Survey
* Your assessment is very important for improving the work of artificial intelligence, which forms the content of this project
Sexually transmitted infection wikipedia , lookup
Brucellosis wikipedia , lookup
Middle East respiratory syndrome wikipedia , lookup
Meningococcal disease wikipedia , lookup
Marburg virus disease wikipedia , lookup
Onchocerciasis wikipedia , lookup
Chagas disease wikipedia , lookup
Schistosomiasis wikipedia , lookup
Surround optical-fiber immunoassay wikipedia , lookup
Creutzfeldt–Jakob disease wikipedia , lookup
Leishmaniasis wikipedia , lookup
Visceral leishmaniasis wikipedia , lookup
Bovine spongiform encephalopathy wikipedia , lookup
Leptospirosis wikipedia , lookup
Eradication of infectious diseases wikipedia , lookup
MATHEMATICAL MODELS OF
PRION PROLIFERATION
Yeast cells infected by prions.
(http://www.mos.org/cst/article/368/7.html)
Prion Proliferation Models Research Team
Meredith Greer (Bates College, Lewiston, Maine, USA)
Hans Engler (Georgetown University, Washington, DC, USA)
Jan Pruss (Martin Luther Universitat, Halle-Wittenberg, Germany)
Laurent Pujo-Menjouet (University of Lyon, Lyon, France)
Gieri Simonett (Vanderbilt University, Nashville, Tennessee, USA)
Christoph Walker (Vanderbilt University, Nashville, Tennessee, USA)
Glenn Webb (Vanderbilt University, Nashville, Tennessee, USA)
Rico Zacher (Martin Luther Universitat, Halle-Wittenberg, Germany)
Transmissible Spongiform Encephalopathies (TSEs)
TSEs are diseases such as Creutzfeld-Jakob disease in humans,
scrapie in sheep, and bovine spongiform encephalopathies in cows.
These diseases are characterized by long incubation periods, lack
of immune response, and invisibility to detection as viruses.
In 1982 Stanley Prusiner postulated that these diseases are caused
not by viruses, but by abnormally shaped proteins, which he called
prions. This hypothesis explains many of the features of the
infectious agents of TSEs, except for their ability to replicate.
Prions lack DNA or RNA , which is the commonly accepted basis
for replication. Current research in this field seeks to explain the
mechanism of prion replication.
The nucleated polymerization theory
J. Jarrett and P. Lansbury, Cell, 1993
M. Eigen, Biophys. Chem, 1996
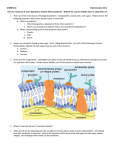
The leading theory of prion replication is nucleated
polymerization. We use the notations for the normal PrPC (prion
protein cellular) and abnormal PrPSc (prion protein scrapies) to
denote the two primary forms of prions. By polymerize we mean
that PrPSc increases its length by attaching units of PrPC in a
string-like fashion.
After a monomer attaches to the polymer, it is converted to the
infectious form. Once the PrPSc is long enough to wrap into a
helical shape (the nucleus), it forms stabilizing bonds that
constitute the polymer strings. These strings can be formed into
lengths of thousands of monomer units.
Replication of prion polymers by splitting
PrPSc polymers may split into two smaller polymers, which
results in two infectious polymers capable of further
lengthening. If after splitting, a smaller polymer falls below the
critical size, however, it degrades immediately into normal
PrPC monomers. The biological processes are
(1) lengthening (by addition of PrPC monomers),
(1) splitting (into two smaller polymer lengths), and
(2) degradation (by metabolic processes)
An infinite system of ODE model
J. Masel, V. Jansen, M. Nowak, Biophys. Chem. 1999
V (t) population of PrP C monomers at time t
ui (t) population of PrP Sc polyomers of length i at time t
U(t)
in0 ui (t), where n is the minimum polymer length
0
d
V (t) V (t) V (t)U(t) 2
dt
i1 ji1i u j (t)
d
ui (t) V (t)(u (t) ui (t)) ui (t)
i1
dt
(i 1) ui (t) 2
ui (t) 0 for i n
0
ji1 u j
for i n
0
A model with continuous polymer length
V(t) = population of normal PrPC monomers at time t
u(x,t) = density of polymers at time t w.r.t. length x in
(x0, ), (where x0 > 0 is the minimum length)
Let
U(t) = total polymer population at time t.
U(t)
u(x,t)dx
x0
Schematic diagram of the model
QuickTime™ and a
TIFF (LZW) decompressor
are needed to see this picture.
