Survey
* Your assessment is very important for improving the workof artificial intelligence, which forms the content of this project
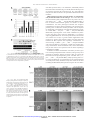
[CANCER RESEARCH 64, 7248 –7255, October 15, 2004] The Nuclear Factor B Subunits RelA/p65 and c-Rel Potentiate but Are Not Required for Ras-Induced Cellular Transformation Julie L. Hanson,1,2 Noel A. Hawke,1 David Kashatus,1,2 and Albert S. Baldwin1,2,3 1 Lineberger Comprehensive Cancer Center, 2Curriculum in Genetics and Molecular Biology, and 3Department of Biology, University of North Carolina, Chapel Hill, North Carolina ABSTRACT Extensive data indicate that oncoproteins, such as oncogenic H-Ras, initiate signal transduction cascades that ultimately lead to the activation of specific transcription factors. We and others have previously demonstrated that Ras activates the inherent transcriptional activation function of the transcription factor nuclear factor B (NF-B). Supportive of the importance of NF-B in transformation, Ras-induced cellular transformation can be suppressed by expression of IB␣, an inhibitor of NF-B, or by dominant-negative forms of the upstream activator IB kinase (IKK). However, conclusive evidence for a requirement for NF-B subunits in oncogenic transformation has not been reported. Furthermore, there is little understanding of the gene targets controlled by NF-B that might support oncogenic conversion. The data presented here demonstrate that, although both p65 and c-Rel enhance the frequency of Rasinduced cellular transformation, these NF-B subunits are not essential for Ras to transform spontaneously immortalized murine fibroblasts. Microarray analysis identified a set of genes induced by Ras that is dependent on NF-B for their expression and that likely play contributory roles in promoting Ras-induced oncogenic transformation. INTRODUCTION Proteins of the Ras family function as important signaling molecules in a variety of cellular processes, including growth, differentiation, and survival (1). Approximately 30% of human tumors have mutations in Ras alleles that result in constitutively active proteins (2). The oncogenic activation of Ras ultimately leads to the chronic stimulation of signaling cascades that promote activation of transcription factors involved in controlling proliferation, differentiation, and apoptosis (1). As an example, experiments have demonstrated that the transcription factor nuclear factor B (NF-B) is activated by oncogenic Ras. Expression of interfering proteins such as the inhibitor of NF-B (IB␣) or dominant-negative forms of IB kinase (IKK) suppresses Ras-induced transformation in vitro (3– 6), suggesting the involvement of NF-B in the transformation process induced by oncogenic Ras expression. NF-B represents a family of dimeric transcription factors that are characterized by a 300-amino-acid region called the Rel homology domain (7–9). The five mammalian NF-B family members are RelA/p65, RelB, c-Rel, p50/p105, and p52/p100, with the RelA/p65 and c-Rel subunits exhibiting strong transactivation potential. In unstimulated cells, IB proteins localize NF-B dimers to the cytoplasmic compartment through masking of nuclear localization sequences of NF-B subunits. Activation of NF-B can occur through various stimuli, including inflammatory cytokines and bacterial and viral infection, as well as oncogenic signals (7–9). Activation of NF-B, Received 12/16/03; revised 7/15/04; accepted 8/10/04. Grant support: Research support was provided by NIH grants RO1 CA73756 and RO1 CA75080 (A. Baldwin), and CA096418 (N. Hawke). We also gratefully acknowledge the Waxman Cancer Research Institute for research support. The costs of publication of this article were defrayed in part by the payment of page charges. This article must therefore be hereby marked advertisement in accordance with 18 U.S.C. Section 1734 solely to indicate this fact. Requests for reprints: Albert Baldwin, Lineberger Comprehensive Cancer Center, CB 7295, University of North Carolina, Chapel Hill, NC 27599. Phone: (919) 966-3652; Fax: (919) 966-0444; E-mail: [email protected]. ©2004 American Association for Cancer Research. through receptors such as tumor necrosis factor receptor 1 (TNFR1), interleukin 1 receptor (IL-1R), or various Toll-like receptors, initiates signal transduction cascades ultimately leading to activation of the IB kinase (IKK) complex. The IKK complex is composed of three subunits, IKK␣, IKK and IKK␥ (7–9). Both IKK␣ and IKK have inducible catalytic activity, whereas IKK␥ is a regulatory subunit. Following its activation, the IKK complex phosphorylates IB␣ and IB at specific serine residues, which targets IB for ubiquitination and degradation by a proteasome-dependent pathway. After IB degradation, the unmasked nuclear localization sequence of NF-B allows nuclear accumulation that, in turn, promotes sequence-specific DNA binding and transcriptional activation of target genes (9). Although nuclear accumulation of NF-B is a key step in the overall activation process controlled by receptor-dependent activation, posttranslational modifications on RelA/p65 also appear to be necessary for the transcriptional competence of NF-B. One such modification, phosphorylation on serine 276, has been shown to be required for stable interactions between RelA/p65 and the transcriptional coactivator CBP and for transcriptional activation of certain NF-B target genes (10, 11). Additional sites of phosphorylation have also been described that appear to contribute to the inherent transcriptional activity of NF-B (12–14). Additionally, Akt, which functions downstream of Ras and phosphatidylinositol 3⬘-kinase (PI3K), can control transcriptional activation function through a mechanism dependent on IKK function and on RelA/p65 phosphorylation, but in a manner that does not promote enhanced nuclear accumulation of NF-B (15, 16). Consistent with this, we have shown that both oncogenic Ras and oncogenic Raf activate an NF-B-dependent reporter in mouse fibroblasts without inducing DNA binding of NF-B as measured by gel mobility shift assay (4, 6). It is noted that human epithelial cells, transformed with oncogenic Ras, exhibit enhanced NF-B DNAbinding activity in association with IB␣ degradation (3). Certain transcription factors, including Ets and NF-B proteins, have been shown to be important for transformation in response to oncoprotein expression (4, 17, 18); however, it was unclear whether transforming proteins elicit a full range of transcription