Survey
* Your assessment is very important for improving the work of artificial intelligence, which forms the content of this project
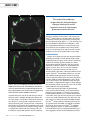
cover story Understanding Trabecular Meshwork Outflow Its role in modulating aqueous humor outflow and IOP. By Swarup S. Swaminathan; Dong-Jin Oh, P h D; Min Hyung Kang, P h D; Ramez I. Haddadin, MD; Guadalupe Villarreal J r , MD; Marc Töteberg-Harms, MD; Ayan Chatterjee; and Douglas J. Rhee, MD A queous humor outflow occurs through two routes, the conventional and uveoscleral pathways. The conventional pathway is responsible for approximately 85% of aqueous outflow.1 The primary constituent is the trabecular meshwork (TM), which consists of seven to eight layers of cellular beams surrounded by extracellular matrix (ECM) in human eyes. The aqueous humor traverses the superficial portions of the TM, known as the corneoscleral TM, to reach the juxtacanalicular connective tissue (JCT) TM, which is an amorphous layer of cells interspersed amongst ECM. The aqueous humor subsequently passes into Schlemm canal and enters the episcleral venous system. A The TM overall and especially the JCT TM are the anatomic location of the highest amount of outflow resistance in the conventional pathway.2,3 TM outflow is primarily mediated by two forces: (1) alterations in the ECM-surrounding cells and (2) alterations within the cells of TM and Schlemm canal’s inner wall that modulate cellular contractility and tension. PARACELLULAR OUTFLOW Numerous studies have demonstrated the impact of increased cellular stiffness on outflow resistance— specifically, an elevation in the number of actin stress fibers, actomyosin fiber bundles, and cross-linked actin B Figure 1. The effect of a Rho kinase inhibitor on actin filaments. Normal TM cells (A) versus TM cells treated with inhibitor (B). Reprinted with permission from Inoue T, Tanihara H. Rho-associated kinase inhibitors: a novel glaucoma therapy [published online ahead of print June 12, 2013]. Prog Retin Eye Res. doi:10.1016/j.preteyeres.2013.05.002. September/October 2013 glaucoma today 27 cover story A “The molecular pathways responsible for the pathologic changes leading to ocular hypertension and subsequent glaucoma remain elusive.” B Figure 2. The eyes of a wild-type control mouse (A) and a SPARC knockout mouse (B). The distribution of the fluorescent tracer demonstrates that outflow through the mouse TM is segmental but is more uniform in the secreted protein acidic and rich in cysteine (SPARC) knockout eye.16 networks within the cells of the TM and inner wall of Schlemm canal.4,5 Multiple compounds inhibit or hinder such changes. Latrunculins, molecules that disrupt actin filaments, cause TM cells to shrink and retract, thereby decreasing IOP.6 Ethacrynic acid, a compound that inhibits the formation of cytoskeleton, alters cellular shape by disrupting actin networks, leading to greater aqueous flow around cells and reducing IOP.7 The intracellular enzyme Rho regulates these intracellular 28 glaucoma today September/October 2013 changes in addition to alterations in cell-ECM interactions.5,8,9 If either Rho or a related enzyme, Rho kinase, is inhibited, TM cells relax significantly, increasing outflow and decreasing IOP (Figure 1).9,10 This discovery led to the development of Rho kinase inhibitors, a novel class of glaucoma medications currently being evaluated in clinical trials. If approved, these drugs will be the first to act directly on the TM to increase aqueous outflow since the release of pilocarpine in the 1870s. ALTERING ECM Changes in the ECM may also alter aqueous outflow. According to multiple research groups, ECM or ECMmodulating proteins appear to be crucial to mediating outflow. For example, gremlin mediates the effects of transforming growth factor-ß2 (TGF-ß2) on ECM deposition,11 whereas cochlin is an ECM protein that appears to sense shear stress in the TM.12 Myocilin influences interactions between TM cells and surrounding ECM,13 and sFRP-1 indirectly modulates ECM protein synthesis.14 Matricellular proteins are secreted and modulate ECM organization and the interaction between TM cells and the ECM. This family of proteins includes SPARC and