Survey
* Your assessment is very important for improving the workof artificial intelligence, which forms the content of this project
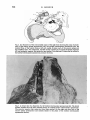
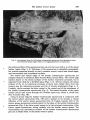
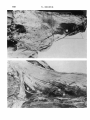
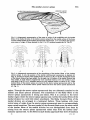
525 J. Anat. (1980), 131, 3, pp. 525-540 With 1O figures Printed in Great Britain A reappraisal of the anatomy of the human lumbar erector spinae NIKOLAI BOGDUK Division of Neurology, Prince Henry Hosptial and Department of Medicine, University of New South Wales, Sydney, Australia (Accepted 2 April 1980) INTRODUCTION In the course of a study of the lumbar dorsal rami, observations were made of the gross anatomy of the lumbar erector spinae muscle. It was found that the observations made were at variance with the descriptions of this muscle given in textbooks. The variance was so marked that it was considered appropriate formally to present the observations and their ramifications, and to reappraise the current interpretation of the anatomy of the lumbar erector spinae. METHODS The lumbar erector spinae was studied by gross dissection in four embalmed adult cadavers. The detailed anatomy of the muscle was studied by systematically resecting its component fascicles and noting their disposition and attachments. The more superficial and lateral fibres were resected first. This revealed the more deeply lying fibres which were then dissected in turn, until the entire erector spinae had been resected. The study was restricted to the fleshy longissimus thoracis and iliocostalis lumborum components of the erector spinae. The spinalis thoracis (dorsi) is mainly aponeurotic in the lumbar region and was not included in the study. The spinalis thoracis and its variations have been fully described previously (Winckler, 1937). RESULTS The lumbar erector spinae is a large muscle mass lying lateral to the multifidus muscle (Fig. 1). It is largely covered by an aponeurosis, the erector spinae aponeurosis, which arises from the dorsal segment of the iliac crest, the sacral and lumbar spinous processes and the intervening supraspinous ligaments. The tendinous fibres of the aponeurosis pass rostrolaterally across the multifidus and erector spinae until they become fleshy. The aponeurosis also gives rise to other fleshy fibres of the lumbar erector spinae from its deep and superficial surfaces (q.v.). On inspection (Fig. 1), the lumbar erector spinae in the dorsal lumbar region appears to be a common muscle mass which, as it enters the dorsal thoracic region splits into medial and lateral divisions which are separated by the iliocostalis thoracis. Dissection of the common muscle mass, however, reveals that the segregation into medial and lateral divisions is also manifest in the lumbar region. The common mass of the lumbar erector spinae is divided into two divisions by the 0021-8782/80/2828-8260 $02.00 © 1980 Anat. Soc. G.B. & I 526 N. BOGDUK .~~~~~~~~~~~~~~~~~~~~ .tus _~~~~~~~~~~~~~ S t ". A l (a)Ontelf,telmaercospaeshasbersctdodmntaeth neurosis~~ 2 relative positions of the iliocostalis thoracis (it) and multifidus (in). The posterior superior iliac spines (ps) and lumbar transverse process (tp) are marked. (B) The lateral division (id) of the lumbar erector spinae (iliocostalis lumborum) has been reflected to demonstrate the cleavage planes about iliocostalis thoracis (it) which separate the lateral division from the medial division (md) (longissimus thoracis). The lumbar erector spinae 527 erector spinae aponeurosis and an intermuscular aponeurosis. This latter aponeurosis has no formal name (Nomina Anatomica, 1977), so it is referred to herein as the lumbar intermuscular aponeurosis. The lumbar intermuscular aponeurosis The lumbar intermuscular aponeurosis stems from the ilium. It is attached to a linear area on the medial aspect of the dorsal segment of the iliac crest just rostral to the posterior superior iliac spine. This line of attachment extends some 2-3 cm ventrolaterally across the medial aspect of the iliac crest (Fig. 2). From this origin 528 N. BOGDUK PS Fig. 2. An illustration of the rostromedial aspect of the right ilium showing the areas of attachment of the erector spinae aponeurosis (esa), the lumbar intermuscular aponeurosis (lia), the lumbar fibres of the lateral division (Id) and medial division (md), of the erector spinae and the multifidus (mf). For orientation, the fifth lumbar vertebra (L5) and the iliolumbar ligament (ill) and posterior superior iliac spine (ps) are