factor-dependent gene expression or whether a limited set of transcription factorregulated genes are induced during transformation. Recently, we demonstrated that oncogenic Ras, although stimulating a NF-Bdependent reporter, suppressed the ability of TNF to activate NF-B in murine fibroblasts (19). The mechanism of suppression of NF-B involved Ras-controlled inhibition of TNF-induced IKK activation and IB degradation. These studies revealed that Ras suppressed the ability of TNF to activate known NF-B-regulated genes such as iNOS, indicating that, although Ras can stimulate NF-B functional activity, it does so in a selective manner. Although it is suggested that NF-B plays a role in Ras-induced transformation, direct genetic evidence is missing. We previously reported that Ras stimulates the activity of the NF-B RelA/p65 transcriptional activation domain (5, 6). Although a specific role for c-Rel in Ras transformation has not been established, c-Rel does have documented transforming potential. Thus, c-Rel has been shown to transform chicken lymphoid cells (20) and recently mouse mammary tumor virus (MMTV)-c-Rel transgenic mice were shown to develop 7248 Downloaded from cancerres.aacrjournals.org on June 11, 2017. © 2004 American Association for Cancer Research. NF-B SUBUNITS POTENTIATE RAS TRANSFORMATION mammary tumors with a high frequency (21). In the present study, we have addressed the roles of the RelA/p65 and c-Rel subunits in controlling Ras-induced cellular transformation. Somewhat surprisingly, neither RelA/p65 nor c-Rel is absolutely required for Rasinduced transformation of immortalized murine fibroblasts. Despite this, each of these NF-B subunits potently enhances the efficacy of oncogenic transformation induced by Ras. We have extended these studies to identify a set of genes induced by Ras that require either p65 or c-Rel for their expression and presumably encode proteins that potentiate the Ras-induced transformation response. MATERIALS AND METHODS contamination. Reverse transcription-PCR (RT-PCR) was then preformed with the Qiagen One-Step RT-PCR kit with gene-specific primers for H-Ras and -actin. In a total volume of 25 L, 0.5 g of total RNA was used to analyze H-Ras expression, and 0.01 g was used to determine the level of -actin expression. Real-time PCR. Total RNA was isolated with the Promega SV total RNA kit. Two g of RNA was reverse transcribed with random hexamers and MMLV-RT (Invitrogen) according to the manufacturer’s protocol. The cDNA was then diluted 1:5 and was used for quantitative PCR. Primers were designed for each gene with consideration of intron/exon borders to favor only mRNAbased PCR products. Each PCR reaction contained 2 L of cDNA, a 2 mol/L concentration of each primer, and Qiagen QuantiTect Syber Green PCR master mix in a 20-L total volume. PCR was preformed in an ABI Prism 7900 sequence detection system (Applied Biosystems, Foster City, CA). All of the reactions were preformed in triplicate and were normalized to the number of copies of 18S detected in each sample. Affymetrix Microarray and Identification of NF-B-Regulated Genes. Seven g of total RNA were used to synthesize cDNA. A custom cDNA kit from Invitrogen was used with a T7-(dT)24 primer for this reaction. Biotinylated cRNA was then generated from the cDNA reaction with the BioArray High Yield RNA Transcript kit. The cRNA was then fragmented in fragmentation buffer [5⫻ fragmentation buffer: 200 mmol/L Tris-acetate (pH 8.1), 500 mmol/L KOAc, 150 mmol/L MgOAc] at 94°C for 35 minutes before the chip hybridization. Fifteen g of fragmented cRNA were then added to a hybridization cocktail (0.05 g/L fragmented cRNA, 50 pmol/L control oligonucleotide B2, BioB, BioC, BioD, and cre hybridization controls, 0.1 mg/ml herring sperm DNA, 0.5 mg/mL acetylated bovine serum albumin, 100 mmol/L 2-(N-morpholino)ethanesulfonic acid (MES), 1 mol/L Na⫹, 20 mmol/L EDTA, 0.01% Tween 20). Ten g of cRNA were used for hybridization. Arrays were hybridized for 16 hours at 45°C in the GeneChip Hybridization Oven 640. The arrays were washed and stained with R-phycoerythrin streptavidin in the GeneChip Fluidics Station 400. After this, the arrays were scanned with the Hewlett Packard GeneArray Scanner. Affymetrix GeneChip Microarray Suite 5.0 software was used for washing, scanning, and basic analysis. The results of these analyses were input into GeneSpring (Silicon Genetics, Redwood City, CA, http://www.silicongenetics.com/cgi/ Products/GeneSpring?index.smf)4 for comparative analysis of expression. The data were “double polished,” i.e., each chip was normalized with a distribution of all of the genes around the 50th percentile (arbitrarily a value of 1), and each individual oligonucleotide, or spot, was normalized across the eight samples by dividing the individual value at each spot by the average of the eight samples. The polished data set was reduced to only those genes that indicated at least four present or marginal calls of the eight samples. The data set was also filtered for absolute, nonnormalized values, of at least 10 (note 0.1 equals 0 or less signal detected) to 21,523 provide an indication of the range of abundances. Subsequent filters for errors (accepting ⬍1 SD between the replicates in two of four conditions) and confidence (accepting only probe sets with t test P values ⬍ 0.5 in two of four conditions) yielded the experimental data set. Applying a final filter for at least a 3-fold change in expression between any two conditions resulted in the final set of dynamically regulated genes, which was then used for cluster analysis with the GeneSpring software. Various algorithms were used to define clusters of genes with similar expression patterns. The cluster of genes reported was identified as Ras induced and NF-B dependent (correlation coefficient of 0.95). Cell Culture and Retroviral Infections. Spontaneously immortalized mouse embryo fibroblasts (MEFs)—wild-type (MEF⫹/⫹), p65⫺/⫺, cRel⫺/⫺, or p65⫺/⫺/cRel⫺/⫺ [p65/cRel double knockout (DKO)]—were maintained in DMEM (Invitrogen, Carlsbad, CA) supplemented with 10% donor calf serum (Invitrogen) and penicillin-streptomycin. MEF lines, stably expressing HRasV12 or vector control, were obtained by infecting various MEF lines with a retrovirus-encoding oncogenic H-Ras or with vector control virus and then selecting with 2 g/mL puromycin. Retrovirus was prepared by cotransfecting 293T cells with pVPack-Eco, pVPack GP (Stratagene) and either pBabe-Puro or pBabe H-RasV12. Twenty-four hours