Survey
* Your assessment is very important for improving the workof artificial intelligence, which forms the content of this project
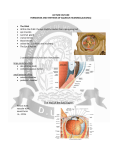
ANATOMY AND PATHOPHYSIOLOGY OF AQUEOUS PRODUCTION AND OUTFLOW* Thomas F. Freddo, O.D., Ph.D. , F.A.A.O. Professor and Director School of Optometry University of Waterloo The following is an overview and introduction to the basic anatomy and physiology of aqueous humor production and drainage. It is not meant to be comprehensive. Anatomy of the Ciliary Body - Intermediate portion of the uvea, Functions include aqueous production, facilitating aqueous drainage and accommodation. Gross Anatomy - Subdivided into 2 regions. Anteriorly the pars plicata, named for the numerous fin-like ciliary processes surrounding the lens. Posteriorly, the ciliary body has a flat inner surface, the pars plana. Histology Ciliary Epithelium: Two layers of epithelial cells -non-pigmented ciliary epithelium (closest to posterior chamber) - pigmented ciliary epithelium faces ciliary body stroma. Ciliary body stroma : a loose, vascularized connective tissue (CT) fills core of ciliary processes, extending between ciliary muscle fibers. The CT spaces in the muscle are the pathway for uveoscleral outflow. Ciliary body microvasculature: the ciliary processes have a dual vascular supply. Anterior arteriole-, large leaky capillaries from which aqueous is made. Aqueous Humor Formation - Two non-independent steps Elaboration of a Plasma Filtrate from the Microvasculature of the Ciliary Body Filtration = fluid forced across a membrane by pressure. Stromal capillaries from the anterior arteriole are highly permeable, owing to fenestrations and lack of tight junctions between endothelial cells. Thus, they are freely permeable, not only to solute and ions, but to plasma proteins as well. Ions, fluid and small molecules of the filtrate leave the fenestrated capillaries driven by the hydrostatic pressure within them. Interstitial fluid pressure increases with IOP. Thus, moderate elevations of IOP can actually suppress aqueous inflow, decreasing IOP. Unfortunately, the compensation is insufficient to protect against elevated IOP in POAG, but still has clinical relevance. Formation and Secretion of Aqueous Humor from the Plasma Filtrate by the Ciliary Epithelium The formation of aqueous humor by the ciliary epithelium is dependent upon two forces: 1) hydrostatic pressure; 2) the oncotic pressure gradient across the ciliary epithelium. Diurnal fluctuation: flow rates highest just after awakening, lowest during sleep. The magnitude of this effect is significant (approximately a 50% decrease), emphasizing the importance of recording time of day when taking tonometric measurements of intraocular pressure. Critical Enzymes in Aqueous Production a) Na+/K+ ATPase Na+/K+ ATPase is the enzyme that drives the Na+/K+ exchange pump. A currently accepted model of this pump (Post-Albers) proposes that the enzyme switches successively between two conformations within the cell membrane. The conformation presented to the cytoplasmic surface exhibits a high affinity for sodium but a low affinity for potassium. The affinities of the alternate conformation, presented to the extracellular surface under normal conditions, are opposite. By successively switching from one conformation to the other, in a cyclical fashion, 3 sodium ions are extruded and 2 potassium ions are imported per ATP molecule. The exchange is thus not electrically neutral and results in a net outward movement of positive charge. Water follows the sodium to produce aqueous humor. Sodium-potassium ATPase activity has been localized in the non-pigmented ciliary epithelium and inhibition of this enzyme with ouabain reduces aqueous secretion. b) Carbonic Anhydrase Carbonic anhydrase plays a central but incompletely understood role in aqueous secretion by generating bicarbonate ions. This enzyme has been localized solely to the non-pigmented layer at the tips of the ciliary processes. The enzyme accelerates the hydration of carbon dioxide to carbonic acid, which dissociates into hydrogen and bicarbonate ions according to the following equation: + H20 + CO2 <--> H2CO3 <--> HCO3 + H Inhibition of carbonic anhydrase decreases not only the entry of bicarbonate ions into the posterior chamber, but also sodium ions, though a direct linkage remains, unclear. One theory regarding the role played by bicarbonate ions in aqueous humor production maintains that HCO3(-) is transported in parallel with Na+, primarily to offset the net outward movement of cations by the Na+/K+ ATPase pump. Receptor Systems Related to Aqueous Production Various receptors have been identified in the ciliary body of various species, The best understood thus far are the alpha and beta adrenergics. Beta Adrenergic Receptors Beta-adrenergic receptors have been localized to the ciliary epithelium. Stimulation of these receptors initiates a signal transduction cascade beginning with activation of a regulatory intramembranous G-protein which then activates the second messenger adenylyl cyclase leading to elevation of cyclic-AMP levels resulting in phosphorylation of protein kinase-A. Phosphorylation of protein kinase-A may directly affect the rate of the Na-K-pump and thus aqueous secretion rate. Beta-antagonists (aka blockers) such as timolol maleate lead to reductions in both aqueous inflow and intraocular pressure. Alpha Adrenergic Receptors Alpha-2 adrenergic receptors are also present in the ciliary