Survey
* Your assessment is very important for improving the workof artificial intelligence, which forms the content of this project
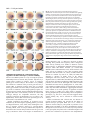
3899 Development 127, 3899-3911 (2000) Printed in Great Britain © The Company of Biologists Limited 2000 DEV3249 Maternally controlled β-catenin-mediated signaling is required for organizer formation in the zebrafish Christina Kelly*, Alvin J. Chin*, Judith L. Leatherman, David J. Kozlowski and Eric S. Weinberg‡ Department of Biology, The University of Pennsylvania, Philadelphia, PA 19104, USA *These two authors contributed equally to the work ‡Author for correspondence ([email protected]) Accepted 2 July; published on WWW 22 August 2000 SUMMARY We have identified and characterized a zebrafish recessive maternal effect mutant, ichabod, that results in severe anterior and dorsal defects during early development. The ichabod mutation is almost completely penetrant, but exhibits variable expressivity. All mutant embryos fail to form a normal embryonic shield; most fail to form a head and notochord and have excessive development of ventral tail fin tissue and blood. Abnormal dorsal patterning can first be observed at 3.5 hpf by the lack of nuclear accumulation of β-catenin in the dorsal yolk syncytial layer, which also fails to express bozozok/dharma/nieuwkoid and znr2/ndr1/squint. At the onset of gastrulation, deficiencies in expression of dorsal markers and expansion of expression of markers of ventral tissues indicate a dramatic alteration of dorsoventral identity. Injection of β-catenin RNA markedly dorsalized ichabod embryos and often completely rescued the phenotype, but no measurable dorsalization was obtained with RNAs encoding upstream INTRODUCTION The dorsal signaling center known as the ‘organizer’ was first recognized in amphibians by Spemann and Mangold (1924) by its ability to induce a second body axis when transplanted to a second embryo. This signaling center also exists in other vertebrate embryos – the teleost ‘embryonic shield’, the avian ‘Hensen’s node’ and the mammalian ‘node’ (reviewed in Harland and Gerhart, 1997). In amphibians, induction of mesoderm and formation of the organizer is dependent on the activity of an earlier vegetal signaling site, often called the Nieuwkoop center (Nieuwkoop, 1969), which is directly dependent on maternal gene activity. Establishment of the Nieuwkoop center requires cortical rotation of the fertilized egg, resulting in a redistribution of maternal determinants (Gerhart et al., 1989; reviewed in Moon and Kimelman, 1998). In Xenopus, formation of the organizer is dependent on at least two pathways involving maternally encoded products. The T-box family transcription factor, VegT, is required for the vegetal-derived signaling that converts cells of the equatorial zone into mesoderm (Zhang et al., 1998), and β-catenin is required for embryonic axis formation (Heasman et al., 1994; Wnt pathway components. In contrast, dorsalization was obtained when RNAs encoding either Bozozok/Dharma/ Nieuwkoid or Znr2/Ndr1/Squint were injected. Moreover, injection of β-catenin RNA into ichabod embryos resulted in activation of expression of these two genes, which could also activate each other. RNA injection experiments strongly suggest that the component affected by the ichabod mutation acts on a step affecting β-catenin nuclear localization that is independent of regulation of β-catenin stability. This work demonstrates that a maternal gene controlling localization of β-catenin in dorsal nuclei is necessary for dorsal yolk syncytial layer gene activity and formation of the organizer in the zebrafish. Key words: β-catenin, Maternal effect, Axis formation, Dorsoventral patterning, Spemann organizer, Nieuwkoop center, Zebrafish, Wnt signaling, Nuclear localization Wylie et al., 1996, reviewed in Sokol, 1999). Depletion of maternal RNA encoding these products results, respectively, in the inability to form mesoderm and the failure to develop axial structures (Heasman et al., 1994; Zhang et al., 1998). In both cases, the Spemann organizer fails to form. Depletion of Xenopus maternal RNA encoding the GSK3-binding protein GBP also causes ventralization (Yost et al., 1998). Since GBP inhibits GSK3 by preventing Axin from binding GSK3 (Farr et al., 2000), thereby resulting in stabilization of β-catenin (Yost et al., 1998), this experiment also provides evidence for maternal factors that regulate β-catenin-mediated dorsal signaling. In frogs, two changes in protein levels have been reported after cortical rotation on the prospective dorsal side in a microtubule-dependent manner: (1) Disheveled accumulates directly after cortical rotation, indicating the involvement of a maternal Wnt pathway in dorsal cell fate establishment (Miller et al., 1999), and (2) GSK3 levels decrease in the dorsal region in early cleavage stages, possibly by a non-Wnt pathway signal mediated by GBP (Dominguez and Green, 2000). Zebrafish and Xenopus share many of the same mechanisms involved in formation of the organizer and dorsal axis. As in frogs, the formation of the zebrafish body axis is dependent on 3900 C. Kelly and others cortical arrays of microtubules (Jesuthasan and Strähle, 1997), which presumably facilitate the cytoplasmic rearrangements required for formation of the yolk syncytial layer (YSL). Additionally, nuclear β-catenin accumulates on the dorsal side of both zebrafish and Xenopus embryos (Schneider et al., 1996). In zebrafish, this nuclear localization is first observed in the dorsal YSL and then later in overlying blastoderm cells, suggesting that the dorsal YSL may be the functional equivalent of the Xenopus Nieuwkoop center (Schneider et al., 1996). Overexpression of β-catenin in zebrafish embryos can induce a secondary axis (Kelly et al., 1995a), as it does in Xenopus (Funayama et al., 1995; Guger and Gumbiner, 1995), indicating a conserved response pathway to β-catenin in these organisms. Further evidence of Nieuwkoop center activity of the zebrafish YSL is its ability to induce and pattern mesoderm after transplantation to the animal pole region of an intact blastula (Mizuno et al., 1996) or in a conjugate with an animal cap (Ober and Schulte-Merker, 1999). Two proteins, the homeodomain transcription factor Dharma/Nieuwkoid (Yamanaka et al., 1998; Koos and Ho, 1998; Fekany et al., 1999) and Nodal-related Znr2/Ndr1 (Erter et al., 1998, Feldman et al., 1998) have recently been shown to act non-cell autonomously in the YSL to induce goosecoid, a marker of the organizer. Dharma/Nieuwkoid is encoded by bozozok (Fekany et al., 1999), which when mutated leads to a ventralized phenotype characterized by loss of notochord, prechordal plate, and anterior and ventral CNS deficiencies (SolnicaKrezel et al., 1996). Znr2/Ndr1 (Erter et al., 1998; Rebagliati et al., 1998) is encoded by the squint gene (Feldman et al., 1998), which when mutated results in ventral CNS defects and cyclopia. Embryos doubly homozygous for mutations in squint and cyclops (another nodal-related gene) completely lack trunk mesoderm and endoderm (Feldman et al., 1998). Normally, LiCl-treated early blastulae are dorsalized by inhibition of GSK3 (Hedgepeth et al., 1997). Thus, the observations that LiCl can enhance bozozok/dharma/nieuwkoid expression (Yamanaka et al., 1998) and that LiCl enhancement of goosecoid expression is blocked in bozozok embryos (Fekany et al., 1999) are suggestive that bozozok/dharma/nieuwkoid is a downstream target of the β-catenin/Tcf pathway in the YSL. Also in support of this relationship, the dharma/nieuwkoid promoter has been reported to contain Tcf/Lef-binding sites (Yamanaka et al., 1998), and nuclear localization of β-catenin is observed in bozozok mutant embryos (Fekany et al., 1999). β-catenin was originally shown to be required for axis formation in Xenopus (Heasman et al., 1994; Wylie et al., 1996), and it was recently established that mice lacking functional β-catenin (Huelsken et al., 2000) or Wnt3 (Liu et al., 1999b) genes also fail to form a normal primary axis. We report here the characterization of a zebrafish maternal effect mutant that severely affects formation of the organizer and subsequent development of dorsal and anterior tissues. We find that mutant embryos fail to accumulate β-catenin in YSL and marginal blastoderm nuclei, providing strong evidence for a requirement for β-catenin-mediated signaling in the zebrafish. Mutant embryos can be dorsalized and rescued by RNAs encoding β-catenin and the downstream proteins Znr2/Ndr1/ Squint and Bozozok/Dharma/Nieuwkoid, indicating that the axis specification pathway downstream of β-catenin is intact in ichabod mutant embryos. However, since injection of RNAs encoding proteins that inhibit GSK3 activity or that increase βcatenin stability fail to dorsalize ichabod embryos, the ichabod gene product most likely functions to ensure nuclear localization of β-catenin, in parallel to the control of β-catenin stability, rather than directly on modulating GSK3 activity or