Dynamics of the monomer population
x0
d
dt
V (t) V (t) V (t) U (t) 2 x (y )k (x, y) u(y,t)dydx
0
x0
= background source of monomers
= degradation rate of monomers
= conversion rate of monomers to polymers
(y) = rate of splitting of monomers to polymers
k(x,y) = probability that a polymer of length y
splits to lengths x and y-x
Dynamics of the polymer population
u(x,t) V (t) u(x,t)
t
x
(x) u(x,t) (x)u(x,t) 2 (y) k (x,y) u(y,t)dy
x
x 0 x , t 0
(x) degradation rate of polymers
k(x,y)dx 0
0
if y x 0 and
k(x,y)dx 1
0
if y x 0
Equations of the model
x0
(1) dtd V (t) V (t) V (t) U (t) 2 x (y )k (x,y) u(y,t)dydx
0
x0
(2) V (0) V0
(3)
u(x,t) V(t) u(x,t)
t
x
(x) u(x,t) (x)u(x,t) 2 (y) k (x,y) u(y,t)dy
x
(4) u(x,0) (x), x 0 x
(5) u(x 0 ,t) 0, t 0
where U (t)
u(x,t)dx
x0
Assumptions on the parameters
(x) x (the rate of splitting is proportional to polymer length
x)
1
k (x, y)
if 0 x y and x y, and k (x, y) 0 if y x or
y
y x. There is an equal probability of a polymer of length
y splitting
to any shorter length x < y (with the other piece having length y x).
Observe
0
0
k (x, y)dx 0 if y x
0
0
and
0
y
k (x, y)dx
0
1
dx = 1 if x 0 y.
y
An associated system of ODEs
d
2
V (t) V (t) V (t)U(t) x U(t)
0
dt
d
U(t) P(t) U(t) 2 x U(t)
0
dt
d
2
P(t) V (t)U(t) P(t) x U(t)
0
dt
where
V (t) total population of monomers at time
t
U(t)
u(x,t)dx total population of polymers at time
x0
P(t)
xu(x,t)dx total population of monomers in the
x0
polymers at time
t
t
Steady states for the associated system of ODEs
The disease free steady state:
V
, U 0, P 0
The disease steady state:
V
(x 0 )2
(x 0 )2
U
(2x 0 )
(x 0 )2
P
Linearization about the disease-free steady state
The linearization about the disease free steady state V = /, U = 0, P = 0
is
2x 0
0
2
x 0 / 0
2
x
/
0
0
The eigenvalues are
,(x )
0
/ ,(x0 )
/
Theorem. The steady state V = /, U = 0, P = 0 is locally exponentially
asymptotically stable if
/ x 0
Linearization about the disease steady state
V
(x 0 )2
(x 0 )2
U
(2x 0 )
(x 0 )2
P
which exists in the positive cone if
/ x 0
The inearization about the disease steady state is
2x 0
0
(2x )
2
(x
)
0
0
(2x 0 )
(x 0 )2
(2x 0 )
(2x 0 )
0
The eigenvalues of the linearization satisfy the characteristic
equation
z3 a1z 2 a2 z a3 0
where
x 0 2 2 ( 4 ) 6x 0 2 2 3
a1
(2x 0 )
2 (x 0 ) (x 0 )
a2
(2x 0 )
2
a3 (x 0 )2
By the Ruth-Hurwitz condition the eigenvalues all have negative real
parts iff
a1 0, a3 0, and a1 a2 a3.
Theorem. The prion disease steady state is locally exponentially
asymptotically stable if
/ x 0
A general model of infection dynamics
dx
z x
dt
dy
y x y x
dt
dz
x yz
dt
Theorem. Let , , > 0 and [ 0, ). If (x(0), y(0), z(0)) 3 ,
then the solution
to the initial value problem exists in
t 0. If , then the (disease free) equilibrium
3 for all
(0, ,0)is
globally asymptotically stable (exponentially if the inequality is
( )
strict). If , then the (disease) equilibrium (
, ,
)
is globally exponentially asymptotically stable.
Lyapunov functionals
If < , then a Lyapunov functional for disease free equilibruim is
1
(x, y,z) (y ) 2 (2 )(x z)
2
If > , then a Lyapunov functional for disease equilibruim is
x
y
(x, y,z) (x x x log ) (y y y log )
x
y
(z z z log )
(y y log y)
z
z
( )
where (x, y,z ) (
,,
).
Application to the prion ODE system
d
dt
V (t) V (t) V (t)U (t) x 0 U (t)
d
dt
U (t) P(t) U (t) 2 x 0U (t)
d
dt
P(t) V (t)U (t) P(t) x 0 2U (t)
2
Theorem. Let (V,U,P) :V 0, U 0, P x U . If
0
(V (0),U(0), P(0)) , then the solution to the initial value
problem exists in for all t 0. If / x ,
0
then the disease free steady state is globally asymptotically
stable in (exponentially if the inequality is strict)
. If
/ x 0 , then the disease steady state is globally
exponentially asymptotically stable in
.
