous filter. In addition, they were able to prevent the rapid rise of macromolecule permeability that was induced by an inhibition of endothelial mitochondrial and glycolytic energy production. Likewise, FXIIIa prevented the increase of myocardial water content induced by low-flow ischemia and subsequent reperfusion on rat heart (275). Nonactivated pFXIII and cFXIII, inactivated FXIIIa and FXIII-B failed to exert any effect in these experiments, suggesting that the effect of FXIIIa is related to its TG activity. Immunohistological investigations demonstrated the deposition of FXIIIa at interfaces of adjacent endothelial cells and between the cells and the filter support. The mechanism of the effect of FXIIIa on endothelial cell permeability has not been revealed. A variety of FXIIIa substrate adhesive proteins, like fibronectin and vitronectin, reside in the intracellular clefts and subcellular matrix; their cross-linking might influence the paracellular pathway for passage of macromolecules through the monolayer. A few clinical and experimental animal studies also support the effect of FXIII on vascular permeability. In a small prospective investigation on newborns and young children with congenital heart disease undergoing open-heart surgery, peri-operative cardiac edema formation was related to the decrease of pFXIII concentration (381). Preoperative treatment with FXIII concentrate reduced the extent of myocardial swelling and the extension of generalized edema. In another small study on children with open-heart surgery, a single intravenous dose of pFXIII reduced severe pleural effusion (333). Czabanka et al. (82) demonstrated that the administration of pFXIII together with endotoxin reduced the increase of vascular permeability in the rat, and leukocytes were not needed for this effect. The findings published, so far, clearly indicate that FXIIIa reduces the permeability of endothelial cell monolayers to macromolecules. Revealing the mechanism by which FXIIIa exerts its effect seems to be a promising and interesting field of future investigations. A few data also suggest that FXIII decreases the enhanced vascular permeability induced by various stimuli in experimental and clinical setting. However, the results of these sporadic observations and their possible clinical impact need to be confirmed by wellplanned and focused clinical and animal studies. C. Cardiac Effect of Factor XIII In 2006 a surprisingly novel aspect of FXIII-A deficiency was reported by Nahrendorf et al. (268). They compared the survival of homozygous and heterozygous FXIII-A knock out mice (FXIII-A⫺/⫺ and FXIII-A⫹/⫺, respectively), Physiol Rev • VOL 91 • JULY 2011 • www.prv.org Downloaded from http://physrev.physiology.org/ by 10.220.32.246 on June 14, 2017 The exact role of FXIII in maintaining pregnancy and the cause of pregnancy losses in women with FXIII deficiency is unknown. Two substrates of FXIIIa, fibrin(ogen) and fibronectin, are important components of the Nitabuch’s fibrinoid layer, the layer in the decidua basalis between the zona compacta and zona spongiosa, where the placenta detaches itself from the uterus at birth (FIGURE 10). Kobayashi et al. (184) reported that FXIII-A was present in the extracellular space of the extravillous cytotrophoblast forming the cytotrophoblastic shell adjacent to Nitabuch’s layer. FXIII-A colocalized with fibrinogen and fibronectin at the Nitabuch’s layer next to the cytotrophoblastic shell (24). The localization of FXIII-A supports its plasmatic origin, while the absence of FXIII-B at these locations suggests that pFXIII became activated and FXIII-B dissociated from FXIII-A. MULTIPLE FUNCTIONS OF FACTOR XIII were detected in the myocardium of patients with infarct rupture (267), and moderately decreased pFXIII levels (57 ⫾ 8%) were reported in three patients presenting with acute myocardial rupture following myocardial infarction (270). These interesting findings could provide a basis for controlled clinical studies on the risk of cardiac rupture following myocardial infarction in individuals with moderately decreased FXIII, including heterozygous FXIII-A-deficient patients. An imbalance in extracellular matrix turnover was also observed. The expression of mRNA for collagen was lower in the infarct area of FXIII-A⫺/⫺ mice, while the level of MMP-9 in the myocardial tissue of FXIII-deficient mice was sevenfold higher than in wild-type mice (268). Substitution with human pFXIII preparation reversed most, but not all, changes observed in FXIII-A⫺/⫺ mice. Postmyocardial infarction remodeling was enhanced and left ventricule enddiastolic volume was increased in spite of replenishment with pFXIII (268). The collagen synthesis remained impaired, and the scar was significantly thinner in these animals. These findings suggest that, although the lack of pFXIII plays the dominant role in the development of cardiac rupture following ligation-induced myocardial infarction in FXIII-A knockout animals, cFXIII present in the cells of cardiac tissue also contributes to the pathomechanism. FXIII-A mRNA and protein have been detected in murine cardiac tissue, and FXIII-A antigen was localized to spindleand round-shaped cells positive for monocyte/macrophage marker by immunohistochemical methods (351, 352). FXIII activity becomes gradually elevated in the myocardial infarct of wild-type mice with a maximum on day 3 (267, 268). Treatment of these animals by extra pFXIII resulted in a series of changes. It attenuated left ventricular dilation, enhanced the recruitment of macrophages in the myocardium, upregulated VEGF, and increased the formation of new microvessels and the synthesis of collagen (267). The same survival rate of homozygous and heterozygous FXIII-A knockout animals after coronary artery ligation is rather unexpected. However, FXIII has been shown to be used up during the acute phase of myocardial infarction (18, 110), and further decrease of the already-low FXIII level in FXIII-A⫹/⫺ mice could result in a situation similar to FXIII-A⫺/⫺ animals. Furthermore, diminished FXIII levels Spontaneous pathological alterations in the cardiac tissue were also observed in FXIII-A⫺/⫺ mice (352). Interestingly, these changes seemed to be gender specific. Only ⬃50% of male FXIII-A⫺/⫺ mice survived 10 mo, while 90% of female mice of the same genotype remained alive. A quarter of deceased mice died of severe intra-thoracic hemorrhage, and large hematomas were found in their hearts. Hemorrhage was also found in the heart of other deceased mice, and all FXIII-A⫺/⫺ males showed widespread hemosiderin deposition and fibrosis in the cardiac tissue. Heart structure and function of FXIII-A knockout mice did not differ significantly from their wild-type counterparts. Fibrosis and hemosiderin deposition were also present, although to a much lower extent, in FXIII-B⫺/⫺ males, but not in females. There is no clear explanation for the gender difference; it was supposed that the more aggressive fighting behavior of male animals could be one of the contributing factors. The above findings suggest that FXIII exerts its cardiac effect by multiple mechanisms. It upregulates the synthesis of extracellular matrix components, and in the case of ischemic tissue damage, it enhances microvessel angiogenesis and modulates the influx of inflammatory cells. Additional mechanisms might also be revealed in the near future. It also remains to be seen to what extent the mechanisms operating in the coronary artery ligation-induced murine infarction model are valid for human myocardial ischemic damage caused by atherothombotic events. Acute myocardial infarction developed on the basis of atherosclerosis is a more complex process in which ischemic cell death is only the end-stage of a series of events. Clinical findings suggest that FXIII might contribute to the events leading to coronary occlusion. Gene expression profiling revealed increased expression of FXIII-A in peripheral blood cells in patients with coronary artery disease (230), and an elevated FXIII level was found to be a risk factor of myocardial infarction in women (40). Detailed discussion of the interrelationship between FXIII and myocardial infarction is beyond the Physiol Rev • VOL 91 • JULY 2011 • www.prv.org 955 Downloaded from http://physrev.physiology.org/ by 10.220.32.246 on June 14, 2017 FXIII-A⫺/⫺ mice supplemented with human pFXIII and wild-type mice following myocardial infarction caused by left coronary artery ligation. FXIII-A⫺/⫺ and FXIII-A⫹/⫺ mice subjected to coronary ligation died within 5 days due to the rupture of the left ventricle wall. No death occurred in sham-operated mice, and none of the wild-type coronary ligated mice died within 5 days (only 40% of them died within 40 days). Daily intravenous replenishment of FXIIIA⫺/⫺ mice with human pFXIII restored survival to the level seen in wild-type controls. Immunohistochemistry demonstrated the presence of FXIII-A in the healing infarct of wild-type and pFXIII supplemented FXIII-A⫺/⫺ mice. Within the infarct of these animals, considerable FXIIIa activity was detected by molecular imaging using 111Inlabeled NH2-terminal ␣2-PI dodecapeptide as FXIIIa substrate (156, 268). In FXIII-A⫺/⫺ mice, a reduced inflammatory response was observed at the site of infarct; on day 2 following ligation, the recruitment of neutrophil granulocytes and macrophages to the infarct decreased by 56 and 54%, respectively (268, 269). The diminished inflammatory response is related to the impaired migration of monocytes from their splenic reservoir to the site of the injury (358). Normally the recruitment of monocytes to the site of myocardial injury operates through angiotensin II-AT1 receptor signaling (358) that very likely involves the intracellular activation of FXIII (1) (see also sects. VB and VIIB). The lack of FXIII could abrogate such a mechanism. MUSZBEK ET AL. scope of this review; for a most recent overview on this topic, see Reference 260. D. Factor XIII in Cartilage and Bone The function of chondrocyte FXIII seems to change according to the location of the cells and the stage of bone development. In the embryonic avian and mouse growth plate of long bones, it is not present in cells of proliferative zone and appears first within the cells of the zone of maturation. Its expression progressively increases until the prehypertrophic zone, and in hypertrophic cells, the expression remains at a high level (280 –282). In articular cartilage, FXIII-A-containing chondrocytes are predominantly localized in the condylar region of tarsus, suggesting that FXIII is also needed in regions that will be exposed to mechanical stress and abrasive forces. On the basis of experiments using cultured hypertrophic chondrocytes transfected with FXIII-Agreen fluorescence protein construct, it was surmised that cell death and lysis are responsible for the externalization of FXIII (280). However, this theory could not be verified by experiments on human embryonic growth plate and articular chondrocytes (165). The incorporation of a lysine-containing rhodamine-labeled tetrapeptide, an amine substrate of FXIIIa, into intracellular proteins and into extracellular matrix components indicated that, at least part of chondrocyte FXIII became activated both in the intracellular and extracellular environment (281, 282). No clear evidence was provided for its intracellular proteolytic activation in chondrocytes. However, just like in platelets, nonproteolytic activation of the zymogen, induced by increased Ca2⫹ concentration, might have been sufficient to bring about TG activity. On the basis of SDS-PAGE analysis, FXIII-A of avian chondrocyte was mistakenly considered as a monomer (282); it is very likely a homodimer, like cFXIII of other cell origin. 956 In normal human knee articular cartilage, some expression of TG-2 and cFXIII was detected in the superficial zone and in the central (chondrocytic) zone of medial menisci (165). Markedly upregulated expression of both TGs by enlarged chondrocytes was observed in the superficial and deep zones of articular cartilage obtained from patients with severe osteoarthritis. A similar finding was obtained with medial meniscal cartilage. Antibody against FXIII-A had a greater neutralizing effect on the TG activity of meniscal cell lysate than anti-TG-2 antibody, which suggests the predominance of cFXIII over TG-2. Because the cell lysates were not treated with thrombin, the actual cFXIII-to-TG-2 ratio could have been significantly higher than that measured by the authors. Interleukin-1 (IL-1) and transforming growth factor- (TGF-), whose activities are upregulated in osteoarthritis, induced increased TG-2 and FXIII-A immunostaining in knee articular and meniscal cartilage slices carried in organ culture (165). In cartilage with severe osteoarthritis, IL-1 was more effective than TGF-. The mechanism of IL-1induced TG expression is not clear. IL-1 induces nitric oxide (NO) production, and NO donors also increase TG expression, although to a lesser extent than IL-1. The results suggest that the effect of IL-1, at least in part, is exerted via increased NO generation. When TC28 cells, an immortalized cell line of human juvenile costal chondrocytes, or cultured meniscal chondrocytes were transfected with FXIII-A or TG-2, the markedly increased TG activity was accompanied by marked increase of calcium precipitates in the matrix of cultured cells (165). This effect of elevated cFXIII and TG-2 did not seem to be attributable to changes in PPi metabolism or cell apoptosis. Increased TG activity was not associated with apoptosis of the TC28 cells or meniscal chondrocytes and failed to in- Physiol Rev • VOL 91 • JULY 2011 • www.prv.org Downloaded from http://physrev.physiology.org/ by 10.220.32.246 on June 14, 2017 FXIII-A has been detected by immunohistochemical and Western blotting methods in hypertrophic chondrocytes of chicken and mouse embryonic long bone’s growth plate (281, 282), as well as in avian, porcine, and human articular chondrocytes (165, 282, 318). Chondrocytes express TG activity, which is due to TG-2 and cFXIII that coexist in these cells (165, 280 –282). The highly increased TG activities following treatment with thrombin also prove the presence of functional cFXIII zymogen in these cells (280, 319). The detection of FXIII-A mRNA indicates that chondrocytes actively synthesize this protein (280, 282, 318). FXIII-A was detected in the cytosol and membrane fractions of cultured porcine chondrocytes, more in those cells obtained from old than from young animals (318). FXIII-A also appeared in the cell culture media and in articular cartilage vesicles that participate in calcium pyrophosphate crystal formation. The mechanism of its externalization is unclear. More recent experiments reported by Johnson et al. (166) shed some more light on the mechanism by which FXIII is involved in the hypertrophic differentiation of chondrocytes. There seems to be an intriguing interplay between cFXIII and coexisting TG-2. Treatment of human articular chondrocytes with cFXIII or with TG-2 increased the expression of VEGF and MMP-13 genes, typically expressed by hypertrophic chondrocytes, and type X collagen synthesis was also enhanced. cFXIII was shown to be a ligand of ␣1-integrin, and extracellularly administered cFXIII induced ␣11-integrin-dependent signaling leading to drastic translocation of TG-2 from the cytosol to the cell surface and to consequent p38 MAP kinase and FAK phosphorylation. The latter events have a central role in transducing maturation to hypertrophy in cultured chondrocytes (241, 373, 387). As it was shown on chondrocytes from FXIII-A and TG-2 knockout mice, exogenous cFXIII required both endogenous cFXIII and TG-2 for its effect on chondrocyte differentiation (166). MULTIPLE FUNCTIONS OF FACTOR XIII duce significant changes in the extracellular PPi or nucleoside triphosphate pyrophosphohydrolase or alkaline phosphatase activity. The cross-linking and stabilization of pericellular calcium binding proteins by TGs (12, 13) might contribute to this process. The data obtained on MC3T3-E1 cells were confirmed and extended on mouse bone by the same group (271). The expression of FXIII-A