body. They are also coupled through G-proteins to adenylyl cyclase, but in a negative fashion that serves to block the elevations of cyclic-AMP levels induced by beta-agonists. Mechanisms remain unclear since both alpha- adrenergic agonists and antagonists have been reported to lower intraocular pressure. Only agonists are commercially available for IOP reduction. Aqueous Outflow The outflow of aqueous humor occurs predominately through 2 pathways, trabecular outflow, which is pressure-dependent and uveoscleral outflow, which is pressure-independent. Anatomy of the Trabecular Meshwork The trabecular meshwork (TM) is a wedge-shaped, band of tissue encircling and bridging the anterior chamber angle. The apex of this wedge is attached to Schwalbe’s line, the peripheral edge of Descemet’s membrane. Expanding posteriorly, the trabecular meshwork attaches to the stromas of the ciliary body and the peripheral iris. Indeed, thin strands of pigmented tissue, called iris processes, often rise from the peripheral iris to meet the trabecular meshwork. Projecting into the base of this wedge is a shelf-like projection of sclera termed the scleral spur Attached to the posterior surface of the scleral spur are the smooth muscle fibers of the longitudinal bundle of the ciliary muscle. When these fibers contract, they pull the spur posteriorly. In doing so, the layers of the corneoscleral meshwork attached to the anterior surface of the spur are spread apart. It is generally assumed that opening the meshwork in this fashion facilitates aqueous drainage and represents the most likely basis for the use of miotics in increasing aqueous outflow to reduce IOP in glaucoma. The Trabecular Meshwork is divided into: Uveal meshwork- structured as endothelial cell-lined beams running from Schwalbe's line to stroma of ciliary body (closer to AC than line on photo-above) Corneoscerlal meshwork - structured as endothelial cell-lined beams running from Schwalbe's line to stroma of ciliary body Closer to Schlemm's canal than line on photo-above). Cribriform or juxtacanalicular meshwork: open connective tissue between corneoscleral beams and inner wall of Schlemm's canal (area of squiggly line on photo above) The Gonioscopic View The principal landmarks evident in a macroscopic view of an open anterior chamber angle include, from superior to inferior, 1) Schwalbe's line -SL (i.e. the peripheral terminus of Descemet's membrane), 2) the trabecular meshwork overlying Schlemm's canal (SC), 3) the scleral spur (SS) and 4) the ciliary body band (CBB). The trabecular meshwork from Schwalbe's line to the scleral spur can be further sub-divided into a minimally pigmented anterior meshwork and a more heavily pigmented posterior meshwork. (see gonio-photo below - courtesy Dr. Rod Gutner). In sagittal sections the anterior portion of the meshwork corresponds to the portion of the meshwork that has no Schlemm’s canal external to it. As a result, this portion of the meshwork sees little aqueous flow and thus its trabecular endothelial cells phagocytose relatively little pigment. It is this anterior, low-flow portion of the meshwork that is ideally targeted in argon laser trabeculoplasty. Extracellular Matrix of the Meshwork For years it was assumed that a gel of hyaluronic acid filled the "open" spaces of the trabecular meshwork, creating the resistance of aqueous outflow that generates normal intraocular pressure. It was also assumed that a progressive increase in such a gel likely caused the increased resistance that characterizes POAG. More recent studies do not support this view since hyaluronate is not found in the open spaces of the meshwork, except in the juxtacanalicular region and contrary to the common theory, hyaluronate levels in the meshwork decrease with age and decrease even further in POAG! Other extracellular matrix proteins have more recently been examined. Two groups of compounds produced by the meshwork that may be involved in regulation of outflow are the matrix metalloproteinases (MMPs) and their inhibitors (TIMPs - tissue inhibitors of metalloproteinases). MMPs are enzymes used by an array of connective tissues to initiate and regulate turnover of extracellular matrix. MMPs, are made in the trabecular meshwork and degrade ECM. TIMPs, also made by the meshwork, serve to limit MMP activity. Whether an imbalance in these forces represents a contributing or initiating event causing a build-up of matrix that blocks the meshwork in POAG remains to be established. Most recently, a novel protein, induced by exposure of trabecular cell cultures to glucocorticoids has been identified, along with its gene. This protein, called TIGR (trabecular meshwork inducible glucocorticoid response (aka myocilin) is a glycoprotein that increases in amount in response to glucocorticoid exposure. The Canal of Schlemm The Canal of Schlemm is a continuous, circumferentially oriented channel, with direct connections to the venous system of the episclera. Despite this connection, blood is usually not seen in the canal unless intraocular pressure falls below that of episcleral venous pressure. The canal usually appears slit-like. The outer wall of Schlemm’s canal directly abuts the sclera. The inner wall of Schlemm’s canal faces the juxtacanalicular region of the meshwork. The inner wall of Schlemm’s canal exhibits unusual structures termed giant vacuoles, the formation of which are pressure-dependent but energy-independent. Very small openings called “pores” are found both in vacuoles and in