Survey
* Your assessment is very important for improving the workof artificial intelligence, which forms the content of this project
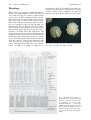
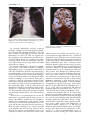
Prokaryotes (2006) 3:919–933 DOI: 10.1007/0-387-30743-5_34 CHAPTER 1.1.19 ehT suneG The Genus Mycobacterium—Medical BEATRICE SAVIOLA AND WILLIAM BISHAI Introduction The genus Mycobacterium encompasses a number of medically important species that exact an alarming toll in human morbidity and mortality. Indeed the World Health Organization (WHO) estimates that nearly one third of the world’s population (1.8 billion people) are infected with Mycobacterium tuberculosis, the cause of tuberculosis (TB) (http://www.who.org). In 1993, the WHO declared a global emergency owing to the fact that TB was epidemic in many areas of the world. Other mycobacterial diseases continue to plague the world’s populations as well. Mycobacterium leprae, the cause of leprosy, persists in developing countries, and other mycobacteria (ordinarily nonpathogens) have now become threats to individuals infected with the human immunodeficiency virus (HIV). Mycobacteria have shaped the course of human history. Indeed until 1900, TB was one of the chief causes of death in Europe and the Americas. The nature of the disease, however, remained poorly understood and as a testimonial to this fact its contagiousness was the subject of heated debate well into the 18th and 19th centuries. For many years, psychological and inherited factors were thought to predispose individuals to the disease. However, others thought that a contagious agent was the culprit. This debate ended only after the bacillus Mycobacterium tuberculosis was identified as the causative agent of TB. Jean Antoine Villemin in 1868 was the first to transmit TB from man to rabbit (Haas and Haas, 1996). As a military doctor, he had observed that many young healthy military personnel housed in barracks eventually succumbed to TB. He also noted that a disproportionate number of individuals with active TB were prisoners, industrial workers, members of cloistered religious orders as well as military personnel; all of them were housed together with many other people. Villemin speculated that TB was transmissible. He used material (gray and soft tubercles) from humans who had succumbed to TB as well as blood and sputum from tuberculous patients to infect rabbits. The inoculated rabbits indeed developed pathologic evidence of tuberculosis. Then in 1882, Robert Koch was the first to view Mycobacterium tuberculosis through a light microscope. He was then able to grow M. tuberculosis in pure culture, infect guinea pigs with the bacilli, and reisolate bacteria from these animals (Adler and Rose, 1996). Thus a new era of mycobacteria research was born. In Europe and the United States, a sanatorium movement was underway by the 1890s to isolate and cure those patients having TB. Edward Livingston Trudeau started one of the first and most successful of the sanatoria in the United States (Davis, 1996). After providing hospice care for his consumptive brother, Dr. Trudeau was stricken with that same disease. He fled to the Adirondack Mountains believing that fresh air, nutritious food and exercise would restore his health. As hoped, his disease symptoms abated and his health consequently improved. He chose to move permanently to Saranac Lake in the Adirondack Mountains where he bought land, built cottages and started the Adirondack Cottage Sanatorium. Dr. Trudeau immersed himself in research and succeeded in cultivating pure Mycobacterium tuberculosis, which he provided to other researchers. Out of the Saranac Lake Laboratory, the Trudeau Institute (http://www.trudeauinstitute. org) for biomedical research was born in 1954. Not until the development of successful antimycobacterial chemotherapy did this and other sanitoria lose their function and close their doors to the public. In 1944 streptomycin was isolated and identified by Dr. Selman Waksman and his graduate student Albert Schatz (Harris, 1996). It marked the first drug in a line of antibiotics having antimycobacterial properties. In 1952 isonicotinic acid hydrazide (INH) was found to have antimycobacterial properties as well and has since become a mainstay in the antibiotic therapy of TB. With the advent of these new advances in the chemotherapy of TB, victory was declared over the disease; however, this declaration would prove premature. Following an extensive period of decline, the incidence of TB in the United States began to rise again in the late 1980s. The l ac i dM e- -mu i re t cabocM y 920 B. Saviola and W. Bishai HIV epidemic combined with an urban drug problem provided fertile ground on which M. tuberculosis could flourish anew. Concomitant with this alarming trend was detection of an increasing incidence of drug-resistant M. tuberculosis. To combat these problems, the WHO as well as government agencies in countries around the world have increased spending in both the public health forum as well as biomedical research. Today, research on M. tuberculosis as well as other members of the genus is providing valuable insight into the basic biology of mycobacterial survival and replication inside the host. Lessons learned through these studies will hopefully provide novel treatments to combat the renewed threat of this age-old enemy. Taxonomy The genus Mycobacterium comprises a number of Gram-positive aerobic bacteria and is the only member of the family Mycobacteriaceae within the order Actinomycetales. The genus shares an unusually high genomic DNA G+C content (62– 70%) and the production of mycolic acids with closely related genera, Nocardia and Corynebacterium, within Actinomycetales. Phylogenetic trees are available which depict genetic relatedness based on homology of the 16S ribosomal gene sequence. Mycobacteria that have highly homologous rRNA sequences are closely related and are on neighboring branches of the tree (Fig. 1). The mycobacterial phylogenetic tree can be further subdivided into fast and slow growing bacteria. The fast growers form colonies on selective media in less than 7 days and the slow growers, in greater than 7 days. In addition, within the genus Mycobacterium a number of species are grouped into complexes (e.g., M. avium and M. tuberculosis complexes) that include bacterial species that have a high degree of genetic similarity as well as cause similar disease syndromes. Habitat Mycobacterium tuberculosis is an obligate pathogen of humans and is rarely identified in other mammals. It is transmitted from person to person, and it has no significant environmental reservoirs. Mycobacterium bovis, which causes tuberculosis in humans and in cattle, has a natural reservoir in ruminants. As a consequence, foodstuffs (including cheese and milk originating from these animals) were often contaminated prior to the introduction of current pasteurization and meat inspection procedures. A number of other medically important mycobacteria are found in the environment. Water serves as the CHAPTER 1.1.19 habitat for a number of mycobacterial species including M. marinum, M. cheloneae, M. fortuitum, M. kansasii and M. avium. Although not yet confirmed, stagnant water may be a habitat for M. ulcerans. Soil may also harbor mycobacteria including M. cheloneae, M. fortuitum, and M. avium. Isolation Medically important mycobacterial species can be isolated and cultured from a number of environmental sources; however, M. tuberculosis is rarely isolated from nonhuman sources. As pulmonary tuberculosis is the most frequent form of the disease, sputum is the most common body specimen submitted for culture—although blood, gastric aspirates and biopsy specimens may also be analyzed. Before sputum samples from patients may be tested