protein in osteoblasts and osteocytes of long bones formed by endochondrial ossification and flat bones formed primarily by intramembraneous ossification was demonstrated by immunohistochemistry and in situ hybridization. FXIII-A mRNA was also detected in the osteoblasts of human tibial subchondral bone plates (327). Interestingly, its expression became significantly upregulated in sclerotic osteoblasts compared with nonsclerotic cells. It is rather surprising that FXIII-A in the bone extract and in MC3T3-E1 cells was found to be a 37 kDa protein (271), as opposed to the 80 kDa FXIII-A in platelets and monocytes. The expression of full-length FXIII-A mRNA in MC3T3-E1 cells and in mouse primary osteoblasts, moreover, the presence of 80 kDa protein in forskolin-treated MC3T3-E1 cells, suggest that the 37 kDa band on the Western blot represents a product of posttranslational proteolytic fragmentation (271). It is a question if the 37 kDa product is the result of a controlled, limited intracellular proteolysis or it is produced during the extraction procedure by proteolysis in the cell lysate. There are several disturbing findings concerning the former possibility. The authors also found, almost exclusively, 37 kDa FXIII-A in mouse hypertrophic chondrocytes. Similarly, a 37 kDa band together with even In conclusion, the studies of the last 8 years, described above, extended FXIII research to a new field, cartilage and bone physiopathology. Now, it is clear that chondrocytes and osteoblasts express FXIII-A, and the expression is influenced by their stage of differentiation, location, and agonists of different cellular functions. In these cells, FXIII-A coexists with another TG, TG-2, and there is an interesting functional interplay between the two enzymes. cFXIII of chondrocytes and osteoblasts becomes externalized and activated, although the biochemistry of neither of these events has been clarified. Experiments with cultured cells suggest that externalized FXIIIa (and TG-2) is involved in extracellular matrix formation, assembly, and stabilization, which seem to be important prerequisites for the process of mineralization. Investigations on FXIII-deficient patients and on FXIII-A knockout mice concerning the structure/development of bone and cartilage has not been carried out in this respect. The few available clinical data support a pathophysiological role of FXIII related to hypertrophic differentiation of chondrocytes and osteoarthritis. In the coming years, more extensive research is expected on this area that might even lay down the scientific foundation of topical recombinant FXIII therapy in certain pathological conditions or in traumatic bone injuries. VII. FACTOR XIII AS AN INTRACELLULAR ENZYME A. Involvement of Factor XIII in Platelet Function Although the presence of FXIII-A in platelets has been known for half a century, only sporadic pieces of information became available on the intracellular function of cFXIII in these cells. Platelet FXIII-A used to be considered only as a source of FXIII-A in pFXIII. However, in the light of slowly accumulating data on the intracellular activation of cFXIII and on the cross-linking of proteins within platelets, this former view could hardly be maintained any more. Physiol Rev • VOL 91 • JULY 2011 • www.prv.org 957 Downloaded from http://physrev.physiology.org/ by 10.220.32.246 on June 14, 2017 In addition to chondrocytes, cFXIII and TG-2 were also detected in cultured MC3T3-E1 (pre)osteoblasts both at mRNA and protein levels (17). Abundant TG activity was observed in the lysate of cells undergoing differentiation, which was highly increased after thrombin treatment, suggesting that cFXIII was the dominant TG in these cells. Treatment of the cells with ascorbic acid, which stimulated collagen synthesis and assembly, greatly enhanced the externalization of cFXIII, although the mechanism of externalization has not been explored. The covalent incorporation of monodansyl cadaverine (MDC), a TG amine substrate, into the extracellular matrix proved that at least part of externalized FXIII existed in active form. Fibronectin was identified as the major TG substrate matrix component. Abrogation of TG activity by cystamine decreased matrix accumulation and mineralization induced by the addition of ascorbic acid plus -glycerophosphate (17). Cystamine also blocked cell differentiation program as indicated by the low level of alkaline phosphatase. It is very likely that the lack of sufficient matrix assembly and alkaline phosphatase level both contributed to the blockade of mineralization. lower molecular weight breakdown products are shown on the Western blot of mouse macrophage lysate. In sharp contrast to these findings, full-length FXIII-A has been demonstrated in chondrocytes (282) and in mouse macrophages (320) without significant proteolytic fragmentation. Furthermore, it was also claimed that the 37 kDa FXIII-A fragment is active without proteolysis by thrombin (271), while in a previous report from the same group thrombin highly increased the TG activity in MC3T3-E1 cell extracts (17). Unless a 37 kDa FXIIIa is isolated from these cells and becomes biochemically well characterized (which sequence of FXIII-A is represented, what is its activity compared with the full-length enzyme, etc.), its intracellular existence and function remain questionable. MUSZBEK ET AL. Agonist-induced platelet aggregation in platelet-rich plasma from patients with FXIII deficiency and from FXIII-A knockout mice was normal (174, 199, 289), and washed FXIIIA-deficient human or mouse platelets showed normal aggregation response to thrombin (199, 262). Although fibrinogen binding to thrombin receptor agonist peptide (TRAP) activated FXIII-A-deficient platelets was somewhat decreased (160), this decrease does not seem to be sufficient to influence the aggregation process. Platelet aggregation is a relatively fast process; it is over in 5 min, while the crosslinking of platelet proteins is much slower (262). This suggests that cross-linked polymers are required only for the later phases of platelet activation, e.g., for spreading of platelets following adhesion or for clot retraction. The cross-linking of contractile proteins, like actin, myosin, vinculin, and filamin (27, 67, 68, 340), supports such a hypothesis. The role of platelet FXIII in clot retraction is not clear. Platelet clot retraction of FXIII-deficient patients was found to be normal (162, 306) or even enhanced (274). In contrast, reduced contractile force was measured in plateletrich plasma clot from a FXIII-deficient patient (57), and the combination of FXIII-A-deficient plasma with normal platelets was sufficient to impair isometric contraction of platelet-rich plasma clot (66). Impaired retraction of clot formed in platelet-rich plasma of FXIII-A knockout mice was observed by Kasahara et al. (174). In the latter experiments, the addition of human pFXIII or rFXIII-A2 to the platelet-rich plasma of FXIII-A knockout mice only partially restored clot retraction, suggesting that cFXIII also contributes to the process (174). However, in an earlier report, FXIII-deficient platelets induced normal clot retraction (306). The controversies concerning the involvement of pFXIII and/or cFXIII in clot retraction might be due to 958 species difference and to the considerable difference between contractile force measurement and clot retraction in a test tube. Most recently, an interesting observation was reported concerning the spreading of platelets on fibrinogen-coated surface (160). FXIII-A-deficient platelets demonstrated an altered spreading phenotype compared with normal platelets. The altered phenotype of FXIII-A-deficient platelets is characterized by higher percentage of platelets emitting filopodial versus lamellipodial extension and by a delay in the spreading process. The involvement of cFXIII in the spreading process is further supported by experiments in which MDC induced a dose-dependent increase in the proportion of control platelets showing filopodial processes (160). MDC might also influence platelet function by means other than the inhibition of protein cross-linking by FXIIIa; it can inhibit calmodulin-dependent mechanisms (76) and modify proteins at their glutamine residues. However, its ineffectiveness on the spreading of FXIII-A-deficient platelets indicates that the alteration of platelet spreading by this compound was due to the inhibition of FXIIIa-mediated protein cross-linking. Kulkarni and Jackson (192) studied the involvement of FXIII in the late stage of platelet spreading on collagen I coated surface. Fully spread platelets undergo a specific sequence of morphological changes, which the authors term sustained calcium-induced platelet (SCIP) morphology. It started with the marked contraction of lamellipodial membranes leading to microvesiculation and eventual fragmentation of the cells into a larger central membrane structure surrounded by smaller membrane bodies. SCIPs bind annexin V, indicating the translocation of phosphatidylserine, the procoagulant phospholipid, to the outer membrane surface. SCIP formation of platelets from a FXIII-A-deficient patient was decreased, and MDC also attenuated this process. It was claimed that a membrane-impermeable FXIIIa inhibitor also exerted an inhibitory effect; however, no evidence was given for the extracellular presence of FXIIIa. It seems that the activation of FXIII and calpain act synergistically in this process. The above findings indicate that the interaction of FXIII with cytoskeletal elements plays a role in its intracellular function. Results published on this area have not been assembled into a comprehensive coherent view; however, important pieces of the puzzle have been revealed. In resting platelets, FXIII-A is uniformly distributed in the platelet cytosol (340, 346, 389); however, during activation or spreading on an adhesive surface, at least part of it becomes translocated to the cytoskeletal fraction (340, 389). In platelets activated by thrombin or Ca2⫹ ionophore, cFXIII translocated to the periphery of the cells within 1 min and became associated with thin ruffle like extensions and/or pseudopods, then gradually diffused back into the central Physiol Rev • VOL 91 • JULY 2011 • www.prv.org Downloaded from http://physrev.physiology.org/ by 10.220.32.246 on June 14, 2017 Experiments with FXIII-A-deficient platelets provided important data on the role of cFXIII in platelet function. As discussed in section IIIA, FXIII becomes activated by the nonproteolytic pathway in human platelets during activation induced by thrombin or Ca2⫹ ionophore (261, 262) and cross-links platelet proteins into high Mr polymers (67, 68, 128, 262). Although only a smaller portion of platelet FXIII becomes activated, due to the very high concentration of cFXIII in these cells, this active fraction is quite sufficient to bring about the cross-linking reaction (261, 340). It is to be emphasized that in human platelets FXIII is the only TG. TG-2 protein and TG-2 mRNA could not be detected in FXIII-A-deficient or normal human platelets, TG activity is negligible in FXIII-A-deficient platelets (160, 243, 255), and no highly cross-linked protein polymers are formed in these cells (262). There might be a species difference in this respect; TG activity in the lysate of platelets from FXIIIA⫺/⫺ mice was higher than in platelet lysate from normal mice, suggesting a compensatory upregulation of another TG (164). It is clear that no such compensatory mechanism exists in humans, and protein cross-linking observed in activated platelet is entirely due to the TG activity of FXIIIa. MULTIPLE FUNCTIONS OF FACTOR XIII located to the cytoskeleton formed during platelet activation. 4) Intracellular FXIIIa is very likely involved in certain phases of the platelet spreading process and might be involved in clot retraction, at least in mice. cFXIII may interact with platelet proteins in two different ways: 1) its inactive or active form may associate with other platelet proteins without using them as substrate and move together during translocation, and 2) FXIIIa can utilize some proteins as glutamine and/or lysine donor substrates. Zhu et al. (389) demonstrated the association of FXIII with the low Mr heat shock protein HSP27 in platelets. In resting platelets, HSP27 was also found in the cytosol, but upon activation, it becomes phosphorylated and associated with the activation-dependent cytoskeleton (240). As opposed to resting platelets, in adhered platelets HSP27, in colocalization with cFXIII, was preferentially confined to the circumferential region consistent with the outermost extensions of spread platelets and to the central region corresponding to condensed granules and contractile elements (389). It is possible that HSP27 escorts FXIII to specific sites within the cells, for instance, to the cytoskeleton assembled de novo during the activation of platelets. Four other proteins, focal adhesion kinase (FAK), gelsolin, myosin IIA, and thrombospondin I, coimmunoprecipitated with cFXIII from the platelet lysate and found in colocalization with FXIII-A both in resting and activated platelets (309). Gelsolin and myosin IIA are cytoskeletal proteins, while the tyrosine kinase FAK translocates to the cytoskeleton in the second wave of cytoskeletal rearrangement (285). Direct association between cFXIII and these proteins should be confirmed in binding experiments with purified proteins. Most recently, the involvement of cFXIII in certain functions of megakaryocytes has also been reported (231). It was shown that megakaryocyte adhesion to type I collagen promoted the spreading of these cells and inhibited proplatelet formation through the release and relocation to the plasma membrane of cellular fibronectin. cFXIII was also shown to be translocated to the outer surface of plasma membrane and cross-linked fibronectin, although the mechanism of its translocation was not revealed. The inhibition of FXIIIa significantly decreased megakaryocyte spreading on type I collagen and reduced actin stress fiber formation and the assembly of fibronectin. These findings suggest that FXIII participates in the mechanism by which type I collagen, in the osteoblastic niche, suppresses megakaryocyte maturation and prevents premature platelet release. The cross-linking of two cytoskeletal elements, filamin and vinculin, during platelet activation has been analyzed in detail (340). A high portion of filamin was cross-linked to at least four different nonidentified platelet proteins in thrombin-activated platelets within 10 min. Blocking TG activity by the -SH reagent iodoacetamide completely abolished filamin cross-linking. Inhibition of fibrinogen binding to platelets and, consequently, platelet aggregation by the peptide Gly-Pro-Arg-Pro decreased the cross-linking of filamin, suggesting that it is an aggregation-induced process. The cross-linking of vinculin was less effective, and it was a platelet aggregation-dependent slower process. In summary, although the exact biochemical mechanism by which FXIII participates in platelet function still needs clarification, the available data allow the following conclusions: 1) a part of platelet FXIII becomes transformed into an active TG during platelet activation by the nonproteolytic activation mechanism. 2) Activated FXIII, the only TG in human platelets, cross-links platelet proteins in the later phases of platelet activation. 