levels. MATERIALS AND METHODS Animals Stocks of Danio rerio were maintained and raised under standard conditions at 28.5°C (Westerfield, 1993). The flhn1 stock that proved to harbor the ichabod mutation was obtained from Dr C. Kimmel of the University of Oregon, and then inbred for several generations in our laboratory prior to the identification of the ichabod phenotype. brass zebrafish, particularly suitable for studies involving in situ hybridization because of delayed and reduced pigmentation and used in this study as ‘wild-type’ in comparisons with ichabod embryos, were originally obtained from EkkWill Waterlife Resources, Gibbonston, FL. DNA constructs The following zebrafish probes were used for in situ hybridization: znr2 (squint/ndr1) (Erter et al., 1998; Rebagliati et al., 1998), nieuwkoid (bozozok/dharma) (Koos and Ho, 1998), goosecoid (Stachel et al.,1993), chordin (Miller-Bertoglio et al., 1997), Otx1 (Li et al., 1994), lim1 (Toyama et al., 1995), bmp4 (Chin et al., 1997), no tail/Brachyury (Schulte-Merker et al., 1992), eve1 (Joly et al., 1993) and gata2 (Detrich et al., 1995). The following constructs were used to prepare RNA for injection: zWnt8 in pT7TS (Kelly et al., 1995b), zFzA in pT3TS (Nasevicius et al., 1998), XDsh in Sp64T (Sokol et al., 1995), Xβ-catenin in pCS2 (modified from Funuyama et al., 1995), zβ-catenin in pZL1 (Kelly et al., 1995a), dominant negative (dn) GSK3 (Xgsk-3K85R) in pCS2 (Pierce and Kimelman, 1995), nieuwkoid in pGEM (Koos and Ho, 1998), znr2/squint in pBS(SK−) (Erter et al., 1998), zebrafish GBP in pCS2+ (Sumoy et al., 1999), Xenopus Axin GID 2 fragment in pCS2MT (Hedgepeth et al., 1999), and Xenopus β-Trcp ∆F in pCS2+MT (Liu et al., 1999a). RNA injections Capped mRNAs were synthesized from linearized plasmid DNA either by using 50 units of the appropriate RNA polymerase (T7, T3, or Sp6) in the presence of 12.5mM m7G (5′)pppGTP cap structure analog (New England Biolabs), or by using the protocol for the mMessage mMachine Kit (Ambion). RNA was diluted 1:1 with 0.5% phenol red/PBS (Sigma) and injected into either the yolk beneath the blastomeres of 1- to 4-cell-stage embryos (for β-catenin RNAs) or into a single blastomere of 8- to 16-cell-stage embryos (for all other RNAs used). Final concentrations of injected RNAs varied from 50-1200 ng/µl, depending on the particular RNA. Embryos were injected with approximately 1 nl of RNA solution (see Fig. 7 for the amount of RNA injected for each species of RNA), containing a final concentration of 0.25% phenol red. In situ hybridization, immunohistochemistry and microscopy In situ hybridization was performed by a modified version (Li et al., 1994) of the protocol of Schulte-Merker et al. (1992). β-catenin was detected immunologically with antibody against Xenopus β-catenin (Schneider et al., 1996) using embryos fixed in 4% paraformaldehyde in PBS. Immunohistochemical staining was carried out after incubation with secondary antibody using preincubated AB complex (ABC Kit, Vector), DAB substrate and hydrogen peroxide. Whole embryos were observed and photographed under a Leica MZ12 stereo dissecting microscope. Maternal control of the zebrafish organizer 3901 RESULTS ichabod is a maternal effect mutation affecting anterior and dorsal development While inbreeding a strain of zebrafish containing the zygotic recessive mutation floating head (flh) (Talbot et al., 1995), we found six female fish that yielded embryos of unexpected phenotype. These embryos lacked anterior structures and notochords and had excessive development of ventral tail fin tissue and blood. Most commonly, the embryos completely lacked heads, and almost all of those embryos that did develop heads had defective forebrains. When these female fish were then bred to wild-type or brass males, batches of embryos entirely abnormal in phenotype were obtained, a result not expected for the zygotic recessive flh mutation. Although failure to form a notochord is a characteristic of flh homozygotes, the other phenotypic characteristics have not been reported for flh. The presence of a strict maternal effect mutation unrelated to flh was suggested by two initial findings: (1) the abnormal embryo phenotype was obtained irrespective of the male parent used in crosses with these six fish, even when the females were outcrossed to wild-type fish, and (2) 99-100% of the embryos from these crosses were abnormal. We set up a breeding scheme to determine if the phenotype could be transmitted genetically, to test whether it was linked to the flh mutation, and to discover whether the putative mutation was dominant or recessive in the female parent (Fig. 1). The six females that transmitted the unusual phenotype (which we denoted ichabod [ich]) were part of a large brood of fish from a single tank that had originally been derived from two pairs of fish (G0 parents), heterozygous for the flh mutation. Since we had not sequestered these four parents, we continued the line by breeding the siblings (F1 siblings) of the six ichabod transmitting females (these original six females could not be used for further genetic analysis since they yielded only lethal embryos). Our reasoning was that, if the trait were recessive, siblings should include females and males heterozygous for the ichabod mutant allele as well as homozygous males, and these siblings should be able to transmit the gene. Alternatively, if the trait were a maternal effect dominant, it would be recovered only when females homozygous for the wild-type allele were crossed with male carriers. We started over 50 F2 families by breeding 50 sibling pairs of fish (F1 siblings). In seven of these families, we were able to recover F2 females that transmitted the ichabod phenotype to their progeny (F3 embryos). In all cases tested, the transmission of ichabod phenotype was observed irrespective of the male to which the females were crossed, confirming that ichabod is a maternal effect mutation. A dominant mode of inheritance would be consistent only with ratios of 1/2 or 1/1 of the F2 females within a particular family yielding the ichabod phenotype. If the mutation were inherited as a dominant trait, a ratio of 1/4 ichabod transmitting females would be impossible to obtain since the female F1 parent would have had to be homozygous for the wild-type allele. A recessive mode of inheritance is consistent with ratios of 1/4 or 1/2 of the F2 females within a particular family being ichabod transmitting fish. In this case, the F1 female parent would be heterozygous and the F1 male parent could be either heterozygous or homozygous for the mutant ichabod allele. Fig. 1. ichabod is a recessive maternal-effect mutation. (Top) Siblings of females that gave rise to ichabod phenotype embryos were used to generate families to test for the mode of inheritance of the trait. A brood of F1 fish, bred from two pairs of flh heterozygote G0 parents, contained six females that transmitted the ichabod phenotype to >99% of their progeny. Pairs of F1 siblings of these six transmitting females were used to generate families of F2 fish. (Bottom) Sibling matings were carried out within each family (e.g., 22 pairwise crosses were performed with progeny of F1 pair no. 2). Matings were also performed between F2 females of family no. 2 and F2 males of family no. 33. In 7 of 33 families, we were able to recover F2 females that transmitted the ichabod phenotype to their progeny (F3 embryos). Ratios of ichabod-transmitting females in four of the families generated from crosses of F1 siblings are presented. The number of F2 females obtained in the other three families was too low (<6) to be informative to analyze the mode of inheritance. In two of the families (no. 7, no. 33), and in a subsequent cross of F1 female no. 2 with male no. 33, the ratios of affected families were consistent only with a recessive maternal effect mode of inheritance. The ratio (A/B) is the number of F2 females that transmitted the ichabod trait (A) divided by the total number of F2 females tested for that family (B). A χ2 test for significance of the ratio of 1/4 gave high consistency for crosses within family no. 7 (P=0.65, χ2=0.2), within family no. 33 (P=0.4, χ2=0.79), and between female no. 2 and male no. 33 (P=0.63, χ2=0.385); but high inconsistency for the significance of a ratio of 1/2 for crosses within family no. 7 (P=0.02, χ2=5.4), within family no. 33 (P<0.0001, χ2=16), and between female no. 2 and male no. 33 (P=0.0015, χ2=10.28). The mutation described here has received the allelic assignment p1 (i.e., ichabodp1). The table in Fig. 1 shows the ratios of ichabod transmitting females in four of the families generated from crosses of siblings of the original F1 fish (in three other ichabod transmitting families, we obtained only two to six adult females, a number too low to be informative to analyze the mode of inheritance). In one family (no. 2), the ratio of transmitting females was close to 1/2, and thus was not informative in distinguishing between recessive and dominant 3902 C. Kelly and others Fig. 