for mycobacterial growth, they must be decontaminated to remove rapidly growing upper respiratory flora. Decontamination procedures take advantage of the thick lipid envelope of mycobacteria and this organism’s natural resistance to chemical agents. N-Acetyl-Lcysteine (0.5–2.0%; NALC) liquefies sputum samples. The compound has minor inhibitory effects on mycobacteria but does allow other antibacterial agents to access and neutralize contaminating organisms. Sodium hydroxide is used to inactivate contaminating bacteria with only modest inhibitory effects against mycobacteria. Once decontaminated, samples may be used to inoculate either a solid or liquid growth medium. The solid growth media are available as either egg- or agar-based. Both types of solid media contain malachite green dye, which inhibits the growth of many other bacteria. The commonly used Löwenstein-Jensen (LJ) medium is egg-based (Fig. 2), whereas Middlebrook 7H10 and 7H11 are agar-based media. Growth can also be assayed in a liquid medium such as Middlebrook 7H9; however, these media are more problematic because overgrowth by contaminating bacteria is more difficult to detect. Growth of mycobacteria from clinical specimens is time-consuming because there is often a lag of three to four weeks before sufficient growth is achieved. To shorten growth detection times, the Bactec (Becton-Dickinson) system may be used (Fig. 2). Mycobacteria are grown in 7H9 that contains 14C-labeled palmitic acid as a carbon source. As the bacteria grow they catabolize the radiolabeled carbon source and convert it to gaseous 14CO2. A specially designed instrument (Becton Dickinson) samples the amount of 14 C above the mycobacterial cultures and converts this information into an index of growth. CHAPTER 1.1.19 The Genus Mycobacterium—Medical 921 M. fortuitum M. farcinogenes M. senegalense M. chelorne M. penegrinum M. neaurum M. diernhoferi M. abscessus M. chitae M. fullax M. aurum M. vaccae M. confluentis M. madagascariense M. flavescens M. smegmatis M. thermoresistible M. phlei rapid growers M. triviaic M. simae M. genavense slow growers M. interjectum M. intermedium M. terrar M. hibernine M. nonchromogenicum M. cookii M. xenapi M. celatum type 1 M. celatum type 2 M. gordonae M. asiaticam M. luberculosis complex M. marinum M. leprne M. scrofmlaceni M. gratrif, M. kansaii M. szulgni M. malmacuse M. intracellulars M. paratuberculosis M. avium Fig. 1. A phylogenetic tree based on 16S rRNA sequences of the genus Mycobacterium depicts closely related species on neighboring branches. Redrawn from Shinnick and Good (1994). The Bactec method reduces detection time in liquid media to 9–14 days for M. tuberculosis (Middleton et al., 1997; Pfyffer et al., 1997). Recently a completely closed system called the mycobacterial growth indicator tube (MGIT) system has been introduced. The tube contains a plastic resin that fluoresces as oxygen levels are depleted by growing bacteria, and an automated fluorescence detector reports growth readings at regular intervals (Saito et al., 1996; Sharp et al., 1996). Identification Once mycobacteria have been isolated from either an environmental or a human source, the speciation process can begin. Acid-fast staining 922 B. Saviola and W. Bishai CHAPTER 1.1.19 Fig. 3. Kinyoun acid-fast stain of Mycobacterium smegmatis. Bacilli appear as small red rods with clumping. Fig. 2. The growth medium Löwenstein Jensen (left) is used to culture mycobacteria. Mycobacterium tuberculosis takes 3 to 4 weeks to produce colonies on this growth medium. The Bactec 12B (right) bottle is used for radiometric detection of mycobacterial growth. Early growth of mycobacteria is detected by the conversion of 14C-labeled palmitate to gaseous 14C-labeled CO2. An automated sampling needle removes a small amount of gas through a rubber septum each day, and a detector measures the radioactivity obtained. Using the Bactec system, mycobacterial growth may be detected in 7–15 days. in combination with light microscopy permits an initial rapid diagnosis of a mycobacteriosis. Mycobacteria are acid fast by nature. They form stable complexes with arylmethane dyes such as fuchsin and auramine O. The phenol used in the primary staining procedure allows the stain to penetrate. The mycolic acids in the cell wall act to retain the stain even after the exposure to acid alcohol or strong mineral acids. The resulting acid-fast mycobacteria can be identified microscopically. The Ziehl-Neelsen and Kinyoun methods are acid stains in which the acid-fast bacilli (AFB) appear red against a blue or green background (Fig. 3). Fluorochrome-staining procedures can be used as well. In these procedures, which use auramine O or auramine-rhodamine, the AFB fluoresce yellow to orange. Fluorescence staining is superior as AFB are more easily identified at a lower magnification. Initial microscopic identification of bacteria, while rapid, is not reliable, has a sensitivity that ranges from 22 to 80%, and does not allow species identification of AFB. Therefore, other methods of mycobacterial identification are necessary (Nolte and Metchock, 1995). A host of biochemical tests can be used to differentiate mycobacterial species. Biochemical tests that distinguish M. tuberculosis from other species include the niacin accumulation test, the 68°C catalase test and the nitrate reduction test (Kent and Kubica, 1985; Nolte and Metchock, 1995). Because M. tuberculosis has a blocked pathway for the conversion of free niacin to nicotinic acid mononucleotide, the bacterium accumulates niacin and excretes it into the culture medium. As confirmation, additional biochemical tests (e.g., the nitrate reduction test) must be performed inasmuch as strains of some mycobacterial species other than M. tuberculosis also accumulate niacin. Mycobacterium tuberculosis is a strongly positive nitrate reducer. Because M. kansasii, M. szulgai and M. fortuitum also reduce nitrate, an additional test (e.g., heat-stable catalase) should be used. The heat-stable catalase test assays the amount of catalase present in the bacterium after it has been resuspended in a buffer and heated to 68°C. Mycobacterium tuberculosis always loses its catalase activity under these conditions. Once AFB have been observed in culture, most clinical labs use a nucleic acid hybridization test rather than the more laborious biochemical tests to speciate the isolate. The Accuprobe assay is available through Gen-Probe (http:// www.gen-probe.com). This assay uses the Hybridization Protection Assay (HPA) where a DNA acridinium-ester-labeled probe hybridizes to a mycobacterial rRNA target sequence. A selection agent cleaves the label away from the unhybridized probe but leaves untouched the probe that is complexed with its target sequence. Light (detected with a luminometer) will be emitted from the DNA probe that is complexed to the rRNA. The advantage of this technique is that it can identify small quantities of mycobacteria due to the many copies of the rRNA target sequence in each mycobacterium. Thus there is an inherent amplification of the signal sequence. Gen-Probe has available probes that are specific for M. avium, M. intracellulare, the M. avium CHAPTER 1.1.19 complex, M. gordonae, M. kansasii and the M. tuberculosis complex. Nucleic Acid Amplification Tests to Detect and Speciate M. tuberculosis Directly from Sputum Even shorter detection times may be achieved using various commercially available nucleic acid amplification assays (NAA) designed to work using sputum. The United States Food and Drug Administration (USFDA) has approved several commercially available amplification tests for use directly with sputum. These tests include the Mycobacterium tuberculosis Direct Test from Gene-Probe (MTD; http://www.gen-probe.com) and the Amplicor (http://www.roche.com/ diagnostics/) Mycobacterium tuberculosis (MTB) Test. The MTD combines Transcription Mediated Amplification, which can amplify target rRNA sequences, with the HPA described in the previous section. This test is approved by the USFDA for use with sputum samples that are both positive and negative for AFB. The MTB test uses the polymerase chain reaction (PCR) to amplify DNA specific to M. tuberculosis. This test is approved by the USFDA for use with sputum samples positive for AFB. The LCx M. tuberculosis Test (http://www.abbott.com/research/ diagnostics.htm) (Abbott) utilizes a ligase chain reaction where two oligos are hybridized to a target sequence from the mycobacteria and are subsequently ligated. The ligated oligos then serve as templates for additional oligos to hybridize and be ligated, resulting in amplification of the target sequence. At present this test is not approved for use with sputum within the United States. A recent study comparing two NAA methods revealed that MTD and LCx had sensitivities of 98.6 and 100%, respectively, and specificities of 99.4 and 99.3% with smear-positive samples (Wang and Tay, 1999). The Amplicor MTB test was analyzed in clinical trials and had a sensitivity of 95.0% and a specificity of 100% with smear-positive sputum samples (Amplicor, 2000). The NAA tests, while costly, are slightly more sensitive than acid-fast staining procedures. These tests hasten care for infected individuals by quickly identifying mycobacteria within about 25–50% of sputum samples that are negative by acid-fast staining but later culture positive. This aspect of the NAA tests is important especially in view of the recent evidence supporting the transmission of M. tuberculosis from patients who did not have acid-fast bacilli in their sputum (Behr et al., 1999). Additionally, in contrast to acid-fast staining, NAA tests provide information about species present within a The Genus Mycobacterium—Medical 923 sputum sample. Thus, M. tuberculosis-infected individuals can be isolated and treated more rapidly, preventing the spread of the bacteria. The NAA tests cannot differentiate live from dead bacteria and therefore may not prove helpful for monitoring response to therapy. Once a bacterial isolate has been identified as M. tuberculosis, it can be further subtyped to the strain level. The ability differentiate M. tuberculosis on the strain level has enhanced our knowledge of the epidemiology of TB. In fact it has led to the identification of large outbreaks of disease traced back to a single person (Edlin et al., 1992; Valway et al., 1998). Strain typing also has lead to the discovery of unsuspected routes of transmission such as medical equipment (Agerton et al., 1997; Michele et al., 1997), embalming (Sterling et al., 2000), or exposure to childhood TB (Curtis et al., 1999). It has been used, as well, to assess the amount of recent transmission of M. tuberculosis in the community, thereby redefining our understanding of the balance between primary active and reactivation TB after a period of latency. Techniques of DNA fingerprinting allow the identification of different M. tuberculosis strains. Virtually all M. tuberculosis strains have 1–26 copies of an insertion sequence IS6110. When a strain’s chromosomal DNA is fragmented with restriction enzymes, the number of copies and location of these insertion sequences within the chromosome produce a restriction length polymorphism (RFLP) pattern that characterizes the strain. In addition, the frequency with which IS6110 moves within the chromosome is low enough to allow identification of specific strains that are the cause of an outbreak. Reasonable identification of a strain can be obtained when the bacterium contains six or more IS6110 elements. In the event that a strain contains less than six insertion elements, a second probe may be used for increased specificity. This second probe is generally for the polymorphic GC-rich repetitive sequence (PGRS). Distinct strains will have a unique pattern of probe hybridization, thus facilitating identification (Harrington and Bishai, 2000). Current research in diagnostics for TB is focusing on techniques that are fast, sensitive and specific. A major challenge is to develop inexpensive tests, which may be used in developing countries where TB is abundant. These tests could potentially identify patients with active TB early, thus permitting the isolation of such individuals and preventing the spread of the bacilli. Earlier identification of those individuals that harbor M. tuberculosis could possibly lower TB rates in many parts of the developing world. 924 B. Saviola and W. Bishai Physiology Mycobacteria are straight or slightly curved nonmotile rods (approximately 0.2–0.6 µm wide by 1.0–10 µm long; Fig. 4), aerobic, nonsporeformers that grow at times as filaments. Two remarkable bacteriologic features define mycobacteria. First, they have a cell wall that is rich in longchain fatty acid esters known as “mycolic acids” that are attached to the cell wall through arabinogalactan (Fig. 5). Mycolic acids are chemically related to wax and give the colonies their characteristic waxy appearance and the cells a tendency to clump and resist dispersion. The waxy nature of the coat renders the bacteria not readily stainable (such as by the Gram method) with aniline dyes, though the bacteria are considered to be Gram positive. Mycobacterial mycolic acids do, however, form strong complexes with some dyes that leave them resistant to decolorization with acid alcohol. Therefore the identification of AFB in a sample is suggestive of CHAPTER 1.1.19 mycobacteria (Fig. 3). In addition, this lipid coat renders the mycobacteria impervious to many aseptic solutions and antibiotics. Second, all mycobacteria grow slowly with generation times Fig. 4. Mycobacterium smegmatis colony morphology after three days of growth from a single bacterium. Branched and capped portion of LAM Free Lipids Purin Mycolic acids Arabinan portion of LAM Pentaarbinasyl Motifs Arabinan Linker Galactan Pepcidaglyean LM portion of LAM PIMs Plasma membrane Polyprenyl sugars Associated plasma membrane proteins The Mycobacterium ruberculosis cell envelope Fig. 5. Schematic representation of the plasma membrane and outer wall of Mycobacterium tuberculosis reveals an abundance of mycolic acids. Phosphatidyl-inositol mannosides (PIMs), lipoarabinomannans (LAMs) and lipomannan (LM) portion of LAM are shown as outer wall constituents. From Brennan and Nikaido (1995). CHAPTER 1.1.19 Genetics The Completed Mycobacterium tuberculosis Genome Genetic research into the virulent nature of M. tuberculosis has long been hampered by the organism’s slow growth. The generation time of M. tuberculosis is 24 hours in contrast to the 20minute generation time of the common laboratory bacterium Escherichia coli. The tendency for mycobacteria to form clumps also leads to significant technical challenges in the research laboratory. As a consequence, traditional genetic screens and selections are difficult to conduct. Therefore, reverse genetic approaches have been emphasized in the study of M. tuberculosis. This technique targets specific proteins homologous to proteins of known function and importance in other organisms for study at a molecular level within a host. As a result, the elucidation and analysis of the whole genome sequence of M. tuberculosis have become increasingly important. In 1998, the complete genome sequence of the M. tuberculosis strain H37Rv was elucidated (Cole et al., 1998). The completion of the genome sequence was a collaborative effort of the Wellcome Trust and the Institute Pasteur. A web site at the Institute Pasteur shows the completed genome. In addition, The Sanger Center maintains a web site where DNA segments and predicted proteins from the