3) A few cytoskeletal proteins have been identified as substrates for FXIIIa in the crosslinking reaction, and at least part of FXIII becomes trans- B. Involvement of Factor XIII in Monocyte/ Macrophage Function Since the discovery of FXIII-A in monocytes and macrophages in 1985 (8, 130, 257), its expression in various monocyte-derived mobile and fixed macrophages has been intensively studied and turned out to be a marker reaction for various subsets of these cells in physiological and pathological settings (6, 304). The possible coexistence of cFXIII with TG-2 in monocytes and macrophages is a somewhat debated question. We could not measure any detectable TG activity in monocytes from four patients with severe FXIII-A deficiency (255), and only negligible TG activity was measured in such monocytes by the highly sensitive [3H]putrescine incorporation assay (72). No TG-2 was found in normal peripheral blood monocytes by Seiving et al. (338), and only minute TG activity was measured in nonactivated human monocyte cell lysate without thrombin activation (239, 242, 254). In the latter studies, Western blotting also detected a small amount of TG-2. The amount of TG-2 was much less than that of FXIII-A in the antigenspecific immunoprecipitates from nonactivated monocytes (16). Most recently, similar FXIII-A and TG-2 mRNA levels were measured in human monocytes; however, in this study, no attempt was made to measure the translated protein levels (161). In conclusion, overwhelming pieces of evidence suggest that in resting nonactivated monocytes, cFXIII is the major TG, and the amount of TG-2, if any, is not significant. However, during differentiation into macrophages and following activation by various agents, major changes occur in the intracellular level of both TGs. In two early studies, opposing results were obtained on the changes of cFXIII in monocyte culture. A three- to fourfold increased FXIII-A activity and antigen levels were measured Physiol Rev • VOL 91 • JULY 2011 • www.prv.org 959 Downloaded from http://physrev.physiology.org/ by 10.220.32.246 on June 14, 2017 area (340). Cytochalasin D that inhibits the polymerization of actin in intact cells abolished the translocation of cFXIII to the platelet periphery. Part of cytoskeleton-associated cFXIII existed in active form. MUSZBEK ET AL. on day 8 (72), while decreasing cFXIII with increasing TG-2 was reported in long-term culture of monocytes (338). More recently, a significant elevation of both FXIII-A mRNA and protein level was observed during the course of monocyte/macrophage differentiation in cell culture that peaked on day 3 (329). It is to be noted that the expression of TG-2 also becomes upregulated during culturing of monocytes (239, 242, 254). It is interesting that in monocytic leukemia cells, cFXIII level was also increased compared with normal peripheral blood monocytes (172). IL-4 is also involved in the in vitro differentiation of monocytes into antigen-presenting dendritic cells. Induction of such differentiation by granulocyte/macrophage-colony stimulating factor (GM-CSF) plus IL-4 resulted in very high intracellular levels of FXIII-A mRNA and protein (15, 161, 362). If dendritic cells were treated by BCG, stimulating the classical pathway, FXIII-A expression returned to the basal level within 2 days (362). Subsets of dendritic cells present in tissues, like subsets of dermal dendrocytes, also express FXIII-A. The characterization of these subsets is reviewed in Reference 304. cFXIII of perivascular macrophages has been implicated in the flow-dependent remodeling of small arteries in mice (30, 388). In this process, the perivascular accumulation of FXIII-A-positive macrophages was dependent on CXCchemokine receptor 3 (CXCR3) signaling by the CXCR3 960 Considering the high number of reports on the expression of FXIII-A in monocytes, macrophages, histiocytes, and dendritic cells, it is surprising that only a few studies have concerned with the intracellular function of cFXIII in these cells. Monocytes from FXIII-deficient patients, which are exempt of cFXIII, showed an impaired capacity of Fc␥, complement, and lectin-like receptor-mediated phagocytosis (329). In cell culture, the expression of FXIII-A and the phagocytosing capacity of monocytes/macrophages increased in parallel, and the competitive FXIIIa substrate, MDC, significantly inhibited receptor-mediated phagocytosis by these cells. DD cells (a human myelomonocytic cell line) were incapable of phagocytosis and did not express FXIII-A antigen. Partial differentiation of the cells, achieved by phorbol ester treatment, resulted in increased phagocytosis in parallel with the expression of FXIII-A (177). Jayo et al. (161) investigated the role of cFXIII in the locomotion of monocyte-derived dendritic cells. MDC inhibited both basal and chemokine (C-C motif) ligand 19 (CCL19)-induced migration of mature dendritic cells, and FXIII-Adeficient dendritic cells showed a reduced chemotactic response to CCL19. CHO cells overexpressing FXIII-A showed enhanced mobility both in trans-well and scratch wound migration assays and displayed membrane blebs and dynamic cell protrusions involved in cell movement. As both phagocytosis and cell locomotion heavily involve intracellular contractile elements, the interaction of cFXIII or its activated form with cytoskeletal components in monocytes/macrophages seems rather feasible. However, this theory still lacks experimental support. An interesting aspect of the intracellular function of cFXIII in monocytes was reported by AbdAlla et al. (1). In monocytes from hypertensive patients, covalently cross-linked angiotensin receptor 1 (AT1) dimers were detected; this dimerization was carried out by nonproteolytically activated cFXIII. The covalent cross-linking of AT1 could be reproduced by the combined addition of angiotensin II and ionomycin, which activated AT1 and, by increasing the intracellular Ca2⫹ concentration, cFXIII. No cross-linking of AT1 was observed in monocytes from FXIII-A-deficient patients, and it was also abolished by angiotensin convertase inhibitor. Cross-linked AT1 dimers displayed enhanced G␣q/11-stimulated signaling, increased internalization, and desensitization. Gln315 in the cytoplasmic side of AT1 provided the Gln substrate for the FXIIIa-catalyzed cross-linking. The appearance of cross-linked AT1 dimers correlated with the enhanced angiotensin II-dependent adhesion of monocytes to endothelial cells and endothelial dysfunction. Physiol Rev • VOL 91 • JULY 2011 • www.prv.org Downloaded from http://physrev.physiology.org/ by 10.220.32.246 on June 14, 2017 Torocsik et al. (362) investigated the changes of FXIII-A expression after activation of monocytes through the classical or alternative pathway. Induction of the classical activation pathway by interferon (IFN)-␥ or Mycobacterium bovis BCG downregulated FXIII-A mRNA and protein expression in cultured macrophages, while activation by IL-4 through the alternative pathway exerted the opposite effect. In the latter case, on days 4 and 5, the increase of FXIII-A mRNA level was more than 40-fold, and FXIII-A protein level became elevated 11-fold on day 5. The expression of TG-2 did not change in these conditions. Similar upregulation of FXIII-A mRNA by IL-4, and to a lesser extent by IL-13, was observed by another group, as well (59), while priming monocytes by lipopolysaccharide decreased FXIII-A expression (290). In another study, the effect of IL-4 on the expression of FXIII-A at the mRNA and protein level was highly increased by the addition of dexamethasone (120). Results of in vitro investigations were also supported by the detection of FXIII-A in tumor-associated macrophages that are considered as alternatively activated, and its absence in macrophages of tuberculotic granuloma that go through the classical activation pathway (362). The expression of FXIIIa substrate fibronectin is also augmented by IL-4/IL-13. The parallel upregulation of cFXIII expression and tissue inhibitor of metalloproteinase 3 (TIMP3) could protect the deposited extracellular matrix protein from degradation (59). ligand, interferon inducible protein-10 (IP-10). This mechanism also contributed to the highly increased production of FXIII-A in CRCR3-positive macrophages (388). However, it is not clear how elevated cFXIII in perivascular macrophages is involved in vascular remodeling and if such a mechanism also operates in humans. MULTIPLE FUNCTIONS OF FACTOR XIII The inhibition of angiotensin II generation or intracellular FXIII activity suppressed the appearance of cross-linked AT1 receptors and symptoms of atherosclerosis in apoEdeficient mice. Follow-up studies confirming these exciting findings are awaited. There is not much information on the release or surface expression of monocyte/macrophage cFXIII. Although there is a general agreement on the cytoplasmic localization of FXIII-A in these cells, a few studies indicated that macrophages in cell culture translocate it to the surface (16, 72, 190). Increased surface staining of U937 cells (a human promonocytic tumor line) for FXIII-A was observed after incubation with phorbol myristate acetate, lipopolysaccharide, and IFN-␥ (190). In a single study, FXIII-A was detected in the culture medium of dendritic cells differentiated from peripheral blood monocytes (15). However, in these studies, dying or dead cells were not excluded as the source of cFXIII appearing on the cell surface or in the medium. Although the surface expression or release of cFXIII from macrophages, especially from resident macrophages and its involvement in organization and remodeling of extracellular matrix by the cross-linking matrix proteins is quite plausible, this hypothesis has not been supported by experimental proof. As mentioned in section IIC, there is evidence for cFXIII entering the alternative secretory pathway in human macrophages (75), but its actual secretion has not been proven. In addition to the possible translocation of cFXIII from the cytoplasm to the cell surface, another type of cFXIII translocation has also been revealed in macrophages. The transient appearance of cFXIII in the nuclei was demonstrated by confocal laser scanning microscopy, Western blotting, and immunoelectron microscopy in the early phase of monocyte/macrophage differentiation (7). On day 2 of culturing, 90% of the cells expressed FXIII-A in the nucleus in association with electrodense areas, but this phenomenon was not observed on any other day. cFXIII in the nucleus somehow became activated, and its TG activity was demonstrated by the incorporation of MDC into nuclear proteins. Nuclear localization of TG-2 has also been observed (208, 291, 295), but the function of neither TG has been revealed in the nuclear environment. Follow-up studies on VIII. SUMMARY, CONCLUSIONS, AND PERSPECTIVES pFXIII is a tetrameric zymogen protransglutaminase, which consists of two potentially active A and two inhibitory/ carrier B subunits (FXIII-A2B2). In the plasma it is bound to the ␥=-chain of fibrinogen through FXIII-B subunits. The synthesis of the two subunits occurs in different cells, and they form complex in the plasma. There is little doubt that the major part of FXIII-A is synthesized by cells of bone marrow origin, while FXIII-B is produced and secreted by hepatocytes. However, it is still unclear what is the contribution of megakaryocytes and monocytes/macrophages to FXIII-A present in pFXIII. The secretory pathway by which FXIII-A is released from the cells also remains to be elucidated, although most recently evidence has been provided on its entering into the nonclassical secretory pathway in macrophages. FXIII-B is in excess in the plasma; ⬃50% of it circulates in free, uncomplexed form. The dimer of FXIII-A, but not FXIII-B, is also present in cells (platelets, monocytes, macrophages, chondrocytes, osteoblasts, osteocytes). The three-dimensional structure of FXIII-A2 has been revealed by X-ray crystallography, and four major structural domains (-sandwich domain, central core domain, and two -barrel domains) and AP-FXIII were identified. The three-dimensional structure of FXIII-B is not known; it contains 10 sushi domains each held together by two pairs of disulfide bonds. pFXIII becomes activated in the final phase of coagulation cascade by thrombin and Ca2⫹. Thrombin cleaves off APFXIII from FXIII-A, which, as demonstrated most recently, dissociates from the truncated molecule (FXIII-A=). The newly formed fibrin ⬃100-fold accelerates this process. In the presence of Ca2⫹, FXIII-B leaves FXIII-A=2, and the latter assumes an enzymatically active configuration (FXIIIA*2). In plasma, the activation of FXIII occurs on the surface of fibrin, and FXIIIa remains associated with it, while FXIII-B is released into the serum. The release of AP-FXIII is not a prerequisite of FXIII activation. At very high unphysiological Ca2⫹ concentration (ⱖ100 mM), FXIII-B dissociates from the nontruncated FXIII-A2, which then becomes enzymatically active (FXIII-A°2). The main role of the truncation of FXIII-A is to bring down the Ca2⫹ requirement for the dissociation step into physiological range. In the absence of FXIII-B, cFXIII can be transformed into FXIII-A°2 at low Ca2⫹ concentration. The main role of FXIII-B seems to prevent FXIII-A2 from going through a slow progressive activation in plasmatic environment. Several pieces of evidence suggest that the nonproteolytic pathway is the main form of cFXIII activation in platelets and in monocytes/macrophages. No major conformational change Physiol Rev • VOL 91 • JULY 2011 • www.prv.org 961 Downloaded from http://physrev.physiology.org/ by 10.220.32.246 on June 14, 2017 In a most recent paper it was demonstrated that cFXIII is involved in the regulation of gene expression in alternatively activated human macrophages (363). In microarray experiments, a significant difference was demonstrated in the IL-4-sensitive gene expression profile of cultured macrophages derived from FXIII-deficient and nondeficient individuals. The most prominent differences were related to immune functions and wound response, suggesting that the functional impairment of macrophages at the level of gene expression regulation plays a role in the wound healing defect of FXIII-deficient patients. the involvement of cFXIII in nuclear processes are required to extend and clarify the significance of this intriguing finding. MUSZBEK ET AL. was detected in the structure of FXIII-A2 during transformation into a TG by X-ray crystallography; however, biochemical evidence and the analogy with the activation of TG-2 indicate that in solution the activation process induces considerable structural changes. The downregulation of FXIIIa has been explored only most recently. Polymorphonuclear granulocytes incorporated in the clot become activated and release several proteolytic enzymes into their surroundings. These proteases, primarily elastase, break down FXIIIa within the plasma clot, even in the presence of serine protease plasma inhibitors. The kinetics of FXIIIa inactivation and fibrinolytic effect of granulocyte proteases ensure the formation of stable fibrin clot, but prevent the production of over-cross-linked plasma clot, which would be difficult to eliminate, when it is not needed any more. Further research on this important field would broaden our knowledge on what is happening with FXIIIa in the plasma clot and in the thrombus. The main function of FXIIIa in the plasma is to cross-link fibrin chains and ␣2-PI to fibrin. Fibrin ␥-chains are crosslinked into dimers at a very early stage of clotting, while fibrin ␣-chains form high-molecular-weight polymers at a much slower rate. The cross-linking of ␣2-PI to fibrin ␣-chains is also rather fast, and the heterodimer becomes progressively incorporated into the highly cross-linked ␣-chain polymers. Fibrin cross-linking stabilizes fibrin and makes it more resistant against shear stress. Both fibrin cross-linking and the covalent incorporation of ␣2-PI into 962 The involvement of FXIII in wound healing has been supported by sporadic clinical data from studies with FXIII-Adeficient patients. Its role in wound healing is now well proven by the impaired wound healing of FXIII-A knockout mice. The promotion of wound healing by FXIII is a complex process; it involves its effects on fibroblasts, macrophages, extracellular matrix, and angiogenesis. FXIIIa enhanced the migration of fibroblasts and macrophages, increased the rate of fibroblasts proliferation, and decreased their apoptosis. Cross-linking of extracellular matrix proteins might modulate these direct effects. The FXIIIa-induced biochemical mechanisms in these events have not been enlightened, yet. The downregulation of TSP-1 has been shown to contribute to the effect of FXIIIa on monocytes. In the last couple of years, the proangiogenic effect of FXIII has been explored to a significant extent. Although nonactivated FXIII