2. ichabod embryos develop anterior and dorsal deficiencies with variable expressivity. Although greater than 99% of the embryos bred from ichabod-transmitting females were larval or embryonic lethal, the phenotypes of affected embryos were variable. Characteristics of classes 1-4 are described in the text. modes of inheritance. In another family (no. 28), 26 adult females were obtained but the ratio of transmitting females was intermediate between 1/1 and 1/2. However, in two of the families (no. 7, no. 33), and in a subsequent cross of F1 female no. 2 with male no. 33, we obtained sufficient numbers of fish and highly statistically significant ratios to establish that the ratios of affected families were consistent only with a recessive mode of inheritance. By following the transmission of ichabod and flh in the families generated from crosses of F1 fish, we were able to show that the two mutations segregated independently, and were thus unlinked (data not shown). In addition, we have recently shown that the ichabod locus is on zebrafish linkage group 19 (C. K. and E. S. W., unpublished results), ruling out ichabod as an allele of flh, which is located on linkage group 13 (Talbot et al., 1995). The map positions of bozozok and dino (two other ventralized mutants) on linkage group 15 (bozozok, Fekany et al., 1999; dino, Fisher et al., 1997) rules out ichabod as an allele of these genes. Penetrance and expressivity of the ichabod mutation As expected for a maternal effect mutation, over 98% of the embryos bred from homozygous ichabod females exhibited an embryonic mutant phenotype and over 99% were embryonic or larval lethal (7849 embryos were scored in 65 crosses). Less than 1% survived and were ‘escapers.’ There is variable expressivity of the mutation and the phenotypes have been classified into the following groups (Fig. 2). Class 1 (34%) are headless embryos lacking a notochord, and are also deficient in trunk development; most embryos in class 1 develop an excess of blood, have large posterior somites and form multiple tail fin structures (Fig. 2A,B), but some appear completely radialized with little trunk development. Class 2 (30%) have defects similar to class 1, but do form a trunk and at least some hindbrain is present (Fig. 2C). Class 3 (28%) lack notochord, have some forebrain and/or midbrain development but are deficient in anterior forebrain and are either cyclopic or completely lack eyes (Fig. 2D). Class 4 (6%) lack notochord, but have recognizable forebrain, midbrain and hindbrain structure, and eyes (Fig. 2E). Class 5 (2%) are indistinguishable from wild type, but are larval lethal (not shown in Fig. 2). This phenotypic series is similar to that obtained by injection of an RNA encoding dominant negative Tcf3 (Pelegri and Maischen, 1998; Sumoy et al., 1999) or by removal of vegetal yolk (Mizuno et al., 1999). The ratio of phenotypes that we obtained varied in repeated crosses of the same parental pair. Generally, young homozygous female parents (3-4 months of age) yielded more severe phenotypes. As these fish aged, milder embryo phenotypes were seen at an increased frequency (data not shown). A similar observation, that severity of embryonic phenotype declines with maternal age, was reported for the janus mutation, which results in a splitting of the embryo during cleavage stage (Abdelilah and Driever, 1997). ichabod embryos are deficient in formation of the organizer Investigation of the morphological development of affected embryos shows that they are deficient in the formation of the embryonic shield (Fig. 3). As the wild-type embryo begins gastrulation at 5-5.5 hpf, a thickening of the germ ring is seen on the future dorsal side (Fig. 3A). This thickening becomes the site of the initial involution of mesodermal and endodermal cells and persists for several hours (Fig. 3B). In ichabod embryos, a transient slight thickening is seen at 5-5.5 hpf (Fig. 3C), but within 30 minutes, the germ ring is again radially symmetric in thickness, although with a more irregular border (Fig. 3D). Although involution or ingression of mesoderm and endoderm cells does occur, there is no obvious dorsal axial mesoderm thickening. This is the earliest morphological defect seen in ichabod phenotype embryos. Curiously, the lack of a visible shield is observed even in embryos that will go on to Fig. 3. The embryonic shield forms transiently in ichabod embryos. The same wild-type (WT) (A,B) and ichabod (ich) (C,D) embryos were photographed at the 50% epiboly (A,C) and 60% (B,D) epiboly stages. Arrows point to a dorsal thickening of the germ ring, characteristic of the forming embryonic shield. In ichabod embryos of all classes, the thickening was only observed transiently. By 60% epiboly, the germ ring appeared to be of uniform thickness around the circumference of the embryo. Irregularities in the shape of the germ ring border are characteristic of ichabod embryos. Maternal control of the zebrafish organizer 3903 develop the less severe phenotypes (classes 3, 4 and 5). Therefore, a morphologically normal embryonic shield is not required for formation of axial mesoderm and the formation of a complete neural tube (including anterior brain). In most ichabod embryos, however, the absence of a thickened embryonic shield is in fact accompanied by the development of severe anterior and dorsal defects. ichabod embryos have reduced dorsal gene expression and have expanded ventral gene expression domains Since ichabod embryos do not form a normal organizer, it was of interest to determine whether genes usually expressed within the YSL and embryonic shield are active in ichabod embryos. In situ hybridization with probes representing genes with dorsal and ventral expression patterns indicate that there is an early alteration of dorsoventral identity in ichabod gastrula embryos. The earliest changes in gene expression patterns were seen in the dorsal yolk syncytial layer (YSL) in the sphere stage blastulae (4 hpf). Transcripts of bozozok/dharma/nieuwkoid and nodal-related znr2/ndr1/squint, genes normally expressed at this time in the YSL (Fig. 4A,C) (Yamanaka et al., 1998; Koos and Ho, 1998; Feldman et al., 1998; Erter et al., 1998; Rebagliati et al., 1998), were not detected in the YSL or elsewhere in 4 hpf ichabod embryos (Fig. 4B,D). (The later ring of marginal squint expression, however, appeared to be normal in ichabod embryos, data not shown). Most ichabod embryos also do not express goosecoid and chordin, which are expressed in wild-type embryos starting at midblastula and are then characteristically expressed in the embryonic shield region (Fig. 4E,I) (Stachel et al., 1993; Schulte-Merker et al., 1994; Miller-Bertoglio et al., 1997). To determine the distribution of phenotypes in the batch of embryos tested by in situ hybridization, some sibling embryos were always allowed to continue development to 24 hpf at which time the distribution of phenotypic classes was scored. By correlating the percentages of ichabod embryos that develop into the various phenotypic classes with the distribution of in situ hybridization expression patterns, we infer that class 1 and 2 embryos express no detectable goosecoid at the 50% epiboly stage (Fig. 4F), that class 3 embryos express the gene only in a few scattered cells (Fig. 4G), and that class 4 embryos show a disorganized pattern of reduced transcript (Fig. 4H) (similar effects were seen at 80% epiboly, data not shown). In the case of chordin, absence of expression at 50% epiboly was correlated with the frequency of class 1 embryos (Fig. 4J) but, unlike goosecoid, expression of chordin was seen in what are probably class 2 embryos (Fig. 4K), although at low levels. Another group of embryos, probably of class 3 phenotype, expressed more chordin transcript in the embryonic shield region (Fig. 4L), but still at lower than wild-type levels (similar effects were seen at 80% epiboly, data not shown). lim1, which is normally expressed in the marginal cells of the embryo from 30-50% epiboly and at higher levels in the hypoblast at early shield stage (Fig. 4M) (Toyama et al., 1995; Toyama and Dawid, 1997), is expressed in the marginal cells of ichabod embryos, but not in any region suggestive of embryonic shield expression Fig. 4. ichabod embryos are deficient in formation of axial tissues, but have expanded expression of ventral markers. Expression of the following genes was monitored by in situ hybridization as follows: bozozok/dharma/nieuwkoid in wild-type (A) and ichabod (B) 4 hpf embryos; znr2/ndr1/squint in wild-type (C) and ichabod (D) 4 hpf embryo; goosecoid in wild-type (E) and ichabod (F-H) embryos at 50% epiboly; chordin in wild-type (I) and ichabod (J-L) embryos at 50% epiboly; lim1in wild-type (M,O) and ichabod (N,P) embryos at 50% epiboly (M,N) and at 95% epiboly (O,P); Otx1 in wild-type (Q) and ichabod (R) embryos at 80% epiboly; no tail/Brachyury in wild-type (S) and ichabod (T) embryos at 85% epiboly; bmp4 in wild-type (U,W) and ichabod (V,X) embryos at 50% epiboly (U,V) and 70% epiboly (W,X); eve1 in wild-type (Y) and ichabod (Z) embryos at 50% epiboly; and gata2 in wild-type (A′) and ichabod (B′) embryos at 50% epiboly. Embryos are shown in the following orientations: dorsal view with animal pole to the top (A-L, O-T), animal pole view with dorsal to the right (M,N,U,V,Y-B′), and lateral view with dorsal to the right (W,X). In the upper right corner of each panel is indicated whether the embryo is wild type or ichabod. The gene probed for expression is indicated in the lower right corner of each panel. 