marked. Note the area (f) where the fat ventral to the lumbar intermuscular aponeurosis contacts the ilium. r lia t kR = e _ L >;! k" '; l, x v *~~~~~~~~~~~4, e Fig. 3. A dorsal view of a dissection of a left lumbar intermuscular aponeurosis (lia). The dorsal lumbar muscles have all been resected leaving an isolated intact intermuscular aponeurosis. The posterior superior iliac spines (ps) have been marked. On the right note how little of the erector spinae aponeurosis (esa) extends lateral to the plane of the lumbar intermuscular aponeurosis (arrow). The lumbar erector spinae 529 Fig. 4. A dorsolateral view of a left lumbar intermuscular aponeurosis (lia) showing its extent into the dorsal lumbar region. The Ll-L2 zygapophysial joint is marked (zj). 41 the tendinous fibres of the aponeurosis fan out into the lower half or so of the dorsal lumbar region (Figs. 3, 4). The plane of the aponeurosis is essentially parasagittal, but rotated somewhat laterally so that it presents rostral, ventral and dorsal edges, and ventromedial and dorsolateral surfaces. The rostral and ventral edges of the lumbar intermuscular aponeurosis are irregular because its tendinous fibres become fleshy muscle fibres. The ventral edge reaches almost to the dorsal surfaces of the lumbar transverse processes, but is separated from them by a fat-filled space (Figs. 5, 6). The fat contacts the dorsal surfaces of the L1-L4 transverse processes at about their middle third. Over this contact area the transverse processes are free of muscle attachments (Figs. 6, 7). Caudally, the fat contacts the ilium ventral to the ventral end of the attachment of the lumbar intermuscular aponeurosis (Fig. 2). The lateral branches of the upper lumbar dorsal rami course through this fat-filled space and, within it, divide into their branches (Fig. SB). Along its dorsal edge the lumbar intermuscular aponeurosis is continuous with the erector spinae aponeurosis (Fig. 5B) and it appears to be actually a ventral reflection of the erector spinae aponeurosis (Fig. 6). Indeed, because little of the erector spinae aponeurosis extends lateral to the plane of the lumbar intermuscular aponeurosis (Fig. 3) the latter would appear to be the principal continuation of the erector spinae aponeurosis, diving ventrally into the lumbar erector spinae. The ventromedial and dorsolateral surfaces of the lumbar intermuscular aponeurosis give rise to fleshy fibres of the lumbar erector spinae. 530 N. BOGDUK AL-- -t The lumbar erector spinae 531 Fig. 6. A diagramatic transverse section of the lower lumbar region showing the relative disposition of the muscles, fasciae and aponeuroses. The erector spinae aponeurosis (esa) covers the interspinalis (I), multifidus (MF), and the medial division (MD) of the lumbar erector spinae, viz. longissimus thoracis (LT). It is continuous ventrally with the lumbar intermuscular aponeurosis (lia) which envelopes the lateral surface of the medial division, segregating it from the lateral division (LD), viz. iliocostalis lumborum (IL). The fat-filled space (fs) ventral to the lumbar intermuscular aponeurosis maintains this segregation as far as the transverse process. The medial division inserts about the accessory process (ap) while the lateral division inserts further laterally on the transverse process. Lateral to the lumbar intermuscular aponeurosis the erector spinae aponeurosis extends for a limited distance over the dorsal surface of the lateral division (cf. Fig. 1). The intertransversarii mediales (IM) are marked. The lumbar intermuscular aponeurosis and erector spinae aponeurosis form a continuous aponeurotic envelope which, in the lumbar region, divides the lumbar erector spinae into a medial and a lateral division (Fig. 6). Medial division of the lumbar erector spinae In the lumbar region the medial division of the lumbar erector spinae lies medial to the lumbar intermuscular aponeurosis and deep to the erector spinae aponeurosis (Fig. 6), while in the thoracic region it lies medial to iliocostalis thoracis (Fig. 1). Within the medial division lumbar and thoracic fibres may be identified. The lumbar fibres are those which insert into lumbar vertebrae. The thoracic fibres insert into thoracic vertebrae and ribs. The lumbar fibres of the medial division of the lumbar erector spinae arise from the medial aspect of the middle third or so of the dorsal segment of the iliac crest and from the medial surface of the lumbar intermuscular aponeurosis (Fig. 8). The attachment to the lumbar intermuscular aponeurosis is