can bind to endothelial cells, the proangiogenic effect is exerted only by the active form. FXIIIa enhances endothelial cell migration and proliferation and suppresses their apoptosis. The role of FXIII in angiogenesis has also been verified in several animal models. The effect of FXIIIa on endothelial cells involves binding to ␣v3 receptor, the interaction of this receptor with VEGFR-2, VEGFR-2 phosphorylation, phosphorylation and activation of Akt and ERKs, the upregulation of the transcription factors c-Jun and Egr-1, and the downregulation of TSP-1. Further exploration of this mechanism is expected, and the proangiogenic effect of FXIIIa or its inhibition could have a prospect of clinical utilization. Clinical findings and experiments with FXIII-A knockout mice prove, without any doubt, that pFXIII is essential for maintaining pregnancy. It is surprising that, in spite of the importance of these observations, hardly any studies explored the mechanism by which pFXIII prevents spontaneous abortions. As fibrinogen deficiency also leads to early pregnancy loss, the cooperation of the two clotting factors seems essential. The cross-linking of fibrinoid components in the placenta (fibrin, fibronectin) is very likely important Physiol Rev • VOL 91 • JULY 2011 • www.prv.org Downloaded from http://physrev.physiology.org/ by 10.220.32.246 on June 14, 2017 FXIIIa, like other TGs, catalyzes a double displacement acyl transfer reaction, in which the acyl donor peptide-bound glutamine residue forms thioester with the active site Cys314 of the core domain, and ammonia is released. In the second step, an acyl acceptor primary amine is cross-linked to the glutamine residue through isopeptide bond, and the active site cysteine becomes deacylated. If the acyl acceptor amine is provided by a peptide-bound lysine residue, two peptide chains become covalently cross-linked. The glutamine substrate specificity of FXIIIa is rather restrictive, only less than two dozen proteins have been identified as FXIIIa substrates. However, the amino acid sequences around the substrate glutamine site show considerable variation, and no consensus sequence could be derived from the known primary structures. The structural requirement of the interaction with FXIIIa was studied only in the case of two peptide substrates. One can conclude from the NMR studies and kinetic experiments with chemical mutant peptides that, in addition to the active site, binding of the substrate peptide to a second interacting site, involving hydrophobic interaction, on the surface of FXIII-A*2 is also a prerequisite of the enzymatic action. Experiments with a series of individual glutamine donor peptide substrate are to be carried out to clarify the structural elements required for effective enzyme-substrate interaction. fibrin protect fibrin against fibrinolysis. The cross-linking of ␣2-PI to the fibrin network seems to play the dominant role in preventing newly formed fibrin from the prompt elimination by plasmin, while fibrin ␣-chain cross-linking could contribute to the increased resistance of mature thrombi to fibrinolysis. pFXIII is essential for maintaining hemostasis; its deficiency results in very severe bleeding diathesis. pFXIII is now considered as a key player in the process of fibrinolysis, which exerts its effect through the two mechanisms discussed above. In addition to ␣2-PI, FXIIIa also cross-links other components of fibrinolysis, like PAI-2, TAFI, and plasminogen, to fibrin or fibronectin; however, the significance of these cross-linking reactions remains to be explored. Further intensive research is expected on this field in the near future. MULTIPLE FUNCTIONS OF FACTOR XIII for the sealing/anchoring effect and barrier function of the Nitabuch’s layer. The proangiogenic effect of FXIIIa might contribute to normal placenta development. Elaboration of these issues could lead to highly important pieces of information and to better understanding of the mechanisms required for maintaining pregnancy. The active form of FXIII-A2 is also an intracellular enzyme. Although its presence in platelets is known for more than a half century and their existence in monocytes/macrophages for a quarter of century, its intracellular function started to be addressed only most recently. During platelet activation part of its cFXIII becomes activated by the nonproteolytic mechanism in the later phase of the activation process. During this process, part of FXIII becomes translocated to the cytoskeleton and cross-links platelet proteins, particularly cytoskeletal proteins. Activated FXIII is involved in certain phases of platelet spreading. Information on the involvement of cFXIII in clot retraction is controversial. It is not clear if partial contribution of cFXIII to the clot retraction in mice could be extrapolated to humans. Intriguing novel findings were reported on monocyte/macrophage cFXIII in the last couple of years. The expression of cFXIII in monocytes activated by the alternative pathway was highly upregulated, while activation through the classical pathway resulted in the opposite effect. The results obtained with cultured monocytes were also confirmed with in situ activated tissue macrophages. The transient appearance of cFXIII in the nucleus on day 2 of monocyte culturing is an intriguing finding that warrants further investigation. Differentiation of monocytes into dendritic cells resulted in very high intracellular level of cFXIII. Impaired receptor-mediated phagocytosis of monocytes from FXIII-deficient patients suggests the involvement of cFXIII in the biochemical mechanism of this process. cFXIII has ACKNOWLEDGMENTS Address for reprint requests and other correspondence: L. Muszbek, Clinical Research Center, Univ. of Debrecen, Medical and Health Science Center, 98 Nagyerdei krt., Debrecen 4032, Hungary (e-mail: [email protected]). GRANTS This review was made possible through research funding from the Hungarian National Research Fund (OTKANKTH CNK 80776), the Hungarian Academy of Sciences (TKI 227), and the Hungarian Ministry of Health (ETT). The work was also supported by the TÁMOP 4.2.1./B-09/ 1/KONV-2010-0007 project. The project is implemented through the New Hungary Development Plan, cofinanced by the European Social Fund. DISCLOSURES No conflicts of interest, financial or otherwise, are declared by the authors. REFERENCES 1. AbdAlla S, Lother H, Langer A, el Faramawy Y, Quitterer U. Factor XIIIA transglutaminase crosslinks AT1 receptor dimers of monocytes at the onset of atherosclerosis. Cell 119:343–354, 2004. 2. Achyuthan KE, Mary A, Greenberg CS. The binding sites on fibrin(ogen) for guinea pig liver transglutaminase are similar to those of blood coagulation factor XIII. Characterization of the binding of liver transglutaminase to fibrin. J Biol Chem 263:14296 – 14301, 1988. 3. Adamson R. Role of macrophages in normal wound healing: an overview. J Wound Care 18:349 –351, 2009. 4. Adany R. Intracellular factor XIII: cellular distribution of factor XIII subunit a in humans. Semin Thromb Hemost 22:399 – 408, 1996. 5. Adany R, Antal M. Three different cell types can synthesize factor XIII subunit A in the human liver. Thromb Haemost 76:74 –79, 1996. 6. Adany R, Bardos H. Factor XIII subunit A as an intracellular transglutaminase. Cell Mol Life Sci 60:1049 –1060, 2003. 7. Adany R, Bardos H, Antal M, Modis L, Sarvary A, Szucs S, Balogh I. Factor XIII of blood coagulation as a nuclear crosslinking enzyme. Thromb Haemost 85:845– 851, 2001. 8. Adany R, Belkin A, Vasilevskaya T, Muszbek L. Identification of blood coagulation factor XIII in human peritoneal macrophages. Eur J Cell Biol 38:171–173, 1985. 9. Adany R, Glukhova MA, Kabakov AY, Muszbek L. Characterisation of connective tissue cells containing factor XIII subunit a. J Clin Pathol 41:49 –56, 1988. Physiol Rev • VOL 91 • JULY 2011 • www.prv.org 963 Downloaded from http://physrev.physiology.org/ by 10.220.32.246 on June 14, 2017 In addition to its role in hemostasis, wound healing/angiogenesis, and maintaining pregnancy, studies in the last decade drew attention to other less evident, less expected, and less explored effects of FXIII. A few reports indicated that FXIIIa reduces vascular permeability, but the mechanism is unknown. Experiments with FXIII-A-deficient mice suggested a protective effect of FXIII against spontaneous cardiac rupture and against cardiac rupture following ligationinduced myocardial infarction. Studies on FXIII in cartilage and bone extended FXIII research to a new field. It is clear that both chondrocytes and osteoblasts express FXIII-A, and its expression is related to the stage of differentiation and the location of these cells. By an unknown mechanism, FXIII becomes externalized and activated. Experiments with cultured cells suggest that it is involved in extracellular matrix assembly, stabilization, and mineralization. Although some of the reported findings need confirmation, research on these novel functions of FXIII is highly promising. also been implicated in the locomotion of dendritic cells. The nonproteolytic activation of cFXIII in monocytes of hypertensive patients and the cross-linking of AT1 resulted in enhanced signaling and adhesion to endothelial cells. The role of this mechanism in atherosclerosis was suggested by experiments with apoE-deficient mice. There is no doubt that further studies on the intracellular function of cFXIII would lead to exciting new findings and provide new data on the role of this intracellular TG. MUSZBEK ET AL. 10. Adany R, Kiss A, Muszbek L. Factor XIII: a marker of mono- and megakaryocytopoiesis. Br J Haematol 67:167–172, 1987. 11. Adany R, Muszbek L. Immunohistochemical detection of factor XIII subunit a in histiocytes of human uterus. Histochemistry 91:169 –174, 1989. 31. Balogh I, Szoke G, Karpati L, Wartiovaara U, Katona E, Komaromi I, Haramura G, Pfliegler G, Mikkola H, Muszbek L. Val34Leu polymorphism of plasma factor XIII: biochemistry and epidemiology in familial thrombophilia. Blood 96:2479 –2486, 2000. 32. Bangert K, Johnsen AH, Christensen U, Thorsen S. Different N-terminal forms of alpha 2-plasmin inhibitor in human plasma. Biochem J 291:623– 625, 1993. 12. Aeschlimann D, Kaupp O, Paulsson M. Transglutaminase-catalyzed matrix crosslinking in differentiating cartilage: identification of osteonectin as a major glutaminyl substrate. J Cell Biol 129:881– 892, 1995. 33. Barkan G. Zur Frage der Reversibilitat der Fibringerinnung II. Biochem Ztschr 139: 291–301, 1923. 13. Aeschlimann D, Mosher D, Paulsson M. Tissue transglutaminase and factor XIII in cartilage and bone remodeling. Semin Thromb Hemost 22:437– 443, 1996. 34. Barry EL, Mosher DF. Binding and degradation of blood coagulation factor XIII by cultured fibroblasts. J Biol Chem 265:9302–9307, 1990. 14. Ahvazi B, Kim HC, Kee SH, Nemes Z, Steinert PM. Three-dimensional structure of the human transglutaminase 3 enzyme: binding of calcium ions changes structure for activation. EMBO J 21:2055–2067, 2002. 35. Barry EL, Mosher DF. Factor XIII cross-linking of fibronectin at cellular matrix assembly sites. J Biol Chem 263:10464 –10469, 1988. 16. Akimov SS, Belkin AM. Cell surface tissue transglutaminase is involved in adhesion and migration of monocytic cells on fibronectin. Blood 98:1567–1576, 2001. 17. Al-Jallad HF, Nakano Y, Chen JL, McMillan E, Lefebvre C, Kaartinen MT. Transglutaminase activity regulates osteoblast differentiation and matrix mineralization in MC3T3–E1 osteoblast cultures. Matrix Biol 25:135–148, 2006. 18. Alkjaersig N, Fletcher AP, Lewis M, Ittyerah R. Reduction of coagulation factor XIII concentration in patients with myocardial infarction, cerebral infarction, and other thromboembolic disorders. Thromb Haemost 38:863– 873, 1977. 19. Ambrus A, Banyai I, Weiss MS, Hilgenfeld R, Keresztessy Z, Muszbek L, Fesus L. Calcium binding of transglutaminases: a 43Ca NMR study combined with surface polarity analysis. J Biomol Struct Dyn 19:59 –74, 2001. 20. Andersen MD, Kjalke M, Bang S, Lautrup-Larsen I, Becker P, Andersen AS, Olsen OH, Stennicke HR. Coagulation factor XIII variants with altered thrombin activation rates. Biol Chem 390:1279 –1283, 2009. 21. Ando Y, Imamura S, Yamagata Y, Kitahara A, Saji H, Murachi T, Kannagi R. Platelet factor XIII is activated by calpain. Biochem Biophys Res Commun 144:484 – 490, 1987. 22. Anwar R, Gallivan L, Edmonds SD, Markham AF. Genotype/phenotype correlations for coagulation factor XIII: specific normal polymorphisms are associated with high or low factor XIII specific activity. Blood 93:897–905, 1999. 23. Ariens RA, Philippou H, Nagaswami C, Weisel JW, Lane DA, Grant PJ. The factor XIII V34L polymorphism accelerates thrombin activation of factor XIII and affects crosslinked fibrin structure. Blood 96:988 –995, 2000. 24. Asahina T, Kobayashi T, Okada Y, Goto J, Terao T. Maternal blood coagulation factor XIII is associated with the development of cytotrophoblastic shell. Placenta 21:388 – 393, 2000. 25. Asahina T, Kobayashi T, Okada Y, Itoh M, Yamashita M, Inamato Y, Terao T. Studies on the role of adhesive proteins in maintaining pregnancy. Horm Res 50 Suppl 2:37– 45, 1998. 26. Asahina T, Kobayashi T, Takeuchi K, Kanayama N. Congenital blood coagulation factor XIII deficiency and successful deliveries: a review of the literature. Obstet Gynecol Surv 62:255–260, 2007. 36. Beck E, Duckert F, Ernst M. The influence of fibrin stabilizing factor on the growth of fibroblasts in vitro and wound healing. Thromb Diath Haemorrh 6:485– 491, 1961. 37. Bendixen E, Borth W, Harpel PC. Transglutaminases catalyze cross-linking of plasminogen to fibronectin and human endothelial cells. J Biol Chem 268:21962–21967, 1993. 38. Bendixen E, Harpel PC, Sottrup-Jensen L. Location of the major epsilon-(gammaglutamyl)lysyl cross-linking site in transglutaminase-modified human plasminogen. J Biol Chem 270:17929 –17933, 1995. 39. Beninati S, Nicolini L, Jakus J, Passeggio A, Abbruzzese A. Identification of a substrate site for transglutaminases on the human protein synthesis initiation factor 5A. Biochem J 305:725–728, 1995. 40. Bereczky Z, Balogh E, Katona E, Czuriga I, Edes I, Muszbek L. Elevated factor XIII level and the risk of myocardial infarction in women. Haematologica 92:287–288, 2007. 41. Biland L, Duckert F. Coagulation factors of the newborn and his mother. Thromb Diath Haemorrh 29:644 – 651, 1973. 42. Bishop PD, Teller DC, Smith RA, Lasser GW, Gilbert T, Seale RL. Expression, purification, and characterization of human factor XIII in Saccharomyces cerevisiae. Biochemistry 29:1861–1869, 1990. 43. Board PG. Genetic polymorphism of the B subunit of human coagulation factor XIII. Am J Hum Genet 32:348 –353, 1980. 44. Board PG, Losowsky MS, Miloszewski KJ. Factor XIII: inherited and acquired deficiency. Blood Rev 7:229 –242, 1993. 45. Board PG, Pierce K, Coggan M. Expression of functional coagulation factor XIII in Escherichia coli. Thromb Haemost 63:235–240, 1990. 46. Board PG, Webb GC, McKee J, Ichinose A. Localization of the coagulation factor XIII A subunit gene (F13A) to chromosome bands 6p24 —-p25. Cytogenet Cell Genet 48:25–27, 1988. 47. Boda Z, Pfliegler G, Muszbek L, Toth A, Adany R, Harsfalvi J, Papp Z, Tornai I, Rak K. Congenital factor XIII deficiency with multiple benign breast tumours and successful pregnancy with substitutive therapy. A case report. Haemostasis 19:348 –352, 1989. 48. Bohn H, Schwick HG. Isolation and characterization of a fibrin-stabilizing factor from human placenta. Arzneimittelforschung 21:1432–1439, 1971. 49. Borth W, Chang V, Bishop P, Harpel PC. Lipoprotein (a) is a substrate for factor XIIIa and tissue transglutaminase. J Biol Chem 266:18149 –18153, 1991. 27. Asijee GM, Muszbek L, Kappelmayer J, Polgar J, Horvath A, Sturk A. Platelet vinculin: a substrate of activated factor XIII. Biochim Biophys Acta 954:303–308, 1988. 50. Bottenus RE, Ichinose A, Davie EW. Nucleotide sequence of the gene for the b subunit of human factor XIII. Biochemistry 29:11195–11209, 1990. 28. Bagoly Z, Fazakas F, Komaromi I, Haramura G, Toth E, Muszbek L. Cleavage of factor XIII by human neutrophil elastase results in a novel active truncated form of factor XIII A subunit. Thromb Haemost 99:668 – 674, 2008. 