3904 C. Kelly and others of these two markers was expanded to encompass the whole marginal zone and ectoderm, respectively (Fig. 4Z,B′). The altered expression pattern of these genes in ichabod embryos indicates an expansion of ventral identity to the dorsal side of the embryo. Fig. 5. β-catenin is not localized in nuclei of ichabod embryos. An antibody prepared against Xenopus β-catenin (Schneider et al., 1996) was used to determine the distribution of β-catenin in wild-type (A,B) and ichabod (C,D) embryos at high (3.5 hpf) (A,C) and sphere (4.0 hpf) (B,D) stages. Nuclear localization of the protein was observed in the dorsal YSL of wild-type embryos at both stages and in cells at the dorsal blastoderm margin (arrows in B) at sphere stage. Nuclear localization was not observed in ichabod embryos. Arrows in C indicate YSL nuclei which appear as holes, devoid of β-catenin staining, whereas the irregularly shaped YSL region does show staining. These unstained nuclei are also detected at 4 hpf, but are more difficult to see. (Fig. 4N). At later stages, when the expression is normally confined to posterior axial mesoderm (Fig. 4O), ichabod embryos completely lack expression of the gene (Fig. 4P). Interestingly, the absence of shield expression at 50% epiboly and axial mesoderm expression at 95% epiboly is observed in ichabod embryos of all classes. Similarly, we found an absence of no tail/Brachyury expression in axial mesoderm of all ichabod embryos, although expression of this gene at the margin is present in all ichabod embryos and is even more pronounced than in wild-type embryos (Fig. 4S,T). Otx1 (Fig. 4Q,R) and Otx2 (data not shown) expression at 80% epiboly, which marks the future forebrain and midbrain in wild-type embryos (Li et al., 1994), was absent in class 1 and class 2 embryos (Fig. 4R), and diminished in class 3 phenotypes (data not shown). These studies of gene expression are consistent in indicating the absence of a normal organizer, a lack of axial mesodermal tissue and a failure to induce anterior neurectoderm in ichabod embryos. We also studied the expression of a number of ventral markers. bmp4 is normally expressed in the ventral half of the shield stage embryo, except for a spot of expression in the shield (Fig. 4U) (Nikaido et al., 1997; Chin et al., 1997). In ichabod embryos of all classes, the expression of bmp4 was expanded to include all of the dorsal side of the embryo (Fig. 4V). The spot of shield expression was absent from all but one embryo examined (possibly a class 4 or 5 embryo). At later stages of gastrulation, bmp4 expression continued to be expressed radially in the marginal half of all ichabod embryos (Fig. 4X). Results were similar for bmp2b expression (data not shown). Two other genes normally expressed in ventral regions at 50% epiboly also showed dorsal expression. eve1 expression is normally restricted to the ventral and lateral cells of the marginal zone (Fig. 4Y) (Joly et al., 1993) and gata2 is normally only expressed in ventral ectoderm (Fig. 4A′) (Detrich et al., 1995). In all ichabod embryos, the expression ichabod embryos are deficient in nuclear localization of β-catenin Since the formation of the organizer was impaired in ichabod embryos and early YSL markers were not expressed, we tested whether β-catenin is appropriately localized in the nuclei on the dorsal side of the blastula embryo, as had been shown to occur in wild-type zebrafish and Xenopus embryos (Schneider et al., 1996; Larabell et al., 1997; Koos and Ho, 1998). As previously reported (Schneider et al., 1996; Koos and Ho, 1998), we found that nuclear localization of β-catenin in wildtype embryos first occurs in the dorsal YSL nuclei at the high stage and then in blastoderm cells starting at sphere stage (Fig. 5A,B). In contrast, ichabod embryos (immunohistochemically stained in parallel) show an absence of any nuclear localization of β-catenin at these stages (Fig. 5C,D). β-catenin appears to be present in these embryos, however, since staining of the cell membrane regions is as intense as in the wild-type embryos and staining of the YSL cytoplasm is apparent (indicated by arrows in Fig. 5C). This cytoplasmic staining permits visualization of the YSL and demonstrates that the layer is very irregularly shaped in ichabod embryos. Rescue of the ichabod phenotype by expression of β-catenin Since nuclear localization of β-catenin was absent in ichabod embryos, we tested whether injection of β-catenin RNA could rescue these embryos. Expression of Xenopus β-catenin resulted in rescue of goosecoid expression (Fig. 6) and in dorsalization of embryonic phenotypes (Fig. 7H). Identical results were also obtained with injection of zebrafish β-catenin RNA (data not shown). Moreover, phenotypic rescue was surprisingly effective; in many experiments, over one third of the injected ichabod embryos developed into fertile adults. Embryos injected with Xenopus β-catenin RNA showed nuclear localization of β-catenin and, interestingly, the region containing the nuclear localized protein was fairly discrete even though the co-injected GFP RNA lineage tracer was located throughout the embryo (data not shown). We then used RT-PCR to amplify β-catenin cDNA from RNA obtained from 2- to 4-cell embryos bred from ichabod homozygous females and compared its sequence with that from a parental wild-type strain. We found that there was no difference in the ORF encoded in the two RT-PCR products; thus, it is highly unlikely that ichabod is a mutation at the known β-catenin locus. This conclusion is supported by the finding of a different map position of the known zebrafish β-catenin gene (chromosome 16, Postlethwait et al., 1998). Upstream Wnt pathway intermediates fail to rescue ichabod embryos Because ichabod embryos could be dorsalized and rescued with β-catenin RNA, we next carried out a series of injections with RNAs encoding various proteins in the canonical Wnt→β-catenin pathway to determine whether a step upstream of β-catenin in the signaling pathway might be inactive in Maternal control of the zebrafish organizer 3905 ichabod mutant embryos. Embryos bred from ichabod homozygous females were injected with RNA and assayed for expression of the organizer marker goosecoid at shield stage (Fig. 6), or allowed to develop to 24 hpf and scored for the degree of anterior and dorsal deficiencies (Fig. 7). For experiments in which we assayed goosecoid expression, we also compared the transcript level with that of uninjected sibling embryos and of injected and non-injected wild-type embryos. For each experiment in which we scored the degree of dorsalization, uninjected siblings were scored for comparison as well. We also tested for the effect of each injected RNA on development of wild-type embryos. As shown in Figs 6 and 7A-C, RNAs encoding zebrafish Wnt8, zebrafish FrizzledA (zFzA) and Xenopus Dishevelled were all capable of activating ectopic goosecoid expression in control wild-type embryos, but did not activate goosecoid expression in ichabod embryos. Each of these RNAs was able to dorsalize (and in some cases, induce secondary axes in) wild-type embryos (data not shown), as predicted from previous studies of Wnt8 and zFzA RNA injections (Nasevicius et al., 1998; Kelly et al., 1995b), but had no dorsalizing effects on ichabod embryos. We also injected an RNA encoding a kinase-dead dominant negative form of Xenopus GSK3 (dnGSK K85→R, Pierce and Kimelman, 1995), previously shown to dorsalize zebrafish embryos when injected at high concentration (Nasevicius et al., 1998). Again, we obtained the expected dorsalizing effects with wild-type embryos, but not with ichabod embryos (Figs 6, 7E). We then tested for the dorsalizing effects of RNAs encoding the GSK3-binding protein GBP (Yost et al., 1998; Sumoy et al., 1999; Farr et al., 2000) or a fragment of Xenopus Axin (GID2, containing amino acids 320-429) that binds and inhibits GSK3 (Hedgepeth et al., 1999). Injected wild-type embryos were stimulated to produce high amounts of ectopic goosecoid transcript (Fig. 6) and were markedly dorsalized in phenotype (data not shown). ichabod embryos, in contrast, failed to activate goosecoid (Fig. 6) and showed an unchanged ventral phenotype distribution (Fig. 7D,F) in response to the injection of zebrafish GBP and Xaxin GID2 RNAs. Since Xaxin GID2 binds to GSK3 and inhibits its kinase activity (Hedgepeth et al., 1999) and GBP inhibits GSK3 by preventing its binding to full-length Axin (Farr et al., 2000) and facilitates the depletion