strong and dense. The muscle fibres are arranged in five laminae, each directed to a certain lumbar vertebra (Figs. 8, 9). The more caudal fibres from the ilium form the deepest laminae. They pass Fig. 5. (A) A lateral view of the right lumbar erector spinae and erector spinae aponeurosis (esa) with an outline of the area dissected and shown in close up in Fig. 5(B). For orientation, the posterior superior iliac spine (ps) and the lateral branches of the L3 and Li dorsal rami (L3 dr, Li dr), where they cross the iliac crest, have been marked. The lateral division (Id) of the lumbar erector spinae (iliocostalis lumborum) was resected to produce Fig. 5 (B). (B) A close up lateral view of a right lumbar intermuscular aponeurosis (lia). Note its continuity with the erector spinae aponeurosis (esa) dorsally; its irregular ventral edge (e), and the fat-filled space (fs) ventral to it through which lateral branches (lb) of the lumbar dorsal rami course. 532 N. BOGDUK Fig. 7. An illustration of the insertions of the lumbar fibres of the medial division (md) and the lateral division (Id) of the lumbar erector spinae. Note the areas (f) on the transverse processes free of muscle attachment which are in contact with the fat ventral to the lumbar intermuscular aponeurosis, and the tendinous (t) insertion into the tips of the accessory processes and the more fleshy insertion about their bases, of the medial division. ventrally and medially to insert into the accessory process and the dorsal surface of the transverse process of the fifth lumbar vertebra (Figs. 7, 9). The next deepest lamina arises from the ilium, just rostral to the origin of the deepest lamina, and from the medial surface of the lumbar intermuscular aponeurosis at its origin (Fig. 8). This lamina covers the deepest one and inserts into the accessory process and adjacent medial half or so of the transverse process of the fourth lumbar vertebra (Figs. 7, 9). The remaining three laminae arise from successively more superficial and rostral parts of the medial surface of the lumbar intermuscular aponeurosis as depicted in Figure 8. They cover one another in turn as they pass to their insertions on the L 3 to LI vertebrae respectively (Fig. 9). In each lamina the more caudal fibres are the more medial. They insert by way of a musculotendinous attachment into an accessory process. Some of these fibres may bypass the accessory process and insert into the adjacent mamillary process and the mamillo-accessory ligament. The more rostral fibres in each lamina lie more laterally and insert by way of a musculoperiosteal attachment into the medial end of the dorsal surface of a transverse process, principally around the root of the accessory process (Fig. 7). The thoracic fibres of the medial division of the lumbar erector spinae arise from the deep surface of the overlying erector spinae aponeurosis in the upper lumbar The lumbar erector spinae lumbar intermuscular aponeurosis 533 posterior superior / iliac spine Fig. 8. A diagramatic representation of the areas of origin of the multifidus and the lumbar fibres of the medial division of the lumbar erector spinae from the medial surface of the ilium, and the medial surface of the lumbar intermuscular aponeurosis. Li to L5 indicate the approximate areas of origin of fibres destined for the LI to L5 vertebrae respectively (cf. Fig. 9). )) 0 00 OQo0 0 A C B Fig. 9. A diagramatic reconstruction of the morphology of the lumbar fibres of the lumbar erector spinae. In (A), the position of the lumbar intermuscular aponeurosis is depicted (cf. (Fig. 3), and the L5 lamina of the medial division is shown. In (B), on the left, the deepest lamina of the lateral division has been added. On the right, the L4 lamina of the medial division has been added. In each division progressively more superficial lamina cover one another in turn until each of the L3 to LI vertebrae receives its own separate lamina, as shown in (C). These lumbar fibres of the erector spinae are covered by the erector spinae aponeurosis and by the thoracic fibres of the lumbar erector spinae which arise from the erector spinae aponeurosis (cf. Fig. 1). region. Through the erector spinae aponeurosis they are ultimately attached to the lumbar and sacral spinous processes. The attachment of the fleshy fibres to the erector spinae aponeurosis is strong and dense. This is in contrast to the lower lumbar region where the erector spinae aponeurosis overlies the lumbar fibres of the medial division but with little direct attachment. The