Convergence to the disease steady state
The parameters are taken from J. Masel, V. Jansen, M. Nowak, Biophys.
Chem. 1999 and R. Rubenstein et al., J. Infect. Dis. 1991. 4400 ,0.3,
5.0,.0001,0.04,x06
QuickTime™ and a
TIFF (LZW) decompressor
are needed to see this picture.
Phase portrait of V(t), U(t), and P(t)
All solutions converge to the disease steady state (V,U,P) = (55, 250, 103,132).
Application to a model of HIV infection
D. Ho et al., Rapid turnover of plasma virions and CD 4
lymphocytes in HIV-1 infection, Nature 1995, and M. Nowak and
R. May, Virus Dynamics, 2000
I(t) = infected CD4+ T cells at time t
T(t) = uninfected CD4+ T cells at time t
V(t) = virus at time t
d
I(t) V (t)T(t) I(t)
dt
d
T(t) T(t) V (t)T(t)
dt
d
V (t) N I(t) V (t)
dt
Asymptotic behavior of the model of HIV infection
Let R0 = N / . R0 is the number of secondary infections
produced by one infectious virus in a totally susceptible
population.
If R0 < 1, then all solutions converge to the disease free steady
state
Tss =/, Iss = 0, Vss = 0.
If R0 > 1, then all solutions converge to the disease steady state
Tss
,
N
I ss
0
R 1
,
N
Vss
0
R 1
The case R0 < 1
Let = .01, = 10, =
10-4.1, N = 250, = .5,
= 24.
R0 = .827.
All solutions converge to
the disease free steady
state
Tss=1000, Iss = 0, Vss = 0.
The case R0 > 1
Let = .01, = 10, = 10-4.1,
N = 1000, = .5, = 24.
R0 = 3.31.
All solutions converge to the
disease steady state
Tss =302, Iss = 14, Vss = 291.
Application to an SEIS epidemic model
S(t) = susceptible population at time t
E(t) = exposed population at time t (infected but not
yet infectious)
I(t) = infectious population at time t
d
S(t) S(t) I(t) S(t) I(t)
dt
d
E(t) I(t) S(t)( ) E(t)
dt
d
I(t) E(t)( ) I(t)
dt
Asymptotic behavior of the SEIS epidemic model
Let R0 = / [].
R0 is the number of secondary infections produced by one infective
in a totally uninfected susceptible population.
If R0 < 1, then all solutions converge to the disease free steady state
Sss =/, Ess = 0, Iss = 0.
If R0 > 1, then all solutions converge to the disease steady state
Sss
.
( )( )
, Ess
( )( )
, Iss
( )
The case R0 < 1
Let = 0.05, = 500,
=0.1, 10-6.9, =0 .2, =
.0003, .1.
R0 = .933.
All solutions converge to the
disease free steady state
Sss =1,666,667, Ess = 0, Iss =0.
The case R0 > 1
Let = .05, = 500, =.1,
10-6.5, = .2, = .0003,
.1.
R0 = 2.34.
All solutions converge to
the endemic steady state
Sss =711,512, Ess = 1228,
Iss = 1637.
Analysis of the prion PDE model
d
2
V (t) ( U(t)) V (t) x U(t)
0
dt
(2) V (0) V
(1)
0
(3)
u(x,t) V (t) u(x,t)
t
x
( x) u(x,t) 2
(4) u(x,0) (x), x x
0
(5) u(x ,t) 0, t 0
0
xu(y,t)dy
The disease steady state problem for the PDE model
(1) ( U ) V x 0 2U 0
(2) V u(x) ( x) u(x) 2 u(y)dy, x x 0
x
(3) u(x 0 ) 0
where V 0, u(x) 0, and U
u(x)dx
x0
x 0 2U
Solve (1) to obtain V
U
Then use (2) and (3) to show that u(x) satisfies
( x)( U )
3 ( U )
u(x)
u
(x)
u(x) 0, x x 0
2
2
( x0 U )
( x0 U )
u(x 0 ) 0
2 U ( U )
u(x 0 )
( x 0 2U )
Solution of the disease steady state problem
Since the value for
U at the disease steady state is
(x0 ) 2
U
,
(2x 0 )
then the disease equilibrium
u(x) must satisfy
( x)
3 2
(1) u(x)
u(x)
u(x) 0, x x 0
( x 0 )2
( x 0 ) 2
(2) u(x 0 ) 0
2 2 ( ( x 0 ) 2 )
(3) u(x 0 )
.
( x 0 ) 2 ( 2 x 0 )
Theorem. Let ( x 0 )2 . The unique solution of (1), (2),(3) is
u(x)
e
- (x -x 0)( 2 + (x +x 0))
2( + x0 ) 2
2 (x x 0)(2 (x x 0))( ( x 0) 2 )
.