of GSK3 (Dominguez and Green, 2000), it is unlikely that the wild-type ichabod gene product has a role in the control of GSK3 phosphorylation of β-catenin and, thus, in the control of β-catenin stability. We also note that our sequencing of GBP cDNA derived from ichabod embryos failed to detect any Fig. 6. β-catenin RNA, but not RNAs encoding upstream Wnt pathway proteins, can rescue goosecoid expression in ichabod embryos. Wild-type (wt) or ichabod embryos were injected with RNAs encoding the indicated proteins (z and X prefixes respectively indicate zebrafish and Xenopus proteins), or were uninjected controls. Embryos were allowed to develop to 50% epiboly and then were assayed for goosecoid expression by in situ hybridization. These experiments were carried out with batches of ichabod embryos that developed almost entirely class 1 and class 2 phenotypes. The lack of goosecoid expression was observed in every ichabod uninjected and injected embryo, except for those embryos injected with Xenopus β-catenin RNA. differences in sequence in the GBP ORF and that GBP maps to a different chromosome than ichabod (LG 16, Gates et al., 1999), thus further ruling out GBP as a candidate for ichabod. A gain-of-function mutation (unlikely from the recessive pattern of inheritance of ichabod) in Axin1 is also ruled out, since this is not on LG19 (M. Hibi, T. Hirano and R. Geisler, personal communication). 3906 C. Kelly and others Fig. 7. Phenotypic rescue of ichabod embryos by injection with β-catenin, zrn2/ndr1/squint, and bozozok/dharma/nieuwkoid RNAs, but not with RNAs encoding upstream Wnt pathway proteins. Frequencies of phenotypic classes of uninjected embryos (black) are compared to those of injected embryos (gray). Each panel represents a separate experiment carried out with a separate batch of embryos. Injected and uninjected embryos were scored at 24 hpf. For the Xenopus β-catenin RNA-injected embryos (H), class 5 embryos include 25 embryos that developed to adulthood (and are thus ‘class 6’, 17% complete phenotypic rescue). For embryo batches injected with znr2/ndr1/squint, or bozozok/dharma/nieuwkoid RNAs, class 5 embryos were only assayed at 24 hpf and we did not test for survival to adulthood. Injection of bozozok/dharma/nieuwkoid RNAs resulted in high percentages of hyperdorsalized embryos (column marked ‘D’ in [I]). RNAs encoding Wnt pathway proteins upstream of βcatenin, dn GSK3, GBP, X-axin GID-2 and XβTrcp∆F all failed to significantly ameliorate the ichabod phenotype. In each case, however, injection of these RNAs into wild-type embryos resulted in hyperdorsalized phenotypes (data not shown). Embryos were injected with approximately 1 nl of RNA solution, which varied in concentration from 50-1200 ng/µl, depending on the particular RNA. The approximate amount of each RNA (in pg) injected per embryo is indicated at the top of each panel, and the number of injected and non-injected control embryos is noted to the right of each panel. To test further the idea that the ichabod mutation affects organizer formation at a step distinct from the regulation of βcatenin stability, we injected mutant embryos with RNA encoding β-Trcp∆F, a dominant negative form of the Fbox/WD40-repeat protein that recruits phosphorylated βcatenin for degradation by the ubiquitination-proteosome pathway (Liu et al., 1999a). Injection of this RNA into Xenopus embryos results in the induction of ectopic axes due to the stabilization of β-catenin by blocking endogenous β-Trcp (Liu et al., 1999a). When we injected this RNA into wild-type zebrafish embryos, we observed the expected expansion of goosecoid expression (Fig. 6) and dorsalization of embryonic phenotype (data not shown), but there were no such effects in injected ichabod embryos (Figs 6, 7G). Thus, under conditions in which β-catenin would be stabilized by blocking the degradation pathway, we still observed no amelioration of the strongly ventralized ichabod phenotype. Taken together, these results strongly suggest that control of βcatenin stability is not impaired in the mutant. Our examination of the β-catenin ORF sequence indicates that the ichabod lesion is not in β-catenin and mapping data rule out the possibility that ichabod is a mutation in β-catenin cis-regulatory elements. These results therefore suggest that the ichabod lesion may affect an as yet poorly understood portion of the signaling pathway affecting β-catenin release from the Axin-GSK3 complex or in the control of transport of β-catenin into the nucleus. Maternal control of the zebrafish organizer 3907 Proteins that act downstream of β-catenin can dorsalize ichabod embryos To test whether two genes that are expressed in the zebrafish dorsal yolk syncytial layer (YSL), znr2/ndr1/squint and bozozok/dharma/nieuwkoid, are activated by nuclear β-catenin, we performed β-catenin RNA injection experiments. As also shown above (Fig. 4B,D), transcripts from these genes were absent from sphere stage ichabod embryos (Fig. 8E,G). Following injection of β-catenin RNA, not only did we observe an increase in both znr2/ndr1/squint and bozozok/dharma/ nieuwkoid transcripts in wild-type embryos (Fig. 8A-D) (as has also been found by Shimuzu et al., 2000), but both of these transcripts could now be detected at sphere stage (4.0 hpf) in ichabod embryos (Fig. 8F,H). These results indicate that ichabod embryos are capable of znr2/ndr1/squint and bozozok/dharma/nieuwkoid expression when exogenous βcatenin is supplied, and it can be concluded that β-catenin can activate the expression of these two genes. We next took advantage of the absence of nuclear β-catenin in ichabod embryos to test whether bozozok/dharma/nieuwkoid could activate znr2/ndr1/squint, and vice versa, without input from the β-catenin signaling pathway. Wild-type embryos injected with znr2/ndr1/squint RNA clearly showed activation of bozozok/dharma/nieuwkoid (Fig. 8I,J) and those injected with bozozok/dharma/nieuwkoid RNA showed an increase in expression znr2/ndr1/squint (Fig. 8K,L), but such activation could have merely enhanced parallel activation by Tcf3/βcatenin in wild-type embryos. When identical experiments were carried out with ichabod embryos, a striking activation was observed of bozozok/dharma/nieuwkoid by znr2/ndr1/ squint (Fig. 8M,N) and znr2/ndr1/squint by bozozok/ dharma/nieuwkoid (Fig. 8O,P). These results suggests that the pathway downstream of β-catenin signaling is not strictly linear, but that at least two key proteins important in organizer establishment can crossregulate their expression. The effects of injection of znr2/ndr1/squint and bozozok/ dharma/nieuwkoid RNAs on expression of goosecoid at shield stage were also examined. Expression of injected znr2/ndr1/ squint or bozozok/dharma/nieuwkoid RNAs had previously been shown to result in enlargement of the organizer region or formation of multiple organizers (Yamanaka et al., 1998; Koos and Ho, 1998; Erter et al., 1998; Feldman et al., 1998). Moreover, when injected into the dorsal YSL region, these RNAs functioned in a non-cell-autonomous fashion to induce goosecoid in the neighboring blastoderm margin (Yamanaka et al., 1998; Koos and Ho, 1998; Erter et al., 1998). We found, as expected, that injection of either of these RNAs markedly increases and expands goosecoid expression in wild-type embryos (Fig. 8Q-S). However, injection of bozozok/dharma/ nieuwkoid or znr2/ndr1/squint RNAs into ichabod embryos only weakly induced goosecoid (Fig. 8U-W). Interestingly, when the two RNAs were co-injected into ichabod embryos, the goosecoid expression domain was expanded to the same extent as when the RNAs were co-injected into wild-type embryos (Fig. 8T,X). Thus, in the absence of nuclear β-catenin, Znr2/Ndr1/Squint and Bozozok/Dharma/Nieuwkoid act synergistically to activate goosecoid. Further support for positioning znr2/ndr1/squint and bozozok/dharma/nieuwkoid downstream of ichabod and βcatenin was provided by examining the degree of phenotypic rescue of ichabod embryos injected with either znr2/ndr1/ squint or bozozok/dharma/nieuwkoid RNAs (Fig. 7I,J). A shift towards less severe phenotypes was obtained after injection of either of these RNAs. In either case, significant numbers of normal-appearing embryos (19-20%) were produced (although a large number of bozozok/dharma/nieuwkoid RNA-injected embryos, but none of the znr2/ndr1/squint RNA-injected embryos, were hyperdorsalized). Phenotypic rescue to class 5/6 embryos was lower, however, than with β-catenin RNA injections. These results also suggest that normal axis formation can proceed even when the level of goosecoid expression in the organizer is absent or very low, since significant numbers of bozozok/dharma/nieuwkoid RNAinjected ichabod embryos formed normal axes and exhibited no obvious dorsal or anterior defects, even though goosecoid transcripts were undetectable. Taken together, our findings clearly place znr2/ndr1/squint and bozozok/dharma/nieuwkoid downstream of ichabod and β-catenin in the dorsal axis pathway. DISCUSSION Maternal control of organizer