fleshy thoracic fibres of the medial division are arranged in a laminated fashion. Those laminae with more rostral areas of origin from the erector spinae aponeurosis pass to correspondingly more rostral insertions. The most caudal of the thoracic fibres cover the most rostral of the lumbar fibres of the medial division but are distinguished from them by arising from the erector spinae aponeurosis rather than from the lumbar intermuscular aponeurosis. As the fibres in each lamina approach their level of insertion 35 ANA 13I 534 N. BOGDUK they divide into medial and lateral groups. Each medial group inserts into the tip and caudal border of a transverse process, while the lateral group inserts into the caudal border of the rib of the same level, between its angle and tubercle. The thoracic fibres of the medial division usually insert into the lower eleven ribs and transverse processes, but variations in this number can occur (see the description of longissimus dorsi by Poirier & Charpy, 1912). Lateral division of the lumbar erector spinae In the thoracic region the lateral division of the lumbar erector spinae lies lateral to the iliocostalis thoracis (Fig. 1), and in the lumbar region it lies lateral to the lumbar intermuscular aponeurosis or more strictly 'external' to the continuous envelope, formed by the erector spinae aponeurosis and the lumbar intermuscular aponeurosis, which covers the medial division (Fig. 6). Within the lateral division, lumbar and thoracic fibres are identifiable. The lumbar fibres are those which insert into lumbar vertebrae. The thoracic fibres insert into ribs. The lumbar fibres arise principally from the medial surface of the rostral end of the dorsal segment of the iliac crest. The area of origin is bounded medially by the attachment of the lumbar intermuscular aponeurosis and dorsolaterally by the attachment of the erector spinae aponeurosis. It extends ventrally almost as far as the attachment of the iliolumbar ligament (Fig. 2). The muscle fibres are arranged in four laminae, each directed to a particular lumbar vertebra (Fig. 9). The deepest lamina passes rostrally and ventrally to the L4 transverse process. Successively more superficial laminae pass rostrally, ventrally, and medially to the L3 to LI transverse processes respectively. The more lateral fibres describe a curve, convex laterally, which produces the rounded lateral contour of the lumbar erector spinae. Each lamina inserts into the dorsal aspect of the tip of its transverse process by way of a stout musculotendinous attachment (Fig. 7). Some of the most lateral fibres insert into the dorsal and middle layers of the thoracolumbar fascia and thereby reach their transverse process. The most medial fibres in each lamina do not arise directly from the ilium but arise from the lateral surface of the lumbar intermuscular aponeurosis. The attachment, however, is more oblique and not as dense as that of the fibres which arise from the medial surface (Fig. 9). The areas of insertions of the lumbar fibres of the lateral division on the L1-L4 transverse processes are separated from the insertions of the lumbar fibres of the medial division by the fat-filled space ventral to the lumbar intermuscular aponeurosis (Figs. 6, 7). This space, therefore, maintains, deeply, the separation into medial and lateral divisions which was afforded more superficially by the lumbar intermuscular aponeurosis. The thoracic fibres of the lateral division of the lumbar erector spinae arise from the erector spinae aponeurosis and lie superficial to the lumbar fibres of the lateral division. Lateral to the dorsal edge of the lumbar intermuscular aponeurosis, fleshy fibres arise from the rostral ends of the tendinous fibres of the erector spinae aponeurosis. Medial to the lumbar intermuscular aponeurosis, fleshy fibres arise from the dorsal surface of the erector spinae aponeurosis, where it covers the thoracic fibres of the medial division. The thoracic fibres of the lateral division pass rostrally to the 12th to 7th (or 6th) ribs. Those fibres to the 12th rib are the most lateral. Those to successively more rostral levels arise progressively more medially across the erector spinae aponeurosis. Those fibres arising from the dorsal surface of the erector spinae aponeurosis pass to the 7th and 6th ribs. The fibres insert into F"l J ne lumbar erector spinae 535 their respective ribs by way of musculotendinous attachments into the inferior border of the angle of the rib. DISCUSSION Although separable into lumbar