posttransfection, the medium on the 293T cells was changed to DMEM containing 2% fetal bovine serum, and the cells were transferred to a 32°C, 5% CO2 environment. Twenty-four and forty-eight hours later, the supernatant was collected, filter sterilized, and used to infect target cells. Retroviral infection of target cells was performed by incubating 1 ⫻ 105 target cells with 1 mL of virus, 1 mL of complete medium with 8 g/mL Polybrene for 2 to 3 hours and then adding 3 mL of complete medium and incubating the cells for 20 hours. Incubation of the target cells with retrovirus was then repeated approximately 24 hours after the initial infection. Forty-eight hours after the second infection, the cells were either split into puromycin selection for stable cell line or were harvested for RNA. Focus Formation Assays. All MEF cell lines were plated at 4 ⫻ 105 cells/60-mm plate 24 hours before transfection. The MEFs were then transfected with the indicated amounts of DNA with GenePorter2 (Gene Therapy Systems) according to the manufacturer’s protocol. Twenty-four hours posttransfection, the medium was changed and then was changed again every 3 to 4 days for two weeks. After 2 weeks, the cells were fixed with 10% acetic acid and 10% methanol and then were stained with a 4% crystal violet-10% methanol solution. Soft Agar Assays. MEF lines, stably maintaining the vector control or expressing H-RasV12, were suspended in a 0.33% bactoagar/DMEM layer at a concentration of 2 ⫻ 104 cells/mL and were plated over a 0.5% bactoagar/ DMEM layer in 6-well plates. Two weeks after plating the cells, the wells were analyzed for soft agar growth. Western Blot Analysis. Western blot analysis was performed by preparing whole cell extracts and then separating 30 to 50 g of total protein by SDS-PAGE. After transferring the separated proteins to nitrocellulose, blots were blocked in Tris-buffered saline-Tween (TBST) with 5% milk and then were incubated in primary antibody [p65 (Rockland), c-Rel, tubulin (Santa Cruz Biotechnology, Santa Cruz, CA)] for either 1 to 2 hours at room temperature or overnight at 4°C. The membranes were then washed in TBST and were incubated for 1 hour in antirabbit horseradish peroxidase-conjugated secondary antibody (Promega, Madison, WI) and were washed again. Protein RESULTS bands were visualized with enhanced chemiluminescence detection (Amersham Life Science). H-RasV12 Induces Transformation of p65ⴚ/ⴚ and c-Relⴚ/ⴚ Northern Blot Analysis. Total RNA was isolated with Trizol (Invitrogen) MEFs. Previous studies examining the involvement of NF-B in according to the manufacturer’s protocol. Ten g of RNA were then electro- Ras-induced transformation used dominant-negative forms of IKK or phoresed on a 1.5% formaldehyde-agarose gel and were transferred to a nylon the IB-super repressor (SR), the trans-dominant-negative inhibitor of membrane according to standard procedures. The RNA was then cross-linked NFB (3, 4). These interfering proteins either inhibit upstream sigto the membrane by UV irradiation (Stratagene) and were probed with a randomly labeled gene specific probe. Hybridization and washing were pre- naling to a variety of NF-B dimers or block activation of any NF-B formed with ExpressHyb (Stratagene, La Jolla, CA) according to the manu- subunit that will associate with IB family members. Use of dominant-negative IKK or IB-SR does not allow for the determination of facturer’s protocol. Reverse Transcription-PCR. Total RNA was isolated with Trizol (In4 vitrogen) according to the manufacturer’s protocol. Four g total RNA were Internet address: http://www.silicongenetics.com/cgi/Products/GeneSpring?index. then treated with RNase-free DNase (Invitrogen) to eliminate any DNA smf. 7249 Downloaded from cancerres.aacrjournals.org on June 11, 2017. © 2004 American Association for Cancer Research. NF-B SUBUNITS POTENTIATE RAS TRANSFORMATION a potential role of specific NF-B subunits in the transformation process. Additionally, overexpression of IB␣ or IB-SR can have effects on cells that are independent of NF-B (22, 23). Therefore, to determine whether NF-B subunits, specifically p65 and c-Rel, have essential roles in Ras-induced transformation, we used genetic knockout-derived MEFs. Spontaneously immortalized MEFs that lacked the p65 or c-Rel subunit of NF-B and that expressed oncogenic Ras were analyzed for the loss of contact inhibition through the use of focus formation assays. MEFs, either p65⫺/⫺ or c-Rel⫺/⫺, were transfected with vector control plasmid (pZip-neo), oncogenic H-Ras (pZip-HRasV12), or oncogenic H-Ras with an expression vector for either p65 or c-Rel. All of the transfections were done in quadruplicate for each condition. At 48 hours posttransfection, one plate was harvested for whole cell protein extraction and another was harvested for total RNA. These extracts were used as controls for protein expression of p65 and c-Rel and for expression of H-Ras RNA (Fig. 1C). At 14 days posttransfection, the two remaining plates were stained and were analyzed for focus formation. As shown in Fig. 1A and B, Ras alone was able to induce a low level of focus formation in both the p65⫺/⫺ and the c-Rel⫺/⫺ MEFs. This suggests that neither of these subunits alone is absolutely required for Ras-induced focus formation, but these experiments do not allow us to determine whether p65 and c-Rel are able to compensate for each other in facilitating Ras-induced transformation. Cotransfection of p65 and Ras into the p65⫺/⫺ MEFs, or c-Rel and Ras into the c-Rel⫺/⫺ MEFs resulted in an increase in focus formation over that of Ras alone (comparable with levels attained with wild-type cells; see ref. 4), indicating that both p65 and c-Rel can enhance Ras-induced cellular transformation (Fig. 1A and B). p65⫺/⫺ and c-Rel⫺/⫺ MEFs transfected with p65 or c-Rel, respectively, without Ras do not display any focus-forming activity (data not shown). Western blot analysis of whole cell extracts for p65 and c-Rel and RT-PCR analysis for H-Ras confirmed the concentration-dependent expression of the transfected proteins and also shows, importantly, that exogenous expression of p65 and c-Rel does not have an effect on expression of exogenous H-Ras (Fig. 1C). Thus, enhanced focus formation after p65 or c-Rel expression cannot be explained by simply inducing higher levels of H-RasV12 expression and must, instead, be derived from inherent p65 or c-Rel activity. The raw data presented in Fig. 1A suggest that p65⫺/⫺ cells have a higher transforming potential than