M. tuberculosis strain H37Rv may be compared to peptides and DNA from other organisms. The Institute for Genomic Research (TIGR) has completed the sequence for another M. tuberculosis strain, CDC1551 (CSU93). At the TIGR web site, one may also compare the chromosomal sequence to other known genes and predicted proteins. In addition, The National Center for Biotechnology Information offers a web site where the M. tuberculosis genome may be accessed and the chromosomal sequence may be visualized on a map. The genome sequence is 4.4 Mbp. Researchers now have at their disposal the information defining every possible drug target, antigen for incorporation into a vaccine, as well as every virulence determinant from M. tuberculosis. Annotation of the H37Rv sequence reveals 3,924 open reading frames (Cole et al., 1998). The genome has no easily identifiable pathogenicity islands as other organisms have. However, many repetitive elements were identified, including IS6110, which have been used as a tool to identify varying 925 strains during outbreaks of M. tuberculosis (Harrington and Bishai, 2000). The mycobacterium contains within its repertoire genes that could allow the bacterium to adapt to a range of environmental conditions, from growth in a rich broth to survival in host macrophages. These genes include 13 putative σ factors, 140 transcriptional regulators, 32 component regulators that presumably sense environmental signals, and 14 protein kinases or phosphatases. The genome also possesses 250 genes involved in a complex lipid metabolism, which the cell needs to synthesize material for its waxy coat (Cole et al., 1998). Several new classes of putative proteins termed “PE” and “PPE” (each with internal repetitive sequences) were also identified in the genome sequence. Indeed, ten percent of the genome is composed of these new potential coding sequences. The PE or PPE class of proteins has been proposed to be involved in modulation of the host immune response (Cole et al., 1998). Epidemiology Tuberculosis causes more deaths than any other infectious organism; 6 million new active cases and 3 million deaths annually are attributed to M. tuberculosis. Indeed, rates of TB in the world are predicted to increase 50% each decade. However, the annual incidence of TB in the United States remains low with 8.7 cases/ 100,000 people. Prior to 1985, TB rates within the United States were steadily declining at approximately 6% per year. In striking contrast to this trend, from 1986 to 1992 the incidence of TB cases rose about 3% per year (Fig. 6). A Reported TB Cases United States, 1975 - 1997 35,000 30,000 Cases that range from 2 hours for M. smegmatis to 12 days for M. leprae. The Genus Mycobacterium—Medical 25,000 20,000 75 77 79 81 83 85 87 89 91 93 95 97 Year CDC Fig. 6. After many years of decline, reported TB cases in the United States began to rise in the late 1980s. Following public health intervention, TB cases are once again dropping CDC. 926 B. Saviola and W. Bishai maximum of 26,673 cases occurred in 1992. Increased government funding for public health programs and increased use of directly observed therapy (DOT) for tuberculosis have decreased rates since 1992 (Chaisson and Bishai, 1997). As a result, in 1998 the United States Centers for Disease Control and Prevention (CDC) reported 18,361 cases. Again, in 1999 the incidence of new TB cases fell to 17,528 cases. Despite the fact that the number of cases in the United States is now dropping, there is the new problem of multidrug-resistant TB. The term multidrug-resistant tuberculosis (MDRTB) is used to describe strains that are resistant to two or more of the five first-line anti-TB drugs: isoniazid, rifampin, pyrazinamide, ethambutol and streptomycin. Indeed the incidence of drug resistance has increased in urban settings and among HIV-infected populations. In fact, during 1991, 33% of all TB strains recovered in New York City were MDRTB. The United States rate of MDRTB, however, has now fallen with improved control measures, but the global problem of drug resistance continues. In an international survey between 1994 and 1997, 12% of incident cases were resistant to at least one drug and 7.6% of TB strains were resistant to isoniazid (Pablos-Mendez et al., 1998). Risk factors for TB include intravenous drug abuse, alcoholism, chronic pulmonary disease, prolonged steroid use, diabetes, renal failure, malnutrition and organ transplant. Infection with HIV is also a risk factor for TB: HIVinfected patients have a 25–50 fold increased risk compared with HIV-negative individuals. A number of social factors also increase a person’s risk of TB including institutional living, urban dwellings, poverty and low educational levels (Chaisson and Bishai, 1997). The incidence of other mycobacterial diseases has increased in recent years. Diseases caused by the Mycobacterium avium complex (MAC) were uncommon prior to the AIDS epidemic. Infection with MAC was mostly seen in individuals that were immunocompromised or had underlying lung disease. Indeed, prior to 1981 the incidence of MAC cervical lymphadenitis and MAC pulmonary infection was approximately 300 and 3,000 cases per year, respectively, in the United States. With the onset of the AIDS epidemic, the incidence of disseminated MAC infection jumped in the mid-1990s to approximately 20,000 individuals per year (Chaisson and Bishai, 1997). The MAC rates, however, appear to have peaked in 1997 and have since fallen due to improved antiretroviral therapy for patients infected with HIV (Palella et al., 1998). CHAPTER 1.1.19 Disease The MYCOBACTERIUM TUBERCULOSIS Complex The M. tuberculosis complex is comprised of M. tuberculosis, M. bovis, M. microti and M. africanum. Only M. tuberculosis and M. bovis are a significant source of human disease (http:// www.hopkins-tb.org). Mycobacterium microti causes disease in voles and was employed as a vaccine in the earlier part of the twentieth century (Wells and Oxon, 1937; Birkhaug, 1946; Wells, 1949; Wells and Wylie, 1954). Mycobacterium africanum causes disease in humans but makes up only a minority of cases of pulmonary TB, and its incidence is isolated to Africa. M. TUBERCULOSIS The main route of infection of M. tuberculosis is by person-to-person inhalation of infectious aerosols. Bacilli are transmitted through speaking, coughing, sneezing and singing. Droplets of infectious M. tuberculosis are sized in the range of 1–10 µm. After inhalation of the droplets, the bacteria travel to the terminal bronchioles and alveoli where they are phagocytosed by alveolar macrophages. Many of these bacilli are killed in the phagosomes after these fuse with lysosomes and become acidified. Mycobacterium tuberculosis, however, is an intracellular pathogen and can efficiently inhibit phagosome-lysosome fusion (Fig. 7). As a result some of the invading M. tuberculosis bacilli survive these initial host defenses. Surviving bacilli grow within the macrophages and are released when the macrophages die. Unactivated macrophages arrive from the blood stream and ingest the newly liberated bacilli, which grow symbiotically within the unactivated macrophages for approximately 3 weeks. Eventually the bacilli lyse the macrophages and spill out into the host tissue. Fig. 7. Macrophages infected with M. bovis BCG are visualized by acid-fast staining. Mycobacteria are small red rods within the blue-stained macrophages. CHAPTER 1.1.19 The Genus Mycobacterium—Medical 927 Fig. 8. Cavitary pulmonary tuberculosis in a 36-year old man. Right upper lobe streaky infiltrates and nodules are seen; two cavities are also present near the right apex. In resistant individuals, released bacterial products stimulate strong cell-mediated immunity through Th1 signaling with INF-γ, Il-2, and Il-12. Activated macrophages that can