and thoracic on the basis of their insertion levels, the lumbar and thoracic fibres of the medial division of the lumbar erector spinae constitute a single muscle. Since the erector spinae aponeurosis and the lumbar intermuscular aponeurosis form a continuous aponeurotic envelope around the medial division, the areas of origin of the lumbar and thoracic fibres are continuous with one another across the deep (concave) surface of this envelope. There is no line of demarcation between the areas on the medial surface of the lumbar intermuscular aponeurosis for the lumbar fibres, and the area on the deep surface of the erector spinae aponeurosis for the thoracic fibres. Thus, all the fibres of the medial division arise from a single, albeit curved, continuous area. Although inserting at different vertebral levels, the insertions of the lumbar and thoracic fibres are homologous to one another. The accessory processes of the lumbar vertebrae represent the 'true' transverse processes, while the so-called transverse processes are the 'costal elements' of the lumbar vertebrae, representative of the ribs. Comparison of the insertions of the lumbar and thoracic fibres of the medial division reveals that at each segmental level both lumbar and thoracic fibres insert into a true transverse process and the proximal end of the adjacent costal element. Thus, from lumbar to thoracic levels, the medial division has a continuous series of homologous insertions. Having both a single continuous area of origin and a continuous series of homologous insertions, the lumbar and thoracic fibres of the medial division must be interpreted as constituents of a single muscle. Similarly, the lumbar and thoracic fibres of the lateral division constitute a single muscle. Their origin is a continuous area extending from the ilium, across the lateral surface of the lumbar intermuscular aponeurosis and onto the dorsal surface of the erector spinae aponeurosis. Across this area of origin there is no demarcation between the areas for the lumbar and thoracic fibres. Thus, all the fibres of the lateral division arise from a single continuous area. The insertions of the lumbar and thoracic fibres are homologous. At both lumbar and thoracic levels the fibres insert at each segmental level into costal elements distal to the area of insertion of the medial division. Having both a continuous area of origin and a continuous series of homologous insertions, the component fibres of the lateral division, like the medial division, must be considered as constituting a single muscle. The two divisions of the lumbar erector spinae are clearly separated morphologically, by the iliocostalis thoracis in the thoracic region, and by the erector spinae aponeurosis and the intermuscular aponeurosis in the lumbar region. Accordingly, the medial and lateral divisions of the lumbar erector spinae should be considered, in both the thoracic and lumbar regions, as separate muscles. This is not the interpretation given in the textbooks. For comparison. the description of Gray's Anatomy (1973) is taken as the standard. This description, apart from changes to modern nomenclature, has not altered from those of early editions of Gray (1872, 1880, 1909). Previous authors (Bryce, 1923; Cleland & Mackay, 1896; Le Double, 1897; Piersol, 1918; Spalteholz, 1923; Tandler, 1919; Testut, 1904; Winckler, 1948; Wood-Jones, 1950) and other modem authors (Basmajian, 1971; Etemadi, 1963; Gardner, Gray & O'Rahilly, 1975; Hamilton, 1976; Lockhart, 35-2 536 N. BOGDUK Hamilton & Fyfe, 1969; Pansky & House, 1969; Romanes, 1972; Sinelnyikov, 1972) presented similar or even more abridged descriptions. Indeed, Last (1963), Tobias & Arnold (1967), Arnold (1968) and Hollinshead (1969) did not describe any form of lumbar insertion for either longissimus thoracis or iliocostalis lumborum. Conspicuously, in the standard description, there is no mention of the lumbar intermuscular aponeurosis or an equivalent. The iliocostalis lumborum and longissimus thoracis are distinguished by their eventual insertions in the thoracic region but no distinction is made of their different origins. The standard description denies an insertion of the iliocostalis lumborum into lumbar vertebrae. The longissimus thoracis is said to occupy the entire dorsal surface of the lumbar transverse processes. Winckler (1948) did describe two separate insertions on the transverse processes, as in the present study, but he ascribed both to longissimus thoracis. Upon comparing the standard description