formation in the zebrafish is mediated by β-catenin We have found that ichabod gene activity is required in the maternal parent, for proper control of the zygotic nuclear localization of β-catenin in the embryo. Phenotypic effects of a mutation in this gene are manifested in a classic maternal effect recessive mode of inheritance. The lack of zygotic requirement for the gene was indicated by our findings that inheritance of the ichabod phenotype was completely dependent on the female parental genome, that the male parental genome had no effect on the distribution of phenotypic classes of ichabod embryos and that crosses of heterozygous parents never yielded abnormal embryos. It has long been appreciated that, in Xenopus, dorsal vegetal cells from pre-midblastula transition embryos can induce mesoderm in conjugated ectodermal tissue (Nieuwkoop, 1969), and can induce a complete axis in UV-ventralized embryos or a second axis in normal embryos (Gimlich and Gerhart, 1984). Antisense-mediated mRNA-depletion experiments have identified β-catenin, GBP (an inhibitor of GSK3) and VegT as maternal factors essential for normal development (Heasman et al., 1994; Yost et al., 1998; Zhang et al., 1998). Since depletion of β-catenin RNA results in failure to form the organizer (Heasman et al., 1994) and GBP RNA depletion causes varying degrees of ventralization (Yost et al., 1998), it can be concluded that maternal gene function is important in regulating β-catenin levels in the Xenopus embryo. In teleosts as well, mesoderm formation and dorsoventral axis establishment are dependent on maternal gene action. In both goldfish and zebrafish, embryos defective in dorsal development were obtained when vegetal yolk was removed shortly after fertilization (Tung et al., 1945; Mizuno et al., 1997, 1999; Ober and Schulte-Merker, 1999), suggesting that essential maternal determinants are localized in this region of the egg. The results reported here clearly show that a consequence of a mutation in ichabod is the failure to properly localize β-catenin in dorsal YSL and blastoderm margin nuclei, and that this defect is entirely dependent on maternal ichabod activity. 3908 C. Kelly and others Fig. 8. znr2/ndr1/squint and bozozok/dharma/nieuwkoid act downstream of ichabod and β-catenin in the zebrafish axial signaling pathway. (A-H) Injection of β-catenin RNA activates expression of znr2/ndr1/squint and bozozok/dharma/nieuwkoid. Uninjected wildtype embryos express nieuwkoid (A) and squint (C) in the future dorsal margin (arrows) at 4 hpf, whereas expression of either gene is not observed in ichabod embryos (E,G). After injection of β-catenin RNA, expression of these two genes in wild-type embryos is greatly expanded (B,D), and expression of the two genes in ichabod embryos is observed in a discrete marginal region (arrows) (F,H). (I-P) znr2/ndr1/squint and bozozok/dharma/nieuwkoid can crossactivate each other in both wild-type and ichabod mutant embryos. Uninjected wild-type embryos express nieuwkoid (I) and squint (K). Injection of squint RNA results in a marked increase in nieuwkoid expression (J) and vice versa (L). The cross activation is also observed in ichabod embryos (M-P). (Q-X) znr2/ndr1/squint and bozozok/dharma/nieuwkoid act synergistically to activate goosecoid in ichabod embryos. Injection of either of these RNAs or a combination of the two RNAs into wild-type embryos causes marked expansion of the region expressing goosecoid (R,S) in comparison with the uninjected control (Q). A batch of ichabod embryos that developed into class 1 and class 2 phenotypes did not express goosecoid (U). Injection of bozozok/dharma/nieuwkoid or znr2/ndr1/squint RNAs activated goosecoid only weakly in these ichabod embryos (V,W), but co-injection of the two RNAs (X) resulted in expression of goosecoid at the same level as in wild-type embryos. The labels in the upper right corner indicate whether the embryo is wild-type or ichabod, the lower left corner, the injected RNA, and the lower right corner, the gene probed for expression. All embryos are at 4 hpf sphere stage and are shown in animal pole views. ichabod acts upstream of β-catenin but not by regulation of GSK3 activity or β-catenin stability We have used RNA injection as a way of testing the epistatic relationship between ichabod and β-catenin. Injection of βcatenin RNA resulted in effective rescue of wild-type phenotype in ichabod embryos. We consider it very unlikely that ichabod is a mutation in the β-catenin gene since the βcatenin ORF is identical in RNAs from eggs of ichabod homozygous mothers and from embryos of the parental wildtype strain. A mutation in non-coding regulatory regions of a β-catenin gene is possible, but our immunohistochemical assay suggests that levels of non-nuclear β-catenin in ichabod and wild-type embryos are comparable. Moreover, the map position of ichabod is on chromosome 19 (C. K. and E. S. W., unpublished), whereas that of the known β-catenin gene is on chromosome 16 (Postlethwait et al., 1998). Nuclear localization and stability of β-catenin can be modulated by the Wnt/wingless signaling pathway (reviewed in Cadigan and Nusse, 1997; Sokol, 1999). Although we were able to dorsalize wild-type embryos by injecting RNAs encoding zWnt8, zFzA or Xenopus Dishevelled, injection of these RNAs into ichabod embryos failed to activate goosecoid (Fig. 6) and had no effect on the degree of dorsalization of mutant phenotype (Fig. 7). Moreover, injection of RNAs encoding three proteins that negatively affect endogenous GSK3 activity similarly were unable to dorsalize ichabod embryos (Figs 6, 7), although they markedly affected wild-type embryos. These three RNAs encoded: (1) a kinase-dead dnGSK3 (Pierce and Kimelman, 1995), which interferes with GSK3 activity by binding to Axin, thus preventing GSK3 association with Axin (Farr et al., 2000), (2) GBP, which can function in a similar way (Farr et al., 2000), but which also facilitates a decrease in GSK3 levels in the embryo (Dominguez and Green, 2000), and (3) the Xenopus Axin fragment (GID-2), which rather than enhancing destabilization of β-catenin by providing a scaffold for GSK3 interaction with β-catenin, as is the case for full-length Axin, binds to GSK3 and inhibits its kinase activity (Hedgepeth et al., 1999). Thus, even a depletion of GSK3 activity in ichabod embryos does not lead to any measurable dorsalization, suggesting that the block in the mutants is not in the pathway of regulation of GSK3 kinase activity or GSK3 level. Since GSK3 is thought to directly phosphorylate β-catenin making it more susceptible to proteolysis (e.g., Yost et al., 1996), our result suggests that βcatenin stability may not be affected in ichabod mutant embryos. Further corroboration that ichabod does not function to regulate β-catenin stability was provided by the failure of injected Xenopus β-Trcp∆F RNA (Liu et al., 1999a) to dorsalize ichabod embryos. β-Trcp binds to β-catenin via its WD40-repeat and binds to the Skp1 ubiquitination complex through its F-box motif (Liu et al., 1999a). If β-Trcp lacks the F-box (β-Trcp∆F), the protein acts as a dominant negative and prevents the recruitment of β-catenin to the ubiquitination- Maternal control of the zebrafish organizer 3909 proteosome complex (Liu et al., 1999a). Our result showing that injection of β-Trcp∆F RNA failed to dorsalize ichabod mutants embryos, while clearly having a dorsalizing effect on wild-type embryos, strongly suggests that inhibition of βcatenin degradation cannot rescue the ichabod phenotype. If the ichabod mutation is not located in the β-catenin gene and ichabod mutants are not impaired in β-catenin stability, then what step might be blocked in the mutant embryos? We propose that the wild-type ichabod protein may function in a step facilitating β-catenin release from the Axin-GSK3-APC complex, or in the promotion of nuclear transport of β-catenin. These functions are not yet understood in β-catenin signaling and it is quite likely that ichabod encodes a novel protein required for stabilized β-catenin to be utilized as a component of the dorsal signaling pathway. The YSL signaling center can form in ichabod embryos injected with β-catenin RNA The finding that expression in the YSL of either Znr2/Ndr1/Squint or Bozozok/Dharma/Nieuwkoid can induce organizer markers in the overlying blastoderm margin has led to the conclusion that these proteins are mediators of Nieuwkoop center activity (Erter et al., 1998; Feldman et al., 1998; Yamanaka et al., 1998; Koos and Ho, 1998; Fekany et al., 1999). We found that ichabod embryos fail to express znr2/ndr1/squint and bozozok/dharma/nieuwkoid in the dorsal YSL and that injection of β-catenin RNA resulted in the activation of expression of these two genes. bozozok/ dharma/nieuwkoid has been posited to be a downstream target of β-catenin signaling in the YSL (Yamanaka et al., 1998; Fekany et al., 1999). Here, we demonstrate that the absence of nuclear β-catenin in the YSL results in loss of expression of this gene and of