51. Broekman MJ, Handin RI, Cohen P. Distribution of fibrinogen, platelet factors 4 and XIII in subcellular fractions of human platelets. Br J Haematol 31:51–55, 1975. 29. Bagoly Z, Haramura G, Muszbek L. Down-regulation of activated factor XIII by polymorphonuclear granulocyte proteases within fibrin clot. Thromb Haemost 98:359 – 367, 2007. 30. Bakker EN, Pistea A, Spaan JA, Rolf T, de Vries CJ, van Rooijen N, Candi E, VanBavel E. Flow-dependent remodeling of small arteries in mice deficient for tissue-type transglutaminase: possible compensation by macrophage-derived factor XIII. Circ Res 99:86 –92, 2006. 964 52. Broker M, Bauml O. New expression vectors for the fission yeast Schizosaccharomyces pombe. FEBS Lett 248:105–110, 1989. 53. Brown LF, Lanir N, McDonagh J, Tognazzi K, Dvorak AM, Dvorak HF. Fibroblast migration in fibrin gel matrices. Am J Pathol 142:273–283, 1993. 54. Bruhn HD, Zurborn KH. Influences of clotting factors (thrombin, factor XIII) and of fibronectin on the growth of tumor cells and leukemic cells in vitro. Blut 46:85– 88, 1983. Physiol Rev • VOL 91 • JULY 2011 • www.prv.org Downloaded from http://physrev.physiology.org/ by 10.220.32.246 on June 14, 2017 15. Akagi A, Tajima S, Ishibashi A, Matsubara Y, Takehana M, Kobayashi S, Yamaguchi N. Type XVI collagen is expressed in factor XIIIa⫹ monocyte-derived dermal dendrocytes and constitutes a potential substrate for factor XIIIa. J Invest Dermatol 118:267– 274, 2002. MULTIPLE FUNCTIONS OF FACTOR XIII 55. Buluk K. An unknown action of blood platelets; preliminary communication. Pol Tyg Lek 10:191, 1955. 56. Carpenter SL, Mathew P. Alpha2-antiplasmin and its deficiency: fibrinolysis out of balance. Haemophilia 14:1250 –1254, 2008. 57. Carr ME ,Jr, Carr SL, Tildon T, Fisher LM, Martin EJ. Batroxobin-induced clots exhibit delayed and reduced platelet contractile force in some patients with clotting factor deficiencies. J Thromb Haemost 1:243–249, 2003. 58. Carrell NA, Erickson HP, McDonagh J. Electron microscopy and hydrodynamic properties of factor XIII subunits. J Biol Chem 264:551–556, 1989. 59. Chaitidis P, O’Donnell V, Kuban RJ, Bermudez-Fajardo A, Ungethuem U, Kuhn H. Gene expression alterations of human peripheral blood monocytes induced by medium-term treatment with the TH2-cytokines interleukin-4 and -13. Cytokine 30: 366 –377, 2005. 60. Chen R, Doolittle RF. Cross-linking sites in human and bovine fibrin. Biochemistry 10:4487– 4491, 1971. 62. Claes L, Burri C, Gerngross H, Mutschler W. Bone healing stimulated by plasma factor XIII. Osteotomy experiments in sheep. Acta Orthop Scand 56:57– 62, 1985. 63. Cleary DB, Doiphode PG, Sabo TM, Maurer MC. A non-reactive glutamine residue of alpha2-antiplasmin promotes interactions with the factor XIII active site region. J Thromb Haemost 7:1947–1949, 2009. 64. Cleary DB, Maurer MC. Characterizing the specificity of activated Factor XIII for glutamine-containing substrate peptides. Biochim Biophys Acta 1764:1207–1217, 2006. 65. Cohen I, Blankenberg TA, Borden D, Kahn DR, Veis A. Factor XIIIa-catalyzed crosslinking of platelet and muscle actin. Regulation by nucleotides. Biochim Biophys Acta 628:365–375, 1980. 78. Credo RB, Curtis CG, Lorand L. Alpha-chain domain of fibrinogen controls generation of fibrinoligase (coagulation factor XIIIa). Calcium ion regulatory aspects. Biochemistry 20:3770 –3778, 1981. 79. Credo RB, Curtis CG, Lorand L. Ca2⫹-related regulatory function of fibrinogen. Proc Natl Acad Sci USA 75:4234 – 4237, 1978. 80. Csosz E, Mesko B, Fesus L. Transdab wiki: the interactive transglutaminase substrate database on web 2.0 surface. Amino Acids 36:615– 617, 2009. 81. Curtis CG, Brown KL, Credo RB, Domanik RA, Gray A, Stenberg P, Lorand L. Calcium-dependent unmasking of active center cysteine during activation of fibrin stabilizing factor. Biochemistry 13 3774 –3780, 1974. 82. Czabanka M, Martin E, Walther A. Role of antithrombin and factor XIII in leukocyteindependent plasma extravasation during endotoxemia: an intravital-microscopic study in the rat. J Surg Res 136:219 –226, 2006. 83. D’Argenio G, Grossman A, Cosenza V, Valle ND, Mazzacca G, Bishop PD. Recombinant factor XIII improves established experimental colitis in rats. Dig Dis Sci 45:987– 997, 2000. 84. Dallabrida SM, Falls LA, Farrell DH. Factor XIIIa supports microvascular endothelial cell adhesion and inhibits capillary tube formation in fibrin. Blood 95:2586 –2592, 2000. 85. Dardik R, Krapp T, Rosenthal E, Loscalzo J, Inbal A. Effect of FXIII on monocyte and fibroblast function. Cell Physiol Biochem 19:113–120, 2007. 86. Dardik R, Leor J, Skutelsky E, Castel D, Holbova R, Schiby G, Shaish A, Dickneite G, Loscalzo J, Inbal A. Evaluation of the pro-angiogenic effect of factor XIII in heterotopic mouse heart allografts and FXIII-deficient mice. Thromb Haemost 95:546 –550, 2006. 66. Cohen I, Gerrard JM, White JG. Ultrastructure of clots during isometric contraction. J Cell Biol 93:775–787, 1982. 87. Dardik R, Livnat T, Seligsohn U. Variable effects of alpha v suppression on VEGFR-2 expression in endothelial cells of different vascular beds. Thromb Haemost 102:975– 982, 2009. 67. Cohen I, Glaser T, Veis A, Bruner-Lorand J. Ca2⫹-dependent cross-linking processes in human platelets. Biochim Biophys Acta 676:137–147, 1981. 88. Dardik R, Loscalzo J, Eskaraev R, Inbal A. Molecular mechanisms underlying the proangiogenic effect of factor XIII. Arterioscler Thromb Vasc Biol 25:526 –532, 2005. 68. Cohen I, Lim CT, Kahn DR, Glaser T, Gerrard JM, White JG. Disulfide-linked and transglutaminase-catalyzed protein assemblies in platelets. Blood 66:143–151, 1985. 69. Cohen I, Young-Bandala L, Blankenberg TA, Siefring GE ,Jr, Bruner-Lorand J. Fibrinoligase-catalyzed cross-linking of myosin from platelet and skeletal muscle. Arch Biochem Biophys 192:100 –111, 1979. 70. Colman RW, Clowes AW, George JN, Goldhaber SZ, Marder VJ. Overview of hemostasis. In: Hemostasis and Thrombosis, edited by Colman RW, Marder VJ, Clowes AW, George JN, Goldhaber SZ. Philadelphia, PA:Lippincott Williams and Wilkins, 2006, p. 3–100 –20. 71. Colman RW, Clowes AW, George JN, Goldhaber SZ, Marder VJ. Thrombin-activatable fibrinolysis inhibitor aka procarboxypeptidase U. In: Hemostasis and Thrombosis, edited by Colman RW, Marder VJ, Clowes AW, George JN, Goldhaber SZ. Philadelphia, PA:Lippincott Williams and Wilkins, 2006, p. 381–100 – 407. 72. Conkling PR, Achyuthan KE, Greenberg CS, Newcomb TF, Weinberg JB. Human mononuclear phagocyte transglutaminase activity cross-links fibrin. Thromb Res 55: 57– 68, 1989. 73. Coopland A, Alkjaersig N, Fletcher AP. Reduction in plasma factor 13 (fibrin stabilizing factor) concentration during pregnancy. J Lab Clin Med 73:144 –153, 1969. 74. Corbett SA, Lee L, Wilson CL, Schwarzbauer JE. Covalent cross-linking of fibronectin to fibrin is required for maximal cell adhesion to a fibronectin-fibrin matrix. J Biol Chem 272:24999 –25005, 1997. 89. Dardik R, Shenkman B, Tamarin I, Eskaraev R, Harsfalvi J, Varon D, Inbal A. Factor XIII mediates adhesion of platelets to endothelial cells through alpha(v)beta(3) and glycoprotein IIb/IIIa integrins. Thromb Res 105:317–323, 2002. 90. Dardik R, Solomon A, Loscalzo J, Eskaraev R, Bialik A, Goldberg I, Schiby G, Inbal A. Novel proangiogenic effect of factor XIII associated with suppression of thrombospondin 1 expression. Arterioscler Thromb Vasc Biol 23:1472–1477, 2003. 91. De Willige SU, Standeven KF, Philippou H, Ariens RA. The pleiotropic role of the fibrinogen gamma= chain in hemostasis. Blood 114:3994 – 4001, 2009. 92. Donga A, Kendrickb B, Kreilga˚rdb L, Matsuurab J, Manningb MC, Carpenterb JF. Spectroscopic study of secondary structure and thermal denaturation of recombinant human factor XIII in aqueous solution. Arch Biochem Biophys 347:213–220, 1997. 93. Doolittle RF. Searching for differences between fibrinogen and fibrin that affect the initiation of fibrinolysis. Cardiovasc Hematol Agents Med Chem 6:181–189, 2008. 94. Duckert F, Jung E, Shmerling DH. A hitherto undescribed congenital haemorrhagic diathesis probably due to fibrin stabilizing factor deficiency. Thromb Diath Haemorrh 5:179 –186, 1960. 95. Facchiano A, Facchiano F. Transglutaminases and their substrates in biology and human diseases: 50 years of growing. Amino Acids 36:599 – 614, 2009. 96. Fairweather D, Cihakova D. Alternatively activated macrophages in infection and autoimmunity. J Autoimmun 33:222–230, 2009. 75. Cordell PA, Kile BT, Standeven KF, Josefsson EC, Pease RJ, Grant PJ. Association of coagulation factor XIII-A with Golgi proteins within monocyte-macrophages: implications for subcellular trafficking and secretion. Blood 115:2674 –2681, 2010. 97. Falls LA, Farrell DH. Resistance of gammaA/gamma= fibrin clots to fibrinolysis. J Biol Chem 272:14251–14256, 1997. 76. Cornwell MM, Juliano RL, Davies PJ. Inhibition of the adhesion of Chinese hamster ovary cells by the naphthylsulfonamides dansylcadaverine and N-(6-aminohexyl)-5chloro-1-naphthylenesulfonamide (W7). Biochim Biophys Acta 762:414 – 419, 1983. 98. Feng Q, Liu Y, Liu K, Byrne S, Liu G, Wang X, Li Z, Ockleford CD. Expression of urokinase, plasminogen activator inhibitors and urokinase receptor in pregnant rhesus monkey uterus during early placentation. Placenta 21:184 –193, 2000. Physiol Rev • VOL 91 • JULY 2011 • www.prv.org 965 Downloaded from http://physrev.physiology.org/ by 10.220.32.246 on June 14, 2017 61. Chung SI. Comparative studies on tissue transglutaminase and factor XIII. Ann NY Acad Sci 202:240 –255, 1972. 77. Cottrell BA, Strong DD, Watt KW, Doolittle RF. Amino acid sequence studies on the alpha chain of human fibrinogen. Exact location of cross-linking acceptor sites. Biochemistry 18:5405–5410, 1979. MUSZBEK ET AL. 99. Fesus L, Metsis ML, Muszbek L, Koteliansky VE. Transglutaminase-sensitive glutamine residues of human plasma fibronectin revealed by studying its proteolytic fragments. Eur J Biochem 154:371–374, 1986. 121. Greenberg CS, Achyuthan KE, Fenton JW ,2nd. Factor XIIIa formation promoted by complexing of alpha-thrombin, fibrin, and plasma factor XIII. Blood 69:867– 871, 1987. 100. Fisher S, Rikover M, Naor S. Factor 13 deficiency with severe hemorrhagic diathesis. Blood 28:34 –39, 1966. 122. Greenberg CS, Achyuthan KE, Rajagopalan S, Pizzo SV. Characterization of the fibrin polymer structure that accelerates thrombin cleavage of plasma factor XIII. Arch Biochem Biophys 262:142–148, 1988. 101. Floridon C, Nielsen O, Holund B, Sunde L, Westergaard JG, Thomsen SG, Teisner B. Localization and significance of urokinase plasminogen activator and its receptor in placental tissue from intrauterine, ectopic and molar pregnancies. Placenta 20:711–721, 1999. 102. Fox BA, Yee VC, Pedersen LC, Le Trong I, Bishop PD, Stenkamp RE, Teller DC. Identification of the calcium binding site and a novel ytterbium site in blood coagulation factor XIII by x-ray crystallography. J Biol Chem 274:4917– 4923, 1999. 103. Francis CW, Marder VJ. Increased resistance to plasmic degradation of fibrin with highly crosslinked alpha-polymer chains formed at high factor XIII concentrations. Blood 71:1361–1365, 1988. 105. Francis RT, McDonagh J, Mann KG. Factor V is a substrate for the transamidase factor XIIIa. J Biol Chem 261:9787–9792, 1986. 106. Gaffney PJ, Whitaker AN. Fibrin crosslinks and lysis rates. Thromb Res 14:85–94, 1979. 107. Gallivan L, Markham AF, Anwar R. The Leu564 factor XIIIA variant results in significantly lower plasma factor XIII levels than the Pro564 variant. Thromb Haemost 82: 1368 –1370, 1999. 108. Gao J, Hooker BS, Anderson DB. Expression of functional human coagulation factor XIII A-domain in plant cell suspensions and whole plants. Protein Expr Purif 37:89 –96, 2004. 109. Gascuel O. BIONJ: an improved version of the NJ algorithm based on a simple model of sequence data. Mol Biol Evol 14:685– 695, 1997. 110. Gemmati D, Federici F, Campo G, Tognazzo S, Serino ML, De Mattei M, Valgimigli M, Malagutti P, Guardigli G, Ferraresi P, Bernardi F, Ferrari R, Scapoli GL, Catozzi L. Factor XIIIA-V34L and factor XIIIB-H95R gene variants: effects on survival in myocardial infarction patients. Mol Med 13:112–120, 2007. 111. Gemmati D, Federici F, Catozzi L, Gianesini S, Tacconi G, Scapoli GL, Zamboni P. DNA-array of gene variants in venous leg ulcers: detection of prognostic indicators. J Vasc Surg 50:1444 –1451, 2009. 112. Gemmati D, Tognazzo S, Catozzi L, Federici F, De Palma M, Gianesini S, Scapoli GL, De Mattei M, Liboni A, Zamboni P. Influence of gene polymorphisms in ulcer healing process after superficial venous surgery. J Vasc Surg 44:554 –562, 2006. 113. Gemmati D, Tognazzo S, Serino ML, Fogato L, Carandina S, De Palma M, Izzo M, De Mattei M, Ongaro A, Scapoli GL, Caruso A, Liboni A, Zamboni P. Factor XIII V34L polymorphism modulates the risk of chronic venous leg ulcer progression and extension. Wound Repair Regen 12:512–517, 2004. 114. Gierhake FW, Papastavrou N, Zimmermann K, Bohn H, Schwick HG. Prophylaxis of post-operative disturbances of wound healing with factor XIII substitution (author’s transl). Dtsch Med Wochenschr 99:1004 –1009, 1974. 115. Gierhake FW, Volkmann W, Becker W, Schwarz H, Schwick HG. Factor XIII concentration and wound healing. Dtsch Med Wochenschr 95:1472–1475, 1970. 116. Girolami A, Cappellato MG, Lazzaro AR, Boscaro M. Type I and type II disease in congenital factor XIII deficiency. A further demonstration of the correctness of the classification. Blut 53:411– 413, 1986. 124. Greenberg CS, Sane DC, Lai TS. Factor XIII and fibrin stabilization. In: Hemostasis and Thrombosis, edited by Colman RW, Clowes AW, Goldhaber SZ, Marder VJ, George JN. Philadelphia, PA:Lippincott Williams and Wilkins, 2006, p. 317–466 –334. 125. Greenberg CS, Shuman MA. The zymogen forms of blood coagulation factor XIII bind specifically to fibrinogen. J Biol Chem 257:6096 – 6101, 1982. 126. Grinnell F, Feld M, Minter D. Fibroblast adhesion to fibrinogen and fibrin substrata: requirement for cold-insoluble globulin (plasma fibronectin). Cell 19:517–525, 1980. 127. Grundmann U, Amann E, Zettlmeissl G, Kupper HA. Characterization of cDNA coding for human factor XIIIa. Proc Natl Acad Sci USA 83:8024 – 8028, 1986. 128. Harsfalvi J, Fesus L, Tarcsa E, Laczko J, Loewy AG. The presence of a covalently cross-linked matrix in human platelets. Biochim Biophys Acta 1073:268 –274, 1991. 129. Hayano Y, Imai N, Karasawa T. Studies on the physiological changes of blood coagulation factor XIII during pregnancy and their significance. Nippon Sanka Fujinka Gakkai Zasshi 34:469 – 477, 1982. 130. Henriksson P, Becker S, Lynch G, McDonagh J. Identification of intracellular factor XIII in human monocytes and macrophages. J Clin Invest 76:528 –534, 1985. 131. Herouy Y, Hellstern MO, Vanscheidt W, Schopf E, Norgauer J. Factor XIII-mediated inhibition of fibrinolysis and venous leg ulcers. Lancet 355:1970 –1971, 2000. 132. Hevessy Z, Haramura G, Boda Z, Udvardy M, Muszbek L. Promotion of the crosslinking of fibrin and alpha 2-antiplasmin by platelets. Thromb Haemost 75:161–167, 1996. 133. Higaki S, Nakano K, Onaka S, Amano A, Tanioka Y, Harada K, Hashimoto S, Sakaida I, Okita K. Clinical significance of measuring blood coagulation factor XIIIA regularly and continuously in patients with Crohn’s disease. J Gastroenterol Hepatol 21:1407– 1411, 2006. 134. Hildenbrand T, Idzko M, Panther E, Norgauer J, Herouy Y. Treatment of nonhealing leg ulcers with fibrin-stabilizing factor XIII: a case report. Dermatol Surg 28:1098 – 1099, 2002. 135. Hirahara K, Shinbo K, Takahashi M, Matsuishi T. Suppressive effect of human blood coagulation factor XIII on the vascular permeability induced by anti-guinea pig endothelial cell antiserum in guinea pigs. Thromb Res 71:139 –148, 1993. 136. Holmes WE, Lijnen HR, Collen D. Characterization of recombinant human alpha 2-antiplasmin and of mutants obtained by site-directed mutagenesis of the reactive site. Biochemistry 26:5133–5140, 1987. 137. Hornyak TJ, Shafer JA. Interactions of factor XIII with fibrin as substrate and cofactor. Biochemistry 31:423– 429, 1992. 138. Hornyak TJ, Shafer JA. Role of calcium ion in the generation of factor XIII activity. Biochemistry 30:6175– 6182, 1991. 