with that given in the present study, in the thoracic region, the medial and lateral divisions of the lumbar erector spinae obviously correspond to the longissimus thoracis and iliocostalis lumborum respectively. Because of their insertions into the lumbar accessory processes, the lumbar fibres of the medial division correspond to the lumbar portion of the longissimus thoracis. Thus, the medial division is clearly longissimus thoracis. However, the standard description implies that the lumbar fibres of the lateral division, since they insert into lumbar transverse processes, should also belong to the longissimus thoracis, since only the longissimus is said to insert into the entire dorsal surfaces of the lumbar transverse processes. However, this is not consistent with the morphology of these fibres. The lumbar fibres of the lateral division are clearly separated from the longissimus thoracis by the lumbar intermuscular aponeurosis and the fat-filled space ventral to it. This space also separates their insertions onto the transverse processes from those of the longissimus. Moreover, the origin of the lumbar lateral fibres is continuous with that of the thoracic lateral division, viz. iliocostalis, and their insertions are homologous with those of the thoracic lateral division. Thus, the lumbar fibres of the lateral division cannot be considered as part of the longissimus. They are more accurately interpreted as parts of the iliocostalis lumborum. The interpretation suggested by the present study is that the common muscle mass of the lumbar erector spinae is divided into medial and lateral divisions by an aponeurotic envelope formed by the erector spinae aponeurosis and the lumbar intermuscular aponeurosis. The medial division is the longissimus thoracis, which arises from the ilium and from the medial surface of the lumbar intermuscular aponeurosis and the deep surface of the erector spinae aponeurosis, and which has a series of insertions into transverse and costal elements of both lumbar and thoracic vertebrae. The lateral division is the iliocostalis lumborum, which arises from the ilium and external surfaces of the erector spinae aponeurosis and the lumbar intermuscular aponeurosis, and has a series of insertions into the lower six or seven ribs and the costal elements of the first four lumbar vertebrae. Interestingly, with respect to comparative anatomy, this interpretation is more consistent with the veterinary literature than is the standard human description. In the cat (Bogorodsky, 1930; Reighard & Jennings, 1934; Bogduk, 1973, 1980; Carlson, 1978), dog (Miller, Christensen & Evans, 1964; Bogduk, 1973) and monkey (Bogduk, 1973; Osman Hill, 1974) the lumbar erector spinae is divided by a lumbar intermuscular septum, homologous to the lumbar intermuscular aponeurosis. This The lumbar erector spinae Cat Dog Monkey Man 537 Fig. 10. Diagrammatic transverse sections illustrating the comparative anatomy of the dorsal lumbar muscles and lumbar intermuscular septa (adapted from Bogduk, 1973). Note how, in man, the lumbar intermuscular aponeurosis is rotated laterally and the area of insertion of the iliocostalis is displaced relatively more laterally. x, dorsal layer of thoracolumbar fascia; y, erector spinae aponeurosis; s, lumbar intermuscular spetum; lia, lumbar intermuscular aponeurosis; il, iliocostalis lumborum; II ,lumbar fibres of longissimus; m, multifidus; i, intertransversarii mediales; sc, sacrocaudalis dorsalis lateralis; ap, accessory process; j, zygapophysial joint. structure is used to define the longissimus thoracis and iliocostalis lumborum in these animals, and the iliocostalis is found to insert into the lumbar transverse processes. In the literature referring to man, only Poirier & Charpy (1912) have noted the lumbar intermuscular aponeurosis. They called it the 'cloison aponevrotique sagittale' and used it, as in the present study, to define the origins of the iliocostalis and longissimus. They highlighted the homology between the lumbar and thoracic fibres of iliocostalis lumborum. Rouviere (1927) and Spalteholz (1923), although not describing a 'cloison' or a lumbar intermuscular aponeurosis, nevertheless described the 'correct' insertions of the iliocostalis lumborum. Braus (1954) illustrated the lumbar insertions of the iliocostalis but only at the LI and L2 levels. Yet he too stressed the homology. As it has not been described in detail previously, the lumbar intermuscular aponeurosis deserves some further comment. This aponeurosis appears to be homologous to the lumbar intermuscular septum of the cat, dog and monkey (loc. cit.). In these animals the lumbar intermuscular septum has an origin, and a relationship to the erector spinae aponeurosis and to the longissimus and iliocostalis similar to those of the lumbar intermuscular aponeurosis in man (Fig. 10). The only major difference is its orientation. In man, the lumbar intermuscular aponeurosis appears to have rotated laterally. In association with this, the insertion of the iliocostalis lumborum has moved laterally, occupying a relatively smaller area on the lumbar transverse processes (Fig. 10). 