Survey
* Your assessment is very important for improving the workof artificial intelligence, which forms the content of this project
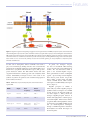
Features Communication in Plants and Microbes Differences and similarities Innate immunity in plants and animals Cara H. Haney (Harvard Medical School and Massachusetts General Hospital, USA), Jonathan Urbach (Massachusetts General Hospital, USA) and Frederick M. Ausubel (Harvard Medical School and Massachusetts General Hospital, USA) Plants and animals must avoid becoming a free meal to microbes, which vastly outnumber eukaryotic life in both quantity and diversity. Adaptive immunity in the strict sense, whereby the host creates an immunological memory after exposure to a pathogen, is limited to vertebrates. Both plants and animals (including insects and mammals) have an innate immune system, which helps protect hosts from the majority of microbes they encounter during their lifetime. Plant and animal innate immune systems recognize an overlapping set of conserved microbe-associated molecular patterns (MAMPs). This observation suggests that the innate immune system in plants and animals may have been derived from a common ancestor. However, the majority of data indicate that innate immunity has arisen independently in plants and animals and that functional overlap is the result of convergent evolution: confronted with the same problem, and given the same molecular tools, plants and animals have independently derived similar solutions. This review discusses the functional and mechanistic details of the innate immune system in plants and animals including receptor-mediated immunity, endolysosomal immunity, and the interplay of the innate immune system and host-associated microbial communities. The evolutionary impetus for an innate immune system Microbes thrive in nearly every environment on the planet, and given nutrient limitations in soil and water, eukaryotes provide an ideal habitat for microbes. Plants are able to synthesize their own carbon, which can be limiting for microbial growth in soil1. Animals forage and collect carbon and other nutrients. A microbe that is lucky enough to live in association with plant roots or inside an animal gut has first access to a potential free meal. Plants and animals also depend on these microbes; they help with nutrient acquisition, and in fact act as another line of defence against pathogenic microbes (discussed below). Despite potential selection pressures for microbes to take more and more nutrients from their hosts, the majority of host-associated microbes are non-pathogenic. This is in part Key words: NF-κB-family transcription factor, pathogen, resistanceprotein, rhizosphere, type III effector. because eukaryotes have sophisticated surveillance systems that monitor the presence of microbes in their environments and within themselves. Once a potential pathogen has been perceived, the innate immune system activates a number of defence responses including production of antimicrobial compounds and localized host cell death. Extracellular perception: transmembrane immune receptors and signalling Plants and animals must contend with similar microbial diversity and consequentially have the ability to recognize conserved MAMPs. These include bacterial flagellin, peptidoglycan and lipopolysaccharide (LPS), and fungal chitin (Table 1). In plants and animals, some MAMPs are recognized by pattern-recognition receptors (PRRs) that contain an extracellular leucine-rich repeat (LRR) domain (Figure 1). After MAMP recognition, receptors in plants and animals signal through mitogenactivated protein kinase (MAPK) cascades, which in turn activate transcription of defence-related genes. In animals and insects, a subset of MAMPs are recognized directly or indirectly by PRRs called Toll receptors in insects and Toll-like receptors (TLRs) in vertebrates. Initially identified in Drosophila Abbreviations: APAF-1, apoptotic protease-activating factor 1; CERK1, chitin elicitor receptor kinase 1; FLS2, Flagellin-Sensing2; LYSM1, lysin motif domain 1; LPS, ARC, NBS domain in APAF-1; NBS, nucleotide-binding site; PRR, pattern-recognition receptor; R-protein, resistance-protein; TIR, Toll/interleukin receptor; TLR, Toll-like receptor. 