139. Hsieh L, Nugent D. Factor XIII deficiency. Haemophilia 14:1190 –1200, 2008. 117. Gladner JA, Nossal R. Effects of crosslinking on the rigidity and proteolytic susceptibility of human fibrin clots. Thromb Res 30:273–288, 1983. 140. Huh MM, Schick BP, Schick PK, Colman RW. Covalent crosslinking of human coagulation factor V by activated factor XIII from guinea pig megakaryocytes and human plasma. Blood 71:1693–1702, 1988. 118. Glaros T, Larsen M, Li L. Macrophages and fibroblasts during inflammation, tissue damage and organ injury. Front Biosci 14:3988 –3993, 2009. 141. Huson DH, Bryant D. Application of phylogenetic networks in evolutionary studies. Mol Biol Evol 23:254 –267, 2006. 119. Glynn LE (Editor) . Tissue Repair and Regeneration. New York:Raven, 1985. 142. Ichinose A, Asahina T, Kobayashi T. Congenital blood coagulation factor XIII deficiency and perinatal management. Curr Drug Targets 6:541–549, 2005. 120. Gratchev A, Kzhyshkowska J, Utikal J, Goerdt S. Interleukin-4 and dexamethasone counterregulate extracellular matrix remodelling and phagocytosis in type-2 macrophages. Scand J Immunol 61:10 –17, 2005. 966 143. Ichinose A, Bottenus RE, Davie EW. Structure of transglutaminases. J Biol Chem 265: 13411–13414, 1990. Physiol Rev • VOL 91 • JULY 2011 • www.prv.org Downloaded from http://physrev.physiology.org/ by 10.220.32.246 on June 14, 2017 104. Francis CW, Marder VJ. Rapid formation of large molecular weight alpha-polymers in cross-linked fibrin induced by high factor XIII concentrations. Role of platelet factor XIII. J Clin Invest 80:1459 –1465, 1987. 123. Greenberg CS, Miraglia CC. The effect of fibrin polymers on thrombin-catalyzed plasma factor XIIIa formation. Blood 66:466 – 469, 1985. MULTIPLE FUNCTIONS OF FACTOR XIII 144. Ichinose A, Davie EW. Characterization of the gene for the a subunit of human factor XIII (plasma transglutaminase), a blood coagulation factor. Proc Natl Acad Sci USA 85:5829 –5833, 1988. 165. Johnson K, Hashimoto S, Lotz M, Pritzker K, Terkeltaub R. Interleukin-1 induces pro-mineralizing activity of cartilage tissue transglutaminase and factor XIIIa. Am J Pathol 159:149 –163, 2001. 145. Ichinose A, Hendrickson LE, Fujikawa K, Davie EW. Amino acid sequence of the a subunit of human factor XIII. Biochemistry 25:6900 – 6906, 1986. 166. Johnson KA, Rose DM, Terkeltaub RA. Factor XIIIA mobilizes transglutaminase 2 to induce chondrocyte hypertrophic differentiation. J Cell Sci 121:2256 –2264, 2008. 146. Ichinose A, McMullen BA, Fujikawa K, Davie EW. Amino acid sequence of the b subunit of human factor XIII, a protein composed of ten repetitive segments. Biochemistry 25:4633– 4638, 1986. 167. Joist JH, Niewiarowski S. Retention of platelet fibrin stabilizing factor during the platelet release reaction and clot retraction. Thromb Diath Haemorrh 29:679 – 683, 1973. 147. Iismaa SE, Holman S, Wouters MA, Lorand L, Graham RM, Husain A. Evolutionary specialization of a tryptophan indole group for transition-state stabilization by eukaryotic transglutaminases. Proc Natl Acad Sci USA 100:12636 –12641, 2003. 168. Kaartinen MT, El-Maadawy S, Rasanen NH, McKee MD. Tissue transglutaminase and its substrates in bone. J Bone Miner Res 17:2161–2173, 2002. 148. Iismaa SE, Mearns BM, Lorand L, Graham RM. Transglutaminases and disease: lessons from genetically engineered mouse models and inherited disorders. Physiol Rev 89: 991–1023, 2009. 169. Kaetsu H, Hashiguchi T, Foster D, Ichinose A. Expression and release of the a and b subunits for human coagulation factor XIII in baby hamster kidney (BHK) cells. J Biochem 119:961–969, 1996. 170. Kappelmayer J, Bacsko G, Birinyi L, Zakany R, Kelemen E, Adany R. Consecutive appearance of coagulation factor XIII subunit A in macrophages, megakaryocytes, liver cells during early human development. Blood 86:2191–2197, 1995. 150. Inbal A, Lubetsky A, Krapp T, Castel D, Shaish A, Dickneitte G, Modis L, Muszbek L. Impaired wound healing in factor XIII deficient mice. Thromb Haemost 94:432– 437, 2005. 171. Kappelmayer J, Bacsko G, Kelemen E, Adany R. Onset and distribution of factor XIII-containing cells in the mesenchyme of chorionic villi during early phase of human placentation. Placenta 15:613– 623, 1994. 151. Inbal A, Muszbek L. Coagulation factor deficiencies and pregnancy loss. Semin Thromb Hemost 29:171–174, 2003. 172. Kappelmayer J, Simon A, Katona E, Szanto A, Nagy L, Kiss A, Kiss C, Muszbek L. Coagulation factor XIII-A. A flow cytometric intracellular marker in the classification of acute myeloid leukemias. Thromb Haemost 94:454 – 459, 2005. 152. Inbal A, Muszbek L, Lubetsky A, Katona E, Levi I, Karpati L, Nagler A. Platelets but not monocytes contribute to the plasma levels of factor XIII subunit A in patients undergoing autologous peripheral blood stem cell transplantation. Blood Coagul Fibrinolysis 15:249 –253, 2004. 173. Karimi M, Bereczky Z, Cohan N, Muszbek L. Factor XIII deficiency. Semin Thromb Hemost 35:426 – 438, 2009. 153. Isogai N, Ueda Y, Kurozumi N, Kamiishi H. Wound healing at the site of microvascular anastomosis: fibrin-stabilizing factor XIII administration and its effects. Microsurgery 11:40 – 46, 1990. 154. Ivaskevicius V, Seitz R, Kohler HP, Schroeder V, Muszbek L, Ariens RA, Seifried E, Oldenburg J. International registry on factor XIII deficiency: a basis formed mostly on European data. Thromb Haemost 97:914 –921, 2007. 155. Iwata H, Kitano T, Umetsu K, Yuasa I, Yamazaki K, Kemkes-Matthes B, Ichinose A. Distinct C-terminus of the B subunit of factor XIII in a population-associated major phenotype: the first case of complete allele-specific alternative splicing products in the coagulation and fibrinolytic systems. J Thromb Haemost 7:1084 –1091, 2009. 156. Jaffer FA, Sosnovik DE, Nahrendorf M, Weissleder R. Molecular imaging of myocardial infarction. J Mol Cell Cardiol 41:921–933, 2006. 157. Jagadeeswaran P, Reddy SV, Haas P. Synthesis of human coagulation factor XIII in yeast. Gene 86:279 –283, 1990. 158. Jansen JW, Haverkate F, Koopman J, Nieuwenhuis HK, Kluft C, Boschman TA. Influence of factor XIIIa activity on human whole blood clot lysis in vitro. Thromb Haemost 57:171–175, 1987. 159. Janus TJ, Lewis SD, Lorand L, Shafer JA. Promotion of thrombin-catalyzed activation of factor XIII by fibrinogen. Biochemistry 22:6269 – 6272, 1983. 174. Kasahara K, Souri M, Kaneda M, Miki T, Yamamoto N, Ichinose A. Impaired clot retraction in factor XIII A subunit-deficient mice. Blood 115:1277–1279, 2010. 175. Katona E, Haramura G, Karpati L, Fachet J, Muszbek L. A simple, quick one-step ELISA assay for the determination of complex plasma factor XIII (A2B2). Thromb Haemost 83:268 –273, 2000. 176. Katona EE, Ajzner E, Toth K, Karpati L, Muszbek L. Enzyme-linked immunosorbent assay for the determination of blood coagulation factor XIII A-subunit in plasma and in cell lysates. J Immunol Methods 258:127–135, 2001. 177. Kavai M, Adany R, Pasti G, Suranyi P, Szucs G, Muszbek L, Bojan F, Szegedi G. Marker profile, enzyme activity, and function of a human myelomonocytic leukemia cell line. Cell Immunol 139:531–540, 1992. 178. Kida M, Souri M, Yamamoto M, Saito H, Ichinose A. Transcriptional regulation of cell type-specific expression of the TATA-less A subunit gene for human coagulation factor XIII. J Biol Chem 274:6138 – 6147, 1999. 179. Kiesselbach TH, Wagner RH. Demonstration of factor XIII in human megakaryocytes by a fluorescent antibody technique. Ann NY Acad Sci 202:318 –328, 1972. 180. Kilian O, Fuhrmann R, Alt V, Noll T, Coskun S, Dingeldein E, Schnettler R, Franke RP. Plasma transglutaminase factor XIII induces microvessel ingrowth into biodegradable hydroxyapatite implants in rats. Biomaterials 26:1819 –1827, 2005. 181. Kimura S, Aoki N. Cross-linking site in fibrinogen for alpha 2-plasmin inhibitor. J Biol Chem 261:15591–15595, 1986. 160. Jayo A, Conde I, Lastres P, Jimenez-Yuste V, Gonzalez-Manchon C. New insights into the expression and role of platelet factor XIII-A. J Thromb Haemost 7:1184 –1191, 2009. 182. Kimura S, Tamaki T, Aoki N. Acceleration of fibrinolysis by the N-terminal peptide of alpha 2-plasmin inhibitor. Blood 66:157–160, 1985. 161. Jayo A, Conde I, Lastres P, Jimenez-Yuste V, Gonzalez-Manchon C. Possible role for cellular FXIII in monocyte-derived dendritic cell motility. Eur J Cell Biol 88:423– 431, 2009. 183. Kiss F, Hevessy Z, Veszpremi A, Katona E, Kiss C, Vereb G, Muszbek L, Kappelmayer JN. Leukemic lymphoblasts, a novel expression site of coagulation factor XIII subunit A. Thromb Haemost 96:176 –182, 2006. 162. Jelenska M, Kopec M, Breddin K. On the retraction of collagen and fibrin induced by normal, defective and modified platelets. Haemostasis 15:169 –175, 1985. 184. Kobayashi T, Asahina T, Okada Y, Terao T. Studies on the localization of adhesive proteins associated with the development of extravillous cytotrophoblast. Trophoblast Res 13:35–53, 1999. 163. Jensen PH, Lorand L, Ebbesen P, Gliemann J. Type-2 plasminogen-activator inhibitor is a substrate for trophoblast transglutaminase and factor XIIIa. Transglutaminasecatalyzed cross-linking to cellular and extracellular structures. Eur J Biochem 214:141– 146, 1993. 185. Kobayashi T, Terao T, Kojima T, Takamatsu J, Kamiya T, Saito H. Congenital factor XIII deficiency with treatment of factor XIII concentrate and normal vaginal delivery. Gynecol Obstet Invest 29:235–238, 1990. 164. Jobe SM, Leo L, Eastvold JS, Dickneite G, Ratliff TL, Lentz SR, Di Paola J. Role of FcRgamma and factor XIIIA in coated platelet formation. Blood 106:4146 – 4151, 2005. 186. Komanasin N, Catto AJ, Futers TS, van Hylckama Vlieg A, Rosendaal FR, Ariens RA. A novel polymorphism in the factor XIII B-subunit (His95Arg): relationship to subunit dissociation and venous thrombosis. J Thromb Haemost 3:2487–2496, 2005. Physiol Rev • VOL 91 • JULY 2011 • www.prv.org 967 Downloaded from http://physrev.physiology.org/ by 10.220.32.246 on June 14, 2017 149. Ikematsu S. An approach to the metabolism of factor XIII. Nippon Ketsueki Gakkai Zasshi 44:1499 –1505, 1981. MUSZBEK ET AL. 209. Lewis BA, Freyssinet JM, Holbrook JJ. An equilibrium study of metal ion binding to human plasma coagulation factor XIII. Biochem J 169:397– 402, 1978. 188. Kopec M, Latallo ZS, Stahl M, Wegrzynowicz Z. The effect of proteolytic enzymes on fibrin stabilizing factor. Biochim Biophys Acta 181:437– 445, 1969. 210. Lewis SD, Janus TJ, Lorand L, Shafer JA. Regulation of formation of factor XIIIa by its fibrin substrates. Biochemistry 24:6772– 6777, 1985. 189. Koseki-Kuno S, Yamakawa M, Dickneite G, Ichinose A. Factor XIII A subunit-deficient mice developed severe uterine bleeding events and subsequent spontaneous miscarriages. Blood 102:4410 – 4412, 2003. 211. Lim BC, Ariens RA, Carter AM, Weisel JW, Grant PJ. Genetic regulation of fibrin structure and function: complex gene-environment interactions may modulate vascular risk. Lancet 361:1424 –1431, 2003. 190. Kradin RL, Lynch GW, Kurnick JT, Erikson M, Colvin RB, McDonagh J. Factor XIII A is synthesized and expressed on the surface of U937 cells and alveolar macrophages. Blood 69:778 –785, 1987. 212. Liu JN, Kung W, Harpel PC, Gurewich V. Demonstration of covalent binding of lipoprotein(a) [Lp(a)] to fibrin and endothelial cells. Biochemistry 37:3949 –3954, 1998. 191. Krarup A, Gulla KC, Gal P, Hajela K, Sim RB. The action of MBL-associated serine protease 1 (MASP1) on factor XIII and fibrinogen. Biochim Biophys Acta 1784:1294 – 1300, 2008. 213. Liu S, Cerione RA, Clardy J. Structural basis for the guanine nucleotide-binding activity of tissue transglutaminase and its regulation of transamidation activity. Proc Natl Acad Sci USA 99:2743–2747, 2002. 192. Kulkarni S, Jackson SP. Platelet factor XIII and calpain negatively regulate integrin alphaIIbbeta3 adhesive function and thrombus growth. J Biol Chem 279:30697–30706, 2004. 214. Lockwood CR, Frayling TM. Combining genome and mouse knockout expression data to highlight binding sites for the transcription factor HNF1alpha. In Silico Biol 3:57–70, 2003. 193. Kurochkin IV, Procyk R, Bishop PD, Yee VC, Teller DC, Ingham KC, Medved LV. Domain structure, stability and domain-domain interactions in recombinant factor XIII. J Mol Biol 248:414 – 430, 1995. 215. Loewy AG, Dahlberg A, Dunathan K, Kriel R, Wolfinger HL ,Jr. Fibrinase. II. Some physical properties. J Biol Chem 236:2634 –2643, 1961. 194. Lahav J, Karniel E, Bagoly Z, Sheptovitsky V, Dardik R, Inbal A. Coagulation factor XIII serves as protein disulfide isomerase. Thromb Haemost 101:840 – 844, 2009. 195. Lai TS, Slaughter TF, Peoples KA, Greenberg CS. Site-directed mutagenesis of the calcium-binding site of blood coagulation factor XIIIa. J Biol Chem 274:24953–24958, 1999. 216. Loewy AG, Dunathan K, Gallant JA, Gardner B. Fibrinase. III. Some enzymatic properties. J Biol Chem 236:2644 –2647, 1961. 217. Loewy AG, Dunathan K, Kriel R, Wolfinger HL ,Jr. Fibrinase. I. Purification of substrate and enzyme. J Biol Chem 236:2625–2633, 1961. 218. Loewy AG, Gallant JA, Dunathan K. Fibrinase. IV. Effect on fibrin solubility. J Biol Chem 236:2648 –2655, 1961. 196. Laki K, Lorand L. On the solubility of fibrin clots. Science 108:280, 1948. 197. Lanir N, Ciano PS, Van de Water L, McDonagh J, Dvorak AM, Dvorak HF. Macrophage migration in fibrin gel matrices. II. Effects of clotting factor XIII, fibronectin, glycosaminoglycan content on cell migration. J Immunol 140:2340 –2349, 1988. 198. Larkin MA, Blackshields G, Brown NP, Chenna R, McGettigan PA, McWilliam H, Valentin F, Wallace IM, Wilm A, Lopez R, Thompson JD, Gibson TJ, Higgins DG. Clustal W and Clustal X version 2.0. Bioinformatics 23:2947–2948, 2007. 219. Loewy AG, Veneziale C, Forman M. Purification of the factor involved in the formation of urea-insoluble fibrin. Biochim Biophys Acta 26:670 – 671, 1957. 220. Lopaciuk S, Lovette KM, McDonagh J, Chuang HY. Subcellular distribution of fibrinogen and factor XIII in human blood platelets. Thromb Res 8:453– 465, 1976. 221. Lorand L. A study on the solubility of fibrin clots in urea. Hung Acta Physiol 1:192–196, 1948. 199. Lauer P, Metzner HJ, Zettlmeissl G, Li M, Smith AG, Lathe R, Dickneite G. Targeted inactivation of the mouse locus encoding coagulation factor XIII-A: hemostatic abnormalities in mutant mice and characterization of the coagulation deficit. Thromb Haemost 88:967–974, 2002. 222. Lorand L. Fibrin clots. Nature 166:694 – 695, 1950. 200. Lawler J. Thrombospondin-1 as an endogenous inhibitor of angiogenesis and tumor growth. J Cell Mol Med 6:1–12, 2002. 224. Lorand L, Jeong JM, Radek JT, Wilson J. Human plasma factor XIII: subunit interactions and activation of zymogen. Methods Enzymol 222:22–35, 1993. 201. Lee IH, Chung SI, Lee SY. Effects of Val34Leu and Val35Leu polymorphism on the enzyme activity of the coagulation factor XIII-A. Exp Mol Med 34:385–390, 2002. 225. Lorand L, Konishi K, Jacobsen A. Transpeptidation mechanism in blood clotting. Nature 194:1148 –1149, 1962. 202. Lee KN, Jackson KW, Christiansen VJ, Chung KH, McKee PA. A novel plasma proteinase potentiates alpha2-antiplasmin inhibition of fibrin digestion. Blood 103:3783– 3788, 2004. 226. Lorenz R, Born P, Olbert P, Classen M. Factor XIII substitution in ulcerative colitis. Lancet 345:449 – 450, 1995. 203. Lee KN, Jackson KW, Christiansen VJ, Lee CS, Chun JG, McKee PA. Why alphaantiplasmin must be converted to a derivative form for optimal function. J Thromb Haemost 5:2095–2104, 2007. 204. Lee KN, Lee CS, Tae WC, Jackson KW, Christiansen VJ, McKee PA. Cross-linking of wild-type and mutant alpha 2-antiplasmins to fibrin by activated factor XIII and by a tissue transglutaminase. J Biol Chem 275:37382–37389, 2000. 205. Lee KN, Lee SC, Jackson KW, Tae WC, Schwartzott DG, McKee PA. Effect of phenylglyoxal-modified alpha2-antiplasmin on urokinase-induced fibrinolysis. Thromb Haemost 80:637– 644, 1998. 206. Lee KN, Tae WC, Jackson KW, Kwon SH, McKee PA. Characterization of wild-type and mutant alpha2-antiplasmins: fibrinolysis enhancement by reactive site mutant. Blood 94:164 –171, 1999. 223. Lorand L, Credo RB, Janus TJ. Factor XIII (fibrin-stabilizing factor). Methods Enzymol 80:333–341, 1981. 227. Loscalzo J, Weinfeld M, Fless GM, Scanu AM. Lipoprotein(a), fibrin binding, plasminogen activation. Arteriosclerosis 10:240 –245, 1990. 