kill M. tuberculosis, as well as T cells, are recruited to the periphery of the infectious focus. The bacteria and cellular debris are contained in a tissue structure called “a caseous granuloma.” The disease will be halted at the stage where this small granuloma is formed. In sensitive individuals, cell-mediated immunity is weak, and bacilli continue to multiply. Macrophages arrive to engulf the bacilli while T cells also accumulate. Because the macrophages are inadequately activated, the M. tuberculosis bacilli also parasitize the recruited macrophages. Cytotoxic T-cells produce toxic substances, which results in damage of host tissues. This cycle is repeated so that the granuloma enlarges. Eventually, tissue damage may result in cavity formation within the lung as well as in liquefaction of the granuloma (Figs. 8 and 9). Mycobacterium tuberculosis bacilli are particularly adept at multiplying in a liquefied cavity from which they are then expelled and spread through coughing and sneezing (Dannenberg, 1993; Dannenberg, 1994). As they have no environmental reservoirs, M. tuberculosis organisms spread from person to person. In addition, fomites are not thought to support the transmission of the disease. Some (25–50%) of the diseased individual’s close contacts will become infected with M. tuberculosis. However, only a minority (5% of those exposed) is susceptible and will develop primary active disease. In contrast, most humans seem to be naturally resistant to M. tuberculosis; these indi- Fig. 9. Lung of a rabbit infected with M. bovis. Cavities are abundant within the lung. viduals harbor the bacilli and develop only a latent infection. Approximately 95% of infected individuals control the spread of M. tuberculosis within the body and disease progression is halted at the stage where a small granuloma is formed. These individuals may have a lifelong latent infection with no symptom other than a positive reaction to the administration of purified protein derivative (PPD) of M. tuberculosis, indicating immune memory of M. tuberculosis infection. Any process that disrupts the immunocompetence of a latently infected individual, however, may cause active infection to develop. The elderly often have a recrudescence of disease after decades of a latent infection due to their waning immunity. Acquiring a viral infection such as HIV may cause an individual to become immunosuppressed and develop active from latent disease. In fact, HIV-infected individuals have a 4–8% yearly risk for reactivation TB whereas immunocompetent individuals have only a 5– 10% lifetime risk (Chaisson and Bishai, 1997). Disease development due to M. tuberculosis infection can be either pulmonary or extrapulmonary. Inasmuch as infection is mainly through the aerosol route, immunocompetent individuals manifest primarily pulmonary disease. Symptoms include cough, chest pain, sputum production, fever, night sweats and hemoptysis in advanced disease. About 15% of TB cases in immunocompetent individuals occur at extrapulmonary sites. Such cases are attributed to reactivation of a latent extrapulmonary focus of infection, because during the initial infection 928 B. Saviola and W. Bishai process transient bacteremia may occur. Occasionally hospital and laboratory workers infect soft tissues through accidental injection with M. tuberculosis-contaminated syringes. In those cases infection is extrapulmonary but may spread by the blood to the lungs and other organs if the immune system cannot check the growth of the bacilli. Immunocompromised individuals (such as the very young, old and those infected with HIV) are more likely to develop extrapulmonary tuberculosis. M. BOVIS Isolated from cattle, M. bovis causes TB in ruminants and occasionally humans. This organism prefers to grow at 37°C and growth of a colony from a single bacillus usually requires 3–4 weeks of culture. Before routine pasteurization was practiced, humans could be infected by drinking M. bovis-contaminated milk. In fact milk was an important reservoir of infectious bacilli. However, since pasteurization became commonplace in the United States and in much of the world, the incidence of M. bovis infection has decreased considerably. Mycobacterium bovis is spread among cattle by an aerosol route and from cattle to humans by either a gastrointestinal or an aerosol route. In humans, M. bovis may cause intra-abdominal TB, cervical lymphadenitis (scrofula) or pulmonary TB (Adler and Rose, 1996). Mycobacterium bovis was used to derive the well-known attenuated vaccine strain, bacille Calmette Guerin (BCG), which is still used in many parts of the world. M. AFRICANUM This slow growing mycobacterium, first described in 1969, is infrequently associated with human pulmonary disease. It causes a disease similar to that caused by M. tuberculosis and M. bovis. Most patients with this disease reside or have resided in Africa. Mycobacterium africanum probably spreads by an aerosol route (Adler and Rose, 1996). THE MYCOBACTERIUM AVIUM COMPLEX The M. avium complex is comprised of three species, M. avium, M. intracellulare and M. paratuberculosis. All three species are common in the environment, as they are isolated from soil, water, food and domestic animals. Categorized as slow growing mycobacteria, these species grow optimally at 37°C and achieve growth in approximately 4– 6 weeks (Havlir and Ellner, 2000). Colonies are nonpigmented and are both opaque and domed or flat and transparent (Toosi and Ellner, 1998). Mycobacterium avium causes a tuberculosis-like infection in chickens, pigeons and other birds, whereas M. paratuberculosis causes Johne’s disease in ruminants, which is a contagious enteritis resulting in progressive wasting and eventual death (Fraser et al., 1986). Mycobacterium CHAPTER 1.1.19 paratuberculosis has not been shown to cause human disease. In humans, M. avium and M. intracellulare can cause pulmonary disease, regional lymphadenitis or disseminated disease. Those individuals who are coinfected with HIV are prone to disseminated disease, whereas those who are not coinfected are likely to have pulmonary disease. Predisposing factors to infection with M. avium include underlying lung disease, HIV infection, chronic obstructive pulmonary disease, chronic bronchitis, healed or active TB, pulmonary mycosis and malignancy. Infection occurs by either inhalation or ingestion of the infectious bacilli. Diagnosis in those affected individuals without HIV is made by radiographic evidence of disease, positive sputum cultures and an acidfast smear. Symptoms are a productive cough, hemoptysis, fever and weight loss (Chaisson and Bishai, 1997). Disseminated M. avium is a late opportunistic infection in those individuals coinfected with HIV. In fact, disseminated MAC generally occurs only in AIDS patients with CD4 cell counts less than 100/mm3 (Chaisson and Bishai, 1997). Symptoms of infection are fever, drenching night sweats and weight loss. The hallmark of a disseminated infection is the high circulating levels of MAC bacteremia, which can reach 104 bacteria/ml. Organ involvement may be widespread and the tissue burden may reach 106 bacteria/g. The gastrointestinal tract is frequently infected, leading to symptoms including nausea, vomiting, watery diarrhea and abdominal pain. Left untreated, the disease will cause progressive clinical deterioration (Havlir and Ellner, 2000). MYCOBACTERIUM LEPRAE Leprosy is particularly dreaded because it may cause bodily deformities leading to social stigmatization. In centuries past, persons suffering from leprosy were ostracized and forced to live away from the general population, in colonies or leprosaria. Today many patients suffering from leprosy are still concerned about the social stigma associated with leprosy. As of 1999, there were 719,332 leprosy