znr2/ndr1/squint as well, and that injection of β-catenin RNA can activate expression of both genes in wildtype embryos and in ichabod embryos deficient in nuclear βcatenin. These results not only confirm the enhancement of expression of these two genes by β-catenin in wild-type embryos (Shimizu et al., 2000), but also, using the ichabod mutants blocked in nuclear β-catenin, directly demonstrate the dependence of bozozok/dharma/nieuwkoid and znr2/ndr1/ squint expression on endogenous β-catenin signaling. A novel aspect of our studies is the demonstration that, in both wildtype embryos and ichabod mutant embryos, each of these genes was capable of inducing the other. Thus, even in the absence of nuclear β-catenin, znr2/ndr1/squint can strongly induce expression of bozozok/dharma/nieuwkoid (Fig. 8M,N), and bozozok/dharma/nieuwkoid can induce expression of znr2/ndr1/squint, albeit more weakly (Fig. 8O,P). A recently published study failed to detect these activations in wild-type embryos, probably because far lower amounts of RNA were injected (Shimizu et al., 2000). Since bozozok/ dharma/nieuwkoid is probably a direct target of Tcf3/β-catenin as judged by the presence of Tcf3-binding sites in the bozozok promoter (Yamanaka et al., 1998), one activation pathway is probably Tcf3/β-catenin→bozozok→squint. Since bozozok expression is normal in maternal-zygotic one eyed pinhead mutant blastulae (Gritsman et al., 1999), direct activation by βcatenin in the absence of squint is sufficient for normal blastula expression of bozozok. However, our results suggest that the Tcf3/β-catenin→squint→bozozok sequence can also operate, although it is possible that squint may not be a direct target of Tcf3/β-catenin. The crossactivation of squint and bozozok might allow persistence of activation of these genes after a decrease of β-catenin signal during development. Injection of either bozozok/dharma/nieuwkoid or znr2/ndr1/ squint RNAs into ichabod mutant embryos only weakly activated goosecoid, but co-injection of the two RNAs strikingly activated expression of this gene (Fig. 8U-X). These results are interesting in two respects. First, although goosecoid levels were very low (or even absent in one experiment) in bozozok/dharma/nieuwkoid RNA-injected ichabod embryos, injection of the RNA strongly dorsalized ichabod embryos and resulted in many normal appearing embryos (Fig. 7J). This finding suggests that axis formation and correct dorsoventral patterning can proceed in zebrafish embryos even with little or no goosecoid expression. This result is in agreement with the findings that mouse embryos homozygous for a null goosecoid allele do not have gastrulation or neural induction defects (Yamada et al., 1995; Rivera-Perez et al., 1995) and chick embryos can develop normally even when manipulated so that their organizer does not express goosecoid (Psychoyos and Stern, 1996). Second, since co-injection of the two RNAs strongly resulted in synergistic activation of goosecoid (Fig. 8T,X), a significant interaction of gene pathways dependent on expression of a TGFβ protein (Znr2/Ndr1/Squint) and Bozozok/Dharma/Nieuwkoid can occur in the region of the blastoderm margin. However, since we obtained significant crossactivation of the two genes (Fig. 8N,P), we might have expected that levels of both proteins present in ichabod embryos injected with only one of these RNAs would be sufficient to synergistically activate goosecoid. Further analysis on the molecular structure of the induced transcripts produced endogenously must be carried out before these findings can be fully understood. The block in β-catenin signaling in ichabod mutant embryos provides us with an opportunity to further examine the spatial and temporal aspects of the network of gene interactions that occur in the YSL and dorsal blastoderm margin. We plan to extend these studies to investigate whether other proteins expressed in the YSL are regulated in linear pathways or whether there is extensive crossactivation of genes expressed in this early signaling center. We thank Gianfranco Bellipanni, Mary Mullins, Michael Granato, Peter Klein, Daniel Kessler and Cecilia Lo for helpful discussions during the course of this work. We are grateful to Dr C. Kimmel for sending us the flh stock of zebrafish and Mary Mullins for supplying us with antibody against β-catenin. cDNA clones were kindly provided to us by R. Ho, M. Rebagliati, C. Wright, D. Grunwald, S. Schulte-Merker, R. Toyama, M. Halpern, G. Kelly, S. Ekker, D. Kimelman, M. Mullins, P. Klein, X. He and D. Kessler. This work was supported by NIH grant RO1 NS34365 to E. S. W., a University of Pennsylvania Research Foundation grant to E. S. W., and NIH training grant fellowships to C. K. and D. J. K. REFERENCES Abdelilah, S. and Driever, W. (1997). Pattern formation in janus-mutant zebrafish embryos. Dev. Biol. 184, 70-84. Cadigan, K. M. and Nusse, R. (1997). Wnt signaling: a common theme in animal development. Genes Dev. 11, 3286-3305. Chin, A. J., Chen, J.-N. and Weinberg, E. S. (1997). Bone morphogenietic protein-4 expression characterizes inductive boundaries in organs of developing zebrafish. Dev. Genes Evol. 207, 107-114. 3910 C. Kelly and others Detrich, H. W., Kieran, M. W., Chan, F. Y., Barone, L. M., Yee, K., Rundstadler, J. A., Pratt, S., Ransom, D. and Zon, L. I. (1995). Extraembryonic hematopoietic-cell migration during vertebrate development. Proc. Natl. Acad. Sci. USA 92, 10713-10717. Dominguez, I. and Green, J.B.A. (2000). Dorsal downregulation of GSK3β by a non-Wnt-like mechanism is an early molecular consequence of cortical rotation in early Xenopus embryos. Development 127, 861-868. Erter, C. E., Solnica-Krezel, L. and Wright, C. V. E. (1998). Zebrafish nodal-related 2 encodes an early mesendodermal inducer signaling from the extraembryonic YSL. Dev. Biol. 204, 361-372. Farr, G.H., Ferkey, D.M., Yost, C., Pierce, S.B., Weaver, C. and Kimelman, D. (2000). Interaction among GSK-3, GBP, axin, and APC in Xenopus axis specification. J. Cell Biol. 148, 691-701. Fekany, K., Yamanaka, Y., Leung, T, Sirotkin, H. I., Topczewski, J., Gates, M. A., Hibi, M., Renucci, A., Stemple, D., Radbill, A., Schier, A. F., Driever, W., Hirano, T., Talbot, W. S. and Solnica-Krezel, L. (1999). The zebrafish bozozok locus encodes Dharma, a homeodomain protein essential for induction of gastrula organizer and dorsoanterior embryonic structures. Development 126, 1427-1438. Feldman, B., Gates, M. A., Egan, E. S., Dougan, S. T., Rennebeck, G., Sirotkin, H. I., Schier, A. F. and Talbot, W. S. (1998). Zebrafish organizer development and germ-layer formation require nodal-related signals. Nature 395, 181-185. Fisher, S., Amacher, S. L. and Halpern, M. E. (1997). Loss of cerebum function ventralizes the zebrafish embryo. Development 124, 1301-1311. Funuyama, N., Fagotto, F., McCrea, P. and Gumbiner, B. M. (1995). Embryonic axis induction by the armadillo repeat domain of beta-catenin – evidence for intracellular signaling. J. Cell Biol. 128, 959-968. Gates, M. A., Kim, L., Egan, E. S., Cardozo, T., Sirotkin, H. I., Dougan, S. T., Lashkari, D., Abagyan, R., Shier, A. F. and Talbot, W. S. (1999). A genetic linkage map for zebrafish: comparative analysis and localization of genes and expressed sequences. Genome Res. 9, 334-347. Gerhart, J., Danilchik, M., Doniach, T., Roberts, S., Rowing, B. and Steward, R. (1989). Cortical rotation of the Xenopus egg: Consequences for the anteroposterior pattern of embryonic dorsal development. Development 107 Supplement, 37-51. Gimlich, R. L. and Gerhart, J. C. (1984). Early cellular interactions promote embryonic axis formation in Xenopus laevis. Dev. Biol. 104, 117-130. Gritsman, K., Zhang, J., Cheng, S., Hecksher, E., Talbot, W.S. and Shier, A.F. (1999). The EGF-CFC protein one-eyed pinhead is essential for nodal signaling. Cell 97, 121-132. Guger, K. A. and Gumbiner, B. M. (1995). β-catenin has wnt-like activity and mimics the Nieuwkoop signaling center in Xenopus dorsal-ventral patterning. Dev. Biol. 172, 115-125. Harland, R. and Gerhart, J. (1997). Formation and function of Spemann’s organizer. Annu. Rev. Cell Dev. Biol. 13, 611-667. Heasman, J., Crawford, A., Goldstone, K., Garner-Hamrick, P., Gumbiner, B., McCrea, P., Kintner, C., Noro, C. Y. and Wylie, C. (1994). Overexpression of cadherins and underexpression of β-catenin inhibit dorsal mesoderm induction in early Xenopus embryos. Cell 79, 791-803. Hedgepeth, C, M., Conrad, L. J., Zhang, J., Huang, H. C., Lee, V. M. and Klein, P. S. (1997). Activation of the Wnt signaling pathway: a molecular mechanism for lithium action. Dev. Biol. 185, 82-91. Hedgepeth, C.M., Deardorff, M.A., Rankin, K. and Klein, P.S. (1999). Regulation of glycogen synthase kinase 3β and downstream Wnt signaling by axin. Mol. Cell. Biol.19, 7147-7157. Huelsken, J., Vogel, R., Brinkmann, V., Eerdman, B., Birchmeier, C. and Birchmeier, W. (2000). Requirement for β-catenin in anterior-posterior axis formation in mice. J. Cell Biol. 148, 567-578. Jesuthasan, S. and Strähle, U. (1997). Dynamic microtubules