do the c-Rel⫺/⫺ cells. This observation could be interpreted to indicate that c-Rel plays a more important role in Ras-induced focus formation than does p65. However, we have analyzed at least two independent spontaneously immortalized cell lines for each genotype and have found that not all cell lines have the same transforming potential as measured by focus formation assay. Some MEF cell lines (p65⫺/⫺ and c-Rel⫺/⫺) will not transform with or without cotransfection of either p65 or c-Rel, which indicates that the absence of p65 or c-Rel is not the reason for the lack of focus formation (data not shown). Furthermore, all MEF lines do not have equal transfection efficiency, presumably because of genetic alterations associated with immortalization. Therefore, it is not appropriate to directly compare the transforming potential of two different spontaneously immortalized cell lines. Thus, our analysis is limited to the analysis of p65-null, c-Rel-null, and double-null MEFs as well as the restored isogenic cell lines. H-RasV12 Induces Transformation of p65/c-Rel DKO MEFs. To further examine the role of p65 and c-Rel in Ras-induced transformation and to analyze the possibility that p65 and c-Rel serve redundant functions in transformation, we performed focus formation assays in p65/c-Rel DKO MEFs. Immunoblotting analysis reveals that Arf and p53 are inducible (by oncogenic Ras expression and by UV exposure, respectively; data not shown), which indicates that these Fig. 1. p65 and c-Rel enhance Ras-induced transformation in p65⫺/⫺ and cRel⫺/⫺ MEFs, respectively. A, focus formation assay of p65⫺/⫺ (top panel) and cRel⫺/⫺ (bottom panel) MEFs transfected with pBabe-Puro (Control) or H-RasV12 alone or with 1 or 2 g of pBabe-p65 (top panel) or pBabe-cRel (bottom panel). B, graphical representation of the number of foci/plate (in duplicate) in the focus formation assay shown in A. C, RT-PCR analysis for H-Ras and -actin (loading control) expression. The data shown are representative of three independent experiments. tumor suppressor proteins are wild-type in these cells (see Discussion). As shown above for the p65 and c-Rel knockout MEFs, oncogenic H-Ras alone was able to induce focus formation in the p65/cRel DKO MEFs (Figs. 2A and B), indicating that p65 and c-Rel are not essential for Ras-induced focus formation in MEFs. Importantly, p65 7250 Downloaded from cancerres.aacrjournals.org on June 11, 2017. © 2004 American Association for Cancer Research. NF-B SUBUNITS POTENTIATE RAS TRANSFORMATION Fig. 2. p65 and c-Rel enhance Ras-induced transformation in p65/c-Rel DKO MEFs. A, focus formation assays in p65/cRel DKO MEFs transfected with pBabe-Puro (Control) or pBabe-H-RasV12 alone or in combination with pBabe-p65, pBabe-c-Rel, or both. B, graphical representation of the number of foci/plate (in duplicate) from the focus formation assays shown in A. C, Immunoblotting for p65 and c-Rel (upper panel). RT-PCR analysis for H-Ras and -actin (loading control) expression (lower panel). The data shown are representative of three independent experiments. and c-Rel expression alone, or in combination, substantially enhance Ras-induced focus formation (Fig. 2A and 2B) without affecting levels of exogenous oncogenic Ras expression (Figs. 2C), again indicating that p65 and c-Rel contribute to the potential for Ras-induced transformation. MEFs Stably Expressing Oncogenic H-Ras Are Morphologically Transformed and Grow in Soft Agar. The results observed in the focus formation assays demonstrate that in the presence of oncogenic H-Ras, p65⫺/⫺, c-Rel⫺/⫺, and p65/cRel DKO MEFs display a loss of normal contact-inhibited growth, one measure of cellular transformation. Two other phenotypes associated with oncogenic transformation are altered cellular morphology and anchorage-independent growth. To determine whether p65 and/or c-Rel are required for morphologic transformation and anchorage-independent growth, we created MEF⫹/⫹ and p65⫺/⫺, c-Rel⫺/⫺, and p65/c-Rel DKO MEFs stably expressing either vector control or H-RasV12 by infecting the cells with a control retrovirus, pBabe-Puro, or a retrovirus expressing pBabe-HRasV12. At 48 hours postinfection, infected cells were selected for 14 days in growth medium supplemented with puromycin (1 g/mL). Drug-resistant colonies were then pooled and examined for Ras expression and morphologic transformation. As observed in Fig. 3A, MEF⫹/⫹, p65⫺/⫺, c-Rel⫺/⫺, and p65/c-Rel DKO cells expressing RasV12, each display a spindly and refractile phenotype characteristic of Ras-transformed cells, which demonstrates that p65 and c-Rel are not required for Ras to induce morphologic transformation of MEFs. A hallmark of cellular transformation is the ability of cells to grow in an anchorage-independent manner. Therefore, soft agar assays were performed to determine whether p65 and/or c-Rel are required for Ras-induced anchorage-independent growth. All MEF lines (⫹/⫹, p65⫺/⫺, c-Rel⫺/⫺, and p65/cRel DKO), stably expressing either vector control or H-RasV12, were suspended in 0.33% agar/medium and were analyzed for colony formation 14 days later. All of the cell lines (including the DKO MEFs) that expressed oncogenic Ras were able to Fig. 3. p65-, cRel-, and p65/cRel-null MEFs stably expressing H-RasV12 display morphologic transformation and anchorage-independent growth. A, morphologic transformation observed in ⫹/⫹, p65⫺/⫺, c-Rel⫺/⫺, and p65/cRel DKO MEFs stably expressing pBabe-H-RasV12 (bottom panel) compared with pBabe-Puro control cells (top panel). B, soft agar assay with MEFs described in A were suspended in 0.33% bactoagar growth medium and were incubated for 14 days at 37°C, 5% CO2. Fourteen days later, the cells were examined for colony formation indicative of anchorage-independent growth. The data shown are representative of three independent experiments, each performed in triplicate. 7251 Downloaded from cancerres.aacrjournals.org on June 11, 2017. © 2004 American Association for Cancer Research. NF-B SUBUNITS POTENTIATE RAS TRANSFORMATION Fig. 4. p65 and c-Rel do not contribute to Ras-induced expression of Cyclin D1 or c-myc. A, RNase protection assay for Cyclin D1 RNA levels. Total RNA isolated from ⫹/⫹ or p65/c-Rel DKO MEFs, infected with either control or HRasV12-expressing retrovirus, was incubated with probes for cyclin D1 and glyceraldehyde-3-phosphate dehydrogenase (GAPDH) (as an internal control) as described under Materials and Methods. B, total RNA (as described for A) was separated by electrophoresis in a formaldehyde-agarose gel and was analyzed by Northern blot with a 32P-labeled c-myc DNA probe. 