1 October 2014 © Biochemical Society Communication in Plants and Microbes Features Figure 1. Flagellin recognition and signalling is suggestive of convergent evolution in MAMP perception in plants and animals. Plant and animal flagellin receptors (FLS2 in plants and TLR5 in animals) recognize distinct epitopes of bacterial flagellin. Transmembrane PRRs in both plants and animals have extracellular LRR receptor domain, but have different cytoplasmic domains. Signalling in plants and animals makes use of a MAPK cascade but the specific molecular players including PRR-interacting proteins and transcription factor families are not conserved. The pathways in both FLS2 and TLR5 signalling are vastly simplified to emphasize specific similarities and differences. for their role in development, TLRs in mammals were found to play a role in immunity by binding molecules such as bacterial LPS, peptidoglycan and flagellin. TLRs have an extracellular LRR receptor domain and a cytoplasmic Toll/interleukin receptor (TIR) protein– protein interaction domain. The TIR domains interact with other cytoplasmic TIR-domain-containing proteins such as MyD88, which activates NF-κB-family transcription factors. This results in the synthesis of cytokines in vertebrates and of antimicrobial peptides in both vertebrates and insects2. Table 1. MAMPs that are perceived by both plants and animals and their cognate PRRs. MAMP Origin Plant receptor(s) Animal receptor(s) Flagellin Bacterial FLS23 TLR57,22 LPS Bacterial Unknown TLR423,24 Peptidoglycan Bacterial CERK1, LYSM1 and LYSM325,26 Nod120 and Nod227 Chitin Fungal CERK128 Unknown In plants, the analogous receptors also have an extracellular LRR-containing receptor domain and an intracellular kinase domain. The best-characterized PRR in plants is the bacterial flagellin receptor, FLS23,4, which binds a 22-amino-acid flagellin peptide5. Upon binding bacterial flagellin, FLS2 interacts with another receptor-like kinase, BAK16. Together FLS2 and BAK1 activate a MAPK cascade, which activates transcription via WRKY family transcription factors4 (Figure 1). Flagellin perception in animals is functionally very similar: flagellin perception by PRRs activates transcription via a MAPK cascade; however, the molecular details are not shared (Figure 1). TLR5 performs the analogous flagellin-perception function in humans7, although it binds to a distinct epitope of bacterial flagellin8. TLR5 interacts with the cytoplasmic TIR-containing MyD88 to activate (i) transcription via a MAPK signalling cascade and (ii) NF-κB-dependent transcription through a MAPK-independent mechanism. Both MyD88 and NF-κB homologues are absent from plants. October 2014 © Biochemical Society 2 Features Communication in Plants and Microbes Figure 2. Presence of NBS and LRR domains in plants, animals and bacteria. LRR (purple) and WD40 repeat (blue) and NBS (green; NB-ARC and NACHT) domains are present in bacterial and archaeal genomes. The NBS–LRR combination is extremely rare or non-existent except in plant disease R-proteins and animal Nod-like immune receptors. Plant and animal innate immune receptors appear to have independently co-opted these domains for defence purposes. This is supported by (i) differing domain structure between the plant and animal NBS-containing proteins, and (ii) apparent functional divergence in proteins with the most similar NBS domains in plants and animals. Note: there are additional NBS-domain-containing proteins found in animals, bacteria and Archaea whose domain structures are not included here. The flagellin-recognition case exemplifies the parallels and distinctions in MAMP perception and PRR-dependent signalling in plants and animals. PRRs in plants and animals share some similarity in protein domains and domain architecture and have similarly structured signalling modules. However, although functionally analogous, the organization of protein domains, and the exact downstream molecular players (such as transcription factors), are not conserved between plants and animals. Taken together, these data suggest evolutionarily independent solutions to the problem of perceiving and responding to bacterial infection9. Intracellular perception: cytoplasmic NBS–LRR receptors Plants and animals have a second class of intracellular innate immune receptors. Like the transmembrane receptors, these