cases registered for treatment. Almost all of these individuals were receiving multidrug therapy. The new case detection rate stayed the same or was increasing with 747,369 new cases detected during 1998 (http://www.who.int/lep). The prevalence of leprosy is not evenly distributed, as much of the disease seen today occurs in the third world. Most cases (72%) are from Asia and Oceania, whereas 18% are from Africa and only 7% are from the Americas (Lockwood and McAdam, 1998). The genome of the 1–8 µm long and 0.3– 0.5 µm wide M. leprae bacillus is approximately 3.2 MB (Gelber and Rea, 2000). The sequence CHAPTER 1.1.19 has been partially determined and is available for inspection (http://www.sanger.ac.uk/ DataSearch). It is speculated that many of the genes essential for the survival of M. leprae in the environment have been deleted, and the bacillus has become an obligate parasite of its human host. Indeed to date no one has been able to continuously culture the bacillus in either in vitro conditions or cell culture. It may be true that the mycobacterium has lost genes essential for survival in these ex vivo conditions and thus has an absolute requirement for host factors (Gelber and Rea, 2000). In fact, the only methods to propagate M. leprae bacilli are via animal infections with either 9-banded armadillos or severe combined immunodeficiency (SCID) mice. Growth of sufficient bacilli for laboratory manipulation takes approximately one year, inasmuch as the generation time for this organism is approximately 14 days in animal tissues. Leprosy is a chronic disease with a long incubation period. This fact is affirmed by the observation that children younger than two years do not have leprosy symptoms. In addition, people residing in nonendemic countries who have visited a site with endemic leprosy may develop the disease many years after the initial exposure. Hence the incubation period is estimated to range from 2–12 years. Like those infected with TB, only about 10% of those infected with leprosy go on to develop the disease (Lockwood and McAdam, 1998). Being a human disease, leprosy has no established natural reservoir for infection. Mycobacterum leprae is probably transmitted from person to person although the reservoirs and their role in transmission remain controversial issues. In 1898 Schaffer noted that leprosy patients discharged large numbers of acid-fast bacilli (AFB) when coughing, sneezing or speaking normally. In fact, if left untreated a lepromatous leprosy patient may dispel 6.8 × 1010 AFB in a single nose blow. Thus leprosy may to be transmitted via an aerosol route where it goes on to infect the lining of the nose (Lockwood and McAdam, 1998). Though M. leprae produces lesions that often involve the extremities and skin, bacilli probably reach these locations by hematogenous spread. In fact people who have leprosy may have granulomata in the lymph nodes, liver, kidney, spleen, bone marrow, adrenals, testes and eyes. Thus the bacilli can spread to remote portions of the body to cause lesions. Although M. leprae spreads through the body via a systemic route, it prefers to grow at temperatures below 37°C and therefore has a tropism for the extremities and skin where its growth can flourish. The outcome of the disease is strongly dependent on the host’s immune response. Upon infection, bacilli are engulfed by macrophages. If the The Genus Mycobacterium—Medical 929 macrophages are sufficiently activated to kill the invading bacilli, then the infection will be cleared. Otherwise, the bacilli will go on to replicate within macrophages, lyse the cells, and infect other macrophages (Gelber and Rea, 2000). The host’s immune system can take one of two pathways. First, the disease is controlled by strong cell-mediated immunity employing helper T cells of the Th1 lineage. These helper T-cells induce the macrophages to kill the bacilli, and INF-γ and IL-2 are found at the site of infection (Lockwood and McAdam, 1998). Organized granulomata are composed of the macrophages surrounded by the T-cells, resulting in what is known as tuberculoid or paucibacillary leprosy. It is known from the days before antibiotic therapy that tuberculoid leprosy often spontaneously heals (Lockwood and McAdam, 1998). However, tuberculoid leprosy also may result in tissue damage from continuous lymphocyte infiltration into the area of infection. Tuberculoid leprosy has a shorter incubation time, generally 2–5 years, and causes few skin lesions. (Lesions when manifest are hypopigmented and asymmetric.) Nerve damage can occur when granulomata are near small dermal sensory and autonomic nerve fibers (Lockwood and McAdam, 1998). Tuberculoid leprosy is not associated with the presence of stainable M. leprae in skin and does not produce upper respiratory signs and symptoms. The reason is probably that strong cell-mediated immunity controls the infection, though the control is at the expense of significant tissue damage. In general, tuberculoid leprosy has a good prognosis with treatment. The other path of infection occurs in patients who do not have good cell-mediated immunity and good initial killing of the M. leprae. In this expression of the disease, named “lepromatous” or “multibacillary” leprosy, bacilli multiply out of control. This form of the disease has a somewhat longer incubation period, 8–12 years. The bacilli may invade Schwann cells, resulting in demyelination of nerves and nerve damage (Lockwood and McAdam, 1998). Granulomata that do form are poorly organized, with high bacterial loads. Bacilli may replicate within dermal cells as evidenced by AFB in the skin. Most (80% of) lepromatous patients have some nasal symptoms due to invasion of the nasal mucosa by M. leprae. Occasionally, without treatment, the bacilli may cause so much destruction that the nasal septum collapses. In addition, denervation and loss of pain sensation results in repetitive trauma and injury to limbs. Destruction can occur over vast areas of limbs that in some instances may necessitate amputation. In between the two extremes or poles, tuberculoid and lepromatous, there is a continuum. These disease states, which are defined patholog- 930 B. Saviola and W. Bishai ically after review of skin biopsy specimens, are referred to as borderline tuberculoid, borderline leprosy and borderline lepromatous. Borderline tuberculoid has some aspects of lepromatous leprosy, whereas borderline lepromatous leprosy has some aspects of tuberculoid leprosy. Borderline leprosy refers to a state in between lepromatous and tuberculoid. Borderline states are extremely unstable and may shift spontaneously between mild and severe disease. Until recently not much was known about the mechanism by which M. leprae interacts with and causes damage to cells of the nervous system. Recent evidence has revealed that M. leprae binds to the basal lamina surrounding Schwann cells through an interaction between a 21-kDa surface protein on M. leprae and laminin-2 within the basal lamina (Rambukkana et al., 1997; Shimoji et al., 1999). Additionally, M. leprae can interact directly with Schwann cells by binding to α-dystroglycan on the Schwann cell surface. This interaction, however, occurs only in the presence of laminin-2 (Rambukkana et al., 1998). These findings have the promise of providing mechanistic information that may aid in the development of improved therapeutics against leprosy. slow-growing MYCOBACTERIUM KANSASII This organism is the second most frequent cause of pulmonary disease by a nontuberculous mycobacteria (Brown and Wallace, 2004). Mycobacterum kansasii is found in environmental water, and water is probably the source of human infection (Iseman, 1998). The clinical presentations of pulmonary M. kansasii are a chronic cough, lowgrade fever, malaise and chest pain. Though