18S RNA is included as a control for RNA loading. Data shown are representative of three independent experiments. form colonies in soft agar, whereas the vector control cells were unable to grow (Fig. 3B). Furthermore, the p65⫺/⫺ MEFs did not show transforming potential on their own as suggested in another study (24), although cell origin may explain the difference. Together these data indicated that p65 and c-Rel are not required for Rasinduced loss of contact-inhibited growth, morphologic transformation, or anchorage-independent growth. Determination of Ras-Induced Genes Controlled by p65 and/or c-Rel. The data thus provided indicated that, although p65 and c-Rel are not required for Ras-induced transformation, these NF-B subunits substantially enhance transformation. Genes that are known to be regulated by NF-B (at least in response to certain stimuli) include c-IAP1/2, TRAF1 and -2, cyclin D1, and c-myc (25). To determine whether p65 and c-Rel regulate cyclin D1 or c-myc expression in response to oncogenic Ras signaling, we infected MEFs⫹/⫹ or p65/ c-Rel DKO MEFs with either control or H-RasV12 retrovirus. At 48 hours postinfection, total RNA was isolated and Northern blot analysis was performed. As expected, oncogenic Ras induced the expression of cyclin D1 and c-myc in MEFs⫹/⫹. Interestingly, inducible expression of cyclin D1 and c-myc was largely unaffected in the p65/cRel DKO MEFs (Fig. 4), indicating that p65 and c-Rel do not contribute substantially to Ras-induced expression of these genes. Furthermore, at least in murine fibroblasts, we did not observe Rasinduced expression of c-IAP1/2 or TRAF1/2 (see Fig. 5B). These data are consistent with our recent report that Ras does not regulate certain NF-B-dependent genes (19). To determine which, if any, Ras-induced genes are NF-B (p65 and/or c-Rel)-dependent, we performed Affymetrix microarray analysis. Total RNA from ⫹/⫹ or p65/c-Rel DKO MEFs was isolated 48 hours post-retroviral infection with control or H-RasV12 virus. The RNA was then amplified and hybridized to the Affymetrix mouse gene array 430A, as described in Materials and Methods. Affymetrix analysis identified 899 differentially regulated genes with the criteria for inclusion (see Materials and Methods) defined by a 2-fold-orgreater change in expression among any two of the four samples. Of these genes, 135 were considered to be specifically Ras-inducible based on a 3-fold increase in Ras-expressing wild-type MEFs over control/wild-type MEFs irrespective of their regulation by RelA/p65 and c-Rel. Clustering algorithms also identified a single subset of 25 similarly regulated genes that represent p65/c-Rel-dependent, Rasinducible genes (Table 1; see below). This collective expression profile demonstrates a requirement for p65/c-Rel to exhibit induced gene expression on oncogenic Ras expression. The largest single cluster identified 88 genes, including c-myc, cyclin D1, HMGCI, and HMGY that were induced by Ras in both the wild-type and p65/cRel-null cells. A more refined clustering of these 88 genes demonstrated that the basal expression levels of 44 genes were unaffected by the loss of p65/c-Rel. The remaining 44 genes segregated into two additional profiles based on repressed or elevated basal expression in the DKO cells versus the wild-type MEFs. Therefore, we have iden- Table 1 Ras-inducible, NF-B-regulated genes Normalized fluorescent intensity† Gene name gb identifier Fold* DKO–H-ras DKO-PURO p65⫹/⫹ PURO p65⫹/⫹ H-ras P value‡ Serpinb2 Tslp Adra2a Tmod2 PCA-1§ Galactin7 Posh PI4P5K-I Or10 Psgl-1/Selp1 Aeg1 Pkp1 Tage4 EST Nppb Stk10 Col18a1 Bsf3 AldH1a3§ Klf3 Adra2c Cgref1 Sox5 RN19§ Oas16 NM_011111 NM_021367 NM_0074717 AK018223 BB560177 AF038562 NM_021506 BC003763 NM_020513 NM_009151 NM_009638 NM_019645 NM_009310 AV302409 NM_008726 NM_009288 NM_009929 NM_019952 BC026667 NM_008453 NM_007418 BC023116 AK015212 AK015966 AB067530 33.32 11.92 6.339 5.895 5.695 5.586 5.315 4.896 4.022 3.925 3.862 3.786 3.712 3.684 3.515 3.228 3.045 2.999 2.809 2.626 2.597 2.501 2.466 2.256 2.079 1.036 0.491 1.063 1.613 0.884 0.921 0.751 1.307 0.843 0.95 0.725 1.151 1.07 0.998 0.995 0.809 0.998 1.112 0.897 0.803 0.965 1.064 0.831 0.885 0.929 0.511 0.79 0.897 0.904 0.94 0.777 0.743 0.926 0.655 0.965 1.004 0.84 0.86 0.905 0.998 0.796 0.767 0.949 0.847 0.821 0.778 0.83 1.023 1.021 1.116 0.989 1.224 0.823 0.838 1.058 0.972 0.856 0.76 1.043 0.877 0.956 0.754 0.768 0.775 0.831 1.074 0.925 0.925 0.958 1.111 1.044 0.887 0.993 0.951 0.856 32.95 14.59 5.217 4.94 6.025 5.43 4.55 3.721 4.195 3.442 3.692 2.855 2.851 2.855 2.921 3.467 2.817 2.774 2.691 2.917 2.711 2.218 2.449 2.145 1.78 0.00936 0.078 0.00518 0.028 0.179 0.0437 0.0231 0.207 0.00204 0.00351 0.251 0.242 0.13 0.0662 0.054 0.021 0.179 0.0256 0.227 0.15 0.0211 0.0112 0.14 0.0129 0.248 Abbreviations: gb, GenBank; DKO, double knockout; PURO, (supplemented with) puromycin. * Fold induction on response to H-ras expression in p65⫹/⫹ MEFs. † Normalization procedure is described in Materials and Methods. ‡ t test P values for significance across two independent experiments involving independent Affymetrix chips. § Gene name based on ⬎80% nucleotide identity to Hs homolog. 7252 Downloaded from cancerres.aacrjournals.org on June 11, 2017. © 2004 American Association for Cancer Research. NF-B SUBUNITS POTENTIATE RAS TRANSFORMATION Fig. 5. Ras-induced expression BSF3 is dependent on p65 and c-Rel. A, quantitative real-time PCR for expression of BSF3, Serpinb2 (A), c-IAP1 and TRAF2 (B) from ⫹/⫹ and p65/c-Rel DKO MEFs infected with either control or HRasV12-expressing virus. Data shown are representative of at least three independent experiments, each performed in triplicate. [Data represented as molecules (mol.) of a transcript versus attomole of 18S.] tified four distinct populations of Ras-inducible genes: (a) p65/c-Rel independent (unaffected basally in DKO cells); (b) p65/c-Rel basally activated (basal levels repressed in DKO cells); (c) p65/c-Rel basally repressed (i.e., basal levels up-regulated in DKO cells); and (d) p65/c-Rel-dependent (unaffected basally in DKO cells). A list of those Ras-induced genes clearly dependent on p65 and/or c-Rel and their relative expression is presented in Table 1. Among the NF-B-dependent genes are: (a) plasminogen activator inhibitor type 2 (Serpin B2) known to be regulated by NF-B and by oncogenic Ras and to suppress TNF-induced apoptosis (26 –28); (b) TSLP (thymic stromal lymphopoietin) which can promote the growth of pre-B cells (29, 30); (c) plakophilin-1 (a member of the armadillo/-catenin family, which is known to be up-regulated in certain cancers (31); (d) Tage4/nectain-like molecule 5 [associated with cell migration (32)], (e) neurotrophin/B-cell stimulating factor 3 [a member of the IL-6 family (33)]; (f) Posh-like molecule [posh is a Rac-regulated scaffold protein associated with JNK and NF-B signaling pathways (34)]; (g) galectin 7, expression of which is associated with a more aggressive phenotype in lymphoma cells (35); (h) Stk10 [a recently described polo-like kinase expressed in hematopoietic tissues (36)]; and (i) PCA-1, a cell surface marker for multiple myeloma (37). In agreement with a recent study (29), we found several genes, including phosholipase A2 group VII, connexin 43, transferrin receptor, ERK3, and Mdm2 to be induced by H-RasV12 in ⫹/⫹ MEFs. Expression of Tage4, shown here to be NF-B-dependent, was also found increased in K-RasV12 transformed NIH3T3 cells by Ikeda et al. (32). Additionally, other Ras-induced genes were identified for which NF-B played a measurable, but less substantial, role in regulating gene expression. These genes and those induced by Ras in a manner independent of NF-B will be reported/deposited elsewhere. To confirm results derived from the microarray studies, we further analyzed the expression of several of the inducible genes with quantitative PCR. Quantitative real-time PCR was performed on cDNA generated from ⫹/⫹ and p65/cRel DKO infected with control or H-RasV12 virus with primers specific to Serpinb2, Bsf3, c-IAP2, and TRAF2. As shown in Fig. 5, SerpinB2 and Bsf3 (both identified as NF-B regulated in the array studies) were found to be induced by Ras in a manner highly dependent on the presence of p65/RelA and/or c-Rel (Fig. 5A). This analysis demonstrated that c-IAP2 and TRAF2, although known to be regulated by NF-B downstream of TNFinduced signaling (25), were not induced by oncogenic H-Ras (Fig. 5B). In fact, Ras repressed c-IAP2 expression. These results, therefore, have identified a gene set induced by oncogenic Ras in a manner highly dependent on the p65 and/or c-Rel subunits of NF-B. These NF-B-dependent genes are likely to encode proteins that promote the efficacy of transformation of cells in response to oncogenic Ras expression. Longer-term experimentation would be required to determine how many of these genes are direct targets for NF-B and whether knockdown of expression of any of these genes would exhibit a measurable effect on Ras-induced transformation efficiency. DISCUSSION Ras-induced cellular transformation is mediated by interactions between Ras and downstream effector proteins such as PI3K, Raf, and RalGEFs (1). Raf is considered to be the most potent of this group in controlling transformation in murine cells, whereas RalGDS appears to play a more important role in Ras-induced transformation of human cells (1, 38). In support of the key role of Raf in transformation of murine fibroblasts, it has been observed that constitutively active Raf or mitogen-activated protein/extracellular signal-regulated kinase [MAP/ERK kinase (MEK)] can cause tumorigenic conversion of immortalized 3T3 cells and that inhibitors of this pathway can block this process (1, 38). Activation of the ERK pathway presumably targets certain transcription factors that promote the changes in gene expression that stimulate oncogenic transformation. Our group and others have shown that NF-B is activated downstream of Rasinduced as well as Raf-induced signaling and that inhibition of NF-B suppresses the ability of Ras to induce oncogenic conversion (3– 6). In this regard, recent reviews have covered the potential involvement of NF-B in promoting oncogenic conversion and progression (39, 40). Importantly, our data with murine fibroblasts for analysis of Rasinduced responses indicate that the activation of NF-B in this process does not involve the traditional IB degradation pathway that is associated with cytokine-induced signaling (19). However, it is noted that oncogenic Ras induces IKK activity and IB degradation in human epithelial cells (3). The results in this article indicate that the two major transactivating 7253 Downloaded from cancerres.aacrjournals.org on June 11, 2017. © 2004 American Association for Cancer Research. NF-B SUBUNITS POTENTIATE RAS TRANSFORMATION NF-B subunits, p65 and c-Rel, are not essential for Ras to induce cellular transformation of murine fibroblasts. However, re-expression of these subunits in p65- or c-Rel-null cells strongly promotes the ability of Ras to induce focus formation (Figs. 1 and 2). These findings help to clarify previous results with expression of IB-SR in which inhibition suppressed, but was unable to completely block, Ras-induced cellular transformation. Furthermore, we identify a gene set that is activated by oncogenic Ras (V12) in murine fibroblasts, the expression of which is highly dependent on either p65 or c-Rel (Table 1). The proposed functions of these gene products are generally consistent with promotion of cell growth and oncogenic transformation. Of the genes identified as NF-B regulated, Bsf3, TSLP, and Stk10, all have documented effects on cellular proliferation. For example, BSF3 and TSLP have both been shown to stimulate the growth of myeloid leukemia cells (29, 33). Although the precise function of serine/threonine kinase Stk10 is unknown, the only known substrate of Stk10, Plk1, is a protein that is involved in cellular proliferation (36). A number of the other genes identified as Ras inducible and NF-B dependent have also been found to exhibit increased expression in cancer. Tage4, a nectin-like protein, was originally found up-regulated in rat colon carcinomas and Min mouse intestinal adenomas (41). More recently, it has been determined that Tage4 interacts with nectin-3 and enhances cell migration (32). Additionally, Plakophilin-1, a component of desmosomes, has been found overexpressed in head and neck squamous cell carcinomas (42, 43). At this point, it is unclear which of the NF-B-regulated genes contribute to the enhancement of Ras-induced cellular transformation. The function of NF-B in potentiating Ras-induced cell transformation could be manifested at several levels, some of which may be independent of Ras-signaling. The simplest hypothesis is that the induction of certain genes by Ras in a manner dependent on p65 and c-Rel promotes the efficacy or likelihood of