thrombospondin-1 and 2, which have essential roles in the regulation of IOP and aqueous outflow.15,16 Both proteins appear to affect collagen fiber formation, which may alter the nature of the ECM around TM cells and change outflow. Other ECM-related alterations in glaucomatous eyes have been found. TGF-ß2, an essential molecule promoting tissue growth, is elevated in the aqueous humor of glaucomatous eyes.17,18 Studies have demonstrated that, when TGF-ß2 is overexpressed in cadaveric human eyes, ECM deposition increases within the TM and reduces the amount of aqueous outflow through the TM.19,20 Various proteins, including connective tissue growth factor and SPARC, are thought to mediate the TGF-ß2–driven increase in ECM.21,22 In addition, protein aggregates known as sheath-derived plaques have been found in the JCT TM of glaucomatous eyes.23 These aggregates contain several ECM proteins, includ- cover story ing elastin, collagen, and various proteoglycans. Studies have also shown that either increasing ECM production or decreasing ECM degradation increases IOP. When SPARC is overexpressed in perfused human eyes, the concentration of certain metalloproteinases (which catalyze the enzymatic degradation of ECM proteins) is decreased, whereas their inhibitors are upregulated.24 Metalloproteinases also play a role in reducing IOP via ECM degradation when activated by the adenosine receptor.25 Agonist compounds for this receptor are currently in clinical trials as potential therapeutic agents. Aqueous outflow does not occur consistently throughout all 360º of the TM. Rather, outflow occurs only in certain sections of the TM, a concept referred to as segmental flow.16,26,27 In mice lacking SPARC, IOP is decreased, and a reduction in IOP is correlated with an increase in the amount of area utilized for outflow (Figure 2).16,28 It appears as though the greater the available area for outflow through the TM, the lower the IOP. Segmental flow may explain why multiple iStent Trabecular Micro-Bypass Stents (Glaukos Corporation) are often required to achieve a substantial reduction in IOP in glaucomatous eyes. FINAL THOUGHTS The TM has been the recent focus of surgical innovation such as ab interno trabeculectomy (Trabectome; NeoMedix Corporation) and the iStent. Results thus far have been limited. Further elucidating the mechanics of TM outflow such as segmental outflow will be essential to identifying the pathophysiologic basis of primary open-angle glaucoma as well as to increasing the success rates of TM bypass procedures. Although investigators have begun to explain outflow physiology in the nonglaucomatous eye, the molecular pathways responsible for the pathologic changes leading to ocular hypertension and subsequent glaucoma remain elusive. Numerous research groups, including the authors’, have aimed at therapeutically inhibiting this disease process. There will be more soon! n Ayan Chatterjee is a medical student at the Perelman School of Medicine at the University of Pennsylvania in Philadelphia. Ramez I. Haddadin, MD, is a cornea fellow at the Massachusetts Eye and Ear Infirmary in Boston. Min Hyung Kang, PhD, is a research scientist at Case Western Reserve University in Cleveland. Dong-Jin Oh, PhD, is an assistant professor at Case Western Reserve University in Cleveland. Douglas J. Rhee, MD, is the chair of the Department of Ophthalmology and Visual Sciences at Case Western Reserve 30 glaucoma today September/October 2013 University in Cleveland. He is an ad hoc consultant to AqueSys, Inc., and Glaukos Corporation. Dr. Rhee may be reached at (216) 844-8590; [email protected]. Swarup S. Swaminathan is a medical student in the Harvard-Massachusetts Institute of Technology Division of Health Sciences & Technology at Harvard Medical School in Boston. Marc Töteberg-Harms, MD, is a clinical and basic research glaucoma fellow at Case Western Reserve University in Cleveland. Guadalupe Villarreal Jr, MD, is a resident at the Wilmer Eye Institute of Johns Hopkins University in Baltimore. 1. Bill A, Phillips CI. Uveoscleral drainage of aqueous humour in human eyes. Exp Eye Res. 1971;12:275-281. 