538 N. BOGDUK In man, the lumbar intermuscular aponeurosis is a ventral reflection of the erector spinae aponeurosis which provides an extensive area of common attachment for the lumbar portions of longissimus and iliocostalis. It is more intimately associated with the longissimus, for, while only a few fibres of iliocostalis arise from it, all the L1-L4 fibres of the longissimus arise exclusively from it. Through the lumbar intermuscular aponeurosis, this considerable number of fibres eventually gains attachment to the ilium, but to a rather limited area on the dorsal segment of the iliac crest. Morphologically, the lumbar intermuscular aponeurosis appears to 'economise' the origin of the longissimus thoracis. Biomechanically, through the aponeurosis, all the LI-L4 fibres of the longissimus exert their pull on a rather small area on the ilium. Clinically, the area of attachment of the lumbar intermuscular aponeurosis to the ilium underlies a common site of tenderness in patients with acute lumbar strain. An entertainable hypothesis is that rather than a nonspecific 'muscle strain' this tenderness may indicate tear or avulsion of the aponeurosis caused by sudden strain of the longissimus thoracis, an injury analogous to 'tennis elbow'. Apart from their purely morphological significance, the details presented in this study are of particular relevance in the biomechanical analysis of forces acting on the lumbar spine. Using the traditional descriptions of the lumbar erector spinae, previous authors have made erroneous or oversimplified analyses of the forces exerted by this muscle. Hutton, Stott & Cyron (1977) stated that the longissimus acts upwards on the lumbar accessory processes. The reverse is true. Farfan (1973) included the 'thoracic' fibres of iliocostalis lumborum in his analysis of muscles acting on the lumbar spine but neglected the 'lumbar' fibres of both longissimus and iliocostalis. It is important to realise that the lumbar erector spinae does not simply span the lumbar region like a 'bowstring' between the sacrum and thorax. The concavity of the lumbar lordosis is filled in by a laminated series of relatively short muscle fibres connecting the ilium to the lumbar transverse processes, with many acting through the lumbar intermuscular aponeurosis. Apart from a rostrocaudal orientation, these fibres, particularly at the L5 level, have a considerable dorsoventral orientation, With such a variety of lengths and orientations one cannot simplify the action of the lumbar erector spinae into a single force. Any analysis must consider this muscle as a continuum of independent fibres, each with a unique orientation. Finally, the laminated nature of the lumbar fibres of the erector spinae suggests that those to a given vertebra could act independently. If such action occurred at the L5 level, for example, the effect would be to extend and retract the L5 vertebra, tending to diminish the lumbar lordosis. It is suggested that such segmental spasm of either the lumbar erector spinae or multifidus (or both) is the mechanism underlying the 'loss of lordosis' commonly seen in patients with low back pain. SUMMARY In the lumbar region the longissimus thoracis and iliocostalis lumborum are separated by the erector spinae aponeurosis and its ventral reflection - the lumbar intermuscular aponeurosis. Lumbar fibres of the longissimus arise from the ilium and the lumbar intermuscular aponeurosis and insert into the accessory processes and proximal ends of the transverse processes of the lumbar vertebrae. The lumbar erector spinae 539 Lumbar fibres of iliocostalis insert into the costal elements of the first four lumbar vertebrae. The lumbar insertions of these muscles are homologous to their thoracic insertions. The lumbar intermuscular aponeurosis is homologous to the lumbar intermuscular septum in the dog, cat and monkey. The details of attachment of the lumbar fibres of the