cellular transformation. This hypothesis is consistent with the microarray data presented in Table 1 and Fig. 5 and with the proposed functions of several of the genes identified as Ras inducible and NF-B dependent. However, based on the known antiapoptotic function of NF-B, we cannot rule out the possibility that the loss of NF-B and certain NF-B–regulated (but potentially not Ras-inducible) genes provides a proapoptotic background for Ras. Thus, oncogenic Ras expression may induce a higher level of apoptosis in p65- and c-Rel-null cells leading to less efficient oncogenic conversion. Additionally, Ras may induce certain antiapoptotic genes in an NF-B-dependent manner. Although the NF-B–regulated antiapoptotic gene c-IAP2 was not found to be inducible by Ras in murine fibroblasts, Serpinb2 has been shown to protect cells from TNF-induced apoptosis and may serve an antiapoptotic function in Ras transformation as well (26). A role for NF-B in suppressing Ras-induced apoptosis has been previously described by our group (5). Another potential role for NF-B in promoting Rasinduced oncogenic transformation is the control of Ras-induced signaling. Thus, NF-B may be required for more potent or persistent activation of Raf-induced signaling, e.g., promoting higher levels of Ras-induced cellular transformation. Additionally, we cannot rule out the possibility that Ras represses a set of genes in an NF-B– dependent manner and that this process also promotes transformation. Combinations of any of these possibilities remain possible and additional studies in these areas are currently underway. Additionally, it must be noted that the genetic phenotype of an immortalized cell may control or modulate the ability of NF-B to regulate specific gene expression and, correspondingly, to contribute to oncogenic transformation. In this regard, we have analyzed the status of the tumor suppressors Arf and p53 in the p65/c-Rel-null MEFs used in our studies. Both proteins appear to be wild-type, because they are inducible by Ras and UV exposure, respectively (data not shown). On the basis of earlier studies, induction of Arf by Ras in these cells would be expected to block NF-B function (44). However, clearly, Ras can use NF-B in a functional manner in the null MEFs restored for NF-B expression, indicating that Arf does not suppress NF-B activity directed toward certain genes. Regarding the genes induced by Ras in a manner dependent on NF-B, it is noteworthy that the majority of these genes have not been identified as NF-B regulated downstream of cytokine-induced signaling. We did not find any of the well-characterized NF-B–regulated cytokine, cytokine receptors, cell adhesion genes, and so forth, associated with immune and inflammatory function as Ras-inducible genes. These results are consistent with our previous hypothesis that Ras induces a selective arm of the NF-B pathway while suppressing the ability of TNF to activate NF-B and NF-B– dependent gene expression in murine fibroblasts (19). Thus, a remaining challenge is to understand how Ras-induced signals converge on NF-B to activate certain genes while simultaneously suppressing the ability of cytokines to activate the more traditional, and better characterized, NF-B-response. It is likely that this key process controlled by oncogenic Ras, selectively activating NF-B in a manner leading to up-regulation of a distinct subset of genes, is an important component in the ability of Ras and potentially other oncoproteins to efficiently transform cells. ACKNOWLEDGMENTS We thank Dr. Alexander Hoffmann (University of California San Diego, La Jolla, CA) for the generous gift of NF-B-null MEFs. REFERENCES 1. Reuther GW, Der CJ. The Ras branch of small GTPases: Ras family members don’t fall far from the tree. Curr Opin Cell Biol 2000;12:57–165. 2. Bos JL. ras oncogenes in human cancer: a review. Cancer Res 1989;49:4682–9. 3. Arsura M, Mercurio F, Oliver AL, Thorgeirsson SS, Sonenshein GE. Role of the IkappaB kinase complex in oncogenic Ras- and Raf-mediated transformation of rat liver epithelial cells. Mol Cell Biol 2000;20:5381–91. 4. Finco TS, Westwick JK, Norris JL, Beg AA, Der CJ, Baldwin AS Jr. Oncogenic Ha-Ras-induced signaling activates NF-kappaB transcriptional activity, which is required for cellular transformation. J Biol Chem 1997;272:24113– 6. 5. Mayo MW, Wang CY, Cogswell PC, et al. Requirement of NF-kappaB activation to suppress p53-independent apoptosis induced by oncogenic Ras. Science (Wash DC) 1997;278:1812–5. 6. Norris JL, Baldwin AS Jr. Oncogenic Ras enhances NF-kappaB transcriptional activity through Raf-dependent and Raf-independent mitogen-activated protein kinase signaling pathways. J Biol Chem 1999;274:13841– 6. 7. Li X, Stark GR. NFkappaB-dependent signaling pathways. Exp Hematol 2002;30: 285–96. 8. Silverman N, Maniatis T. NF-kappaB signaling pathways in mammalian and insect innate immunity. Genes Dev 2001;15:2321– 42. 9. Ghosh S, Karin M. Missing pieces in the NF-kappaB puzzle. Cell 2002;109(Suppl): S81–96. 10. Zhong H, SuYang H, Erdjument-Bromage H, Tempst P, Ghosh S. The transcriptional activity of NF-kappaB is regulated by the IkappaB-associated PKAc subunit through a cyclic AMP-independent mechanism. Cell 1997;89:413–24. 11. Zhong H, Voll RE, Ghosh S. Phosphorylation of NF-kappa B p65 by PKA stimulates transcriptional activity by promoting a novel bivalent interaction with the coactivator CBP/p300. Mol Cell 1998;1:661–71. 12. Wang D, Baldwin AS Jr. Activation of nuclear factor-kappaB-dependent transcription by tumor necrosis factor-alpha is mediated through phosphorylation of RelA/p65 on serine 529. J Biol Chem 1998;273:29411– 6. 13. Wang D, Westerheide SD, Hanson JL, Baldwin AS Jr. Tumor necrosis factor alpha-induced phosphorylation of RelA/p65 on Ser529 is controlled by casein kinase II. J Biol Chem 2000;275:32592–7. 14. Sakurai H, Chiba H, Miyoshi H, Sugita T, Toriumi W. IkappaB kinases phosphorylate NF-kappaB p65 subunit on serine 536 in the transactivation domain. J Biol Chem 1999;274:30353– 6. 15. Madrid LV, Mayo MW, Reuther JY, Baldwin AS Jr. Akt stimulates the transactivation potential of the RelA/p65 subunit of NF-kappa B through utilization of the Ikappa B kinase and activation of the mitogen-activated protein kinase p38. J Biol Chem 2001;276:18934 – 40. 16. Sizemore N, Lerner N, Dombrowski N, Sakurai H, Stark GR. Distinct roles of the Ikappa B kinase alpha and beta subunits in liberating nuclear factor kappa B 7254 Downloaded from cancerres.aacrjournals.org on June 11, 2017. © 2004 American Association for Cancer Research. NF-B SUBUNITS POTENTIATE RAS TRANSFORMATION 17. 