2. Overby DR, Stamer WD, Johnson M. The changing paradigm of outflow resistance generation: towards synergistic models of the JCT and inner wall endothelium. Exp Eye Res. 2009;88:656-670. 3. Maepea O, Bill A. Pressures in the juxtacanalicular tissue and Schlemm’s canal in monkeys. Exp Eye Res. 1992;54:879-883. 4. Wettschureck N, Offermanns S. Rho/Rho-kinase mediated signaling in physiology and pathophysiology. J Mol Med (Berl). 2002;80:629-638. 5. Rao PV, Deng P, Sasaki Y, Epstein DL. Regulation of myosin light chain phosphorylation in the trabecular meshwork: role in aqueous humour outflow facility. Exp Eye Res. 2005;80:197-206. 6. Peterson JA, Tian B, Geiger B, Kaufman PL. Effect of latrunculin-B on outflow facility in monkeys. Exp Eye Res. 2000;70:307-313. 7. Erickson-Lamy K, Schroeder A, Epstein DL. Ethacrynic acid induces reversible shape and cytoskeletal changes in cultured cells. Invest Ophthalmol Vis Sci. 1992;33:2631-2640. 8. Rao PV, Deng PF, Kumar J, Epstein DL. Modulation of aqueous humor outflow facility by the Rho kinase-specific inhibitor Y-27632. Invest Ophthalmol Vis Sci. 2001;42:1029-1037. 9. Rao PV, Deng P, Maddala R, et al. Expression of dominant negative Rho-binding domain of Rho-kinase in organ cultured human eye anterior segments increases aqueous humor outflow. Mol Vis. 2005;11:288-297. 10. Tian B, Kaufman PL. Effects of the Rho kinase inhibitor Y-27632 and the phosphatase inhibitor calyculin A on outflow facility in monkeys. Exp Eye Res. 2005;80:215-225. 11. Wordinger RJ, Fleenor DL, Hellberg PE, et al. Effects of TGF-beta2, BMP-4, and gremlin in the trabecular meshwork: implications for glaucoma. Invest Ophthalmol Vis Sci. 2007;48:1191-1200. 12. Bhattacharya SK. Focus on molecules: cochlin. Exp Eye Res. 2006;82:355-356. 13. Resch ZT, Fautsch MP. Glaucoma-associated myocilin: a better understanding but much more to learn. Exp Eye Res. 2009;88:704-712. 14. Wang WH, McNatt LG, Pang IH, et al. Increased expression of the WNT antagonist sFRP-1 in glaucoma elevates intraocular pressure. J Clin Invest. 2008;118:1056-1064. 15. Haddadin RI, Oh DJ, Kang MH, et al. Thrombospondin-1 (TSP1)-null and TSP2-null mice exhibit lower intraocular pressures. Invest Ophthalmol Vis Sci. 2012;53:6708-6717. 16. Swaminathan SS, Oh DJ, Kang MH, et al. Secreted protein, acidic and rich in cysteine (SPARC)-null mice exhibit more uniform outflow. Invest Ophthalmol Vis Sci. 2013;54:2035-2047. 17. Inatani M, Tanihara H, Katsuta H, et al. Transforming growth factor-beta 2 levels in aqueous humor of glaucomatous eyes. Graefes Arch Clin Exp Ophthalmol. 2001;239:109-113. 18. Tripathi RC, Li J, Chan WF, Tripathi BJ. Aqueous humor in glaucomatous eyes contains an increased level of TGF-beta 2. Exp Eye Res. 1994;59:723-727. 19. Fleenor DL, Shepard AR, Hellberg PE, et al. TGFbeta2-induced changes in human trabecular meshwork: implications for intraocular pressure. Invest Ophthalmol Vis Sci. 2006;47:226-234. 20. Gottanka J, Chan D, Eichhorn M, et al. Effects of TGF-beta2 in perfused human eyes. Invest Ophthalmol Vis Sci. 2004;45:153-158. 21. Kang MH, Oh DJ, Rhee DJ. Regulation of SPARC by transforming growth factor β-2 in human trabecular meshwork. Invest Ophthalmol Vis Sci. 2013;54:2523-2532. 22. Fuchshofer R, Tamm ER. Modulation of extracellular matrix turnover in the trabecular meshwork. Exp Eye Res. 2009;88:683-688. 23. Tektas OY, Lutjen-Drecoll E. Structural changes of the trabecular meshwork in different kinds of glaucoma. Exp Eye Res. 2009;88:769-775. 24. Oh DJ, Kang MH, Ooi YH, et al. Overexpression of SPARC in human trabecular meshwork increases intraocular pressure and alters extracellular matrix. Invest Ophthalmol Vis Sci. 2013;54:3309-3319. 25. Avila MY, Stone RA, Civan MM. A(1)-, A(2A)- and A(3)-subtype adenosine receptors modulate intraocular pressure in the mouse. Br J Pharmacol. 2001;134:241-245. 26. Hann CR, Bahler CK, Johnson DH. Cationic ferritin and segmental flow through the trabecular meshwork. Invest Ophthalmol Vis Sci. 2005;46:1-7. 27. Lu Z, Overby DR, Scott PA, et al. The mechanism of increasing outflow facility by rho-kinase inhibition with Y-27632 in bovine eyes. Exp Eye Res. 2008;86:271-281. 28. Haddadin RI, Oh DJ, Kang MH, et al. SPARC-null mice exhibit lower intraocular pressures. Invest Ophthalmol Vis Sci. 2009;50:3771-3777.