erector spinae and of the lumbar intermuscular aponeurosis should be taken into account in biomechanical analyses of the lumbar vertebral column. The author extends his thanks to Professor F. W. D. Rost of the Department of Anatomy, University of New South Wales, and Professor M. J. Blunt of the Department of Anatomy, University of Sydney, for providing the facilities used in performing this and related studies. The assistance of Mrs F. Rubiu, Mr J. Van Loedersloot and the Department of Medical Illustration, University of New South Wales, in the preparation of the illustrations and photographs in this paper is gratefully acknowledged. REFERENCES ARNOLD, M. (1968). Reconstructive Anatomy. Philadelphia: W. B. Saunders. BASMAJIAN, J. V. (1971). (Ed.). Grant's Method of Anatomy. Baltimore: Williams & Wilkins. BOGDUK, N. (1973). The comparative anatomy of the dorsal lumbosacral region. Thesis, University of Sydney. BOGDUK, N. (1980). The dorsal lumbosacral muscles of the cat. Anatomischer Anzeiger 148, 55-67. BOGORODSKY, B. W. (1930). Der laterale Strange der Dorsalmuskulatur bei den fleischfressenden Tieren. Anatomischer Anzeiger 69, 83-121. BRAUS, H. (1954). Anatomie des Menschen. Berlin: Springer-Verlag. BRYCE, T. H. (1923). Myology. Vol. IV, Pt. Il. In Quain's Elements of Anatomy. 11th edition, (ed. E. S. Schafer, J. Symington & T. H. Bryce). New York: Longmans, Green & Co. CARLSON, H. (1978). Morphology and contraction properties of cat lumbar back muscles. Actaphysiologica scandinavica 103, 180-197. CLELAND, J. & MACKAY, J. V. (1896). Human Anatomy. General and Descriptive. Glasgow: MacLehose. ETEMADI, S. A. (1963). Observations on the musculature and innervation of the human back. M.Sc. Thesis, University of London. FARFAN, H. F. (1973). Mechanical Disorders of the Low Back. Philadelphia: Lea & Febiger. GARDNER, E., GRAY, D. J. & O'RAHILLY, R. (1975). Anatomy. 4th edition. Philadelphia: Saunders. GRAY, H. (1872). Anatomy. Descriptive and Surgical. 6th edition (ed. T. Holmes). London: Longmans, Green & Co. GRAY, H. (1880). Anatomy. Descriptive and Surgical. 9th edition (ed. T. Holmes). London: Longmans, Green & Co. GRAY, H. (1909). Anatomy. Descriptive and Applied. 17th edition (ed. R. Howden). London: Longmans, Green & Co. GRAY, H. (1973). Anatomy. Descriptive and Applied. 35th edition (ed. R. Warwick & P. L. Williams). London: Longmans. HAMILTON, W. J. (1976) (Ed.). Textbook of Human Anatomy. 2nd edition. London: Macmillan. HOLLINSHEAD, W. H. (1969). Anatomy for Surgeons. Vol. 3. Tne Back and Limbs. New York: Harper & Row. HUTTON, W. C., STOTT, J. R. R. & CYRON, B. M. (1977). Is spondylosis a fatigue fracture? Spine 2, 202-209 LAST, R. J. (1963). Anatomy-Regional and Applied. 3rd edition. London: Churchill. LE DOUBLE, A. F. (1897). Traite des Variations du Systeme Musculaire de l'Homme. Paris: Schleicher Freres. LOCKHART, R. D., HAMILTON, G. F. & FYFE, F. W. (1969). Anatomy of the Human Body. 2nd edition. London: Faber. MILLER, M. E., CHRISTENSEN, G. C. & EVANS, H. E. (1964). Anatomy of the Dog. Philadelphia: Saunders. OSMAN HILL, W. C. (1974). Primates. Comparative Anatomy and Taxonomy. Vol. VII. Cynopithecinae. Edinburgh: University Press. PANSKY, B. & HOUSE, E. L. (1969). Review of Gross Anatomy. 2nd edition. London: Macmillan. PIERSOL, G. A. (1918). Human Anatomy. 7th edition. Philadelphia: Lippincott. 540 N. BOGDUK POIRIER, P. & CHARPY, A. (1912). Traite d'Anatomie Humaine. Tome 1IT. Fasc. I. 3rd edition. Paris: Masson. REIGHARD, J. & JENNINGS, H. S. (1934). Anatomy of the Cat. 3rd edition. New York: Holt. ROMANES, G. J. (1972) (Ed.). Cunningham's Textbook of Anatomy. 11th edition. London: Oxford University Press. ROUVIERE, H. (1927). Anatomie Humaine. Tome I. 2nd edition. Paris: Masson. SINELNYIKOV, R. D. (1972). Atlas Anatomii Cheloveka. Moscow: Meditsina. SPALTEHOLZ, W. (1923). Hand Atlas of Human Anatomy. 4th edition. Philadelphia: Lippincott. TANDLER, J. (1919). Lehrbuch der systematischer Anatomie. Leipzig: Vagel. TESTUT, L. (1904). Traite d'Anatomie Humaine. 5th edition. Paris: Doin. TOBIAS, P. V. & ARNOLD, M. (1967). Man's Anatomy. Vol. 3. 2nd edition. Johannesburg: Witwatersrand University Press. WINCKLER, C. (1937). La structure du muscle spinalis dorsi chez l'homme. Archives d'Anatomie, d'Histologie et d'Embryologie 23, 183-206. WINCKLER, G. (1948). Les muscles profonds du dos chez l'homme. Archives d'Aratomie, d'Histologie et d'Embryologie 31, 1-58. WOOD-JONES, F. (1950) (Ed.). Buchanan's Manual of Anctomy. 8th edition. London: Balliere, Tindal & Cox.