18. 19. 20. 21. 22. 23. 24. 25. 26. 27. 28. 29. (NF-kappa B) from Ikappa B and in phosphorylating the p65 subunit of NF-kappa B. J Biol Chem 2002;277:3863–9. Langer SJ, Bortner DM, Roussel MF, Sherr CJ, Ostrowski MC. Mitogenic signaling by colony-stimulating factor 1 and ras is suppressed by the ets-2 DNA-binding domain and restored by myc overexpression. Mol Cell Biol 1992;12:5355– 62. Yang BS, Hauser CA, Henkel G, et al. Ras-mediated phosphorylation of a conserved threonine residue enhances the transactivation activities of c-Ets1 and c-Ets2. Mol Cell Biol 1996;16:538 – 47. Hanson JL, Anest V, Reuther-Madrid J, Baldwin AS. Oncoprotein suppression of tumor necrosis factor-induced NF kappa B activation is independent of Raf-controlled pathways. J Biol Chem 2003;278:34910 –7. Gilmore TD, Cormier C, Jean-Jacques J, Gapuzan ME. Malignant transformation of primary chicken spleen cells by human transcription factor c-Rel. Oncogene 2001; 20:7098 –103. Romieu-Mourez R, Kim DW, Shin SM, et al. Mouse mammary tumor virus c-rel transgenic mice develop mammary tumors. Mol Cell Biol 2003;23:5738 –54. Viatour P, Legrand-Poels S, van Lint C, et al. Cytoplasmic IkappaBalpha increases NF-kappaB-independent transcription through binding to histone deacetylase (HDAC) 1 and HDAC3. J Biol Chem 2003;278:46541– 8. Zhou M, Gu L, Zhu N, Woods WG, Findley HW. Transfection of a dominantnegative mutant NF-kappaB inhibitor (IkappaBm) represses p53-dependent apoptosis in acute lymphoblastic leukemia cells: interaction of IkBm and p53. Oncogene 2003;22:8137– 44. Gapuzan ME, Yufit PV, Gilmore TD. Immortalized embryonic mouse fibroblasts lacking the RelA subunit of transcription factor NF-kappaB have a malignantly transformed phenotype. Oncogene 2002;21:2484 –92. Baldwin AS. Control of oncogenesis and cancer therapy resistance by the transcription factor NF-kappaB. J Clin Investig 2001;107:241– 6. Dickinson JL, Bates EJ, Ferrante A, Antalis TM. Plasminogen activator inhibitor type 2 inhibits tumor necrosis factor alpha-induced apoptosis. Evidence for an alternate biological function. J Biol Chem 1995;270:27894 –904. Mahony D, Kalionis B, Antalis TM. Plasminogen activator inhibitor type-2 (PAI-2) gene transcription requires a novel NF-kappaB-like transcriptional regulatory motif. Eur J Biochem 1999;263:765–72. Cohen RL, Niclas J, Lee WM, et al. Effects of cellular transformation on expression of plasminogen activator inhibitors 1 and 2. Evidence for independent regulation. J Biol Chem 1989;264:8375– 83. Sims JE, Williams DE, Morrissey PJ, et al. Molecular cloning and biological characterization of a novel murine lymphoid growth factor. J Exp Med 2000;192:671– 80. 30. Quentmeier H, Drexler HG, Fleckenstein D, et al. Cloning of human thymic stromal lymphopoietin (TSLP) and signaling mechanisms leading to proliferation. Leukemia (Baltimore) 2001;15:1286 –92. 31. Papagerakis S, Shabana AH, Depondt J, Gehanno P, Forest N. Immunohistochemical localization of plakophilins (PKP1, PKP2, PKP3, and p0071) in primary oropharyngeal tumors: correlation with clinical parameters. Hum Pathol 2003;34:565–72. 32. Ikeda W, Kakunaga S, Itoh S, et al. Tage4/Nectin-like molecule-5 heterophilically trans-interacts with cell adhesion molecule Nectin-3 and enhances cell migration. J Biol Chem 2003;278:28167–72. 33. Senaldi G, Varnum BC, Sarmiento U, et al. Novel neurotrophin-1/B cell-stimulating factor-3: a cytokine of the IL-6 family. Proc Natl Acad Sci USA 1999;96:11458 – 63. 34. Tapon N, Nagata K, Lamarche N, Hall A. A new rac target POSH is an SH3containing scaffold protein involved in the JNK and NF-kappaB signalling pathways. EMBO J 1998;17:1395– 404. 35. Moisan S, Demers M, Mercier J, Magnaldo T, Potworowski EF, St-Pierre Y. Upregulation of galectin-7 in murine lymphoma cells is associated with progression toward an aggressive phenotype. Leukemia (Baltimore) 2003;17:751–9. 36. Walter SA, Cutler RE Jr, Martinez R, Gishizky M, Hill RJ. Stk10, a new member of the polo-like kinase kinase family highly expressed in hematopoietic tissue. J Biol Chem 2003;278:18221– 8. 37. Ruiz-Arguelles G, San Miguel J. Cell surface markers in multiple myeloma. Mayo Clin Proc 1994;69:684 –90. 38. Hamad NM, Elconin JH, Karnoub AE, et al. Distinct requirements for Ras oncogenesis in human versus mouse cells. Genes Dev 2002;16:2045–57. 39. Gilmore TD. The Rel/NF-kappaB/IkappaB signal transduction pathway and cancer. Cancer Treat Res 2003;115:241– 65. 40. Lin A, Karin M. NF-kappaB in cancer: a marked target. Semin Cancer Biol 2003; 13:107–14. 41. Chadeneau C, LeCabellec M, LeMoullac B, Meflah K, Denis MG. Over-expression of a novel member of the immunoglobulin superfamily in Min mouse intestinal adenomas. Int J Cancer 1996;68:817–21. 42. Takahashi T, Sano B, Nagata T, et al. Polo-like kinase 1 (PLK1) is overexpressed in primary colorectal cancers. Cancer Sci 2003;94:148 –52. 43. Elez R, Piiper A, Giannini CD, Brendel M, Zeuzem S. Polo-like kinase1, a new target for antisense tumor therapy. Biochem Biophys Res Commun 2000;269:352– 6. 44. Rocha S, Campbell K, Perkins N. p53 and Mdm2-independent repression of NFkappaB transactivation by the ARF tumor suppressor. Mol Cell 2003;12:15–25. 7255 Downloaded from cancerres.aacrjournals.org on June 11, 2017. © 2004 American Association for Cancer Research. The Nuclear Factor κB Subunits RelA/p65 and c-Rel Potentiate but Are Not Required for Ras-Induced Cellular Transformation Julie L. Hanson, Noel A. Hawke, David Kashatus, et al. Cancer Res 2004;64:7248-7255. Updated version Cited articles Citing articles E-mail alerts Reprints and Subscriptions Permissions Access the most recent version of this article at: http://cancerres.aacrjournals.org/content/64/20/7248 This article cites 43 articles, 24 of which you can access for free at: http://cancerres.aacrjournals.org/content/64/20/7248.full.html#ref-list-1 This article has been cited by 14 HighWire-hosted articles. Access the articles at: /content/64/20/7248.full.html#related-urls Sign up to receive free email-alerts related to this article or journal. To order reprints of this article or to subscribe to the journal, contact the AACR Publications Department at [email protected]. To request permission to re-use all or part of this article, contact the AACR Publications Department at [email protected]. Downloaded from cancerres.aacrjournals.org on June 11, 2017. © 2004 American Association for Cancer Research.