receptors have homologous domains in plants and animals: they both have nucleotidebinding site (NBS) domains adjacent to either an N-terminal LRR domain (in both plants and animals) or a WD40-repeat domain (in animals only; Figure 2). The C-terminal domains are variable in animals but typically contain either a TIR or a coiled-coil domain in plants. The variation of C-terminal domains, and divergent functions and molecular partners (discussed below) suggest that plants and animals have both co-opted an ancestral NBS-domain-containing protein independently to perform similar functions. NBS-containing proteins in plants and animals have diverse functions and divergent NBS domains. There are 14 mammalian NBScontaining proteins including APAF-1 (apoptotic protease-activating factor 1) and Nod proteins. Nod1 and Nod2, which contain LRR domains, recognize peptidoglycan degradation products and like 3 October 2014 © Biochemical Society TLRs, activate NF-κB-like transcription factors. APAF-1 (which has a WD40-repeat domain and has homologues in all sequenced animal genomes including CED-4 in Caenorhabditis elegans), interacts with caspases to trigger programmed cell death and does not function in innate immunity10. The NBS domains in APAF-1 and Nod proteins are divergent (called NB-ARC and NACHT domains respectively) and appear to have originated in bacteria11,12 (Figure 2). In plants, NBS–LRR proteins, also called resistance-proteins (or ‘R-proteins’), were initially identified because they conferred resistance to specific pathogens. Many successful Gram-negative plant pathogens have type III secretion systems that translocate effector proteins directly into their host cells. In contrast with MAMP recognition by animal Nod genes, plant R-genes recognize type III effectors or the damage the effectors cause to the host cell13. In addition to functional divergence of NBS-containing proteins in plants and animals, phylogenetic analysis of NBS domains in plants, animals and bacteria also points to distinct origins for plant and animal intracellular receptors11,12 (J. Urbach and F.M. Ausubel, unpublished work) (Figure 2). The individual domains found in NBS-containing proteins (LRR, NB-ARC, NACHT, etc.) are found in bacterial and archaeal genomes, indicating that they have ancient origins11 (Figure 2). Phylogenetic analysis reveals that the NBS domain in plant R-genes is more similar to the NB-ARC domain in animal APAF-1 (involved in apoptosis) than it is to the NACHT domain in the functionally similar animal Nod genes (J. Urbach and F.M. Ausubel, unpublished work). Collectively, functional and molecular data support convergent evolution for NBS-containing proteins in plants and animals. Intracellular innate immunity and endolysosomal trafficking Animal macrophages function in the endocytosis and destruction of potential pathogens. When immunity is successful, invading microbes are routed to the host lysosome, where the extreme pH and lytic enzymes provide a powerful means of bacterial extermination. A successful pathogen must escape this sure destruction and often does so by convincing the host that it is cargo to be routed elsewhere in the cell. In addition to using type III effectors to block the PRRs, pathogens such as Salmonella and Shigella are able to use their type III effectors to modulate host endolysosomal trafficking14. In animals, endolysosomal immunity also aids in immunity to toxins and viruses; this is not a wellstudied area in plants and is not discussed here. In plants, there are no known intracellular bacterial pathogens, and direct endocytosis of microbes is not considered to be a routine part of the innate immune system. However, there is some indication that endolysosomal immunity is functional in plants. Symbiotic nitrogenfixing bacteria, collectively referred to as ‘rhizobia’, live intracellularly in leguminous plants (including beans, peas and alfalfa). Most rhizobia are alpha-proteobacteria, and interestingly, the closest relatives of many rhizobia are intracellular animal pathogens and symbionts, including the mammalian pathogen Brucella and insect symbiont Wolbachia. Successful infection among plant- and animal-associated α-proteobacteria depends on a common set of bacterial genes and traits including a modified LPS, a type III secretion