228. Luscher EF. Fibrin-stabilizing factor from thrombocytes. Schweiz Med Wochenschr 87:1220 –1221, 1957. 229. Lynch GW, Slayter HS, Miller BE, McDonagh J. Characterization of thrombospondin as a substrate for factor XIII transglutaminase. J Biol Chem 262:1772–1778, 1987. 230. Ma J, Liew CC. Gene profiling identifies secreted protein transcripts from peripheral blood cells in coronary artery disease. J Mol Cell Cardiol 35:993–998, 2003. 231. Malara A, Gruppi C, Rebuzzini P, Visai L, Perotti C, Moratti R, Balduini C, Tira ME, Balduini A. Megakaryocyte-matrix interaction within bone marrow: new roles for fibronectin and factor XIII-A. Blood. In press. 207. Leifheit HJ, Cleve H. Analysis of the genetic polymorphism of coagulation factor XIIIB (FXIIIB) by isoelectric focusing. Electrophoresis 9:426 – 429, 1988. 232. Marinescu A, Cleary DB, Littlefield TR, Maurer MC. Structural features associated with the binding of glutamine-containing peptides to Factor XIII. Arch Biochem Biophys 406:9 –20, 2002. 208. Lesort M, Attanavanich K, Zhang J, Johnson GV. Distinct nuclear localization and activity of tissue transglutaminase. J Biol Chem 273:11991–11994, 1998. 233. Marx G, Korner G, Mou X, Gorodetsky R. Packaging zinc, fibrinogen, factor XIII in platelet alpha-granules. J Cell Physiol 156:437– 442, 1993. 968 Physiol Rev • VOL 91 • JULY 2011 • www.prv.org Downloaded from http://physrev.physiology.org/ by 10.220.32.246 on June 14, 2017 187. Komaromi I, Bagoly Z, Muszbek L. Factor XIII: novel structural and functional aspects. J Thromb Haemost 9:9 –20, 2011. MULTIPLE FUNCTIONS OF FACTOR XIII 234. Matsuka YV, Medved LV, Migliorini MM, Ingham KC. Factor XIIIa-catalyzed crosslinking of recombinant alpha C fragments of human fibrinogen. Biochemistry 35:5810 – 5816, 1996. 255. Muszbek L, Adany R, Kavai M, Boda Z, Lopaciuk S. Monocytes of patients congenitally deficient in plasma factor XIII lack factor XIII subunit a antigen and transglutaminase activity. Thromb Haemost 59:231–235, 1988. 235. McDonagh J, Fukue H. Determinants of substrate specificity for factor XIII. Semin Thromb Hemost 22:369 –376, 1996. 256. Muszbek L, Adany R, Mikkola H. Novel aspects of blood coagulation factor XIII. I. Structure, distribution, activation, and function. Crit Rev Clin Lab Sci 33:357– 421, 1996. 236. McDonagh J, McDonagh RP. Alternative pathways for the activation of factor XIII. Br J Haematol 30:465– 477, 1975. 237. McDonagh RP ,Jr, McDonagh J, Duckert F. The influence of fibrin crosslinking on the kinetics of urokinase-induced clot lysis. Br J Haematol 21:323–332, 1971. 238. McDonagh RP, McDonagh J, Petersen TE, Thogersen HC, Skorstengaard K, SottrupJensen L, Magnusson S, Dell A, Morris HR. Amino acid sequence of the factor XIIIa acceptor site in bovine plasma fibronectin. FEBS Lett 127:174 –178, 1981. 257. Muszbek L, Adany R, Szegedi G, Polgar J, Kavai M. Factor XIII of blood coagulation in human monocytes. Thromb Res 37:401– 410, 1985. 258. Muszbek L, Ariens RA, Ichinose A. Factor XIII: recommended terms and abbreviations. J Thromb Haemost 5:181–183, 2007. 259. Muszbek L, Bagoly Z, Bereczky Z, Katona E. The involvement of blood coagulation factor XIII in fibrinolysis and thrombosis. Cardiovasc Hematol Agents Med Chem 6:190 – 205, 2008. 260. Muszbek L, Bereczky Z, Bagoly Z, Shemirani AH, Katona E.Factor XIII and atherothrombotic diseases. Semin Thromb Hemost 36:18 –33. 240. Mendelsohn ME, Zhu Y, O’Neill S. The 29-kDa proteins phosphorylated in thrombinactivated human platelets are forms of the estrogen receptor-related 27-kDa heat shock protein. Proc Natl Acad Sci USA 88:11212–11216, 1991. 261. Muszbek L, Haramura G, Polgar J. Transformation of cellular factor XIII into an active zymogen transglutaminase in thrombin-stimulated platelets. Thromb Haemost 73: 702–705, 1995. 241. Merz D, Liu R, Johnson K, Terkeltaub R. IL-8/CXCL8 and growth-related oncogene alpha/CXCL1 induce chondrocyte hypertrophic differentiation. J Immunol 171:4406 – 4415, 2003. 262. Muszbek L, Polgar J, Boda Z. Platelet factor XIII becomes active without the release of activation peptide during platelet activation. Thromb Haemost 69:282–285, 1993. 242. Metha K, Turpin J, Lopez-Berestein G. Induction of tissue transglutaminase in human peripheral blood monocytes by intracellular delivery of retinoids. J Leukoc Biol 41:341– 348, 1987. 243. Mikkola H, Muszbek L, Laiho E, Syrjala M, Hamalainen E, Haramura G, Salmi T, Peltonen L, Palotie A. Molecular mechanism of a mild phenotype in coagulation factor XIII (FXIII) deficiency: a splicing mutation permitting partial correct splicing of FXIII A-subunit mRNA. Blood 89:1279 –1287, 1997. 263. Muszbek L, Yee VC, Hevessy Z. Blood coagulation factor XIII: structure and function. Thromb Res 94:271–305, 1999. 264. Mutch NJ, Koikkalainen JS, Fraser SR, Duthie KM, Griffin M, Mitchell J, Watson HG, Booth NA. Model thrombi formed under flow reveal the role of factor XIII-mediated cross-linking in resistance to fibrinolysis. J Thromb Haemost 8:2017–2024, 2010. 265. Nagy B ,Jr, Simon Z, Bagoly Z, Muszbek L, Kappelmayer J. Binding of plasma factor XIII to thrombin-receptor activated human platelets. Thromb Haemost 102:83– 89, 2009. 244. Mikkola H, Syrjala M, Rasi V, Vahtera E, Hamalainen E, Peltonen L, Palotie A. Deficiency in the A-subunit of coagulation factor XIII: two novel point mutations demonstrate different effects on transcript levels. Blood 84:517–525, 1994. 266. Nagy JA, Henriksson P, McDonagh J. Biosynthesis of factor XIII B subunit by human hepatoma cell lines. Blood 68:1272–1279, 1986. 245. Mirochnik Y, Kwiatek A, Volpert OV. Thrombospondin and apoptosis: molecular mechanisms and use for design of complementation treatments. Curr Drug Targets 9:851– 862, 2008. 267. Nahrendorf M, Aikawa E, Figueiredo JL, Stangenberg L, van den Borne SW, Blankesteijn WM, Sosnovik DE, Jaffer FA, Tung CH, Weissleder R. Transglutaminase activity in acute infarcts predicts healing outcome and left ventricular remodelling: implications for FXIII therapy and antithrombin use in myocardial infarction. Eur Heart J 29:445– 454, 2008. 246. Mishima Y, Nagao F, Ishibiki K, Matsuda M, Nakamura N. Factor XIII in the treatment of postoperative refractory wound-healing disorders. Results of a controlled study. Chirurg 55:803– 808, 1984. 247. Moaddel M, Falls LA, Farrell DH. The role of gamma A/gamma= fibrinogen in plasma factor XIII activation. J Biol Chem 275:32135–32140, 2000. 248. Moaddel M, Farrell DH, Dougherty MA, Frieol MG. Interactions of human fibrinogens with factor XIII: roles of calcium and the gamma peptide. Biochemistry 39:6698 – 6705, 2000. 249. Mortensen SB, Sottrup-Jensen L, Hansen HF, Rider D, Petersen TE, Magnusson S. Sequence location of a putative transglutaminase crosslinking site in human alpha 2-macroglobulin. FEBS Lett 129:314 –317, 1981. 268. Nahrendorf M, Hu K, Frantz S, Jaffer FA, Tung CH, Hiller KH, Voll S, Nordbeck P, Sosnovik D, Gattenlohner S, Novikov M, Dickneite G, Reed GL, Jakob P, Rosenzweig A, Bauer WR, Weissleder R, Ertl G. Factor XIII deficiency causes cardiac rupture, impairs wound healing, and aggravates cardiac remodeling in mice with myocardial infarction. Circulation 113:1196 –1202, 2006. 269. Nahrendorf M, Sosnovik DE, Waterman P, Swirski FK, Pande AN, Aikawa E, Figueiredo JL, Pittet MJ, Weissleder R. Dual channel optical tomographic imaging of leukocyte recruitment and protease activity in the healing myocardial infarct. Circ Res 100:1218 –1225, 2007. 270. Nahrendorf M, Weissleder R, Ertl G. Does FXIII deficiency impair wound healing after myocardial infarction? PLoS One 1:e48, 2006. 250. Mosesson MW. Fibrinogen and fibrin structure and functions. J Thromb Haemost 3:1894 –1904, 2005. 271. Nakano Y, Al-Jallad HF, Mousa A, Kaartinen MT. Expression and localization of plasma transglutaminase factor XIIIA in bone. J Histochem Cytochem 55:675– 685, 2007. 251. Mosesson MW, Siebenlist KR, Amrani DL, DiOrio JP. Identification of covalently linked trimeric and tetrameric D domains in crosslinked fibrin. Proc Natl Acad Sci USA 86:1113–1117, 1989. 272. Naski MC, Lorand L, Shafer JA. Characterization of the kinetic pathway for fibrin promotion of alpha-thrombin-catalyzed activation of plasma factor XIII. Biochemistry 30:934 –941, 1991. 252. Mosesson MW, Siebenlist KR, Hernandez I, Lee KN, Christiansen VJ, McKee PA. Evidence that alpha2-antiplasmin becomes covalently ligated to plasma fibrinogen in the circulation: a new role for plasma factor XIII in fibrinolysis regulation. J Thromb Haemost 6:1565–1570, 2008. 273. Niewiarowski S, Kirby EP, Brudzynski TM, Stocker K. Thrombocytin, a serine protease from Bothrops atrox venom. 2. Interaction with platelets and plasma-clotting factors. Biochemistry 18:3570 –3577, 1979. 253. Mosnier LO, Bouma BN. Regulation of fibrinolysis by thrombin activatable fibrinolysis inhibitor, an unstable carboxypeptidase B that unites the pathways of coagulation and fibrinolysis. Arterioscler Thromb Vasc Biol 26:2445–2453, 2006. 254. Murtaugh MP, Arend WP, Davies PJ. Induction of tissue transglutaminase in human peripheral blood monocytes. J Exp Med 159:114 –125, 1984. 274. Niewiarowski S, Markiewicz M, Nath N. Inhibition of the platelet-dependent fibrin retraction by the fibrin stabilizing factor (FSF, factor 13). J Lab Clin Med 81:641– 650, 1973. 275. Noll T, Wozniak G, McCarson K, Hajimohammad A, Metzner HJ, Inserte J, Kummer W, Hehrlein FW, Piper HM. Effect of factor XIII on endothelial barrier function. J Exp Med 189:1373–1382, 1999. Physiol Rev • VOL 91 • JULY 2011 • www.prv.org 969 Downloaded from http://physrev.physiology.org/ by 10.220.32.246 on June 14, 2017 239. Mehta K, Lopez-Berestein G, Moore WT, Davies PJ. Interferon-gamma requires serum retinoids to promote the expression of tissue transglutaminase in cultured human blood monocytes. J Immunol 134:2053–2056, 1985. MUSZBEK ET AL. 276. Nor JE, Mitra RS, Sutorik MM, Mooney DJ, Castle VP, Polverini PJ. Thrombospondin-1 induces endothelial cell apoptosis and inhibits angiogenesis by activating the caspase death pathway. J Vasc Res 37:209 –218, 2000. 297. Peschen M, Thimm C, Weyl A, Weiss JM, Kurz H, Augustin M, Simon JC, Schopf E, Vanscheidt W. Possible role of coagulation factor XIII in the pathogenesis of venous leg ulcers. Vasa 27:89 –93, 1998. 277. Nossel HL, Lanzkowsky P, Levy S, Mibashan RS, Hansen JD. A study of coagulation factor levels in women during labour and in their newborn infants. Thromb Diath Haemorrh 16:185–197, 1966. 298. Peter A, Lilja H, Lundwall A, Malm J. Semenogelin I and semenogelin II, the major gel-forming proteins in human semen, are substrates for transglutaminase. Eur J Biochem 252:216 –221, 1998. 278. Nurden AT, Kunicki TJ, Dupuis D, Soria C, Caen JP. Specific protein and glycoprotein deficiencies in platelets isolated from two patients with the gray platelet syndrome. Blood 59:709 –718, 1982. 299. Pettersen EF, Goddard TD, Huang CC, Couch GS, Greenblatt DM, Meng EC, Ferrin TE. UCSF chimera: a visualization system for exploratory research and analysis. J Comput Chem 25:1605–1612, 2004. 279. Nurminskaya M, Kaartinen MT. Transglutaminases in mineralized tissues. Front Biosci 11:1591–1606, 2006. 300. Philippou H, Rance J, Myles T, Hall SW, Ariens RA, Grant PJ, Leung L, Lane DA. Roles of low specificity and cofactor interaction sites on thrombin during factor XIII activation. Competition for cofactor sites on thrombin determines its fate. J Biol Chem 278:32020 –32026, 2003. 280. Nurminskaya M, Magee C, Nurminsky D, Linsenmayer TF. Plasma transglutaminase in hypertrophic chondrocytes: expression and cell-specific intracellular activation produce cell death and externalization. J Cell Biol 142:1135–1144, 1998. 301. Pinkas DM, Strop P, Brunger AT, Khosla C. Transglutaminase 2 undergoes a large conformational change upon activation. PLoS Biol 5:e327, 2007. 302. Polgar J, Hidasi V, Muszbek L. Non-proteolytic activation of cellular protransglutaminase (placenta macrophage factor XIII). Biochem J 267:557–560, 1990. 282. Nurminskaya MV, Recheis B, Nimpf J, Magee C, Linsenmayer TF. Transglutaminase factor XIIIA in the cartilage of developing avian long bones. Dev Dyn 223:24 –32, 2002. 303. Poon MC, Russell JA, Low S, Sinclair GD, Jones AR, Blahey W, Ruether BA, Hoar DI. Hemopoietic origin of factor XIII A subunits in platelets, monocytes, and plasma. Evidence from bone marrow transplantation studies. J Clin Invest 84:787–792, 1989. 283. Ogasawara MS, Aoki K, Katano K, Ozaki Y, Suzumori K. Factor XII but not protein C, protein S, antithrombin III, or factor XIII is a predictor of recurrent miscarriage. Fertil Steril 75:916 –919, 2001. 304. Quatresooz P, Paquet P, Hermanns-Le T, Pierard GE. Molecular mapping of Factor XIIIa-enriched dendrocytes in the skin (review). Int J Mol Med 22:403– 409, 2008. 284. Ogawa T, Morioka Y, Inoue T, Takano M, Tsuda S. Involvement of blood coagulation factor XIII in burn healing in the carbon tetrachloride-induced hepatic injury model in rats. Inflamm Res 44:264 –268, 1995. 285. Ohmori T, Yatomi Y, Asazuma N, Satoh K, Ozaki Y. Involvement of proline-rich tyrosine kinase 2 in platelet activation: tyrosine phosphorylation mostly dependent on alphaIIbbeta3 integrin and protein kinase C, translocation to the cytoskeleton and association with Shc through Grb2. Biochem J 347:561–569, 2000. 286. Okamoto M, Yamamoto T, Matsubara S, Kukita I, Takeya M, Miyauchi Y, Kambara T. Factor XIII-dependent generation of 5th complement component(C5)-derived monocyte chemotactic factor coinciding with plasma clotting. Biochim Biophys Acta 1138:53– 61, 1992. 287. Ortner E, Schroeder V, Walser R, Zerbe O, Kohler HP. Sensitive and selective detection of free FXIII activation peptide: a potential marker of acute thrombotic events. Blood 115:5089 –5096, 2010. 288. Otto HH, Schirmeister T. Cysteine proteases and their inhibitors. Chem Rev 97:133– 172, 1997. 289. Ozsoyl US, Hicsonmez G. Platelet aggregation in congenital factor XIII deficiency. Acta Haematol 56:314 –317, 1976. 290. Pabst MJ, Pabst KM, Handsman DB, Beranova-Giorgianni S, Giorgianni F. Proteome of monocyte priming by lipopolysaccharide, including changes in interleukin-1beta and leukocyte elastase inhibitor. Proteome Sci 6:13, 2008. 305. Radek JT, Jeong JM, Wilson J, Lorand L. Association of the A subunits of recombinant placental factor XIII with the native carrier B subunits from human plasma. Biochemistry 32:3527–3534, 1993. 306. Rao KM, Newcomb TF. Clot retraction in a factor XIII free system. Scand J Haematol 24:142–148, 1980. 307. Reed GL, Houng AK. The contribution of activated factor XIII to fibrinolytic resistance in experimental pulmonary embolism. Circulation 99:299 –304, 1999. 308. Reed GL, Matsueda GR, Haber E. Platelet factor XIII increases the fibrinolytic resistance of platelet-rich clots by accelerating the crosslinking of alpha 2-antiplasmin to fibrin. Thromb Haemost 68:315–320, 1992. 309. Rex S, Beaulieu LM, Perlman DH, Vitseva O, Blair PS, McComb ME, Costello CE, Freedman JE. Immune versus thrombotic stimulation of platelets differentially regulates signalling pathways, intracellular protein-protein interactions, and alpha-granule release. Thromb Haemost 102:97–110, 2009. 310. Rider DM, McDonagh J. Resistance of factor XIII to degradation or activation by plasmin. Biochim Biophys Acta 675:171–177, 1981. 311. Rijken DC, Lijnen HR. New insights into the molecular mechanisms of the fibrinolytic system. J Thromb Haemost 7:4 –13, 2009. 312. Ritchie H, Lawrie LC, Crombie PW, Mosesson MW, Booth NA. Cross-linking of plasminogen activator inhibitor 2 and alpha 2-antiplasmin to fibrin(ogen). J Biol Chem 275:24915–24920, 2000. 291. Park D, Choi SS, Ha KS. Transglutaminase 2: a multi-functional protein in multiple subcellular compartments. Amino Acids 39:619 – 631, 2010. 313. Ritchie H, Robbie LA, Kinghorn S, Exley R, Booth NA. Monocyte plasminogen activator inhibitor 2 (PAI-2) inhibits u-PA-mediated fibrin clot lysis and is cross-linked to fibrin. Thromb Haemost 81:96 –103, 1999. 292. Patthy L. Detecting homology of distantly related proteins with consensus sequences. J Mol Biol 198:567–577, 1987. 314. Robbins KC. A study on the conversion of fibrinogen to fibrin. Am J Physiol 142:581– 588, 1944. 