and specification of the zebrafish embryonic axis. Curr. Biol. 185, 82-91. Joly, J. S., Joly, C., Schulte-Merker, S., Boulekbache, H. and Condamine, H. (1993). The ventral and posterior expression of the zebrafish homeobox gene eve1 is perturbed in dorsalized and mutant embryos. Development 119, 1261-1275. Kelly, G. M., Erezyilmaz, D. F. and Moon, R. T. (1995a). Induction of a secondary embryonic axis in zebrafish occurs following the overexpression of β-catenin. Mech. Dev. 53, 261-273. Kelly, G. M., Greenstein, P., Erezyilmaz, D. F. and Moon, R. T. (1995b). Zebrafish wnt8 and wnt8b share a common activity but are involved in distinct developmental pathways. Development 121, 1787-1799. Koos, D. S. and Ho, R. K. (1998). The nieuwkoid gene characterizes and mediates a Nieuwkoop-center-like activity in the zebrafish. Curr. Biol. 8, 1199-1206 Larabell, C. A., Torres, M., Rowning, R. A., Yost, C., Miller, J. R., Wu, M., Kimelman, D. and Moon, R. T. (1997). Establishment of the dorsoventral axis in Xenopus embryos is presaged by early asymmetries in beta-catenin that are modulated by the Wnt signaling pathway. J. Cell Biol. 136, 1123-1136. Li, Y., Allende, M. L., Finkelstein, R. and Weinberg, E. S. (1994). Expression of two zebrafish orthodenticle-related genes in the embryonic brain. Mech. Dev. 48, 229-244. Liu, C., Kato, Y., Zhang, Z., Do, V.M., Yanker, B.A. and He, X. (1999a). β-Trcp couples β-catenin phosphorylation-degradation and regulates Xenopus axis formation. Proc. Natl. Acad. Sci. USA 96, 6273-6278. Liu, P., Wakamiya, M., Shea, M.J., Albrecht, U., Behringer, R.R. and Bradley, A. (1999b). Requirement for Wnt3 in vertebrate axis formation. Nature Genet. 22, 361-365. Miller, J.R., Rowning, B.A., Larabell, C.A., Yang-Snyder, J.A., Bates, R.L. and Moon, R.T. (1999). Establishment of the dorsal-ventral axis in Xenopus embryos coincides with the dorsal enrichment of Dishevelled that is dependent on cortical rotation. J. Cell Biol. 146, 427-437. Miller-Bertoglio, V. E., Fisher, S., Sanchez, A., Mullins, M. C. and Halpern, M. E. (1997). Differential regulation of chordin expression domains in mutant zebrafish. Dev. Biol. 192, 537-550. Mizuno, T., Yamaha, E., Wakahara, M., Kuroiwa, A. and Takeda, H. (1996). Mesoderm induction in zebrafish. Nature 383, 131-132. Mizuno, T., Yamaha, E. and Yamazaki, F. (1997). Localized axis determinant in the early cleavage embryo of goldfish, Carassius auratus. Dev. Genet. Evol. 206, 389-396. Mizuno, T., Yamaha, E., Kuroiwa, A. and Takeda, H. (1999). Removal of vegetal yolk causes dorsal deficiencies and impairs dorsal-inducing ability of the yolk cell in zebrafish. Mech. Dev. 81, 51-63. Moon, R. T. and Kimelman, D. (1998). From cortical rotation to organizer gene expression: Toward a molecular explanation of axis specification in Xenopus. Bioessays 20, 536-545. Nasevicius, A., Hyatt, T., Kim, H., Guttman, J., Walsh, E., Sumanas, S., Wang, Y. and Ekker, S. C. (1998). Evidence for a frizzled-mediated wnt pathway required for zebrafish dorsal mesoderm formation. Development 125, 4283-4292. Nieuwkoop, P. D. (1969). The formation of the mesoderm in urodelean amphibians, II: the origin of the dorso-ventral polarity of the mesoderm. Wilhelm Roux’s Arch. Dev. Biol. 163, 298-315. Nikaido, M., Tada, M., Safi, T. and Ueno, N. (1997). Conservation of BMP signaling in zebrafish mesoderm patterning. Mech. Dev. 61, 75-88. Ober, E. O. and Schulte-Merker, S. (1999). Signals from the yolk cell induce mesoderm, neurectoderm, the trunk organizer, and the notochord in zebrafish. Dev. Biol. 215, 167-181. Pelegri, F. and Maischein, H. M. (1998). Function of zebrafish β-catenin and TCF-3 in dorsoventral patterning. Mech. Dev. 77, 63-74. Pierce, S. B. and Kimelman, D. (1995). Regulation of Spemann organizer formation by the intracellular kinase Xgsk-3. Development 121, 755765. Postlethwait, J. H., Yan, Y. L., Gates, M. A., Horne, S., Amores, A., Brownlie, A., Donovan, A., Egan, E. S., Force, A., Gong, Z. Y., et al. (1998). Vertebrate genome evolution and the zebrafish gene map. Nature Genet. 18, 345-349. Psychoyos, D. and Stern, C. D. (1996). Restoration of the organizer after radical ablation of Hensen’s node and the anterior primitive streak in the chick embryo. Development 122, 3263-3273. Rebagliati, M. R., Toyama, R., Fricke, C., Haffter, P. and Dawid, I. B. (1998). Zebrafish nodal-related genes are implicated in axial patterning and establishing left-right asymmetry. Dev. Biol. 199, 261-272. Rivera-Perez, J.A., Mallo, M., Gendron-Maguire, M., Gridley, T., Behringer, R.R. (1995). Goosecoid is not an essential component of the mouse gastrula organizer but is required for craniofacial and rib development. Development 121, 3005-3012. Schneider, S., Steinbesser, H., Warga, R. M. and Hausen, P. (1996). βcatenin translocation into nuclei demarcates the dorsalizing centers in frog and fish embryos. Mech. Dev. 57, 191-198. Schulte-Merker, S., Ho, R. K., Hermann, B. G. and Nüsslein-Volhard, C. (1992). The protein product of the mouse T-gene is expressed in nuclei of the germ ring and the notochord of the early embryo. Development 116, 1021-1032. Schulte-Merker, S., Hammerschmidt, M., Beuchle, E., Cho, K. W., DeRobertis, E. M. and Nüsslein-Volhard, C. (1994). Expression of zebrafish goosecoid and no tail gene-products in wild-type and mutant not tail embryos. Development 120, 843-852. Maternal control of the zebrafish organizer 3911 Shimizu, T., Yamanaka, Y., Ryu, S.-L., Hashimoto, H., Yabe, T., Hirata, T., Bae, Y.-K., Hibi, M., Hirano, T. (2000). Cooperative roles of Bozozok/Dharma and Nodal-related proteins in the formation of the dorsal organizer in zebrafish. Mech. Dev. 91, 293-303. Sokol, S. Y. (1999). Wnt signaling and dorsoventral axis specification in vertebrates. Curr. Opin. Genet. Dev. 9, 405-410. Sokol, S. Y., Klingensmith, J., Perrimon, N. and Itoh, K. (1995). Dorsalizing and neuralizing properties of Xdsh, a meternally expressed Xenopus homolog of dishevelled. Development 121, 1637-1647. Solnica-Krezel, L., Stemple, D. L., Mountcastle-Shah, E., Rangini, Z., Neuhauss, S. C. F., Malicki, J., Schier, A., Stainier, D. Y. R., Zwartkruis, F., Abdelilah, S. and Driever, W. (1996). Mutations affecting cell fates and cellular rearrangements during gastrulation in zebrafish. Development 123, 117-128. Spemann, H. and Mangold, H. (1924). Uber Induktion von Embryonanlagen durch Implantation artfremder Organisatoren. Wilhelm Roux Arch. EntwMech. Org. 100, 599-638. Stachel, S. E., Grunwald, D. J. and Myers, P. Z. (1993). Lithium perturbation and goosecoid expression identify a dorsal specification pathway in the pregastrula zebrafish. Development 117, 1261-1274. Sumoy, L., Kiefer, J. and Kimelman, D. (1999). Conservation of intracellular Wnt signaling components in dorsal-ventral axis formation in zebrafish. Dev. Genes Evol. 209, 48-58. Talbot, W. S., Trevarrow, B., Halpern, M. E., Melby, A. E., Farr, G., Postlethwait, J. H., Jowett, T., Kimmel, C. B. and Kimelman, D. (1995). A homeobox gene essential for zebrafish notochord development. Nature 378, 150-157. Toyama, R. and Dawid, I. B. (1997). lim6, a novel LIM homeobox gene in the zebrafish: comparison of its expression pattern with lim1. Dev. Dyn. 209, 406-417. Toyama, R., O’Connell, M. L., Wright, C. V. E., Kuehn, M. R. and Dawid, I. B. (1995). nodal induces ectopic goosecoid and lim1 expression and axis duplication in zebrafish. Development 121, 383-391. Tung, T.-C., Chang, C.-Y. and Tung, Y.-F. Y. (1945). Experiments on the developmental potencies of blastoderms and fragments of teleostean eggs separated latitudinally. Proc. Zool. Soc. 115, 175-188. Westerfield, M. (1993). The Zebrafish Book: A Guide for the Laboratory Use of Zebrafish (Brachydanio rerio). Eugene, OR: Univ. of Oregon Press. Wylie, C., Kofron, M., Payne, C., Anderson, R., Hosobuchi, M., Joseph, E. and Heasman, J. (1996). Maternal β-catenin establishes a ‘dorsal signal’ in early Xenopus embryos. Development 122, 2987-2996. Yamada, G., Mansouri, A., Torres, M., Stuart, E.T., Blum, M., Schultz, M., De Robertis E.M. and Gruss, P. (1995). Targeted mutation of the murine goosecoid gene results in craniofacial defects and neonatal death. Development 121, 2917-2922. Yamanaka, Y., Mizuna, T., Sasai, Y., Khishi, M., Takeda, H., Kim, C.-H., Hibi, M. and Hirano, T. (1998). A novel homeobox gene, dharma, can induce the organizer in a non cell-autonomous manner. Genes Dev. 12, 23452353. Yost, C., Torres, M., Miller, J. R., Huang, E., Kimelman, D. and Moon, R. T. (1996). The axis-inducing activity, stability, and subcellular distribution of β-catenin is regulated in Xenopus embryos by glycogen synthase kinase 3. Genes Dev. 10, 1443-1454. Yost, C., Farr, G. H. III, Pierce, S. B., Ferkey, D. M., Chen, M. M. and Kimelman, D. (1998). GBP, an inhibitor of GSK-3, is implicated in Xenopus development and oncogenesis. Cell 93, 1031-1041. Zhang, J., Houston, D. W., King, M. L., Payne, C., Wylie, C. and Haesman, J. (1998). The role of maternal VegT in establishing the primary germ layers in Xenopus embryos. Cell 94, 515-524.