clinical manifestations are similar to those caused by M. tuberculosis, a chest x-ray usually reveals upper-zone disease with fewer fibronodular regions than found with either M. tuberculosis or MAC. Predisposing factors for infection with M. kansasii are smoking-induced chronic bronchitis, chronic inorganic dust exposure, and chronic obstructive pulmonary disease (Toosi and Ellner, 1998). Although extrapulmonary infections are rare, M. kansasii can cause diffuse cutaneous disease as well as cervical lymphadenitis, especially in children. Patients with AIDS or with impaired cellular immunity may also develop a disseminated infection. MYCOBACTERIUM MARINUM M. marinum causes a cutaneus disease in humans known as swimming pool granuloma, fish handler’s nodule or surfer’s nodule. As implied above, water is the major environmental reservoir for this mycobacterial species (Toosi and Ellner, 1998). The bacterium and therefore most cases of infection are mainly found in the coastal areas of North America (Ise- CHAPTER 1.1.19 man, 1998). Humans contract the disease from environmental water, the handling of domestic and wild fish, or the cleaning and maintenance of fish tanks. Mycobacterium marinum lesions appear on the extremities of affected individuals, as the bacillus is most adept at growing at a temperature of 32°C (Brown and Wallace, 2004). Interestingly, M. marinum is closely related to M. tuberculosis on a genetic level. Because M. marinum is not transmitted to humans through an aerosol route and is not considered a biosafety level 3 (BSL3) organism, it has been used as a model system for the study of M. tuberculosis pathogenesis (Ramakrishnan et al., 2000). In animal tissues, M. marinum bacilli infect macrophages and this leads to granuloma formation. As this species of mycobacterium causes disease in both fish and frogs, the nature of its pathogenesis has been investigated in these animal models. Thus, although M. marinum is not a major public health problem, research into its basic biology is ongoing due to its genetic similarity with M. tuberculosis. MYCOBACTERIUM SCROFULACEUM This slowly growing mycobacterium, first isolated in 1956, has an optimal growth temperature of 37°C and, upon initial isolation, can take approximately 4 to 6 weeks to achieve growth. It is a leading cause of scrofula or cervical lymphadenitis in children 1 to 5 years of age. Infrequently it may cause progressive pulmonary disease as well as bone and soft tissue disease and, in some instances, disseminated disease. MYCOBACTERIUM FORTUITUM, M. CHELONAE and M. ABSCESSUS These organisms comprise a group of rapidly growing mycobacteria that are found in soil samples as well as in water; however, their mode of transmission to humans remains unknown (Iseman, 1998). These mycobacteria cause localized infection of the skin, soft tissues and bones. They also may cause pulmonary disease typified by a persistent hacking cough, lowgrade fever, chills, malaise and mucopurulent secretions. MYCOBACTERIUM ULCERANS During the first half of the 20th century this mycobacterial disease was recognized in Bairnsdale, Australia. The syndrome produced an ulcer, subsequently named the “Bairnsdale ulcer,” or “Buruli ulcer,” that was characterized by a chronic progressive ulceration of the skin (Fig. 10). Acid-fast bacilli, later identified as M. ulcerans, were eventually cultivated from lesions of infected individuals. Although no bacteria have been cultivated from environmental sources, the use of polymerase chain reaction (PCR) has identified M. ulcerans in water sources (Ross et al., 1997). Thus it is CHAPTER 1.1.19 The Genus Mycobacterium—Medical 931 the Buruli ulcer (Johnson et al., 1999). The fact that tissue necrosis occurs at sites distal to areas of colonization by M. ulcerans seems to imply that a factor that can diffuse to distant tissues is involved. It has recently been discovered that a polyketide toxin named “mycolactone” is responsible for histopathologic changes distal to the site of infection (George et al., 1998; George et al., 1999). This toxin was present in the culture filtrates of growing M. ulcerans, indicating that it is secreted into the milieu of the bacterium. When this toxin is injected into guinea pig skin, ulcers similar to those caused by M. ulcerans are formed. In addition, exposure of L929 murine fibroblasts to the toxin arrests their cell cycle. Thus, as a toxinogenic microorganism, M. ulcerans has a mode of pathogenesis that is very different from that of other members of the genus Mycobacterium. 6-2-A2. About 1/10 of the ulcers are on the trunk. The advancing edge of the ulcer is frequently hyperpigmiented as seen here. AFIP 74-4472. Fig. 10. A young boy infected with M. ulcerans has a buruli ulcer covering a portion of his torso. From Connor et al. (1976). hypothesized that M. ulcerans is acquired from the environment via water. Indeed it has been noted that people inhabiting areas close to a body of stagnant water in tropical regions are more at risk for developing a Buruli ulcer (Ross et al., 1997; Johnson et al., 1999). The hallmark of the disease is a skin ulcer that begins as a small nodule but (if left untreated) enlarges to encompass a large surface area. The ulcers are themselves painless, suggesting that there is some nerve involvement. Inflammatory response at the site of colonization by the AFB or the site of ulceration appears to be minimal, even though necrosis of the subcutaneous fat tissue is extensive. In contrast to M. tuberculosis, M. ulcerans grows as microcolonies extracellularly during human infection. In addition, treatment of the disease is not trivial. Antibiotic therapy has variable efficacy, and therefore the best treatment is surgical excision after the ulcer has been identified at an early nodule stage. Large ulcers require extensive surgery with skin grafting. In extreme cases where muscle tissue is involved, amputation of an affected limb may be required (Iseman, 1998). It has long been suspected that a toxin mediates the extensive tissue damage associated with Prevention Vaccination with the attenuated M. bovis strain bacille Callmette Guerin (BCG) is common in many parts of the world; however, protection rates due to this vaccine vary from 0– 80%. Whereas BCG probably provides little protection against adult pulmonary TB, it does appear to protect against childhood disseminated TB (Colditz et al., 1994). An additional problem of vaccination is that (once vaccinated with BCG) individuals will have a positive reaction to the administration of purified protein derivative (PPD) from M. tuberculosis, excluding its use as a diagnostic tool. Therefore once a person has been vaccinated, it is impossible to know if they have been infected with M. tuberculosis subsequently and therefore if their risk of developing active disease is substantial. Thus in the United States vaccination is not recommended as a measure to control M. tuberculosis spread. In the United States and many European nations, it is preferable to periodically administer PPD to persons at high risk of becoming infected with M. tuberculosis and to treat prophylactically with antibiotics. Vaccination, however, is useful in reducing miliary and meningeal disease in children in third world countries where tuberculosis rates are high and mass screening is impractical. Literature Cited Adler, J., and D. Rose. 1996. Tuberculosis. In: W. N. Rom and S. Garay (Eds.) Transmission and Pathogenesis of Tuberculosis. Little, Brown and Company. New York, NY. Agerton, T., S. Valway, et al. 1997. Transmission of a highly drug-resistant strain (strain W1) of Mycobacterium tuberculosis: Community outbreak and nosocomial transmission via a contaminated bronchoscope. JAMA 278(13):1073–1077. 