system, homologous two- Communication in Plants and Microbes Features Figure 3. Functional conservation of innate immune pathways in plants and animals. On a functional and conceptual level, innate immune pathways are conserved in plants and animals. Innate immune pathways include (1) extracellular MAMP recognition by PRRs, which activates transcription of defence-related genes; (2) intracellular MAMP and effector recognition by NBS–LRRs, which activate transcription of defence-related genes; (3) trafficking of microbes to the lytic compartment (in plants this is only known to be relevant to nitrogen-fixing bacteria); and (4) pathogen protection by association with a stable microbial community. In the case of (1) and (2), molecular data indicate that functional similarities have arisen through convergent evolution9,11. component systems and an RpoN family extracytoplasmic σ-factor15: shared virulence determinants between plant- and animal-associated α-proteobacteria may indicate common host cell targets. Rhizobia that are able to infect but lack certain virulence determinants (type III secretion, RpoN, etc.) are unable to persist within their host cell. Similarly, RpoN is required for Burkholderia cenocepacia to avoid fusion to the animal lysosome16. Plant endosomal compartments containing nitrogen-fixing bacteria undergo a natural senescence where they fuse to the plant lytic vacuole (analogous to the animal lysosome); this process requires a plant Rab7 small GTPase, also required for endolysosomal trafficking in animals17. At the functional level, this indicates a shared mechanism of endolysosomal immunity. Immunity and commensal microbial communities Plants and animals both associate with rich microbial communities, on which they depend for defence, nutrient acquisition and normal development. In animals, there is a growing body of evidence indicating that the innate immune system is essential for structuring these communities, and for preventing commensals from turning into pathogens. This is most clearly apparent in immunocompromised individuals where normally commensal microbes can cause disease. The molecular and biochemical details of how the innate immune system contributes to maintaining a balance with commensal microbes is just beginning to be elucidated in animals. A number of antimicrobial compounds are essential for limiting the numbers of microbes in the human gut or maintaining the balance of bacterial phyla. These include C-type lectins involved in restricting bacterial proliferation near the epithelial cell layer18, and defensins required for limiting growth of Bacteroidetes19. The role of the plant innate immune system in establishing and maintaining microbial communities is an area of active research in our and other laboratories. In plants, there is a huge body of evidence indicating that the microbiome itself is a natural component of the plant innate October 2014 © Biochemical Society 4 Features Communication in Plants and Microbes immune system. Root-associated bacteria, including Bacillus and Pseudomonas spp., and fungi, including Trichoderma, have long been known to protect their plant hosts from pathogens through a variety of mechanisms, including (i) the production of antimicrobial compounds, (ii) induction of host defences and (iii) out-competing pathogens in the rhizosphere20,21. There is some evidence in animals that the gut microbiome may play a protective function against potential pathogens. After disruption of normal gut microbes (through antibiotic treatment, for instance), normally commensal microbes such as Clostridium difficile can become pathogenic, suggesting the microbiome normally functions to keep C. difficile in a non-pathogenic state. The molecular mechanisms underlying the specificity of host–microbiome associations and how they contribute to protection against pathogens are areas of active research. Perspectives and conclusions Innate immunity in plants and animals has functionally analogous features. These include (i) recognition of MAMPs via PRRs and activation of defence responses by transmembrane; (ii) intracellular PRRs; (iii) endolysosomal trafficking to destroy invading microbes; and (iv) give-and-take with host-associated microbial communities (Figure 3). In cases where the molecular details are well