non-vacuole containing areas of the inner wall. Pores are presumed to represent the actual flow pathways used by aqueous to traverse the inner wall of Schlemm’s canal. How they form remains uncertain. From Schlemm’s Canal to the Episclera From Schlemm’s canal, aqueous humor flows by one of two pathways to join the venous blood of the episclera. Elevations of episcleral venous pressure (e.g carotid-cavernous sinus fistula) result in proportional elevations of IOP. The primary pathway from Schlemm’s canal to the episcleral vasculature begins with collector channels that leave the outer wall and lead to a complex system of tortuous pathways that obliquely traverse the thickness of the sclera. During this tortuous passage, aqueous and blood are mixed. A smaller volume of aqueous humor bypasses the complex pathway outlined above by entering aqueous veins (of Ascher). Few in number, the aqueous veins lead from either the outer wall of Schlemm’s canal or an external collector channel, directly to the episclera and thus the blood and aqueous have not mixed by the time they reach the episclera. Aqueous veins are readily identified by biomicroscopy as blood vessels at the medual or lateral limbus appearing to contain red blood only at the edges of the vessel wall, with a clear stream in the middle. The clear stream is aqueous. Uveoscleral Outflow A smaller but nonetheless important fraction of total aqueous outflow leaves the eye in a pressure independent fashion. In this pathway aqueous passing below the scleral spur enters the ciliary body band. The aqueous passes through the strips of connective tissue that weave between the muscle fibers of the ciliary muscle, finally reaching the inner sirface of the sclera. This aqueous ultimately leaves through the sclera directly (uveoscleral outflow) or through the scleral emissaria provided for the vortex veins (uveovortex flow). Current estimates are that in the normal human eye, uveoscleral flow accounts for about 10% of the total. The uveoscleral pathway has been the subject of increased interest with the discovery that several prostaglandin analogs are capable of augmenting uveoscleral outflow, to reduce IOP, possibly by solubilizing the connective tissue matrix of the ciliary muscle allowing easier flow through this pathway, Fluid Mechanics of Trabecular Outflow - The Goldmann Equation It is clear that an array of basic factors, some known and some yet to be elucidated, all contribute to the measured intraocular pressure at any point in time. How the major factors involved in this process interact is, nonetheless, very useful. These major factors include the following: 1. the measured intraocular pressure 2. the pressure in the venous system of the episclera 3. the flow rate of aqueous humor 4. a factor that represents the ability of aqueous to leave the eye This latter factor is referred to as facility of outflow, which is the inverse of outflow resistance. These factors where first incorporated into simplified mathematical constructs by Goldmann as follows: P = IOP - Episcleral Venous Pressure, and Facility of trabecular outflow = Aqueous flow rate/ P where intraocular and episcleral venous pressures are measured in mm Hg and aqueous flow is in l/minute at steady state. These equations assume that trabecular outflow occurs in an entirely passive manner as a bulk flow down a pressure gradient. They also assume that all outflow is trabecular which, of course is not true but represents a convenient simplification. Under steadystate conditions, episcleral pressure varies very little and thus is a negligible variable. The facility of outflow through the trabecular meshwork is predominant. Under these circumstances the equations can be simplified as follows: Flowin = Flowout= C trab P + Flowuveoscleral Kaufman has provided an illustrative example of this modified equation using measured values for the normal human eye (Kaufman PL: Pressure-dependent outflow. In: The Glaucomas. Ritch R, Shields, MB and Krupin T, 2nd Ed, VI, Cptr 16, p 309, CV Mosby Co, St Louis, MO, 1996). Flow in = Flow out = C(trab) = 2.5 l/min 0.3 l/min/mmHg Intraocular pressure = Episcleral venous pressure = P = (16-9) = F(uveoscleral) = 16 mmHg 9 mmHg 7 0.4 l/min Substituting these values into the equation F (in) = F(out) = C (trab) P + F(uveoscleral) gives: 2.5 = 0.3(7) + 0.4 ****************************************************************************** * The material presented in this hand-out is modified and excerpted from several Chapters on the same topics written by the author in various glaucoma textbooks including those by Lewis and Fingeret, Sassani, Ritch, et al, Morrison and Leopold, in Duane's and on the Ophthalmic Hyperlink CME Website. Several of these are cited below as sources of supplemental reading. 1. Freddo, T.: "Anatomy and Physiology Related to Aqueous Humor Production and Outflow". Primarv Care of the Glaucomas, cptr 3, Fingeret, M. and Lewis, T., eds. Appleton and Lange, 1993 (and revised 1999). 2. Morrison, J.C., Van Buskirk, E.M. and Freddo, T. F.: "Anatomy, Microcirculation and Ultrastructure of the Ciliary Body". Glaucoma, v 1, cptr 3, Ritch, R., Shields, M.B., Krupin, T., eds. C.V. Mosby, 1989 (and revised 1994). 3. Freddo, T: "Anatomy and Pathophysiology of Aqueous Production and Outflow" in: Ophthalmic Fundamentals: Glaucoma. J. Sassani, ed., Slack, Inc., 1999. 4. Freddo T and Gong H: Anatomy of the Ciliary Body and Outflow Pathways, In: Duane’s Clinical Ophthalmology, E. Jaeger, ed. Cptr #43, 2002 (CD-ROM).