293. Patthy L. Evolution of the proteases of blood coagulation and fibrinolysis by assembly from modules. Cell 41:657– 663, 1985. 294. Pedersen LC, Yee VC, Bishop PD, Le Trong I, Teller DC, Stenkamp RE. Transglutaminase factor XIII uses proteinase-like catalytic triad to crosslink macromolecules. Protein Sci 3:1131–1135, 1994. 295. Peng X, Zhang Y, Zhang H, Graner S, Williams JF, Levitt ML, Lokshin A. Interaction of tissue transglutaminase with nuclear transport protein importin-alpha3. FEBS Lett 446:35–39, 1999. 296. Penzes K, Kover KE, Fazakas F, Haramura G, Muszbek L. Molecular mechanism of the interaction between activated factor XIII and its glutamine donor peptide substrate. J Thromb Haemost 7:627– 633, 2009. 970 315. Robinson BR, Houng AK, Reed GL. Catalytic life of activated factor XIII in thrombi. Implications for fibrinolytic resistance and thrombus aging. Circulation 102:1151– 1157, 2000. 316. Rodeghiero F, Castaman GC, Di Bona E, Ruggeri M, Dini E. Successful pregnancy in a woman with congenital factor XIII deficiency treated with substitutive therapy. Report of a second case. Blut 55:45– 48, 1987. 317. Romanic AM, Arleth AJ, Willette RN, Ohlstein EH. Factor XIIIa cross-links lipoprotein(a) with fibrinogen and is present in human atherosclerotic lesions. Circ Res 83: 264 –269, 1998. 318. Rosenthal AK, Masuda I, Gohr CM, Derfus BA, Le M. The transglutaminase, Factor XIIIA, is present in articular chondrocytes. Osteoarthritis Cartilage 9:578 –581, 2001. Physiol Rev • VOL 91 • JULY 2011 • www.prv.org Downloaded from http://physrev.physiology.org/ by 10.220.32.246 on June 14, 2017 281. Nurminskaya MV, Linsenmayer TF. Immunohistological analysis of transglutaminase factor XIIIA expression in mouse embryonic growth plate. J Orthop Res 20:575–578, 2002. MULTIPLE FUNCTIONS OF FACTOR XIII 319. Rosenthal AK, Mosesson MW, Gohr CM, Masuda I, Heinkel D, Seibenlist KR. Regulation of transglutaminase activity in articular chondrocytes through thrombin receptor-mediated factor XIII synthesis. Thromb Haemost 91:558 –568, 2004. 320. Roth WJ, Fleit HB, Chung SI, Janoff A. Characterization of two distinct transglutaminases of murine bone marrow-derived macrophages: effects of exposure of viable cells to cigarette smoke on enzyme activity. J Leukoc Biol 42:9 –20, 1987. 321. Ryan AW, Hughes DA, Tang K, Kelleher DP, Ryan T, McManus R, Stoneking M. Natural selection and the molecular basis of electrophoretic variation at the coagulation F13B locus. Eur J Hum Genet 17:219 –227, 2009. 322. Sabo TM, Brasher PB, Maurer MC. Perturbations in factor XIII resulting from activation and inhibition examined by solution based methods and detected by MALDI-TOF MS. Biochemistry 46:10089 –10101, 2007. 323. Saito M, Asakura H, Yoshida T, Ito K, Okafuji K, Matsuda T. A familial factor XIII subunit B deficiency. Br J Haematol 74:290 –294, 1990. 324. Sakata Y, Aoki N. Cross-linking of alpha 2-plasmin inhibitor to fibrin by fibrin-stabilizing factor. J Clin Invest 65:290 –297, 1980. 326. Sakata Y, Mimuro J, Aoki N. Differential binding of plasminogen to crosslinked and noncrosslinked fibrins: its significance in hemostatic defect in factor XIII deficiency. Blood 63:1393–1401, 1984. 327. Sanchez C, Deberg MA, Bellahcene A, Castronovo V, Msika P, Delcour JP, Crielaard JM, Henrotin YE. Phenotypic characterization of osteoblasts from the sclerotic zones of osteoarthritic subchondral bone. Arthritis Rheum 58:442– 455, 2008. 328. Sane DC, Moser TL, Pippen AM, Parker CJ, Achyuthan KE, Greenberg CS. Vitronectin is a substrate for transglutaminases. Biochem Biophys Res Commun 157:115–120, 1988. 329. Sarvary A, Szucs S, Balogh I, Becsky A, Bardos H, Kavai M, Seligsohn U, Egbring R, Lopaciuk S, Muszbek L, Adany R. Possible role of factor XIII subunit A in Fcgamma and complement receptor-mediated phagocytosis. Cell Immunol 228:81–90, 2004. 330. Schroder V, Kohler HP. Effect of factor XIII Val34Leu on alpha2-antiplasmin incorporation into fibrin. Thromb Haemost 84:1128 –1130, 2000. 331. Schroeder V, Durrer D, Meili E, Schubiger G, Kohler HP. Congenital factor XIII deficiency in Switzerland: from the worldwide first case in 1960 to its molecular characterisation in 2005. Swiss Med Wkly 137:272–278, 2007. 341. Shemirani AH, Haramura G, Bagoly Z, Muszbek L. The combined effect of fibrin formation and factor XIII A subunit Val34Leu polymorphism on the activation of factor XIII in whole plasma. Biochim Biophys Acta 1764:1420 –1423, 2006. 342. Siebenlist KR, Meh DA, Mosesson MW. Protransglutaminase (factor XIII) mediated crosslinking of fibrinogen and fibrin. Thromb Haemost 86:1221–1228, 2001. 343. Siebenlist KR, Mosesson MW. Evidence of intramolecular cross-linked A alpha.gamma chain heterodimers in plasma fibrinogen. Biochemistry 35:5817–5821, 1996. 344. Siebenlist KR, Mosesson MW. Progressive cross-linking of fibrin gamma chains increases resistance to fibrinolysis. J Biol Chem 269:28414 –28419, 1994. 345. Siebenlist KR, Mosesson MW, Hernandez I, Bush LA, Di Cera E, Shainoff JR, Di Orio JP, Stojanovic L. Studies on the basis for the properties of fibrin produced from fibrinogen-containing gamma= chains. Blood 106:2730 –2736, 2005. 346. Sixma JJ, van den Berg A, Schiphorst M, Geuze HJ, McDonagh J. Immunocytochemical localization of albumin and factor XIII in thin cryo sections of human blood platelets. Thromb Haemost 51:388 –391, 1984. 347. Skorstengaard K, Halkier T, Hojrup P, Mosher D. Sequence location of a putative transglutaminase cross-linking site in human vitronectin. FEBS Lett 262:269 –274, 1990. 348. Sobel JH, Gawinowicz MA. Identification of the alpha chain lysine donor sites involved in factor XIIIa fibrin cross-linking. J Biol Chem 271:19288 –19297, 1996. 349. Sorensen ES, Rasmussen LK, Moller L, Jensen PH, Hojrup P, Petersen TE. Localization of transglutaminase-reactive glutamine residues in bovine osteopontin. Biochem J 304: 13–16, 1994. 350. Souri M, Kaetsu H, Ichinose A. Sushi domains in the B subunit of factor XIII responsible for oligomer assembly. Biochemistry 47:8656 – 8664, 2008. 351. Souri M, Koseki-Kuno S, Takeda N, Degen JL, Ichinose A. Administration of factor XIII B subunit increased plasma factor XIII A subunit levels in factor XIII B subunit knockout mice. Int J Hematol 87:60 – 68, 2008. 352. Souri M, Koseki-Kuno S, Takeda N, Yamakawa M, Takeishi Y, Degen JL, Ichinose A. Male-specific cardiac pathologies in mice lacking either the A or B subunit of factor XIII. Thromb Haemost 99:401– 408, 2008. 332. Schroeder V, Vuissoz JM, Caflisch A, Kohler HP. Factor XIII activation peptide is released into plasma upon cleavage by thrombin and shows a different structure compared to its bound form. Thromb Haemost 97:890 – 898, 2007. 353. Standeven KF, Carter AM, Grant PJ, Weisel JW, Chernysh I, Masova L, Lord ST, Ariens RA. Functional analysis of fibrin ␥-chain cross-linking by activated factor XIII: determination of a cross-linking pattern that maximizes clot stiffness. Blood 110:902–907, 2007. 333. Schroth M, Meissner U, Cesnjevar R, Weyand M, Singer H, Rascher W, Klinge J. Plasmatic factor XIII reduces severe pleural effusion in children after open-heart surgery. Pediatr Cardiol 27:56 – 60, 2006. 354. Stirling Y, Woolf L, North WR, Seghatchian MJ, Meade TW. Haemostasis in normal pregnancy. Thromb Haemost 52:176 –182, 1984. 334. Schwartz ML, Pizzo SV, Hill RL, McKee PA. Human Factor XIII from plasma and platelets. Molecular weights, subunit structures, proteolytic activation, and crosslinking of fibrinogen and fibrin. J Biol Chem 248:1395–1407, 1973. 355. Streit M, Velasco P, Riccardi L, Spencer L, Brown LF, Janes L, Lange-Asschenfeldt B, Yano K, Hawighorst T, Iruela-Arispe L, Detmar M. Thrombospondin-1 suppresses wound healing and granulation tissue formation in the skin of transgenic mice. EMBO J 19:3272–3282, 2000. 335. Seelig GF, Folk JE. Noncatalytic subunits of human blood plasma coagulation factor XIII. Preparation and partial characterization of modified forms. J Biol Chem 255: 8881– 8886, 1980. 336. Seitz R, Duckert F, Lopaciuk S, Muszbek L, Rodeghiero F, Seligsohn U. ETRO Working Party on Factor XIII questionnaire on congenital factor XIII deficiency in Europe: status and perspectives. Study Group Semin Thromb Hemost 22:415– 418, 1996. 337. Seitz R, Leugner F, Katschinski M, Immel A, Kraus M, Egbring R, Goke B. Ulcerative colitis and Crohn’s disease: factor XIII, inflammation and haemostasis. Digestion 55: 361–367, 1994. 338. Seiving B, Ohlsson K, Linder C, Stenberg P. Transglutaminase differentiation during maturation of human blood monocytes to macrophages. Eur J Haematol 46:263–271, 1991. 339. Semba U, Chen J, Ota Y, Jia N, Arima H, Nishiura H, Yamamoto T. A plasma protein indistinguishable from ribosomal protein S19: conversion to a monocyte chemotactic factor by a factor XIIIa-catalyzed reaction on activated platelet membrane phosphatidylserine in association with blood coagulation. Am J Pathol 176:1542–1551, 2010. 356. Sugimura Y, Hosono M, Wada F, Yoshimura T, Maki M, Hitomi K. Screening for the preferred substrate sequence of transglutaminase using a phage-displayed peptide library: identification of peptide substrates for TGASE 2 and Factor XIIIA. J Biol Chem 281:17699 –17706, 2006. 357. Sumi Y, Ichikawa Y, Nakamura Y, Miura O, Aoki N. Expression and characterization of pro alpha 2-plasmin inhibitor. J Biochem 106:703–707, 1989. 358. Swirski FK, Nahrendorf M, Etzrodt M, Wildgruber M, Cortez-Retamozo V, Panizzi P, Figueiredo JL, Kohler RH, Chudnovskiy A, Waterman P, Aikawa E, Mempel TR, Libby P, Weissleder R, Pittet MJ. Identification of splenic reservoir monocytes and their deployment to inflammatory sites. Science 325:612– 616, 2009. 359. Takahashi N, Takahashi Y, Putnam FW. Primary structure of blood coagulation factor XIIIa (fibrinoligase, transglutaminase) from human placenta. Proc Natl Acad Sci USA 83:8019 – 8023, 1986. 360. Tamaki T, Aoki N. Cross-linking of alpha 2-plasmin inhibitor to fibrin catalyzed by activated fibrin-stabilizing factor. J Biol Chem 257:14767–14772, 1982. Physiol Rev • VOL 91 • JULY 2011 • www.prv.org 971 Downloaded from http://physrev.physiology.org/ by 10.220.32.246 on June 14, 2017 325. Sakata Y, Aoki N. Significance of cross-linking of alpha 2-plasmin inhibitor to fibrin in inhibition of fibrinolysis and in hemostasis. J Clin Invest 69:536 –542, 1982. 340. Serrano K, Devine DV. Intracellular factor XIII crosslinks platelet cytoskeletal elements upon platelet activation. Thromb Haemost 88:315–320, 2002. MUSZBEK ET AL. 376. Weisel JW, Litvinov RI. The biochemical and physical process of fibrinolysis and effects of clot structure and stability on the lysis rate. Cardiovasc Hematol Agents Med Chem 6:161–180, 2008. 362. Torocsik D, Bardos H, Nagy L, Adany R. Identification of factor XIII-A as a marker of alternative macrophage activation. Cell Mol Life Sci 62:2132–2139, 2005. 377. Weiss MS, Metzner HJ, Hilgenfeld R. Two non-proline cis peptide bonds may be important for factor XIII function. FEBS Lett 423:291–296, 1998. 363. Torocsik D, Szeles L, Paragh G ,Jr, Rakosy Z, Bardos H, Nagy L, Balazs M, Inbal A, Adany R. Factor XIII-A is involved in the regulation of gene expression in alternatively activated human macrophages. Thromb Haemost 104:709 –717, 2010. 378. Wells PS, Anderson JL, Scarvelis DK, Doucette SP, Gagnon F. Factor XIII Val34Leu variant is protective against venous thromboembolism: a huge review and metaanalysis. Am J Epidemiol 164:101–109, 2006. 364. Turner BT ,Jr, Maurer MC. Evaluating the roles of thrombin and calcium in the activation of coagulation factor XIII using H/D exchange and MALDI-TOF MS. Biochemistry 41:7947–7954, 2002. 379. Wolpl A, Lattke H, Board PG, Arnold R, Schmeiser T, Kubanek B, Robin-Winn M, Pichelmayr R, Goldmann SF. Coagulation factor XIII A and B subunits in bone marrow and liver transplantation. Transplantation 43:151–153, 1987. 365. Turner BT ,Jr, Sabo TM, Wilding D, Maurer MC. Mapping of factor XIII solvent accessibility as a function of activation state using chemical modification methods. Biochemistry 43:9755–9765, 2004. 380. Wozniak G, Dapper F, Alemany J. Factor XIII in ulcerative leg disease: background and preliminary clinical results. Semin Thromb Hemost 22:445– 450, 1996. 366. Ueyama M, Urayama T. The role of factor XIII in fibroblast proliferation. Jpn J Exp Med 48:135–142, 1978. 381. Wozniak G, Noll T, Akinturk H, Thul J, Muller M. Factor XIII prevents development of myocardial edema in children undergoing surgery for congenital heart disease. Ann NY Acad Sci 936:617– 620, 2001. 367. Valnickova Z, Enghild JJ. Human procarboxypeptidase U, or thrombin-activable fibrinolysis inhibitor, is a substrate for transglutaminases. Evidence for transglutaminasecatalyzed cross-linking to fibrin. J Biol Chem 273:27220 –27224, 1998. 382. Yee VC, Pedersen LC, Bishop PD, Stenkamp RE, Teller DC. Structural evidence that the activation peptide is not released upon thrombin cleavage of factor XIII. Thromb Res 78:389 –397, 1995. 368. Van Wersch JW, Vooijs ME, Ubachs JM. Coagulation factor XIII in pregnant smokers and non-smokers. Int J Clin Lab Res 27:68 –71, 1997. 383. Yee VC, Pedersen LC, Le Trong I, Bishop PD, Stenkamp RE, Teller DC. Threedimensional structure of a transglutaminase: human blood coagulation factor XIII. Proc Natl Acad Sci USA 91:7296 –7300, 1994. 369. Vanscheidt W, Hasler K, Wokalek H, Niedner R, Schopf E. Factor XIII-deficiency in the blood of venous leg ulcer patients. Acta Derm Venereol 71:55–57, 1991. 370. Voko Z, Bereczky Z, Katona E, Adany R, Muszbek L. Factor XIII Val34Leu variant protects against coronary artery disease. A meta-analysis. Thromb Haemost 97:458 – 463, 2007. 371. Walter M, Nyman D, Krajnc V, Duckert F. The activation of plasma factor XIII with the snake venom enzymes ancrod and batroxobin marajoensis. Thromb Haemost 38:438 – 446, 1977. 372. Wang DL, Annamalai AE, Ghosh S, Gewirtz AM, Colman RW. Human platelet factor V is crosslinked to actin by FXIIIa during platelet activation by thrombin. Thromb Res 57:39 –57, 1990. 373. Wang G, Beier F. Rac1/Cdc42 and RhoA GTPases antagonistically regulate chondrocyte proliferation, hypertrophy, and apoptosis. J Bone Miner Res 20:1022–1031, 2005. 374. Wartiovaara U, Mikkola H, Szoke G, Haramura G, Karpati L, Balogh I, Lassila R, Muszbek L, Palotie A. Effect of Val34Leu polymorphism on the activation of the coagulation factor XIII-A. Thromb Haemost 84:595– 600, 2000. 375. Webb GC, Coggan M, Ichinose A, Board PG. Localization of the coagulation factor XIII B subunit gene (F13B) to chromosome bands 1q31–32.1 and restriction fragment length polymorphism at the locus. Hum Genet 81:157–160, 1989. 972 384. Yorifuji H, Anderson K, Lynch GW, Van de Water L, McDonagh J. B protein of factor XIII: differentiation between free B and complexed B. Blood 72:1645–1650, 1988. 385. Zamboni P, De Mattei M, Ongaro A, Fogato L, Carandina S, De Palma M, Tognazzo S, Scapoli GL, Serino ML, Caruso A, Liboni A, Gemmati D. Factor XIII contrasts the effects of metalloproteinases in human dermal fibroblast cultured cells. Vasc Endovascular Surg 38:431– 438, 2004. 386. Zamboni P, Gemmati D. Clinical implications of gene polymorphisms in venous leg ulcer: a model in tissue injury and reparative process. Thromb Haemost 98:131–137, 2007. 387. Zhang R, Murakami S, Coustry F, Wang Y, de Crombrugghe B. Constitutive activation of MKK6 in chondrocytes of transgenic mice inhibits proliferation and delays endochondral bone formation. Proc Natl Acad Sci USA 103:365–370, 2006. 388. Zhou J, Tang PC, Qin L, Gayed PM, Li W, Skokos EA, Kyriakides TR, Pober JS, Tellides G. CXCR3-dependent accumulation and activation of perivascular macrophages is necessary for homeostatic arterial remodeling to hemodynamic stresses. J Exp Med 207:1951–1966, 2010. 389. Zhu Y, Tassi L, Lane W, Mendelsohn ME. Specific binding of the transglutaminase, platelet factor XIII, to HSP27. J Biol Chem 269:22379 –22384, 1994. Physiol Rev • VOL 91 • JULY 2011 • www.prv.org Downloaded from http://physrev.physiology.org/ by 10.220.32.246 on June 14, 2017 361. Tognazzo S, Gemmati D, Palazzo A, Catozzi L, Carandina S, Legnaro A, Tacconi G, Scapoli GL, Zamboni P. Prognostic role of factor XIII gene variants in nonhealing venous leg ulcers. J Vasc Surg 44:815– 819, 2006.