3
( + x 0) ( + 2x 0)
Analysis of the PDE model
Theorem. Let Z + = + L1 ((x ,); xdx). The model generates a global
0
semiflow in Z +. If / ( x ) 2 , then the disease free equilibrium
0
( / ,0) is globally asymptotically stable,
case of strict inequality. If
and even exponentially in the
/ ( x ) 2, then the unique disease
0
V *,u*(x)
equilibrium is globally asymptotically stable in
where
Z + \ + {0},
V * ( + x ) 2 / ,
0
*
u (x)
e
- (x-x0)(2 + (x+x0))
2( + x0)2
(x x )(2 (x x ))( ( x ) )
.
( + x ) ( +2x )
2
2
0
0
3
0
0
0
Ideas of the proof
(1) The solution V(t) can be considered known. Let w(t) = V(t). w(t)
converges exponentially to w* = / in the disease free case and to w* =
( x0)2/ in the disease case.
(2) First consider the autonomous equation for u(x,t), where w(t) = w*. Prove
that that there is a strongly continuous, linear, positive, contraction
(exponentially in the disease free case) semigroup e-t L, t > 0 in the space
X = L1((x0, );x dx) associated with the autonomous equation.
(3) Prove that the resolvent of L is compact in X, and thus has only point
spectrum in the closed right-half plane. Show that 0 is the only eigenvalue
of L on the imaginary axis, it is simple, the ergodic projection P onto the
kernel on N(L) of L along the range R(L) of L exists and is rank one, find
a formula for P, and prove that e-t L converges strongly to P in X.
(4) Use the method of characteristics to prove that the nonautonomous
equation for u(x,t) is well-posed, obtain bounds for ux(.,t) in X, and use the
convergence of w(t) to w* to show that u(.,t) converges in X to the
equililbrium u*.
Model simulation compared to experimental data of
scrapie associated fibril counts
SAF measurements after intracerebral injection of the 139A scrapie strain into
Compton white mice from Rubenstein et al., J. Infect. Dis., 1991. The arrow
indicates the onset of symptoms. The parameters of the simulation are 4400 ,
0.3,5.0,.0001,0.04,x06.
QuickTime™ and a
TIFF (LZW) decompressor
are needed to see this picture.
Evolution of the
polymer density u(x,t)
QuickTime™ and a
TIFF (LZW) decompressor
are needed to see this picture.
QuickTime™ and a
TIFF (LZW) decompressor
are needed to see this picture.
The polymer density u(x,t)
converges to the steady state.
Evolution of the mean length P(t)/U(t) of the polymer density
QuickTime™ and a
TIFF (LZW) decompressor
are needed to see this picture.
The length of the average polymer u(x,t) increases fast initially and then
slowly decreases due to the depletion of the PrPC monomer population.
Linear dependence on
the log scale of the
incubation times vs the
inoculum dose
QuickTime™ and a
TIFF (LZW) decompressor
are needed to see this picture.
QuickTime™ and a
TIFF (LZW) decompressor
are needed to see this picture.
The linear dependence of
the incubation times on the
log scale for nine orders of
magnitude of inoculum
dilutions.
More general models
(1) Allow the parameters and to depend on polymer length x.
(2) Allow the splitting kernel k(x,y) to have a more general form
Hypothesis :
(x) and (x) L ((x ,);) and k (y, x) 0is measurable
0
existence of unique strong solution
Hypothesis :
there exists 1 and L ((x 0 ,);) such that
(x) 0 as x and (x) (x) (x) x a.e. x (x ,),
for each > 0 there exists > 0 such that
(x) x
sup
(y) k (y, x) dy a.e. x (x ,)
||
0
x
x
0
existence of weak solution
0
References
H. Engler, J. Pruss, and G. Webb, Analysis of a model for the dynamics of prions II, to appear in
J. Math. Anal. Appl.
M. Greer, L. Pujo-Menjouet, and G. Webb, A mathematical analysis of the dynamics of prion
proliferation, to appear.
J. Masel, V. Jansen, and M. Nowak, Quantifying the kinetic parameters of prion replication,
Biophysical Chemistry 77 (1999) 139-152.
Nowak, M., et al. Prion infection dynamics, Integrative Biology 1 (1998) 3-15.
Prusiner, S. Molecular biology of prion diseases, Science 252 (1991) 1515-1522.
J. Pruss, L. Pujo-Menjouet, G. Webb, and R. Zacher, Analysis of a model for the dynamics of
prions, to appear in Discr. Cont. Dyn. Sys.
Rubenstein, R. et al., Scrapie-infected spleens: analysis of infectivity, scrapie-associated fibrils,
and protease-resistant proteins, J. Infect. Dis. 164, (1999) 29-35.
Simonett, G. and Walker, C., On the solvability of a mathematical model of prion proliferation,
to appear.