932 B. Saviola and W. Bishai Amplicor in press. Mycobacterium Tuberculosis Test Insert Information. Behr, M. A., S. A. Warren, et al. 1999. Transmission of Mycobacterium tuberculosis from patients smear-negative for acid-fast bacilli. Lancet 353(9151):444–449. Birkhaug, K. 1946. Vaccination with the vole bacillus (Wells). American Review of Tuberculosis 54:41–50. Brennan, P. J., and H. Nikaido. 1995. The envelope of mycobacteria. Ann. Rev. Biochem. 64:29–63. Brown, B., and R. Wallace. 2004. Infections due to nontuberculous mycobacteria. In: G. Mandell, J. Bennett, and R. Dolin (Eds.) Principles and Practices of Infectious Diseases. Churchill Livingstone. Philadelphia, PA. Chaisson, R. E., and W. R. Bishai (Eds.). 1997. Mycobacterium avium and tuberculosis infection: Management in patients with HIV disease. Clinical Care Options for HIV Continuum of Care Series Medical Care Collaborative and Healthcare Communications Group. Colditz, G. A., T. F. Brewer, et al. 1994. Efficacy of BCG vaccine in the prevention of tuberculosis. JAMA 271(9):698–702. Cole, S. T., R. Brosch, et al. 1998. Deciphering the biology of Mycobacterium tuberculosis from the complete genome sequence. Nature 393(6685):537–544. Connor, D., W. Meyers, et al. 1976. Infection by Mycobacterium ulcerans. In: C. Binford and D. Connor (Eds.) Pathology of Tropical and Extraordinary Diseases. Armed Forces Institute of Pathology. Washington DC. Curtis, A. B., R. Ridzon, et al. 1999. Extensive transmission of Mycobacterium tuberculosis from a child. N. Engl. J. Med. 341(20):1491–1495. Dannenberg Jr., A. M. 1993. Immunopathogenesis of pulmonary tuberculosis. Hosp. Pract. (Off. Ed.) 28(1): 51–58. Dannenberg Jr., A. M. 1994. Roles of cytotoxic delayed-type hypersensitivity and macrophage-activating cellmediated immunity in the pathogenesis of tuberculosis. Immunobiology 191(4–5):461–473. Davis, A. 1996. History of the Sanitorium Movement. In: W. N. Rom and S. Garay (Eds.) Tuberculosis. Little, Brown and Company. New York, NY. Edlin, B. R., J. I. Tokars, et al. 1992. An outbreak of multidrug-resistant tuberculosis among hospitalized patients with the acquired immunodeficiency syndrome. N. Engl. J. Med. 326(23):1514–1521. Fraser, C. M., A. Mays, et al. 1986. The Merk Veterinary Manual. Merk & Co. Rahway, NJ. Gelber, R., and T. Rea. 2000. Mycobacterium leprae (Leprosy, Hansen’s Disease). In: G. Mandell, J. Bennett, and R. Dolin (Eds.) Principles and Practices of Infectious Diseases. Churchill Livingstone. Philadelphia, PA. George, K. M., L. P. Barker, et al. 1998. Partial purification and characterization of biological effects of a lipid toxin produced by Mycobacterium ulcerans. Infect. Immunol. 66(2):587–593. George, K. M., D. Chatterjee, et al. 1999. Mycolactone: A polyketide toxin from Mycobacterium ulcerans required for virulence. Science 283(5403):854–857. Haas, F., and S. Haas. 1996. The origins of Mycobacterium tuberculosis and the notion of its contagiousness. In: W. N. Rom and S. Garay (Eds.) Tuberculosis. Little, Brown and Company. New York, NY. Harrington, S., and W. Bishai. 2000. Molecular epidemiology and infectious diseases. In: K. Kitzenberg (Ed.) Infectious Disease Epidemiology: Theory and Practice. CHAPTER 1.1.19 Harris, W. 1996. Chemotherapy of tuberculosis: The beginning. In: W. N. Rom and S. Garay (Eds.) Tuberculosis. Little, Brown and Company. New York, NY. Havlir, D., and J. Ellner. 2000. Mycobacterium avium complex. In: G. Mandell, J. Bennett, and R. Dolin (Eds.) Principles and Practices of Infectious Diseases. Churchill Livingstone. Philadelphia, PA. Iseman, M. 1998. Nontuberculous mycobacterial infections. S. Gorbach, J. Bartlett, and N. Blacklow. (Eds.) Infectious Diseases. Sauders and Company. Philadelphia, PA. Johnson, P. D., T. P. Stinear, et al. 1999. Mycobacterium ulcerans—a mini-review. J. Med. Microbiol. 48(6):511– 513. Kent, P. T., and G. P. Kubica. 1985. Public Health Mycobacteriology: A Guide for the Level III Laboratory. Department of Health and Human Services. Atlanta, GA. Lockwood, D., and K. McAdam. 1998. Leprosy. In: S. Gorbach, J. Bartlett, and N. Blacklow. Infectious Diseases. Saunders and Company. Philadelphia, PA. Michele, T. M., W. A. Cronin, et al. 1997. Transmission of Mycobacterium tuberculosis by a fiberoptic bronchoscope: Identification by DNA fingerprinting. JAMA 278(13):1093–1095. Middleton, A. M., M. V. Chadwick, et al. 1997. Evaluation of a commercial radiometric system for primary isolation of mycobacteria over a fifteen-year period. Eur. J. Clin. Microbiol. Infect. Dis. 16(2):166–170. Nolte, F., and B. Metchock. 1995. Mycobacterium. In: P. Murray, E. Baron, M. Pfaller, F. Tenover, and R. Yolken (Eds.) Manual of Clinical Microbiology. ASM Press. Washington DC. Pablos-Mendez, A., M. C. Raviglione, et al. 1998. Global surveillance for antituberculosis-drug resistance, 1994– 1997: World Health Organization, International Union against Tuberculosis and Lung Disease Working Group on Anti-Tuberculosis Drug Resistance Surveillance. N. Engl. J. Med. 338(23):1641–1649. Palella Jr., F. J., K. M. Delaney, et al. 1998. Declining morbidity and mortality among patients with advanced human immunodeficiency virus infection: HIV Outpatient Study Investigators. N. Engl. J. Med. 338(13):853– 860. Pfyffer, G. E., C. Cieslak, et al. 1997. Rapid detection of mycobacteria in clinical specimens by using the automated BACTEC 9000 MB system and comparison with radiometric and solid-culture systems. J. Clin. Microbiol. 35(9):2229–2234. Ramakrishnan, L., N. A. Federspiel, et al. 2000. Granulomaspecific expression of Mycobacterium virulence proteins from the glycine-rich PE-PGRS family. Science 288(5470):1436–1439. Rambukkana, A., J. L. Salzer, et al. 1997. Neural targeting of Mycobacterium leprae mediated by the G domain of the laminin-alpha2 chain. Cell 88(6):811–821. Rambukkana, A., H. Yamada, et al. 1998. Role of alphadystroglycan as a Schwann cell receptor for Mycobacterium leprae. Science 282(5396):2076–2079. Ross, B. C., L. Marino, et al. 1997. Development of a PCR assay for rapid diagnosis of Mycobacterium ulcerans infection. J. Clin. Microbiol. 35(7):1696–1700. Saito, H., Y. Kashiwabara, et al. 1996. Rapid detection of acid-fast bacilli with Mycobacteria Growth Indicator Tube (MGIT). Kekkaku 71(6):399–405. Sharp, S. E., C. A. Suarez, et al. 1996. Evaluation of the mycobacteria growth indicator tube compared to Septi- CHAPTER 1.1.19 Chek AFB for the detection of mycobacteria. Diagn. Microbiol. Infect. Dis. 25(2):71–75. Shimoji, Y., V. Ng, et al. 1999. A 21–kDa surface protein of Mycobacterium leprae binds peripheral nerve laminin-2 and mediates Schwann cell invasion. Proc. Natl. Acad. Sci. USA 96(17):9857–9862. Shinnick, T. M., and R. C. Good. 1994. Mycobacterial taxonomy. Eur. J. Clin. Microbiol. Infect. Dis. 13(11):884–901. Sterling, T. R., D. S. Pope, et al. 2000. Transmission of Mycobacterium tuberculosis from a cadaver to an embalmer. N. Engl. J. Med. 342(4):246–248. Toosi, Z., and J. Ellner. 1998. Mycobacterium tuberculosis and other mycobacteria. In: S. Gorbach, J. Bartlett, and N. Blacklow (Eds.) Infectious Diseases. Sauders and Company. Philadelphia, PA. The Genus Mycobacterium—Medical 933 Valway, S. E., M. P. Sanchez, et al. 1998. An outbreak involving extensive transmission of a virulent strain of Mycobacterium tuberculosis. N. Engl. J. Med. 338(10):633– 639. Wang, S. X., and L. Tay. 1999. Evaluation of three nucleic acid amplification methods for direct detection of Mycobacterium tuberculosis complex in respiratory specimens. J. Clin. Microbiol. 37(6):1932–1934. Wells, A. Q., and D. M. Oxon. 1937. Tuberculosis in wild voles. Lancet 1:1221. Wells, A. Q. 1949. Vaccination with the murine type of tubercle bacillus (vole bacillus). Lancet 53–55. Wells, A. Q., and J. A. H. Wylie. 1954. Vaccination against tuberculosis with the vole bacillus. British Medical Bulletin 10:96–100.