understood (mainly PRRs), all evidence points to convergent evolution of mechanisms in innate immunity in plants and animals. It remains to be seen if the same is true for the role of the endolysosomal pathways in innate immunity and if common mechanisms are involved in shaping plant- and animalassociated microbial communities. ■ Cara Haney is funded by the Gordon and Betty Moore Foundation through Grant GBMF [grant number 2550.01] to the Life Sciences Research Foundation. Jonathan Urbach and Cara Haney are supported in part by National Institutes of Health [grant number R37GM48707], awarded to Frederick Ausubel. Cara Haney graduated from Cornell University in 2003 with a BS in plant science and received her PhD in cell and molecular biology from Stanford University in 2011. Dr Haney is a Gordon and Betty Moore Foundation postdoctoral fellow of the Life Sciences Research Foundation. She is currently a postdoctoral fellow in the Ausubel Lab at Harvard Medical School/Massachusetts General Hospital studying beneficial host-associated microbes. email: [email protected] Jonathan Urbach received his BA from Cornell University in 1989 and a PhD in chemistry from Harvard University in 2000. He is currently a bioinformatics specialist in the Molecular Biology Department at Massachusetts General Hospital. email: [email protected] Frederick Ausubel is a Professor of genetics and molecular biology at Harvard Medical School and Massachusetts General Hospital. Dr Ausubel was elected to the National Academy of Sciences in 1994 and received the 2014 Thomas Hunt Morgan Medal for lifetime achievement in the field of genetics. His laboratory focuses on innate immunity in model systems including Arabidopsis and C. elegans. email: ausubel@molbio. mgh.harvard.edu References 1. Aldén, L., Demoling, F. and Bååth, E. (2001) Appl. Environ. Microbiol. 67, 1830–1838 2. Kawai, T. and Akira, S. (2010) Nat. Immunol. 11, 373–384 3. Gomez-Gomez, L. and Boller, T. (2000) Mol. Cell 5, 1003–1011 4. Robatzek, S. and Wirthmueller, L. (2013) Protoplasma 250, 671–681 5. Felix, G. Duran, J.D., Volko, S. and Boller, T. (1999) Plant J. 18, 265–276 6. Heese, A., Hann, D.R., Gimenez-Ibanez, S. et al. (2007) Proc. Natl. Acad. Sci. U.S.A. 104, 12217–12222 7. Hayashi, F., Smith, K.D., Ozinsky, A. et al. (2001) Nature 410, 1099–1103 8. Donnelly, M.A. and Steiner, T.S. (2002) J. Biol. Chem. 277, 40456–40461 9. Ausubel, F.M. (2005) Nat. Immunol. 6, 973–979 10. Zou, H., Henzel, W.J., Liu, X., Lutschg, A. and Wang, X. (1997) Cell 90, 405–413 11. Yue, J.X., Meyers, B.C., Chen, J.Q., Tian, D. and Yang, S. (2012) New Phytol. 193, 1049–1063 12. Leipe, D.D., Koonin, E.V. and Aravind, L. (2004) J. Mol. Biol. 343, 1–28 13. Bent, A.F. and Mackey, D. (2007) Annu. Rev. Phytopathol. 45, 399–436 14. Jacob, F., Vernaldi, S. and Maekawa, T. (2013) Front. Immunol. 4, 297 15. Batut, J., Andersson, S.G. and O’Callaghan, D. (2004) Nat. Rev. Microbiol. 2, 933–945 16. Saldias, M.S., Lamothe, J., Wu, R. and Valvano, M.A. (2008) Infect. Immun. 76, 1059–1067 5 October 2014 © Biochemical Society 17. Limpens, E., Ivanov, S., van Esse, W., Voets, G., Fedorova, E. and Bisseling, T. (2009) Plant Cell 21, 2811–2828 18. Vaishnava, S., Yamamoto, M., Severson, K.M. et al. (2011) Science 334, 255–258 19. Salzman, N.H., Hung, K., Haribhai, D. et al. (2010) Nat. Immunol. 11, 76–83 20. Couillerot, O., Prigent-Combaret, C., Caballero-Mellado, J. and MoënneLoccoz, Y. (2009) Lett. Appl. Microbiol. 48, 505–512 21. Kim, Y.C., Leveau, J., McSpadden Gardener, B.B., Pierson, E.A., Pierson, 3rd, L.S. and Ryu, C.M. (2011) Appl. Environ. Microbiol. 77, 1548–1555 22. Zipfel, C., Kunze, G., Chinchilla, D., Caniard, A., Jones, J.D., Boller, T. and Felix, G. (2006) Cell 125, 749–760 23. Poltorak, A., He, X., Smirnova, I. et al. (1998) Science 282, 2085–2088 24. Chamaillard, M., Hashimoto, M., Horie, Y. et al (2003) Nat. Immunol. 4, 702–707 25. Willmann, R., Lajunen, H.M., Erbs, G. et al. (2011) Proc. Natl. Acad. Sci. U.S.A. 108, 19824–19829 26. Liu, B., Li, J.F., Ao, Y. et al. (2012) Plant Cell 24, 3406–3419 27. Girardin, S.E., Boneca, I.G., Viala, J., Chamaillard, M., Labigne, A., Thomas, G., Philpott, D.J. and Sansonetti, P.J. (2003) J. Biol. Chem. 278, 8869–8872 28. Miya, A., Albert, P., Shinya, T. et al. (2007) Proc. Natl. Acad. Sci. U.S.A. 104, 19613–19618